Nutrition in Cancer Therapy in the Elderly—An Epigenetic Connection?

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Management of Older Adult Cancer Patients
3. Nutrition, Aging, and Cancer in the Elderly
4. Nutrition and Cancer Therapy in the Elderly
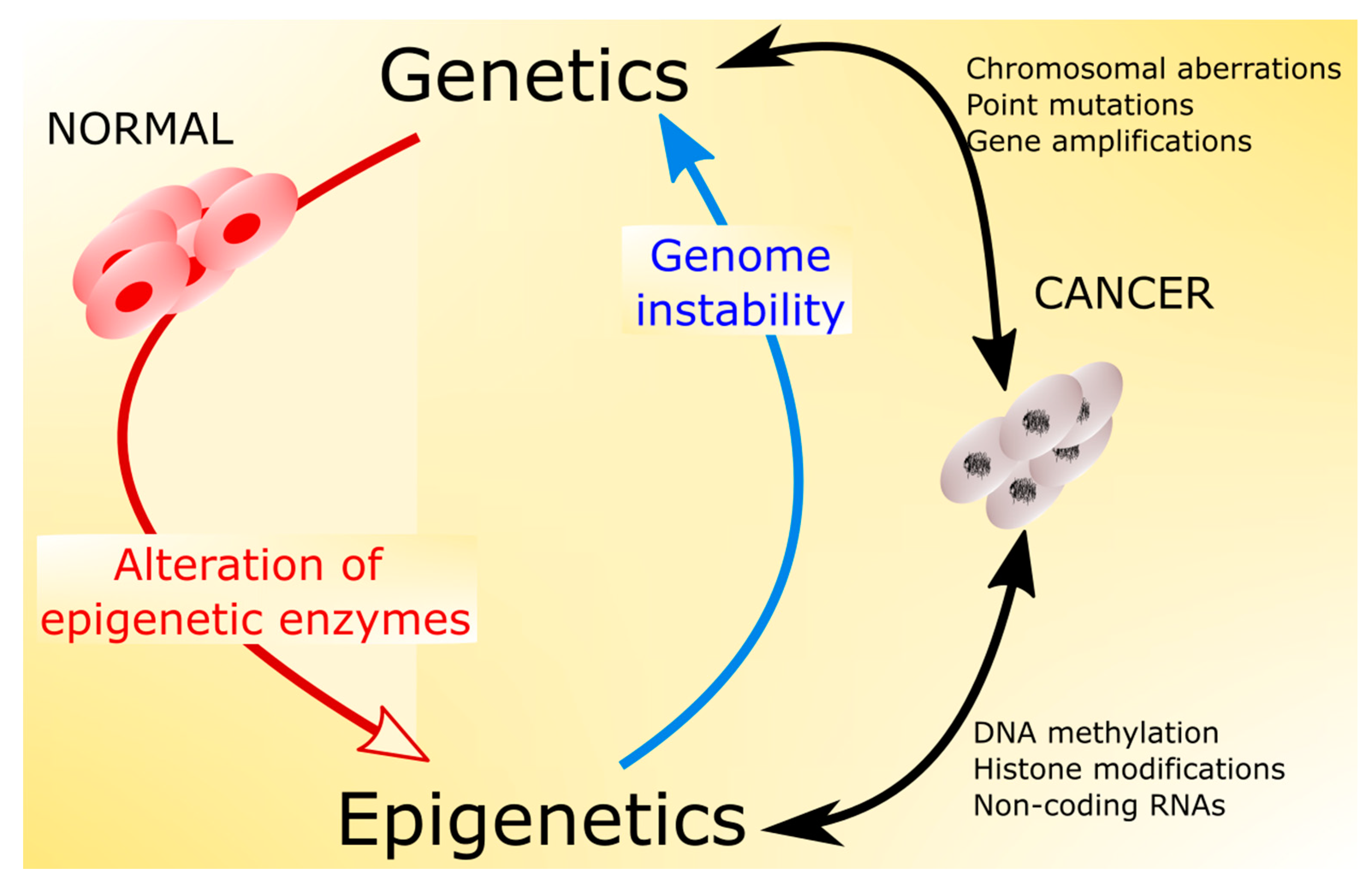
5. Epigenetic Mechanisms in Cancer Transformation and Aging
6. Epigenetic Link between Nutrition, Aging, and Cancer
7. Summary, Conclusions, and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- White, M.C.; Holman, D.M.; Boehm, J.E.; Peipins, L.A.; Grossman, M.; Henley, S.J. Age and Cancer Risk. Am. J. Prev. Med. 2014, 46, S7–S15. [Google Scholar] [CrossRef] [PubMed]
- Presley, C.J.; Dotan, E.; Soto-Perez-De-Celis, E.; Jatoi, A.; Mohile, S.G.; Won, E.; Alibhai, S.; Kilari, D.; Harrison, R.; Klepin, H.D.; et al. Gaps in nutritional research among older adults with cancer. J. Geriatr. Oncol. 2016, 7, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Paillaud, E.; Caillet, P.; Campillo, B.; Bories, P.N. Increased risk of alteration of nutritional status in hospitalized elderly patients with advanced cancer. J. Nutr. Health Aging 2006, 10, 91–95. [Google Scholar] [PubMed]
- Kagohara, L.T.; Stein-O’Brien, G.L.; Kelley, D.; Flam, E.; Wick, H.C.; Danilova, L.V.; Easwaran, H.; Favorov, A.V.; Qian, J.; Gaykalova, D.A.; et al. Epigenetic regulation of gene expression in cancer: Techniques, resources and analysis. Briefings Funct. Genom. 2017, 17, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Tiffon, C. The Impact of Nutrition and Environmental Epigenetics on Human Health and Disease. Int. J. Mol. Sci. 2018, 19, 3425. [Google Scholar] [CrossRef] [PubMed]
- Klepin, H.; Mohile, S.; Hurria, A. Geriatric assessment in older patients with breast cancer. J. Natl. Compr. Cancer Netw. 2009, 7, 226–236. [Google Scholar] [CrossRef][Green Version]
- Welsh, T.J.; Gordon, A.L.; Gladman, J.R. Comprehensive geriatric assessment a guide for the non-specialist. Int. J. Clin. Pract. 2013, 68, 290–293. [Google Scholar] [CrossRef]
- Li, X.; Ploner, A.; Wang, Y.; Magnusson, P.K.; Reynolds, C.; Finkel, D.; Pedersen, N.L.; Jylhävä, J.; Hägg, S. Longitudinal trajectories, correlations and mortality associations of nine biological ages across 20-years follow-up. eLife 2020, 9. [Google Scholar] [CrossRef]
- Ravasco, P. Nutrition in Cancer Patients. J. Clin. Med. 2019, 8, 1211. [Google Scholar] [CrossRef]
- Vellas, B.; Guigoz, Y.; Garry, P.J.; Nourhashemi, F.; Bennahum, D.; Lauque, S.; Albarede, J.-L. The mini nutritional assessment (MNA) and its use in grading the nutritional state of elderly patients. Nutrition 1999, 15, 116–122. [Google Scholar] [CrossRef]
- Howlader, N.N.A.; Krapcho, M.; Miller, D.; Brest, A.; Yu, M.; Ruhl, J.; Tatalovich, Z.; Mariotto, A.; Lewis, D.R.; Chen, H.S.; et al. (Eds.) SEER Cancer Statistics Review; National Cancer Institute: Bethesda, MD, USA, 2019. [Google Scholar]
- Soubeyran, P.; Fonck, M.; Blanc-Bisson, C.; Blanc, J.-F.; Ceccaldi, J.; Mertens, C.; Imbert, Y.; Cany, L.; Vogt, L.; Dauba, J.; et al. Predictors of Early Death Risk in Older Patients Treated with First-Line Chemotherapy for Cancer. J. Clin. Oncol. 2012, 30, 1829–1834. [Google Scholar] [CrossRef]
- Thompson, K.L.; Elliott, L.; Fuchs-Tarlovsky, V.; Levin, R.M.; Voss, A.C.; Piemonte, T. Oncology Evidence-Based Nutrition Practice Guideline for Adults. J. Acad. Nutr. Diet. 2017, 117, 297–310.e47. [Google Scholar] [CrossRef] [PubMed]
- Aaldriks, A.A.; Van Der Geest, L.G.M.; Giltay, E.J.; Le Cessie, S.; Portielje, J.E.; Tanis, B.C.; Nortier, J.W.; Maartense, E. Frailty and malnutrition predictive of mortality risk in older patients with advanced colorectal cancer receiving chemotherapy. J. Geriatr. Oncol. 2013, 4, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Liuu, E.; Saulnier, P.-J.; Gand, E.; Ragot, S.; Valero, S.; Jamet, A.; Hadjadj, S.; Paccalin, M. Frailty and diabetes status in older patients with cancer: Impact on mortality in the ANCRAGE cohort. Aging Clin. Exp. Res. 2020, 32, 1809–1819. [Google Scholar] [CrossRef]
- Kaleru, T.; Vankeshwaram, V.K.; Maheshwary, A.; Mohite, D.; Khan, S. Diabetes Mellitus in the Middle-Aged and Elderly Population (>45 Years) and Its Association with Pancreatic Cancer: An Updated Review. Cureus 2020, 12, e8884. [Google Scholar] [CrossRef] [PubMed]
- Forcados, G.E.; James, D.B.; Sallau, A.B.; Muhammad, A.; Mabeta, P.L. Oxidative Stress and Carcinogenesis: Potential of Phytochemicals in Breast Cancer Therapy. Nutr. Cancer 2017, 69, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.K.; Bin Lee, S.; Won, J.; Choi, H.Y.; Kim, K.; Yang, G.-M.; Dayem, A.A.; Cho, S.G. Correlation between Oxidative Stress, Nutrition, and Cancer Initiation. Int. J. Mol. Sci. 2017, 18, 1544. [Google Scholar] [CrossRef]
- Bryll, A.; Krzyściak, W.; Jurczak, A.; Chrzan, R.; Lizoń, A.; Urbanik, A. Changes in the Selected Antioxidant Defense Parameters in the Blood of Patients after High Resolution Computed Tomography. Int. J. Environ. Res. Public Health 2019, 16, 1476. [Google Scholar] [CrossRef]
- Kozakiewicz, M.; Kornatowski, M.; Krzywińska, O.; Kędziora-Kornatowska, K. Changes in the blood antioxidant defense of advanced age people. Clin. Interv. Aging 2019, 14, 763–771. [Google Scholar] [CrossRef]
- Kushi, L.H.; Doyle, C.; McCullough, M.; Rock, C.L.; Demark-Wahnefried, W.; Bandera, E.V.; Gapstur, S.; Patel, A.V.; Andrews, K.; Gansler, T.; et al. American Cancer Society guidelines on nutrition and physical activity for cancer prevention. CA Cancer J. Clin. 2012, 62, 30–67. [Google Scholar] [CrossRef]
- Norman, H.A.; Butrum, R.R.; Feldman, E.; Heber, D.; Nixon, D.; Picciano, M.F.; Rivlin, R.; Simopoulos, A.; Wargovich, M.J.; Weisburger, E.K.; et al. The Role of Dietary Supplements during Cancer Therapy. J. Nutr. 2003, 133, 3794S–3799S. [Google Scholar] [CrossRef]
- Rock, C.L.; Doyle, C.; Demark-Wahnefried, W.; Meyerhardt, J.; Courneya, K.S.; Schwartz, A.L.; Bandera, E.V.; Hamilton, K.K.; Grant, B.; McCullough, M.; et al. Nutrition and physical activity guidelines for cancer survivors. CA Cancer J. Clin. 2012, 62, 242–274. [Google Scholar] [CrossRef] [PubMed]
- Ambrosone, C.B.; Zirpoli, G.R.; Hutson, A.D.; McCann, W.E.; McCann, S.E.; Barlow, W.E.; Kelly, K.M.; Cannioto, R.; Sucheston-Campbell, L.E.; Hershman, D.L.; et al. Dietary Supplement Use During Chemotherapy and Survival Outcomes of Patients with Breast Cancer Enrolled in a Cooperative Group Clinical Trial (SWOG S0221). J. Clin. Oncol. 2020, 38, 804–814. [Google Scholar] [CrossRef] [PubMed]
- DeWys, W.D.; Begg, C.; Lavin, P.T.; Band, P.R.; Bennett, J.M.; Bertino, J.R.; Cohen, M.H.; Douglass, H.O.; Engstrom, P.F.; Ezdinli, E.Z.; et al. Prognostic effect of weight loss prior tochemotherapy in cancer patients. Am. J. Med. 1980, 69, 491–497. [Google Scholar] [CrossRef]
- Wigmore, S.J.; Plester, C.E.; Ross, J.A.; Fearon, K.C. Contribution of anorexia and hypermetabolism to weight loss in anicteric patients with pancreatic cancer. Br. J. Surg. 1997, 84, 196–197. [Google Scholar]
- Ligibel, J.A.; Alfano, C.M.; Hershman, D.; Ballard, R.M.; Bruinooge, S.S.; Courneya, K.S.; Daniels, E.C.; Demark-Wahnefried, W.; Frank, E.S.; Goodwin, P.J.; et al. Recommendations for Obesity Clinical Trials in Cancer Survivors: American Society of Clinical Oncology Statement. J. Clin. Oncol. 2015, 33, 3961–3967. [Google Scholar] [CrossRef]
- Ligibel, J.A.; Alfano, C.M.; Hershman, D.L.; Merrill, J.K.; Basen-Engquist, K.; Bloomgarden, Z.T.; Demark-Wahnefried, W.; Dixon, S.; Hassink, S.G.; Jakicic, J.M.; et al. American Society of Clinical Oncology Summit on Addressing Obesity Through Multidisciplinary Provider Collaboration: Key Findings and Recommendations for Action. Obesity 2017, 25, S34–S39. [Google Scholar] [CrossRef]
- Wysokiński, A.; Sobow, T.; Kłoszewska, I.; Kostka, T. Mechanisms of the anorexia of aging—A review. AGE 2015, 37. [Google Scholar] [CrossRef]
- Morley, J.E. Anorexia of aging: Physiologic and pathologic. Am. J. Clin. Nutr. 1997, 66, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Brocco, D.; Florio, R.; De Lellis, L.; Veschi, S.; Grassadonia, A.; Tinari, N.; Cama, A. The Role of Dysfunctional Adipose Tissue in Pancreatic Cancer: A Molecular Perspective. Cancers 2020, 12, 1849. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Hontoria, P.L.; Pérez-Matute, P.; Fernández-Galilea, M.; Bustos, M.; Etxeberria, U.; Moreno-Aliaga, M.J. Role of obesity-associated dysfunctional adipose tissue in cancer: A molecular nutrition approach. Biochim. Biophys. Acta (BBA) Bioenerg. 2011, 1807, 664–678. [Google Scholar] [CrossRef] [PubMed]
- Naimo, G.D.; Gelsomino, L.; Catalano, S.; Mauro, L.; Andò, S. Interfering Role of ERα on Adiponectin Action in Breast Cancer. Front. Endocrinol. 2020, 11, 66. [Google Scholar] [CrossRef]
- Cotogni, P.; Caccialanza, R.; Pedrazzoli, P.; Bozzetti, F.; De Francesco, A. Monitoring Response to Home Parenteral Nutrition in Adult Cancer Patients. Health 2020, 8, 183. [Google Scholar] [CrossRef] [PubMed]
- Kanesvaran, R.; Li, H.; Koo, K.-N.; Poon, D. Analysis of Prognostic Factors of Comprehensive Geriatric Assessment and Development of a Clinical Scoring System in Elderly Asian Patients with Cancer. J. Clin. Oncol. 2011, 29, 3620–3627. [Google Scholar] [CrossRef] [PubMed]
- Lagro, J.; Timmer-Bonte, J.; Maas, H.A.A.M. Predictors of Early Death Risk in Older Patients Treated with First-Line Chemotherapy for Cancer and the Importance of Geriatric Assessment. J. Clin. Oncol. 2012, 30, 4443. [Google Scholar] [CrossRef]
- Extermann, M.; Boler, I.; Reich, R.R.; Lyman, G.H.; Brown, R.H.; DeFelice, J.; Levine, R.M.; Lubiner, E.T.; Reyes, P.; Schreiber, F.J.; et al. Predicting the risk of chemotherapy toxicity in older patients: The Chemotherapy Risk Assessment Scale for High-Age Patients (CRASH) score. Cancer 2011, 118, 3377–3386. [Google Scholar] [CrossRef]
- Ferrat, E.; Paillaud, E.; Laurent, M.; Le Thuaut, A.; Caillet, P.; Tournigand, C.; Lagrange, J.-L.; Canouï-Poitrine, F.; Bastuji-Garin, S. Predictors of 1-Year Mortality in a Prospective Cohort of Elderly Patients with Cancer. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2015, 70, 1148–1155. [Google Scholar] [CrossRef]
- Drareni, K.; Bensafi, M.; Giboreau, A.; Dougkas, A. Chemotherapy-induced taste and smell changes influence food perception in cancer patients. Support. Care Cancer 2020, 1–8. [Google Scholar] [CrossRef]
- Donaldson, S.S. Nutritional consequences of radiotherapy. Cancer Res. 1977, 37, 2407–2413. [Google Scholar]
- Arends, J.J.; Baracos, V.V.; Bertz, H.H.; Bozzetti, F.; Calder, P.P.; Deutz, N.; Erickson, N.N.; Laviano, A.A.; Lisanti, M.M.; Lobo, D.N.D.; et al. ESPEN expert group recommendations for action against cancer-related malnutrition. Clin. Nutr. 2017, 36, 1187–1196. [Google Scholar] [CrossRef]
- Peñas, R.D.L.; Majem, M.; Perez-Altozano, J.; Virizuela, J.A.; Cancer, E.; Diz, P.; Donnay, O.; Hurtado, A.; Jimenez-Fonseca, P.; Ocon, M.J. SEOM clinical guidelines on nutrition in cancer patients (2018). Clin. Transl. Oncol. 2019, 21, 87–93. [Google Scholar] [CrossRef]
- Hoppe, S.; Rainfray, M.; Fonck, M.; Hoppenreys, L.; Blanc, J.-F.; Ceccaldi, J.; Mertens, C.; Blanc-Bisson, C.; Imbert, Y.; Cany, L.; et al. Functional Decline in Older Patients with Cancer Receiving First-Line Chemotherapy. J. Clin. Oncol. 2013, 31, 3877–3882. [Google Scholar] [CrossRef] [PubMed]
- Georlee, G.M.; Abiram, U.; Dat, P.N.; Tuan, N.K.; Mehrotra, S. Home-modification interventions addressing falls and participation in activities of daily living among older adults: A scoping review protocol. BMJ Open 2020, 10, e039742. [Google Scholar] [CrossRef] [PubMed]
- Novelli, I.R.; Araújo, B.A.D.; Grandisoli, L.F.; Furtado, E.C.G.; Aguchiku, E.K.N.; Bertocco, M.C.G.; Sudbrak, T.P.; De Araújo, I.C.; Bosko, A.C.F.; Damasceno, N.R.T. Nutritional Counseling Protocol for Colorectal Cancer Patients after Surgery Improves Outcome. Nutr. Cancer 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Paleri, V.; Urbano, T.G.; Mehanna, H.; Repanos, C.; Lancaster, J.; Roques, T.; Patel, M.; Sen, M. Management of neck metastases in head and neck cancer: United Kingdom National Multidisciplinary Guidelines. J. Laryngol. Otol. 2016, 130, S161–S169. [Google Scholar] [CrossRef]
- Presley, C.J.; Krok-Schoen, J.L.; Wall, S.A.; Noonan, A.M.; Jones, D.C.; Folefac, E.; Williams, N.; Overcash, J.; Rosko, A.E. Implementing a multidisciplinary approach for older adults with Cancer: Geriatric oncology in practice. BMC Geriatr. 2020, 20, 1–9. [Google Scholar] [CrossRef]
- Yang, Z.-H.; Dang, Y.-Q.; Ji, G. Role of epigenetics in transformation of inflammation into colorectal cancer. World J. Gastroenterol. 2019, 25, 2863–2877. [Google Scholar] [CrossRef]
- Koch, A.; Joosten, S.C.; Feng, Z.; De Ruijter, T.C.; Draht, M.X.; Melotte, V.; Smits, K.M.; Veeck, J.; Herman, J.G.; Van Neste, L.; et al. Analysis of DNA methylation in cancer: Location revisited. Nat. Rev. Clin. Oncol. 2018, 15, 459–466. [Google Scholar] [CrossRef]
- Bell, C.G.; Lowe, R.; Adams, P.D.; Baccarelli, A.A.; Beck, S.; Bell, J.T.; Christensen, B.C.; Gladyshev, V.N.; Heijmans, B.T.; Horvath, S.; et al. DNA methylation aging clocks: Challenges and recommendations. Genome Biol. 2019, 20, 1–24. [Google Scholar] [CrossRef]
- Zheng, Y.; Joyce, B.T.; Liu, L.; Zhang, Z.; Kibbe, W.A.; Zhang, W.; Hou, L. Prediction of genome-wide DNA methylation in repetitive elements. Nucleic Acids Res. 2017, 45, 8697–8711. [Google Scholar] [CrossRef]
- Demetriadou, C.; Koufaris, C.; Kirmizis, A. Histone N-alpha terminal modifications: Genome regulation at the tip of the tail. Epigenetics Chromatin 2020, 13, 1–13. [Google Scholar] [CrossRef]
- Ferrand, J.; Plessier, A.; Polo, S.E. Control of the chromatin response to DNA damage: Histone proteins pull the strings. Semin. Cell Dev. Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.G.; Allis, C.D.; Chi, P. Chromatin remodeling and cancer, part I: Covalent histone modifications. Trends Mol. Med. 2007, 13, 363–372. [Google Scholar] [CrossRef]
- Slack, F.J.; Chinnaiyan, A.M. The Role of Non-coding RNAs in Oncology. Cell 2019, 179, 1033–1055. [Google Scholar] [CrossRef] [PubMed]
- Fuke, C.; Shimabukuro, M.; Petronis, A.; Sugimoto, J.; Oda, T.; Miura, K.; Miyazaki, T.; Ogura, C.; Okazaki, Y.; Jinno, Y. Age Related Changes in 5-methylcytosine Content in Human Peripheral Leukocytes and Placentas: An HPLC-based Study. Ann. Hum. Genet. 2004, 68, 196–204. [Google Scholar] [CrossRef]
- Mays-Hoopes, L.L. DNA Methylation in Aging and Cancer. J. Gerontol. 1989, 44, 35–36. [Google Scholar] [CrossRef]
- Feinberg, A.P.; Vogelstein, B. Hypomethylation distinguishes genes of some human cancers from their normal counterparts. Nat. Cell Biol. 1983, 301, 89–92. [Google Scholar] [CrossRef]
- Bennett-Baker, P.E.; Wilkowski, J.; Burke, D.T. Age-associated activation of epigenetically repressed genes in the mouse. Genetics 2003, 165, 2055–2062. [Google Scholar]
- Ahuja, N.; Issa, J.P. Aging, methylation and cancer. Histol. Histopathol. 2000, 15, 835–842. [Google Scholar]
- Issa, J.-P. Epigenetic variation and human disease. J. Nutr. 2002, 132, 2388S–2392S. [Google Scholar] [CrossRef]
- Issa, J.-P. Age-related epigenetic changes and the immune system. Clin. Immunol. 2003, 109, 103–108. [Google Scholar] [CrossRef]
- Issa, J.-P.; Ahuja, N.; Toyota, M.; Bronner, M.P.; Brentnall, T.A. Accelerated age-related CpG island methylation in ulcerative colitis. Cancer Res. 2001, 61, 3573–3577. [Google Scholar] [PubMed]
- Issa, J.-P.J.; Ottaviano, Y.L.; Celano, P.; Hamilton, S.R.; Davidson, N.E.; Baylin, S.B. Methylation of the oestrogen receptor CpG island links ageing and neoplasia in human colon. Nat. Genet. 1994, 7, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Ciccarone, F.; Tagliatesta, S.; Caiafa, P.; Zampieri, M. DNA methylation dynamics in aging: How far are we from understanding the mechanisms? Mech. Ageing Dev. 2018, 174, 3–17. [Google Scholar] [CrossRef]
- Pinel, C.; Prainsack, B.; McKevitt, C. Markers as mediators: A review and synthesis of epigenetics literature. BioSocieties 2017, 13, 276–303. [Google Scholar] [CrossRef]
- Fang, M.Z.; Wang, Y.; Ai, N.; Hou, Z.; Sun, Y.; Lu, H.; Welsh, W.; Yang, C.S. Tea polyphenol (-)-epigallocatechin-3-gallate inhibits DNA methyltransferase and reactivates methylation-silenced genes in cancer cell lines. Cancer Res. 2003, 63, 7563–7570. [Google Scholar]
- Li, Y.; Liu, L.; Tollefsbol, T.O. Glucose restriction can extend normal cell lifespan and impair precancerous cell growth through epigenetic control of hTERT and p16 expression. FASEB J. 2009, 24, 1442–1453. [Google Scholar] [CrossRef]
- Blasiak, J. Senescence in the pathogenesis of age-related macular degeneration. Cell. Mol. Life Sci. 2020, 77, 789–805. [Google Scholar] [CrossRef]
- Arancio, W.; Pizzolanti, G.; Genovese, S.I.; Pitrone, M.; Giordano, C. Epigenetic Involvement in Hutchinson-Gilford Progeria Syndrome: A Mini-Review. Gerontology 2014, 60, 197–203. [Google Scholar] [CrossRef]
- Burtner, C.R.; Kennedy, B.K. Progeria syndromes and ageing: What is the connection? Nat. Rev. Mol. Cell Biol. 2010, 11, 567–578. [Google Scholar] [CrossRef]
- Pal, S.; Tyler, J.K. Epigenetics and aging. Sci. Adv. 2016, 2, e1600584. [Google Scholar] [CrossRef]
- Pegoraro, G.; Kubben, N.; Wickert, U.; Göhler, H.; Hoffmann, K.; Misteli, T. Ageing-related chromatin defects through loss of the NURD complex. Nat. Cell Biol. 2009, 11, 1261–1267. [Google Scholar] [CrossRef] [PubMed]
- Degirmenci, U.; Lei, S. Role of lncRNAs in Cellular Aging. Front. Endocrinol. 2016, 7, 151. [Google Scholar] [CrossRef]
- Jin, L.; Song, Q.; Zhang, W.; Geng, B.; Cai, J. Roles of long noncoding RNAs in aging and aging complications. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2019, 1865, 1763–1771. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Chen, X.; Inukai, S.; Zhao, H.; Slack, F.J. Age-associated changes in expression of small, noncoding RNAs, including microRNAs, in C. elegans. RNA 2011, 17, 1804–1820. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Slack, F.J. Ageing and the small, non-coding RNA world. Ageing Res. Rev. 2013, 12, 429–435. [Google Scholar] [CrossRef]
- Heid, J.; Cencioni, C.; Ripa, R.; Baumgart, M.; Atlante, S.; Milano, G.; Scopece, A.; Kuenne, C.; Guenther, S.; Azzimato, V.; et al. Age-dependent increase of oxidative stress regulates microRNA-29 family preserving cardiac health. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef]
- Raz, V.; Kroon, R.H.M.J.M.; Mei, H.; Riaz, M.; Buermans, H.; Lassche, S.; Horlings, C.; De Swart, B.; Kalf, J.; Harish, P.; et al. Age-Associated Salivary MicroRNA Biomarkers for Oculopharyngeal Muscular Dystrophy. Int. J. Mol. Sci. 2020, 21, 6059. [Google Scholar] [CrossRef]
- Turunen, T.A.; Roberts, T.C.; Laitinen, P.; Väänänen, M.-A.; Korhonen, P.; Malm, T.; Ylä-Herttuala, S.; Turunen, M.P. Changes in nuclear and cytoplasmic microRNA distribution in response to hypoxic stress. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Ford, D. Honeybees and cell lines as models of DNA methylation and aging in response to diet. Exp. Gerontol. 2013, 48, 614–619. [Google Scholar] [CrossRef]
- Ford, D.; Ions, L.J.; Alatawi, F.; Wakeling, L.A. The potential role of epigenetic responses to diet in ageing. Proc. Nutr. Soc. 2011, 70, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.J.; Robertson, N.A.; Rather, M.I.; Thomson, J.P.; McBryan, T.; Sproul, D.; Wang, T.; Brock, C.; Clark, W.; Ideker, T.; et al. Diverse interventions that extend mouse lifespan suppress shared age-associated epigenetic changes at critical gene regulatory regions. Genome Biol. 2017, 18, 1–16. [Google Scholar] [CrossRef]
- Longo, V.D.; Antebi, A.; Bartke, A.; Barzilai, N.; Brown-Borg, H.M.; Caruso, C.; Curiel, T.J.; De Cabo, R.; Franceschi, C.; Gems, D.; et al. Interventions to Slow Aging in Humans: Are We Ready? Aging Cell 2015, 14, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Mercken, E.M.; Carboneau, B.A.; Krzysik-Walker, S.M.; De Cabo, R. Of mice and men: The benefits of caloric restriction, exercise, and mimetics. Ageing Res. Rev. 2012, 11, 390–398. [Google Scholar] [CrossRef] [PubMed]
- McCay, C.M.; Crowell, M.F.; Maynard, L.A. The Effect of Retarded Growth Upon the Length of Life Span and Upon the Ultimate Body Size. J. Nutr. 1935, 10, 63–79. [Google Scholar] [CrossRef]
- Daniel, M.; Tollefsbol, T.O. Epigenetic linkage of aging, cancer and nutrition. J. Exp. Biol. 2015, 218, 59–70. [Google Scholar] [CrossRef]
- Colman, R.J.; Anderson, R.M.; Johnson, S.C.; Kastman, E.K.; Kosmatka, K.J.; Beasley, T.M.; Allison, D.B.; Cruzen, C.; Simmons, H.A.; Kemnitz, J.W.; et al. Caloric Restriction Delays Disease Onset and Mortality in Rhesus Monkeys. Science 2009, 325, 201–204. [Google Scholar] [CrossRef]
- Colman, R.J.; Beasley, T.M.; Kemnitz, J.W.; Johnson, S.C.; Weindruch, R.; Anderson, R.M. Caloric restriction reduces age-related and all-cause mortality in rhesus monkeys. Nat. Commun. 2014, 5, 3557. [Google Scholar] [CrossRef]
- Mattison, J.A.; Roth, G.S.; Beasley, T.M.; Tilmont, E.M.; Handy, A.M.; Herbert, R.L.; Longo, D.L.; Allison, D.B.; Young, J.E.; Bryant, M.; et al. Impact of caloric restriction on health and survival in rhesus monkeys from the NIA study. Nat. Cell Biol. 2012, 489, 318–321. [Google Scholar] [CrossRef]
- Hwangbo, D.-S.; Lee, H.-Y.; Abozaid, L.S.; Min, K.-J. Mechanisms of Lifespan Regulation by Calorie Restriction and Intermittent Fasting in Model Organisms. Nutrients 2020, 12, 1194. [Google Scholar] [CrossRef]
- Cantó, C.; Auwerx, J. Calorie restriction: Is AMPK a key sensor and effector? Physiology 2011, 26, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Wang, R.; Wang, H.; Zhang, Y.; Zhao, Z. Long-term caloric restriction activates the myocardial SIRT1/AMPK/PGC-1α pathway in C57BL/6J male mice. Food Nutr. Res. 2020, 64, 64. [Google Scholar] [CrossRef] [PubMed]
- Zullo, A.; Simone, E.; Grimaldi, M.; Musto, V.; Mancini, F.P. Sirtuins as Mediator of the Anti-Ageing Effects of Calorie Restriction in Skeletal and Cardiac Muscle. Int. J. Mol. Sci. 2018, 19, 928. [Google Scholar] [CrossRef]
- Ng, G.Y.-Q.; Fann, D.Y.-W.; Jo, D.-G.; Sobey, C.G.; Arumugam, T.V. Dietary Restriction and Epigenetics: Part I. Cond. Med. 2019, 2, 284–299. [Google Scholar]
- Maegawa, S.; Lu, Y.; Tahara, T.; Lee, J.T.; Madzo, J.; Liang, S.; Jelinek, J.; Colman, R.J.; Issa, J.-P.J. Caloric restriction delays age-related methylation drift. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hernando-Herraez, I.; Evano, B.; Stubbs, T.; Commere, P.-H.; Bonder, M.J.; Clark, S.; Andrews, S.; Tajbakhsh, S.; Reik, W. Ageing affects DNA methylation drift and transcriptional cell-to-cell variability in mouse muscle stem cells. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gensous, N.; Franceschi, C.; Santoro, A.; Milazzo, M.; Garagnani, P.; Bacalini, M.G. The Impact of Caloric Restriction on the Epigenetic Signatures of Aging. Int. J. Mol. Sci. 2019, 20, 2022. [Google Scholar] [CrossRef]
- Hadad, N.; Unnikrishnan, A.; Jackson, J.A.; Masser, D.R.; Otalora, L.; Stanford, D.R.; Richardson, A.; Freeman, W.M. Caloric restriction mitigates age-associated hippocampal differential CG and non-CG methylation. Neurobiol. Aging 2018, 67, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Ng, G.Y.-Q.; Fann, D.Y.-W.; Jo, D.-G.; Sobey, C.G.; Arumugam, T.V. Epigenetic Regulation by Dietary Restriction: Part II. Cond. Med. 2019, 2, 300–310. [Google Scholar]
- Weber, D.D.; Aminazdeh-Gohari, S.; Kofler, B. Ketogenic diet in cancer therapy. Aging 2018, 10, 164–165. [Google Scholar] [CrossRef] [PubMed]
- Cullingford, T.E. The ketogenic diet; fatty acids, fatty acid-activated receptors and neurological disorders. Prostaglandins Leukot. Essent. Fat. Acids 2004, 70, 253–264. [Google Scholar] [CrossRef]
- Shimazu, T.; Hirschey, M.D.; Newman, J.; He, W.; Shirakawa, K.; Le Moan, N.; Grueter, C.A.; Lim, H.; Saunders, L.R.; Stevens, R.D.; et al. Suppression of Oxidative Stress by -Hydroxybutyrate, an Endogenous Histone Deacetylase Inhibitor. Science 2012, 339, 211–214. [Google Scholar] [CrossRef]
- Veech, R.L.; Chance, B.; Kashiwaya, Y.; Lardy, H.A.; Cahill, G.F. Ketone Bodies, Potential Therapeutic Uses. IUBMB Life 2001, 51, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Bozzetti, F.; Stanga, Z. Does nutrition for cancer patients feed the tumour? A clinical perspective. Crit. Rev. Oncol. 2020, 153, 103061. [Google Scholar] [CrossRef] [PubMed]
- Hardy, T.M.; Tollefsbol, T.O. Epigenetic diet: Impact on the epigenome and cancer. Epigenomics 2011, 3, 503–518. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Shishodia, S. Molecular targets of dietary agents for prevention and therapy of cancer. Biochem. Pharmacol. 2006, 71, 1397–1421. [Google Scholar] [CrossRef]
- Ayissi, V.B.O.; Ebrahimi, A.; Schluesenner, H. Epigenetic effects of natural polyphenols: A focus on SIRT1-mediated mechanisms. Mol. Nutr. Food Res. 2014, 58, 22–32. [Google Scholar] [CrossRef]
- Hurria, A.; Wildes, T.; Blair, S.L.; Browner, I.S.; Cohen, H.J.; DeShazo, M.; Dotan, E.; Edil, B.H.; Extermann, M.; Ganti, A.K.P.; et al. Senior Adult Oncology, Version 2.2014. J. Natl. Compr. Cancer Netw. 2014, 12, 82–126. [Google Scholar] [CrossRef]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Signal Transduct. Target. Ther. 2020, 5, 1–17. [Google Scholar] [CrossRef]
- Carlos-Reyes, Á.; López-González, J.S.; Meneses-Flores, M.; Gallardo-Rincón, D.; Ruíz-García, E.; Marchat, L.A.; La Vega, H.A.-D.; De La Cruz, O.N.H.; López-Camarillo, C. Dietary Compounds as Epigenetic Modulating Agents in Cancer. Front. Genet. 2019, 10, 79. [Google Scholar] [CrossRef]
- Fila, M.; Chojnacki, C.; Chojnacki, J.; Blasiak, J. Is an “Epigenetic Diet” for Migraines Justified? The Case of Folate and DNA Methylation. Nutrients 2019, 11, 2763. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blasiak, J.; Chojnacki, J.; Pawlowska, E.; Szczepanska, J.; Chojnacki, C. Nutrition in Cancer Therapy in the Elderly—An Epigenetic Connection? Nutrients 2020, 12, 3366. https://doi.org/10.3390/nu12113366
Blasiak J, Chojnacki J, Pawlowska E, Szczepanska J, Chojnacki C. Nutrition in Cancer Therapy in the Elderly—An Epigenetic Connection? Nutrients. 2020; 12(11):3366. https://doi.org/10.3390/nu12113366
Chicago/Turabian StyleBlasiak, Janusz, Jan Chojnacki, Elzbieta Pawlowska, Joanna Szczepanska, and Cezary Chojnacki. 2020. "Nutrition in Cancer Therapy in the Elderly—An Epigenetic Connection?" Nutrients 12, no. 11: 3366. https://doi.org/10.3390/nu12113366
APA StyleBlasiak, J., Chojnacki, J., Pawlowska, E., Szczepanska, J., & Chojnacki, C. (2020). Nutrition in Cancer Therapy in the Elderly—An Epigenetic Connection? Nutrients, 12(11), 3366. https://doi.org/10.3390/nu12113366