Association between Iron Intake and Diabetic Peripheral Neuropathy in Type 2 Diabetes: Significance of Iron Intake and the Ratio between Iron Intake and Polyunsaturated Fatty Acids Intake


Abstract
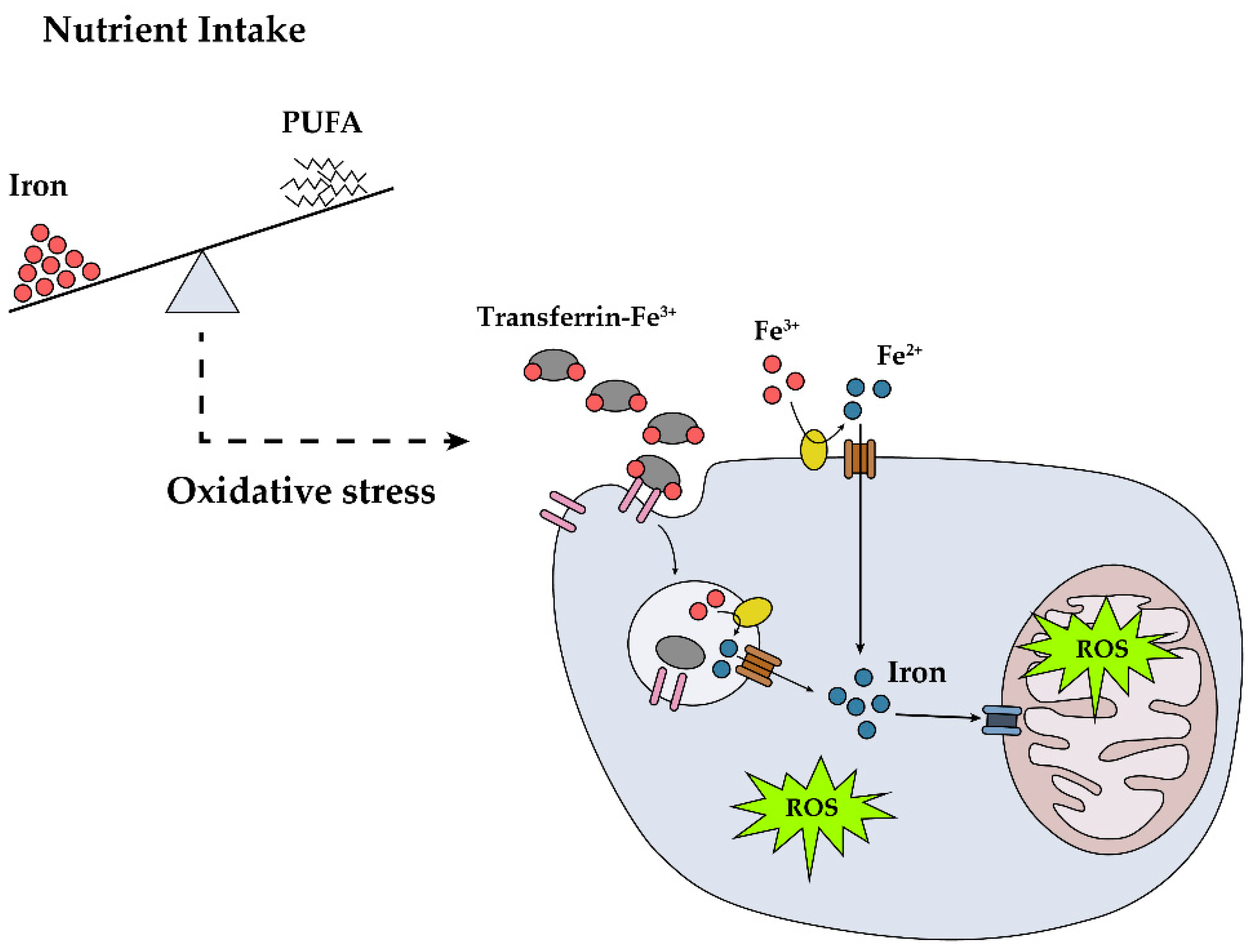
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Assessment of Dietary Intake
2.3. Assessment of DPN
2.4. Anthropometric and Biochemical Measurements
2.5. Statistical Analysis
3. Results
3.1. Characteristics of the Participants
3.2. Iron Intake, Iron/PUFA Ratio, and DPN
3.3. Biochemical Markers of Iron Status and Oxidative Stress Markers of DPN
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cameron, N.E.; Eaton, S.E.; Cotter, M.A.; Tesfaye, S. Vascular factors and metabolic interactions in the pathogenesis of diabetic neuropathy. Diabetologia 2001, 44, 1973–1988. [Google Scholar] [CrossRef] [PubMed]
- Pop-Busui, R.; Boulton, A.J.; Feldman, E.L.; Bril, V.; Freeman, R.; Malik, R.A.; Sosenko, J.M.; Ziegler, D. Diabetic neuropathy: A position statement by the American diabetes association. Diabetes Care 2017, 40, 136–154. [Google Scholar] [CrossRef] [PubMed]
- Papanas, N.; Ziegler, D. Risk factors and comorbidities in diabetic neuropathy: An update 2015. Rev. Diabet. Stud. 2015, 12, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Oh, T.J.; Lee, J.E.; Choi, S.H.; Jang, H.C. Association between body fat and diabetic peripheral neuropathy in middle-aged adults with type 2 diabetes mellitus: A preliminary report. J. Obes. Metab. Syndr. 2019, 28, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.M.; Russell, J.W.; Low, P.; Feldman, E.L. Oxidative stress in the pathogenesis of diabetic neuropathy. Endocr. Rev. 2004, 25, 612–628. [Google Scholar] [CrossRef]
- Mijnhout, G.S.; Kollen, B.J.; Alkhalaf, A.; Kleefstra, N.; Bilo, H.J. Alpha lipoic acid for symptomatic peripheral neuropathy in patients with diabetes: A meta-analysis of randomized controlled trials. Int. J. Endocrinol. 2012, 2012, 456279. [Google Scholar] [CrossRef]
- Evert, A.B.; Dennison, M.; Gardner, C.D.; Garvey, W.T.; Lau, K.H.K.; MacLeod, J.; Mitri, J.; Pereira, R.F.; Rawlings, K.; Robinson, S.; et al. Nutrition therapy for adults with diabetes or prediabetes: A consensus report. Diabetes Care 2019, 42, 731–754. [Google Scholar] [CrossRef]
- Davies, M.J.; D’Alessio, D.A.; Fradkin, J.; Kernan, W.N.; Mathieu, C.; Mingrone, G.; Rossing, P.; Tsapas, A.; Wexler, D.J.; Buse, J.B. Management of hyperglycaemia in type 2 diabetes, 2018. A consensus report by the American Diabetes Association (ADA) and the European Association for the Study of Diabetes (EASD). Diabetologia 2018, 61, 2461–2498. [Google Scholar] [CrossRef]
- Salas-Salvado, J.; Bullo, M.; Estruch, R.; Ros, E.; Covas, M.I.; Ibarrola-Jurado, N.; Corella, D.; Aros, F.; Gomez-Gracia, E.; Ruiz-Gutierrez, V.; et al. Prevention of diabetes with mediterranean diets: A subgroup analysis of a randomized trial. Ann. Intern. Med. 2014, 160, 1–10. [Google Scholar] [CrossRef]
- Chiu, T.H.T.; Pan, W.H.; Lin, M.N.; Lin, C.L. Vegetarian diet, change in dietary patterns, and diabetes risk: A prospective study. Nutr. Diabetes 2018, 8, 12. [Google Scholar] [CrossRef]
- Tay, J.; Thompson, C.H.; Luscombe-Marsh, N.D.; Wycherley, T.P.; Noakes, M.; Buckley, J.D.; Wittert, G.A.; Yancy, W.S., Jr.; Brinkworth, G.D. Effects of an energy-restricted low-carbohydrate, high unsaturated fat/low saturated fat diet versus a high-carbohydrate, low-fat diet in type 2 diabetes: A 2-year randomized clinical trial. Diabetes Obes. Metab. 2018, 20, 858–871. [Google Scholar] [CrossRef]
- Lv, W.S.; Zhao, W.J.; Gong, S.L.; Fang, D.D.; Wang, B.; Fu, Z.J.; Yan, S.L.; Wang, Y.G. Serum 25-hydroxyvitamin d levels and peripheral neuropathy in patients with type 2 diabetes: A systematic review and meta-analysis. J. Endocrinol. Invest. 2015, 38, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Shillo, P.; Selvarajah, D.; Greig, M.; Gandhi, R.; Rao, G.; Wilkinson, I.D.; Anand, P.; Tesfaye, S. Reduced vitamin d levels in painful diabetic peripheral neuropathy. Diabet. Med. 2019, 36, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Jain, A.; Rohatgi, A. An observational study of vitamin b12 levels and peripheral neuropathy profile in patients of diabetes mellitus on metformin therapy. Diabetes Metab. Syndr. 2018, 12, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Gorson, K.C.; Ropper, A.H. Additional causes for distal sensory polyneuropathy in diabetic patients. J. Neurol. Neurosurg. Psychiatry 2006, 77, 354–358. [Google Scholar] [CrossRef]
- Guo, W.; Zhou, Q.; Jia, Y.; Xu, J. Cluster and factor analysis of elements in serum and urine of diabetic patients with peripheral neuropathy and healthy people. Biol. Trace Elem. Res. 2020, 194, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Cairo, G.; Bernuzzi, F.; Recalcati, S. A precious metal: Iron, an essential nutrient for all cells. Genes Nutr. 2006, 1, 25–39. [Google Scholar] [CrossRef]
- Simcox, J.A.; McClain, D.A. Iron and diabetes risk. Cell Metab. 2013, 17, 329–341. [Google Scholar] [CrossRef]
- Shi, Z.; Zhou, M.; Yuan, B.; Qi, L.; Dai, Y.; Luo, Y.; Holmboe-Ottesen, G. Iron intake and body iron stores, anaemia and risk of hyperglycaemia among Chinese adults: The prospective Jiangsu Nutrition Study (JIN). Public Health Nutr. 2010, 13, 1319–1327. [Google Scholar] [CrossRef]
- Eshak, E.S.; Iso, H.; Maruyama, K.; Muraki, I.; Tamakoshi, A. Associations between dietary intakes of iron, copper and zinc with risk of type 2 diabetes mellitus: A large population-based prospective cohort study. Clin. Nutr. 2018, 37, 667–674. [Google Scholar] [CrossRef]
- Baum, P.; Kosacka, J.; Estrela-Lopis, I.; Woidt, K.; Serke, H.; Paeschke, S.; Stockinger, M.; Kloting, N.; Bluher, M.; Dorn, M.; et al. The role of nerve inflammation and exogenous iron load in experimental peripheral diabetic neuropathy (PDN). Metabolism 2016, 65, 391–405. [Google Scholar] [CrossRef] [PubMed]
- Kosacka, J.; Woidt, K.; Toyka, K.V.; Paeschke, S.; Kloting, N.; Bechmann, I.; Bluher, M.; Thiery, J.; Ossmann, S.; Baum, P.; et al. The role of dietary non-heme iron load and peripheral nerve inflammation in the development of peripheral neuropathy (PN) in obese non-diabetic leptin-deficient ob/ob mice. Neurol. Res. 2019, 41, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Paeschke, S.; Baum, P.; Toyka, K.V.; Bluher, M.; Koj, S.; Kloting, N.; Bechmann, I.; Thiery, J.; Kosacka, J.; Nowicki, M. The role of iron and nerve inflammation in diabetes mellitus type 2-induced peripheral neuropathy. Neuroscience 2019, 406, 496–509. [Google Scholar] [CrossRef] [PubMed]
- Richard, D.; Kefi, K.; Barbe, U.; Bausero, P.; Visioli, F. Polyunsaturated fatty acids as antioxidants. Pharmacol. Res. 2008, 57, 451–455. [Google Scholar] [CrossRef]
- Meyer, K.A.; Kushi, L.H.; Jacobs, D.R., Jr.; Folsom, A.R. Dietary fat and incidence of type 2 diabetes in older Iowa women. Diabetes Care 2001, 24, 1528–1535. [Google Scholar] [CrossRef]
- Salmeron, J.; Hu, F.B.; Manson, J.E.; Stampfer, M.J.; Colditz, G.A.; Rimm, E.B.; Willett, W.C. Dietary fat intake and risk of type 2 diabetes in women. Am. J. Clin. Nutr. 2001, 73, 1019–1026. [Google Scholar] [CrossRef]
- Tao, M.; McDowell, M.A.; Saydah, S.H.; Eberhardt, M.S. Relationship of polyunsaturated fatty acid intake to peripheral neuropathy among adults with diabetes in the National Health and Nutrition Examination Survey (NHANES) 1999–2004. Diabetes Care 2008, 31, 93–95. [Google Scholar] [CrossRef]
- Park, S.; Kim, S.; Lee, S.H.; Choe, J.; Choi, Y. Development of 9th revision Korean food composition table and its major changes. Korean J. Community Nutr. 2018, 23, 352–365. [Google Scholar] [CrossRef]
- Feldman, E.L.; Stevens, M.J.; Thomas, P.K.; Brown, M.B.; Canal, N.; Greene, D.A. A practical two-step quantitative clinical and electrophysiological assessment for the diagnosis and staging of diabetic neuropathy. Diabetes Care 1994, 17, 1281–1289. [Google Scholar] [CrossRef]
- Won, J.C.; Im, Y.J.; Lee, J.H.; Kim, C.H.; Kwon, H.S.; Cha, B.Y.; Park, T.S. Clinical phenotype of diabetic peripheral neuropathy and relation to symptom patterns: Cluster and factor analysis in patients with type 2 diabetes in Korea. J. Diabetes Res. 2017, 2017, 5751687. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Fang, A.; Yu, S.; Shen, X.; Li, K. Dietary nonheme, heme, and total iron intake and the risk of diabetes in adults: Results from the China Health and Nutrition Survey. Diabetes Care 2020, 43, 776–784. [Google Scholar] [CrossRef] [PubMed]
- Peterlin, B.; Globocnik Petrovic, M.; Makuc, J.; Hawlina, M.; Petrovic, D. A hemochromatosis-causing mutation c282y is a risk factor for proliferative diabetic retinopathy in Caucasians with type 2 diabetes. J. Hum. Genet. 2003, 48, 646–649. [Google Scholar] [CrossRef]
- Alfrey, A.C.; Froment, D.H.; Hammond, W.S. Role of iron in the tubulo-interstitial injury in nephrotoxic serum nephritis. Kidney Int. 1989, 36, 753–759. [Google Scholar] [CrossRef]
- Facchini, F.S.; Saylor, K.L. A low-iron-available, polyphenol-enriched, carbohydrate-restricted diet to slow progression of diabetic nephropathy. Diabetes 2003, 52, 1204–1209. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhang, L.; Xu, Z.; Chen, W. Neurotoxic effects of iron overload under high glucose concentration. Neural Regen. Res. 2013, 8, 3423–3433. [Google Scholar]
- Newsholme, P.; Keane, K.N.; Carlessi, R.; Cruzat, V. Oxidative stress pathways in pancreatic beta-cells and insulin-sensitive cells and tissues: Importance to cell metabolism, function, and dysfunction. Am. J. Physiol. Cell Physiol. 2019, 317, C420–C433. [Google Scholar] [CrossRef] [PubMed]
- Pitocco, D.; Tesauro, M.; Alessandro, R.; Ghirlanda, G.; Cardillo, C. Oxidative stress in diabetes: Implications for vascular and other complications. Int. J. Mol. Sci. 2013, 14, 21525–21550. [Google Scholar] [CrossRef]
- Emerit, J.; Beaumont, C.; Trivin, F. Iron metabolism, free radicals, and oxidative injury. Biomed. Pharmacother. 2001, 55, 333–339. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Rossi, R.; Colombo, R.; Giustarini, D.; Milzani, A. Biomarkers of oxidative damage in human disease. Clin. Chem. 2006, 52, 601–623. [Google Scholar] [CrossRef]
- Li, J.; Zhang, H.; Xie, M.; Yan, L.; Chen, J.; Wang, H. NSE, a potential biomarker, is closely connected to diabetic peripheral neuropathy. Diabetes Care 2013, 36, 3405–3410. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.F.; Casellini, C.M.; Parson, H.K.; Obrosova, I.G.; Yorek, M.; Vinik, A.I. Role of peroxynitrite in the development of diabetic peripheral neuropathy. Diabetes Care 2015, 38, e100–e101. [Google Scholar] [CrossRef] [PubMed]
- Bigagli, E.; Lodovici, M. Circulating oxidative stress biomarkers in clinical studies on type 2 diabetes and its complications. Oxid. Med. Cell Longev. 2019, 2019, 5953685. [Google Scholar] [CrossRef]
- Gupta, S.; Gambhir, J.K.; Kalra, O.; Gautam, A.; Shukla, K.; Mehndiratta, M.; Agarwal, S.; Shukla, R. Association of biomarkers of inflammation and oxidative stress with the risk of chronic kidney disease in type 2 diabetes mellitus in north Indian population. J. Diabetes Complications 2013, 27, 548–552. [Google Scholar] [CrossRef]
- Tatsch, E.; De Carvalho, J.A.; Hausen, B.S.; Bollick, Y.S.; Torbitz, V.D.; Duarte, T.; Scolari, R.; Duarte, M.M.; Londero, S.W.; Vaucher, R.A.; et al. Oxidative DNA damage is associated with inflammatory response, insulin resistance and microvascular complications in type 2 diabetes. Mutat. Res. 2015, 782, 17–22. [Google Scholar] [CrossRef]
- Ganz, T.; Nemeth, E. Hepcidin and iron homeostasis. Biochim. Biophys. Acta 2012, 1823, 1434–1443. [Google Scholar] [CrossRef]
- Enko, D.; Wagner, H.; Kriegshauser, G.; Kimbacher, C.; Stolba, R.; Worf, E.; Halwachs-Baumann, G. Hepcidin-25 vs. Conventional clinical biomarkers in the diagnosis of functional iron deficiency. Eur. J. Haematol. 2015, 95, 507–513. [Google Scholar] [CrossRef]
- Hurrell, R.; Egli, I. Iron bioavailability and dietary reference values. Am. J. Clin. Nutr. 2010, 91, 1461S–1467S. [Google Scholar] [CrossRef] [PubMed]
- Das, N.K.; Schwartz, A.J.; Barthel, G.; Inohara, N.; Liu, Q.; Sankar, A.; Hill, D.R.; Ma, X.; Lamberg, O.; Schnizlein, M.K.; et al. Microbial metabolite signaling is required for systemic iron homeostasis. Cell Metab. 2020, 31, 115–130. [Google Scholar] [CrossRef]
- O’Brien, P.D.; Sakowski, S.A.; Feldman, E.L. Mouse models of diabetic neuropathy. ILAR J. 2014, 54, 259–272. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance — A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef]
- Sokola-Wysoczanska, E.; Wysoczanski, T.; Wagner, J.; Czyz, K.; Bodkowski, R.; Lochynski, S.; Patkowska-Sokola, B. Polyunsaturated fatty acids and their potential therapeutic role in cardiovascular system disorders — A review. Nutrients 2018, 10, 1561. [Google Scholar] [CrossRef]
- Sakai, C.; Ishida, M.; Ohba, H.; Yamashita, H.; Uchida, H.; Yoshizumi, M.; Ishida, T. Fish oil omega-3 polyunsaturated fatty acids attenuate oxidative stress-induced DNA damage in vascular endothelial cells. PLoS ONE 2017, 12, e0187934. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Kawasaki, R.; Rogers, S.; Man, R.E.; Itakura, K.; Xie, J.; Flood, V.; Tsubota, K.; Lamoureux, E.; Wang, J.J. The associations of dietary intake of polyunsaturated fatty acids with diabetic retinopathy in well-controlled diabetes. Invest. Ophthalmol. Vis. Sci. 2015, 56, 7473–7479. [Google Scholar] [CrossRef]
- Chewcharat, A.; Chewcharat, P.; Rutirapong, A.; Papatheodorou, S. The effects of omega-3 fatty acids on diabetic nephropathy: A meta-analysis of randomized controlled trials. PLoS ONE 2020, 15, e0228315. [Google Scholar] [CrossRef]
- ORIGIN Trial Investigators. N-3 fatty acids and cardiovascular outcomes in patients with dysglycemia. N. Engl. J. Med. 2012, 367, 309–318. [Google Scholar] [CrossRef]
- ASCEND Study Collaborative Group. Effects of n-3 fatty acid supplements in diabetes mellitus. N. Engl. J. Med. 2018, 379, 1540–1550. [Google Scholar] [CrossRef]
- Aung, T.; Halsey, J.; Kromhout, D.; Gerstein, H.C.; Marchioli, R.; Tavazzi, L.; Geleijnse, J.M.; Rauch, B.; Ness, A.; Galan, P.; et al. Associations of omega-3 fatty acid supplement use with cardiovascular disease risks: Meta-analysis of 10 trials involving 77917 individuals. JAMA Cardiol. 2018, 3, 225–234. [Google Scholar] [CrossRef]
- Yokoyama, M.; Origasa, H.; Matsuzaki, M.; Matsuzawa, Y.; Saito, Y.; Ishikawa, Y.; Oikawa, S.; Sasaki, J.; Hishida, H.; Itakura, H.; et al. Effects of eicosapentaenoic acid on major coronary events in hypercholesterolaemic patients (JELIS): A randomised open-label, blinded endpoint analysis. Lancet 2007, 369, 1090–1098. [Google Scholar] [CrossRef]
- Bhatt, D.L.; Steg, P.G.; Miller, M.; Brinton, E.A.; Jacobson, T.A.; Ketchum, S.B.; Doyle, R.T., Jr.; Juliano, R.A.; Jiao, L.; Granowitz, C.; et al. Cardiovascular risk reduction with icosapent ethyl for hypertriglyceridemia. N. Engl J. Med. 2019, 380, 11–22. [Google Scholar] [CrossRef]
- Patterson, E.; Wall, R.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. Health implications of high dietary omega-6 polyunsaturated fatty acids. J. Nutr. Metab. 2012, 2012, 539426. [Google Scholar] [CrossRef]
- Naughton, S.S.; Mathai, M.L.; Hryciw, D.H.; McAinch, A.J. Linoleic acid and the pathogenesis of obesity. Prostaglandins Other Lipid Mediat. 2016, 125, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, R.; Warnakula, S.; Kunutsor, S.; Crowe, F.; Ward, H.A.; Johnson, L.; Franco, O.H.; Butterworth, A.S.; Forouhi, N.G.; Thompson, S.G.; et al. Association of dietary, circulating, and supplement fatty acids with coronary risk: A systematic review and meta-analysis. Ann. Intern. Med. 2014, 160, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.; Al-Khudairy, L.; Abdelhamid, A.S.; Rees, K.; Brainard, J.S.; Brown, T.J.; Ajabnoor, S.M.; O’Brien, A.T.; Winstanley, L.E.; Donaldson, D.H.; et al. Omega-6 fats for the primary and secondary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2018, 7, CD011094. [Google Scholar]
- Cameron, N.E.; Cotter, M.A. Effects of an extracellular metal chelator on neurovascular function in diabetic rats. Diabetologia 2001, 44, 621–628. [Google Scholar] [CrossRef]


{kind=link}
| Characteristic | DPN (–) (n = 79) | DPN (+) (n = 68) | p Value | ||||
|---|---|---|---|---|---|---|---|
| Male, n (%) | 45 (56.9) | 41 (60.3) | 0.683 | ||||
| Age (years) | 57.5 | ± | 9.0 | 60.1 | ± | 9.8 | 0.090 |
| Height (cm) | 163.4 | ± | 8.1 | 163.3 | ± | 9.2 | 0.938 |
| Body weight (kg) | 66.1 | ± | 11.0 | 68.7 | ± | 11.1 | 0.158 |
| BMI (kg/m2) | 24.7 | ± | 3.2 | 25.7 | ± | 2.9 | 0.053 |
| Systolic BP (mmHg) | 128 | ± | 13 | 132 | ± | 15 | 0.155 |
| Diastolic BP (mmHg) | 75 | ± | 9 | 75 | ± | 9 | 0.971 |
| Diabetes duration (years) | 9.4 | ± | 7.2 | 10.4 | ± | 7.0 | 0.377 |
| FPG (mmol/L) | 7.5 | ± | 1.6 | 7.9 | ± | 2.3 | 0.222 |
| HbA1c (mmol/mol) | 54.6 | ± | 13.1 | 57.9 | ± | 15.3 | 0.154 |
| HbA1c (%) | 7.1 | ± | 1.2 | 7.4 | ± | 1.4 | 0.153 |
| Total cholesterol (mmol/L) | 4.3 | ± | 1.0 | 3.9 | ± | 0.8 | 0.002 |
| Triglyceride (mmol/L) a | 1.3 | ± | 0.0 | 1.4 | ± | 0.0 | 0.283 |
| HDL–cholesterol (mmol/L) a | 1.2 | ± | 0.0 | 1.2 | ± | 0.0 | 0.070 |
| LDL–cholesterol (mmol/L) | 2.5 | ± | 0.7 | 2.2 | ± | 0.6 | 0.006 |
| Urea nitrogen (mmol/L) | 5.6 | ± | 1.5 | 5.9 | ± | 2.3 | 0.378 |
| Creatinine (μmol/L) | 68.1 | ± | 18.6 | 74.3 | ± | 23.0 | 0.094 |
| eGFR (mL min−1 (1.73 m)−2) | 97.4 | ± | 21.1 | 90.1 | ± | 22.2 | 0.042 |
| AST (U/L)a | 26.0 | ± | 1.4 | 27.2 | ± | 1.4 | 0.437 |
| ALT (U/L) | 28.9 | ± | 17.1 | 27.1 | ± | 12.3 | 0.469 |
| Insulin (pmol/L) | 58.1 | ± | 32.2 | 63.9 | ± | 33.0 | 0.293 |
| HOMA-IR | 2.6 | ± | 1.6 | 3.2 | ± | 1.9 | 0.065 |
| HOMA-B a | 38.8 | ± | 2.3 | 40.9 | ± | 2.0 | 0.670 |
| MNSI-Q (score) | 1.7 | ± | 2.0 | 2.9 | ± | 2.1 | <0.001 |
| MNSI-PE (score) | 1.3 | ± | 0.6 | 3.5 | ± | 0.7 | <0.001 |
| Smoking status, n (%) | 0.350 | ||||||
| Never smoker | 33 (41.8) | 36 (52.9) | |||||
| Ex-smoker | 30 (38.0) | 19 (27.9) | |||||
| Current smoker | 16 (20.3) | 13 (19.1) | |||||
| Alcohol, n (%) | 43 (54.4) | 34 (50.0) | 0.592 | ||||
| Variable | DPN (–) (n = 79) | DPN (+) (n = 68) | p Value | p Value a | ||||
|---|---|---|---|---|---|---|---|---|
| TE intake (kJ) | 6255.9 | ± | 1812.5 | 6218.3 | ± | 1412.6 | 0.889 | NA |
| Carbohydrate (% TE) | 61.5 | ± | 12.2 | 63.4 | ± | 10.7 | 0.327 | NA |
| Protein (% TE) | 16.5 | ± | 3.5 | 16.5 | ± | 3.2 | 0.934 | NA |
| Fat (% TE) | 23.7 | ± | 9.2 | 22.9 | ± | 9.3 | 0.612 | NA |
| SFA (g) | 13.4 | ± | 8.1 | 12.6 | ± | 7.1 | 0.500 | 0.434 |
| MUFA (g) | 13.0 | ± | 7.9 | 13.0 | ± | 7.9 | 0.987 | 0.887 |
| PUFA (g) | 10.2 | ± | 5.2 | 9.2 | ± | 4.7 | 0.211 | 0.155 |
| Omega-6 PUFA (g) | 8.6 | ± | 4.5 | 7.6 | ± | 3.9 | 0.144 | 0.092 |
| Omega-3 PUFA (g) | 1.4 | ± | 0.8 | 1.3 | ± | 1.0 | 0.637 | 0.653 |
| Omega-6/Omega-3 PUFA ratio | 7.2 | ± | 2.7 | 7.3 | ± | 3.1 | 0.811 | NA |
| PUFA/SFA ratio | 0.9 | ± | 0.6 | 0.9 | ± | 0.5 | 0.698 | NA |
| MUFA/SFA ratio | 1.0 | ± | 0.2 | 1.0 | ± | 0.2 | 0.253 | NA |
| Iron (mg) | 9.9 | ± | 3.6 | 10.9 | ± | 4.0 | 0.130 | 0.041 |
| Iron/PUFA ratio (mg/g) | 1.1 | ± | 0.4 | 1.4 | ± | 0.8 | 0.005 | NA |
| Iron/omega-6 PUFA ratio (mg/g) | 1.3 | ± | 0.5 | 1.7 | ± | 1.1 | 0.006 | NA |
| Iron/omega-3 PUFA ratio (mg/g) | 9.3 | ± | 5.5 | 12.1 | ± | 7.8 | 0.016 | NA |
| Vitamin B1 (mg) | 0.9 | ± | 0.4 | 0.8 | ± | 0.3 | 0.635 | 0.625 |
| Vitamin B6 (mg) | 0.4 | ± | 0.2 | 0.4 | ± | 0.2 | 0.922 | 0.871 |
| Vitamin B12 (µg) | 3.7 | ± | 3.1 | 3.7 | ± | 3.5 | 0.988 | 0.967 |
| Vitamin D (µg) | 4.2 | ± | 4.2 | 3.1 | ± | 4.0 | 0.116 | 0.118 |
| Dietary fiber (g) | 21.7 | ± | 8.2 | 23.3 | ± | 8.1 | 0.240 | 0.156 |
| Variable | Coefficient | p Value |
|---|---|---|
| Carbohydrate (% TE) | 0.095 | 0.250 |
| Protein (% TE) | 0.014 | 0.870 |
| Fat (% TE) | −0.059 | 0.480 |
| SFA (g) a | −0.078 | 0.347 |
| MUFA (g) a | −0.037 | 0.661 |
| PUFA (g) a | −0.118 | 0.155 |
| Omega-6 PUFA a | −0.146 | 0.079 |
| Omega-3 PUFA a | 0.003 | 0.976 |
| Omega-6/Omega-3 PUFA ratio | −0.023 | 0.780 |
| PUFA/SFA ratio | 0.003 | 0.969 |
| MUFA/SFA ratio | 0.029 | 0.731 |
| Iron (mg) a | 0.262 | 0.001 |
| Iron/PUFA ratio (mg/g) | 0.276 | 0.001 |
| Iron/omega-6 PUFA ratio (mg/g) | 0.271 | 0.001 |
| Iron/omega-3 PUFA ratio (mg/g) | 0.204 | 0.013 |
| Vitamin B1 (mg) a | −0.075 | 0.368 |
| Vitamin B6 (mg) a | 0.021 | 0.806 |
| Vitamin B12 (µg) a | 0.056 | 0.501 |
| Vitamin D (µg) a | −0.089 | 0.285 |
| Dietary fiber (g) a | 0.151 | 0.068 |
| Variable | OR | 95% CI | p Value |
|---|---|---|---|
| Iron intake | |||
| Model 1 | 1.126 | 1.003, 1.264 | 0.044 |
| Model 2 | 1.147 | 1.014, 1.298 | 0.029 |
| Model 3 | 1.152 | 1.012, 1.311 | 0.032 |
| Model 4 | 1.136 | 0.995, 1.297 | 0.059 |
| Model 5 | 1.139 | 0.998, 1.301 | 0.053 |
| Iron/PUFA ratio | |||
| Model 1 | 2.628 | 1.324, 5.216 | 0.006 |
| Model 2 | 2.375 | 1.168, 4.830 | 0.017 |
| Model 3 | 2.283 | 1.066, 4.887 | 0.034 |
| Model 4 | 2.215 | 1.032, 4.757 | 0.041 |
| Model 5 | 2.214 | 1.034, 4.742 | 0.041 |
| Iron/omega-6 PUFA ratio | |||
| Model 1 | 2.321 | 1.287, 4.186 | 0.005 |
| Model 2 | 2.136 | 1.160, 3.934 | 0.015 |
| Model 3 | 2.096 | 1.089, 4.032 | 0.027 |
| Model 4 | 2.037 | 1.058, 3.922 | 0.033 |
| Model 5 | 2.046 | 1.063, 3.935 | 0.032 |
| Iron/omega-3 PUFA ratio | |||
| Model 1 | 1.069 | 1.012, 1.130 | 0.018 |
| Model 2 | 1.051 | 0.992, 1.112 | 0.090 |
| Model 3 | 1.054 | 0.995, 1.117 | 0.073 |
| Model 4 | 1.055 | 0.995, 1.118 | 0.074 |
| Model 5 | 1.052 | 0.993, 1.115 | 0.082 |
| Variable | DPN (–) (n = 79) | DPN (+) (n = 68) | p Value | ||||
|---|---|---|---|---|---|---|---|
| Transferrin (μmol/L) | 31.6 | ± | 4.5 | 32.1 | ± | 4.3 | 0.494 |
| Ferritin (μg/L) | 150.2 | ± | 120.4 | 140.8 | ± | 109.5 | 0.627 |
| Iron (μmol/L) | 19.2 | ± | 7.0 | 19.2 | ± | 6.8 | 0.975 |
| TIBC (μmol/L) | 60.6 | ± | 8.2 | 61.9 | ± | 7.9 | 0.325 |
| TSAT (%) | 31.9 | ± | 10.5 | 31.7 | ± | 11.9 | 0.897 |
| Hepcidin (ng/mL) | 3.2 | ± | 1.7 | 3.0 | ± | 1.7 | 0.525 |
| NSE (ng/mL) | 12.6 | ± | 3.6 | 12.3 | ± | 2.6 | 0.573 |
| Nitrotyrosine (nmol/L) a | 10.0 | ± | 2.2 | 9.0 | ± | 2.0 | 0.276 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.; Song, Y.; Oh, T.J.; Choi, S.H.; Jang, H.C. Association between Iron Intake and Diabetic Peripheral Neuropathy in Type 2 Diabetes: Significance of Iron Intake and the Ratio between Iron Intake and Polyunsaturated Fatty Acids Intake. Nutrients 2020, 12, 3365. https://doi.org/10.3390/nu12113365
Kim K, Song Y, Oh TJ, Choi SH, Jang HC. Association between Iron Intake and Diabetic Peripheral Neuropathy in Type 2 Diabetes: Significance of Iron Intake and the Ratio between Iron Intake and Polyunsaturated Fatty Acids Intake. Nutrients. 2020; 12(11):3365. https://doi.org/10.3390/nu12113365
Chicago/Turabian StyleKim, Kyuho, YoonJu Song, Tae Jung Oh, Sung Hee Choi, and Hak Chul Jang. 2020. "Association between Iron Intake and Diabetic Peripheral Neuropathy in Type 2 Diabetes: Significance of Iron Intake and the Ratio between Iron Intake and Polyunsaturated Fatty Acids Intake" Nutrients 12, no. 11: 3365. https://doi.org/10.3390/nu12113365
APA StyleKim, K., Song, Y., Oh, T. J., Choi, S. H., & Jang, H. C. (2020). Association between Iron Intake and Diabetic Peripheral Neuropathy in Type 2 Diabetes: Significance of Iron Intake and the Ratio between Iron Intake and Polyunsaturated Fatty Acids Intake. Nutrients, 12(11), 3365. https://doi.org/10.3390/nu12113365