Vitamins and Minerals for Energy, Fatigue and Cognition: A Narrative Review of the Biochemical and Clinical Evidence


{kind=link}
{kind=link}
Abstract
1. Introduction
2. Energy and Fatigue Are Subjective Perceptions Supported by an Objective Physiological Basis
2.1. Different Definitions Exist for Concepts Such as ‘Energy’ or ‘Fatigue’
2.2. Fatigue, Lack of Energy and Lower Physical and Cognitive Performance
2.3. What Are the Physiological Bases of Physical and Mental Fatigue
2.4. Fatigue May Also Result from A Dysfunction in The Supply of Oxygen to Muscles and Brain
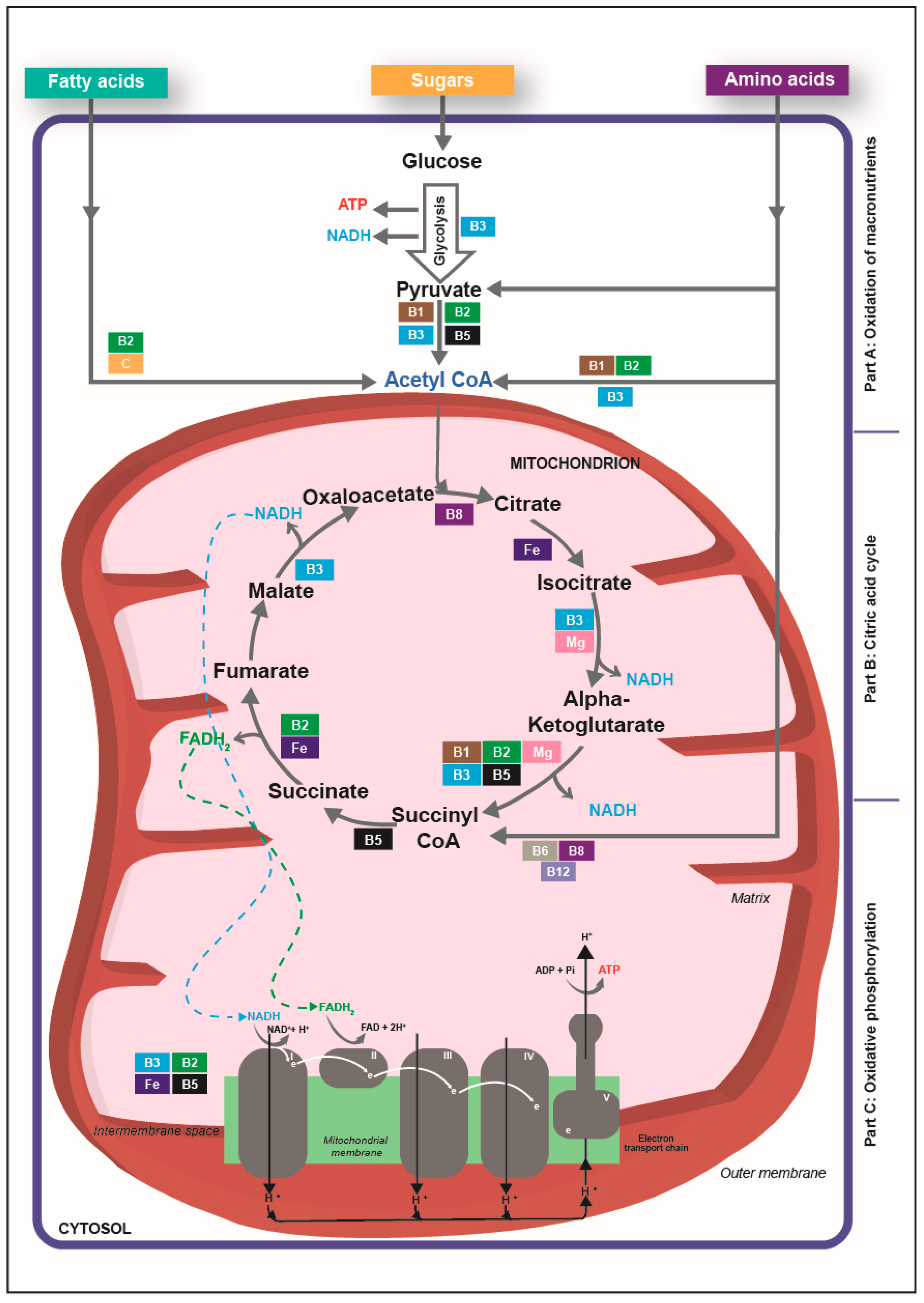
3. Vitamins and Minerals Have Critical Roles in Cellular Energy Production
3.1. Overview of Cellular Energy Production
3.2. The Interplay of B Vitamins in Cellular Energy Production
3.3. Vitamin C, Iron and Magnesium Are Also involved in Energy-Yielding Metabolism
4. Vitamins and Minerals Are Important for Regulating Oxygen in the Body
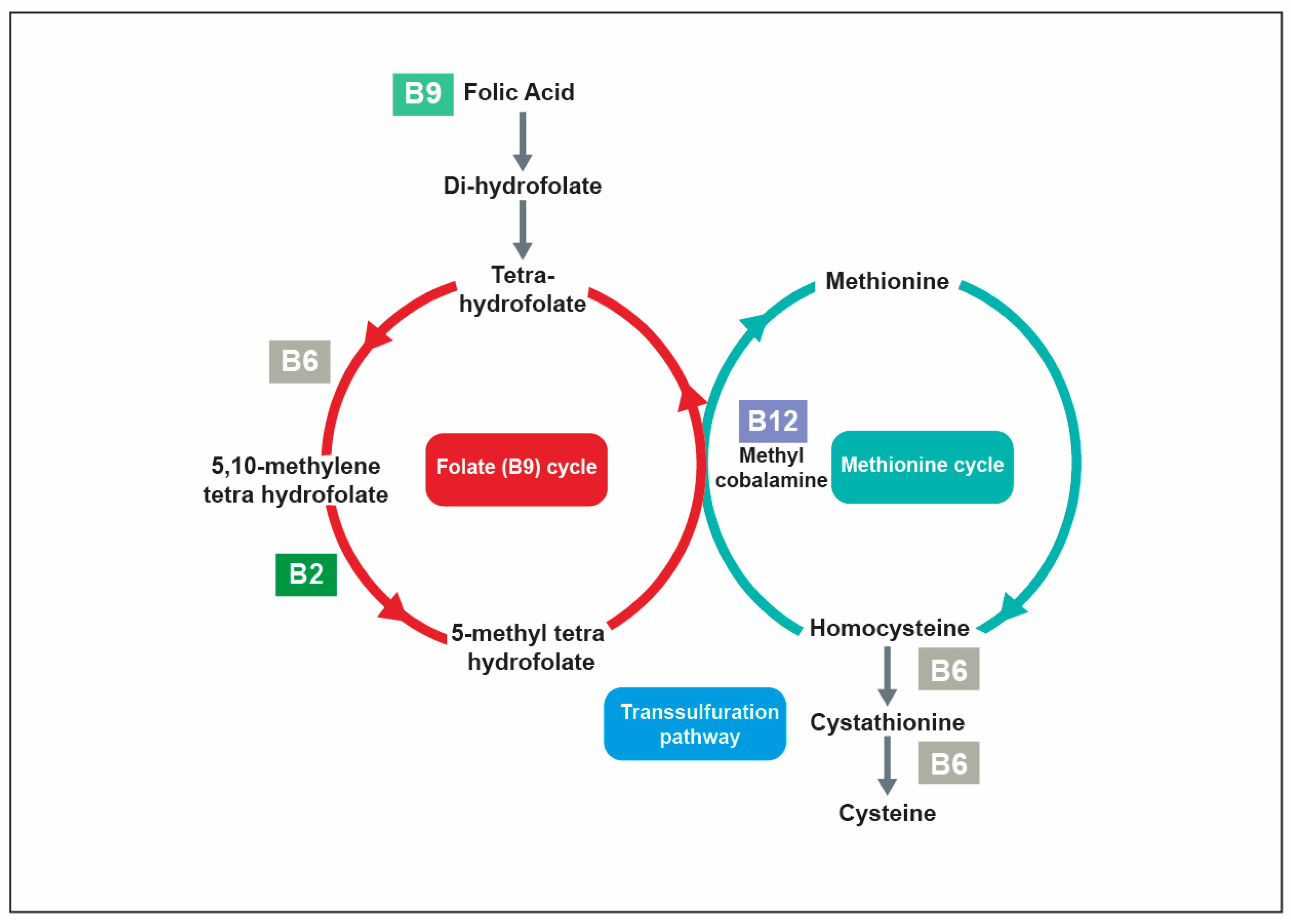
4.1. Oxygen Transport Requires Iron and Vitamins B6, B9 and B12
4.2. Riboflavin, Vitamin C, Iron, Magnesium and Zinc and Oxidative Stress
5. Vitamins and Minerals Are Critical for the Structure and Function of Brain Cells
5.1. Vitamins and Minerals Are Involved in Neuronal Structures
5.2. The Synthesis of Neurotransmitters Is Dependent on Vitamins B1, B5, B6, B9 and C
5.3. Vitamins B3 and B5, Iron, Magnesium and Zinc Are Important for Neurotransmission
6. Clinical Evidence of the Role of Vitamins and Minerals on Physical Fatigue
6.1. Impact of Inadequate Status of Vitamins and Minerals on Physical Fatigue
6.1.1. Inadequate Status of Individual B Vitamins and Physical Fatigue
6.1.2. Inadequate Status in Vitamin C and Physical Fatigue
6.1.3. Consequences of Inadequate Status in Iron and Magnesium on Physical Fatigue
6.2. Effects of Supplementation with Vitamins and Minerals on Physical Fatigue
6.2.1. Supplementation with Individual B Vitamins and Physical Fatigue
6.2.2. Supplementation with Vitamin C and Physical Fatigue
6.2.3. Supplementation with Iron and Magnesium and Physical Fatigue
7. Clinical Evidence of the Role of Vitamins and Minerals on Mental Fatigue and Cognitive or Psychological Functions
7.1. Impact of Vitamins and Minerals Frank Deficiencies on Mental Fatigue and Cognitive or Psychological Functions
7.1.1. Deficiencies in Individual B Vitamins and Mental Fatigue and Cognitive or Psychological Functions
7.1.2. Deficiency in Vitamin C and Minerals and Mental Fatigue and Cognitive or Psychological Functions
7.2. Relationships between Vitamin and Mineral Dietary Intake or Status and Mental Fatigue and Cognitive or Psychological Functions
7.2.1. Intake or Status in Individual B Vitamin and Mental Fatigue and Cognitive or Psychological Functions
7.2.2. Intake or Status in Vitamin C, Iron, Magnesium and Zinc and Mental Fatigue and Cognitive or Psychological Function
7.3. Effect of Supplementation with Vitamins and Minerals on Mental Fatigue and Cognitive or Psychological Functions
7.3.1. Supplementation with Individual B Vitamins on Mental Fatigue and Cognitive or Psychological Functions
7.3.2. Supplementation with Vitamin C, Iron, Magnesium or Zinc and Mental Fatigue and Cognitive or Psychological Functions
7.3.3. Effect of Supplementation with Multiple Micronutrients on Mental Fatigue and Cognitive or Psychological Functions
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- EFSA. Scientific Opinion on principles for deriving and applying Dietary Reference Values. EFSA J. 2010, 8, 1458. [Google Scholar]
- European Commission. Commission Regulation (EU) No 432/2012 of 16 May 2012 establishing a list of permitted health claims made on foods, other than those referring to the reduction of disease risk and to children’s development and health. Off. J. Eur. Union 2012, 136, 1–40. [Google Scholar]
- Griffiths, J.K. Vitamin deficiencies. In Hunter’s Tropical Medicine and Emerging Infectious Disease, 9th ed.; Magil, A.J., Hill, D.R., Solomon, T., Ryan, E.T., Eds.; Elsevier: London, UK, 2013; pp. 997–1002. [Google Scholar]
- Rippin, H.L.; Hutchinson, J.; Jewell, J.; Breda, J.J.; Cade, J.E. Adult Nutrient Intakes from Current National Dietary Surveys of European Populations. Nutrients 2017, 9, 1288. [Google Scholar] [CrossRef] [PubMed]
- Pedroza-Tobias, A.; Hernandez-Barrera, L.; Lopez-Olmedo, N.; Garcia-Guerra, A.; Rodriguez-Ramirez, S.; Ramirez-Silva, I.; Villalpando, S.; Carriquiry, A.; Rivera, J.A. Usual Vitamin Intakes by Mexican Populations. J. Nutr. 2016, 146, 1866S–1873S. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Health of Turkey. Dietary Guidelines. Publication n°1046; Ministry of Health of Turkey: Ankara, Turkey. Available online: https://dosyasb.saglik.gov.tr/Eklenti/10922,17ocaktuberingilizcepdf.pdf?0 (accessed on 2 January 2020).
- Dietary Guidelines Advisory Committee. Scientific Report of the 2015 Dietary Guidelines Advisory Committee: Advisory Report to the Secretary of Health and Human Services and the Secretary of Agriculture; U.S. Department of Agriculture, Agricultural Research Service: Washington, DC, USA, 2015.
- Bailey, R.L.; Gahche, J.J.; Miller, P.E.; Thomas, P.R.; Dwyer, J.T. Why US adults use dietary supplements. JAMA Int. Med. 2013, 173, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Sekhri, K.; Kaur, K. Public knowledge, use and attitude toward multivitamin supplementation: A cross-sectional study among general public. Int. J. Appl. Basic Med. Res. 2014, 4, 77–80. [Google Scholar] [CrossRef]
- Ryan, R.M.; Frederick, C. On energy, personality, and health: Subjective vitality as a dynamic reflection of well-being. J. Pers. 1997, 65, 529–565. [Google Scholar] [CrossRef]
- Aouizerat, B.E.; Gay, C.L.; Lerdal, A.; Portillo, C.J.; Lee, K.A. Lack of energy: An important and distinct component of HIV-related fatigue and daytime function. J. Pain Symptom Manag. 2013, 45, 191–201. [Google Scholar] [CrossRef]
- van Steenbergen, E.; van Dongen, J.M.; Wendel-Vos, G.C.; Hildebrandt, V.H.; Strijk, J.E. Insights into the concept of vitality: Associations with participation and societal costs. Eur. J. Public Health 2016, 26, 354–359. [Google Scholar] [CrossRef]
- Gledhill, J. A qualitative study of the characteristics and representation of fatigue in a French speaking population of cancer patients and healthy subjects. Eur. J. Oncol. Nurs. 2005, 9, 294–312, discussion 294–312. [Google Scholar] [CrossRef]
- Engberg, I.; Segerstedt, J.; Waller, G.; Wennberg, P.; Eliasson, M. Fatigue in the general population-associations to age, sex, socioeconomic status, physical activity, sitting time and self-rated health: The northern Sweden MONICA study 2014. BMC Public Health 2017, 17, 654. [Google Scholar] [CrossRef] [PubMed]
- Enoka, R.M.; Duchateau, J. Translating Fatigue to Human Performance. Med. Sci. Sports Exerc. 2016, 48, 2228–2238. [Google Scholar] [CrossRef] [PubMed]
- Bucher, E.; Sandbakk, O.; Donath, L.; Roth, R.; Zahner, L.; Faude, O. Exercise-induced trunk fatigue decreases double poling performance in well-trained cross-country skiers. Eur. J. Appl. Physiol. 2018, 118, 2077–2087. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.F.; Lee, W.C.; Chen, Y.A.; Hsue, B.J. Fatigue-Induced Changes in Movement Pattern and Muscle Activity During Ballet Releve on Demi-Pointe. J. Appl. Biomech. 2016, 32, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Le Mansec, Y.; Pageaux, B.; Nordez, A.; Dorel, S.; Jubeau, M. Mental fatigue alters the speed and the accuracy of the ball in table tennis. J. Sports Sci. 2018, 36, 2751–2759. [Google Scholar] [CrossRef]
- Hanna, T.N.; Zygmont, M.E.; Peterson, R.; Theriot, D.; Shekhani, H.; Johnson, J.O.; Krupinski, E.A. The Effects of Fatigue From Overnight Shifts on Radiology Search Patterns and Diagnostic Performance. J. Am. Coll. Radiol. 2018, 15, 1709–1716. [Google Scholar] [CrossRef]
- Mollicone, D.; Kan, K.; Mott, C.; Bartels, R.; Bruneau, S.; van Wollen, M.; Sparrow, A.R.; Van Dongen, H.P.A. Predicting performance and safety based on driver fatigue. Accid. Anal. Prev. 2019, 126, 142–145. [Google Scholar] [CrossRef]
- Rudin-Brown, C.M.; Harris, S.; Rosberg, A. How shift scheduling practices contribute to fatigue amongst freight rail operating employees: Findings from Canadian accident investigations. Accid. Anal. Prev. 2019, 126, 64–69. [Google Scholar] [CrossRef]
- Sievertsen, H.H.; Gino, F.; Piovesan, M. Cognitive fatigue influences students’ performance on standardized tests. Proc. Natl. Acad. Sci. USA 2016, 113, 2621–2624. [Google Scholar] [CrossRef]
- Mizuno, K.; Tanaka, M.; Fukuda, S.; Imai-Matsumura, K.; Watanabe, Y. Relationship between cognitive functions and prevalence of fatigue in elementary and junior high school students. Brain Dev. 2011, 33, 470–479. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, G.; Wang, Y.; Ai, Y.; Pinto-Martin, J.; Liu, X. Sleep problems, fatigue, and cognitive performance in Chinese kindergarten children. J. Pediatr. 2012, 161, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ying, Z.; Bosy-Westphal, A.; Zhang, J.; Schautz, B.; Later, W.; Heymsfield, S.B.; Muller, M.J. Specific metabolic rates of major organs and tissues across adulthood: Evaluation by mechanistic model of resting energy expenditure. Am. J. Clin. Nutr. 2010, 92, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Sahlin, K.; Tonkonogi, M.; Soderlund, K. Energy supply and muscle fatigue in humans. Acta Physiol. Scand. 1998, 162, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Sargeant, A.J. Structural and functional determinants of human muscle power. Exp. Physiol. 2007, 92, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Herculano-Houzel, S. Scaling of brain metabolism with a fixed energy budget per neuron: Implications for neuronal activity, plasticity and evolution. PLoS ONE 2011, 6, e17514. [Google Scholar] [CrossRef]
- Harris, J.J.; Jolivet, R.; Attwell, D. Synaptic energy use and supply. Neuron 2012, 75, 762–777. [Google Scholar] [CrossRef]
- Bordone, M.P.; Salman, M.M.; Titus, H.E.; Amini, E.; Andersen, J.V.; Chakraborti, B.; Diuba, A.V.; Dubouskaya, T.G.; Ehrke, E.; Freitas, A.E.; et al. The energetic brain—A review from students to students. J. Neurochem. 2019, 151, 139–165. [Google Scholar] [CrossRef]
- Raichle, M.E. The restless brain: How intrinsic activity organizes brain function. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef]
- Petit, J.M.; Magistretti, P.J. Regulation of neuron-astrocyte metabolic coupling across the sleep-wake cycle. Neuroscience 2016, 323, 135–156. [Google Scholar] [CrossRef]
- Hogan, A.M.; Pit-ten Cate, I.M.; Vargha-Khadem, F.; Prengler, M.; Kirkham, F.J. Physiological correlates of intellectual function in children with sickle cell disease: Hypoxaemia, hyperaemia and brain infarction. Dev. Sci. 2006, 9, 379–387. [Google Scholar] [CrossRef]
- Rink, C.; Khanna, S. Significance of brain tissue oxygenation and the arachidonic acid cascade in stroke. Antioxid. Redox Signal. 2011, 14, 1889–1903. [Google Scholar] [CrossRef] [PubMed]
- Mairbaurl, H. Red blood cells in sports: Effects of exercise and training on oxygen supply by red blood cells. Front. Physiol. 2013, 4, 332. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.L.; Jonassaint, C.; Sharrett, A.R.; Mosley, T.H.; Astor, B.C.; Selvin, E.; Coresh, J.; Gottesman, R.F. Hemoglobin, Anemia, and Cognitive Function: The Atherosclerosis Risk in Communities Study. J. Gerontol. A Biol. Sci. Med. Sci. 2016, 71, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Acaster, S.; Dickerhoof, R.; DeBusk, K.; Bernard, K.; Strauss, W.; Allen, L.F. Qualitative and quantitative validation of the FACIT-fatigue scale in iron deficiency anemia. Health Qual. Life Outcomes 2015, 13, 60. [Google Scholar] [CrossRef]
- Scholey, A.B.; Harper, S.; Kennedy, D.O. Cognitive demand and blood glucose. Physiol. Behav. 2001, 73, 585–592. [Google Scholar] [CrossRef]
- Scholey, A.B.; Laing, S.; Kennedy, D.O. Blood glucose changes and memory: Effects of manipulating emotionality and mental effort. Biol. Psychol. 2006, 71, 12–19. [Google Scholar] [CrossRef]
- Fairclough, S.H.; Houston, K. A metabolic measure of mental effort. Biol. Psychol. 2004, 66, 177–190. [Google Scholar] [CrossRef]
- Scholey, A.B.; Benson, S.; Sela-Venter, S.; Mackus, M.; Moss, M.C. Oxygen Administration and Acute Human Cognitive Enhancement: Higher Cognitive Demand Leads to a More Rapid Decay of Transient Hyperoxia. J. Cogn. Enhanc. 2019, 71, 1–6. [Google Scholar] [CrossRef]
- Sobrero, A.; Puglisi, F.; Guglielmi, A.; Belvedere, O.; Aprile, G.; Ramello, M.; Grossi, F. Fatigue: A main component of anemia symptomatology. Semin. Oncol. 2001, 28, 15–18. [Google Scholar] [CrossRef]
- Butte, N.F.; Caballero, B. Energy needs: Assessment and requirements. In Modern Nutrition in Health and Disease, 11th ed.; Ross, A.C., Caballero, B., Cousins, R.J., Tucker, K.L., Ziegler, T.R., Eds.; Wolters Kluwer-Lippincott, Williams & Wilkins: Baltimore, MD, USA, 2014; pp. 88–101. [Google Scholar]
- Alberts, B.; Johnson, A.; Lewis, J.; Morgan, D.; Raff, M.; Roberts, K.; Walter, P. Cell Chemistry and Bioenergetics (chapter 2). In Molecular Biology of the Cell; Garland-Sciences-CRCPress: New York, NY, USA, 2017; pp. 43–108. [Google Scholar]
- Depeint, F.; Bruce, W.R.; Shangari, N.; Mehta, R.; O’Brien, P.J. Mitochondrial function and toxicity: Role of the B vitamin family on mitochondrial energy metabolism. Chem. Biol. Interact. 2006, 163, 94–112. [Google Scholar] [CrossRef]
- Dhir, S.; Tarasenko, M.; Napoli, E.; Giulivi, C. Neurological, Psychiatric, and Biochemical Aspects of Thiamine Deficiency in Children and Adults. Front. Psychiatry 2019, 10, 207. [Google Scholar] [CrossRef] [PubMed]
- Said, H.M.; Ross, A.C. Riboflavin. In Modern Nutrition in Health and Disease, 11th ed.; Ross, A.C., Caballero, B., Cousins, R.J., Eds.; Lippincott Williams & Wilkins: Baltimore, MD, USA, 2014; pp. 325–330. [Google Scholar]
- Mackey, A.; Davis, S.; Gregory, J. Vitamin B6. In Modern Nutrition in Health and Disease; Shils, M., Shike, M., Ross, A., Caballero, B., Cousins, R., Eds.; Lippincott Williams and Wilkins: Baltimore, MD, USA, 2006; pp. 194–210. [Google Scholar]
- Zempleni, J.; Wueratne, S.S.K.; Kurosihi, T. Biotin. In Present Knowledge in Nutrition; Erdman, J.W., Macdonald, I.A., Zeisel, S.H., Eds.; Wiley-Blackwell: Washington, DC, USA, 2012; pp. 359–374. [Google Scholar]
- Mock, D.M. Biotin: Physiology, dietary sources, and requirements. In Encyclopedia of Human Nutrition, 3rd ed.; Caballero, B., Ed.; Academic Press: Waltham, MA, USA, 2013; pp. 182–190. [Google Scholar]
- Green, R.; Allen, L.H.; Bjorke-Monsen, A.L.; Brito, A.; Gueant, J.L.; Miller, J.W.; Molloy, A.M.; Nexo, E.; Stabler, S.; Toh, B.H.; et al. Vitamin B12 deficiency. Nat. Rev. Dis. Primers 2017, 3, 17040. [Google Scholar] [CrossRef] [PubMed]
- Johnston, C.S.; Vitamin, C. Present Knowledge in Nutrition; Erdman, J.W., Macdonald, I.A., Zeisel, S.H., Eds.; Wiley-Blackwell: Washington, DC, USA, 2012; pp. 248–260. [Google Scholar]
- Levin, M.; Katz, A.; Padayatty, S.J.; Vitamin, C. Modern Nutrition in Health and Disease; Shils, M.E., Shike, M., Ross, A.C., Caballero, B., Cousins, R.J., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2006; pp. 507–524. [Google Scholar]
- Aggett, P.J. Iron. In Present Knowledge in Nutrition, 10th ed.; Erdman, J.W., Macdonald, I.A., Zeisel, S.H., Eds.; Wiley-Blackwell: Washington, DC, USA, 2012; pp. 506–520. [Google Scholar]
- Pilchova, I.; Klacanova, K.; Tatarkova, Z.; Kaplan, P.; Racay, P. The Involvement of Mg2+ in Regulation of Cellular andMitochondrial Functions. Oxid. Med. Cell. Longev. 2017, 2017, 6797460. [Google Scholar] [CrossRef] [PubMed]
- Volpe, S.L. Magnesium. In Present Knowledge in Nutrition, 10th ed.; Erdman, J.W., Macdonald, I.A., Zeisel, S.H., Eds.; International Life Sciences Institute, John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; pp. 459–474. [Google Scholar]
- Rude, R.K. Magnesium. In Modern Nutrition in Health and Disease, 11th ed.; Ross, C.A., Caballero, B., Cousins, R.J., Tucker, K.L., Ziegler, T.R., Eds.; Lippincott Williams & Wilkins: Baltimore, MD, USA, 2014; pp. 159–175. [Google Scholar]
- Yamanaka, R.; Tabata, S.; Shindo, Y.; Hotta, K.; Suzuki, K.; Soga, T.; Oka, K. Mitochondrial Mg(2+) homeostasis decides cellular energy metabolism and vulnerability to stress. Sci. Rep. 2016, 6, 30027. [Google Scholar] [CrossRef]
- Brown, M.J.; Beier, K. Vitamin B6 Deficiency (Pyridoxine). In StatPearls; StatPearls Publishing LLC: Treasure Island, FL, USA, 2019. [Google Scholar]
- Bailey, L.B.; Caudill, M.A. Folate. In Present Knowledge in Nutrition; Erdman, J.W., Macdonald, I.A., Zeisel, S.H., Eds.; Wiley-Blackwell: Washington, DC, USA, 2012; pp. 321–342. [Google Scholar]
- Hariz, A.; Bhattacharya, P.T. Megaloblastic Anemia. In StatPearls; StatPearls Publishing LLC: Treasure Island, FL, USA, 2019. [Google Scholar]
- Hoffbrand, A.V.; Jackson, B.F. Correction of the DNA synthesis defect in vitamin B12 deficiency by tetrahydrofolate: Evidence in favour of the methyl-folate trap hypothesis as the cause of megaloblastic anemia in vitamin B12 deficiency. Br. J. Haematol. 1993, 83, 643–647. [Google Scholar] [CrossRef]
- Butnariu, M. The oxygen paradox. J. Pharmacogenom. Pharmacoproteom. 2012, 3, 1–3. [Google Scholar] [CrossRef]
- Rivlin, R.S. Riboflavin (Vitamin B2). In Handbook of Vitamins, 4th ed.; Zempleni, J., Rucker, R.B., McCormick, D.B., Suttie, J.W., Eds.; CRC Press: Boca raton, FL, USA, 2007; pp. 233–252. [Google Scholar]
- Ballaz, S.J.; Rebec, G.V. Neurobiology of vitamin C: Expanding the focus from antioxidant to endogenous neuromodulator. Pharmacol. Res. 2019, 146, 104321. [Google Scholar] [CrossRef]
- Traber, M.G.; Vitamin, E. Present Knowledge in Nutrition, 10th ed.; Erdman, J.W., Macdonald, I.A., Zeisel, S.H., Eds.; Wiley-Blackwell: Washington, DC, USA, 2012; pp. 214–229. [Google Scholar]
- Sies, H.; Stahl, W.; Sundquist, A.R. Antioxidant functions of vitamins. Vitamins E and C, beta-carotene, and other carotenoids. Ann. N. Y. Acad. Sci. 1992, 669, 7–20. [Google Scholar] [CrossRef]
- Zheltova, A.A.; Kharitonova, M.V.; Iezhitsa, I.N.; Spasov, A.A. Magnesium deficiency and oxidative stress: An update. Biomedicine (Taipei) 2016, 6, 20. [Google Scholar] [CrossRef]
- Rayssiguier, Y.; Durlach, J.; Gueux, E.; Rock, E.; Mazur, A. Magnesium and ageing. I. Experimental data: Importance of oxidative damage. Magnes. Res. 1993, 6, 369–378. [Google Scholar]
- Morais, J.B.; Severo, J.S.; Santos, L.R.; de Sousa Melo, S.R.; de Oliveira Santos, R.; de Oliveira, A.R.; Cruz, K.J.; do Nascimento Marreiro, D. Role of Magnesium in Oxidative Stress in Individuals with Obesity. Biol. Trace. Elem. Res. 2017, 176, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Rock, E.; Astier, C.; Lab, C.; Malpuech, C.; Nowacki, W.; Gueux, E.; Mazur, A.; Rayssiguier, Y. Magnesium deficiency in rats induces a rise in plasma nitric oxide. Magnes. Res. 1995, 8, 237–242. [Google Scholar] [PubMed]
- King, J.C.; Cousins, R.J. Zinc. In Modern Nutrition in Health and Disease, 11th ed.; Shils, M.E., Shike, M., Ross, A.C., Caballero, B., Cousins, R.J., Eds.; Lippincott Williams and Wilkins: Baltimore, MD, USA, 2006; pp. 189–205. [Google Scholar]
- Holt, R.R.; Uriu-Adams, J.Y.; Keen, C.L. Zinc. In Present Knowledge in Nutrition; Erdman, J.W., Macdonald, I.A., Zeisel, S.H., Eds.; Wiley-Blackwell: Washington, DC, USA, 2012; pp. 521–539. [Google Scholar]
- Prasad, A.S.; Bao, B. Molecular Mechanisms of Zinc as a Pro-Antioxidant Mediator: Clinical Therapeutic Implications. Antioxidants 2019, 8, 164. [Google Scholar] [CrossRef] [PubMed]
- Carocci, A.; Catalano, A.; Sinicropi, M.S.; Genchi, G. Oxidative stress and neurodegeneration: The involvement of iron. Biometals 2018, 31, 715–735. [Google Scholar] [CrossRef] [PubMed]
- Grochowski, C.; Litak, J.; Kamieniak, P.; Maciejewski, R. Oxidative stress in cerebral small vessel disease. Role of reactive species. Free Radic. Res. 2018, 52, 1–13. [Google Scholar] [CrossRef]
- Udipi, S.A.; Ghugre, P.S.; Gokhale, C. Iron, Oxidative Stress and Health. In Oxidative Stress—Molecular Mechanisms and Biological Effects; Lushchak, V., Ed.; InTech: London, UK, 2012; pp. 73–108. ISBN 978-953-51-0554-1. Available online: http://www.intechopen.com/books/oxidative-stress-molecular-mechanisms-and-biological-effects/iron-oxidative-stress-and-health-authors (accessed on 9 January 2020).
- Daher, R.; Manceau, H.; Karim, Z. Iron metabolism and the role of the iron-regulating hormone hepcidin in health and disease. Presse Med. 2017, 46, e272–e278. [Google Scholar] [CrossRef]
- Bourre, J.M. Effects of nutrients (in food) on the structure and function of the nervous system: Update on dietary requirements for brain. Part 1: Micronutrients. J. Nutr. Health Aging 2006, 10, 377–385. [Google Scholar]
- Bryan, J.; Osendarp, S.; Hughes, D.; Calvaresi, E.; Baghurst, K.; van Klinken, J.W. Nutrients for cognitive development in school-aged children. Nutr. Rev. 2004, 62, 295–306. [Google Scholar] [CrossRef]
- Moore, K.; Hughes, C.F.; Ward, M.; Hoey, L.; McNulty, H. Diet, nutrition and the ageing brain: Current evidence and new directions. Proc. Nutr. Soc. 2018, 77, 152–163. [Google Scholar] [CrossRef]
- Ba, A. Metabolic and structural role of thiamine in nervous tissues. Cell Mol. Neurobiol. 2008, 28, 923–931. [Google Scholar] [CrossRef]
- Trumbo, P.R. Pantothenic acid. In Modern Nutrition in Health and Disease, 11th ed.; Ross, A.C., Caballero, B., Cousins, R.J., Eds.; Lippincott Williams & Wilkins: Baltimore, MD, USA, 2014; pp. 351–357. [Google Scholar]
- Guilland, J.C.; Lequeu, B. Mécanismes d’action moléculaire et rôles des vitamines. In Encyclopedie des Vitamines; Lavoisier, T.-D., Ed.; Editions Médicales Internationales: Paris, France, 2009; Volume 1, pp. 353–659. [Google Scholar]
- Selhub, J.; Bagley, L.C.; Miller, J.; Rosenberg, I.H. B vitamins, homocysteine, and neurocognitive function in the elderly. Am. J. Clin. Nutr. 2000, 71, 614S–620S. [Google Scholar] [CrossRef]
- Bottiglieri, T.; Laundy, M.; Crellin, R.; Toone, B.K.; Carney, M.W.; Reynolds, E.H. Homocysteine, folate, methylation, and monoamine metabolism in depression. J. Neurol. Neurosurg. Psychiatry 2000, 69, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Harrison, F.E.; May, J.M. Vitamin C function in the brain: Vital role of the ascorbate transporter SVCT2. Free Radic. Biol. Med. 2009, 46, 719–730. [Google Scholar] [CrossRef]
- Munoz, P.; Humeres, A. Iron deficiency on neuronal function. Biometals 2012, 25, 825–835. [Google Scholar] [CrossRef] [PubMed]
- Sensi, S.L.; Paoletti, P.; Bush, A.I.; Sekler, I. Zinc in the physiology and pathology of the CNS. Nat. Rev. Neurosci. 2009, 10, 780–791. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, J.A.; Parrott, J. New considerations on the neuromodulatory role of thiamine. Pharmacology 2012, 89, 111–116. [Google Scholar] [CrossRef]
- Rivera-Calimlim, L.; Hartley, D.; Osterhout, D. Effects of ethanol and pantothenic acid on brain acetylcholine synthesis. Br. J. Pharmacol. 1988, 95, 77–82. [Google Scholar] [CrossRef]
- Clayton, P.T. B6-responsive disorders: A model of vitamin dependency. J. Inherit. Metab. Dis. 2006, 29, 317–326. [Google Scholar] [CrossRef]
- Dakshinamurti, K.; Paulose, C.S.; Viswanathan, M.; Siow, Y.L.; Sharma, S.K.; Bolster, B. Neurobiology of pyridoxine. Ann. N. Y. Acad. Sci. 1990, 585, 128–144. [Google Scholar] [CrossRef]
- Kennedy, D.O. B Vitamins and the Brain: Mechanisms, Dose and Efficacy—A Review. Nutrients 2016, 8, 68. [Google Scholar] [CrossRef]
- Kirkland, J.B. Niacin. In Modern Nutrition in Health and Disease, 11th ed.; Ross, A.C., Caballero, B., Cousins, R.J., Eds.; Lippincott Williams & Wilkins: Baltimore, MD, USA, 2014; pp. 331–340. [Google Scholar]
- Hidalgo, C.; Carrasco, M.A.; Munoz, P.; Nunez, M.T. A role for reactive oxygen/nitrogen species and iron on neuronal synaptic plasticity. Antioxid. Redox Signal. 2007, 9, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, A.E.; Sarlo, G.L.; Holton, K.F. The Role of Magnesium in Neurological Disorders. Nutrients 2018, 10, 730. [Google Scholar] [CrossRef]
- Scholey, A. Nutrients for neurocognition in health and disease: Measures, methodologies and mechanisms. Proc. Nutr. Soc. 2018, 77, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, K.C.; Bourassa, M.W.; Adamolekun, B.; Bergeron, G.; Bettendorff, L.; Brown, K.H.; Cox, L.; Fattal-Valevski, A.; Fischer, P.R.; Frank, E.L.; et al. Thiamine deficiency disorders: Diagnosis, prevalence, and a roadmap for global control programs. Ann. N. Y. Acad. Sci. 2018, 1430, 3–43. [Google Scholar] [CrossRef]
- Shi, Z.; Zhen, S.; Wittert, G.A.; Yuan, B.; Zuo, H.; Taylor, A.W. Inadequate riboflavin intake and anemia risk in a Chinese population: Five-year follow up of the Jiangsu Nutrition Study. PLoS ONE 2014, 9, e88862. [Google Scholar] [CrossRef]
- Aljaadi, A.M.; How, R.E.; Loh, S.P.; Hunt, S.E.; Karakochuk, C.D.; Barr, S.I.; McAnena, L.; Ward, M.; McNulty, H.; Khor, G.L.; et al. Suboptimal Biochemical Riboflavin Status Is Associated with Lower Hemoglobin and Higher Rates of Anemia in a Sample of Canadian and Malaysian Women of Reproductive Age. J. Nutr. 2019, 149, 1952–1959. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, J.; Schwartz, R.A.; Hegyi, V. Pellagra: Dermatitis, dementia, and diarrhea. Int. J. Dermatol. 2004, 43, 1–5. [Google Scholar] [CrossRef]
- Hodges, R.E.; Ohlson, M.A.; Bean, W.B. Pantothenic acid deficiency in man. J. Clin. Investig. 1958, 37, 1642–1657. [Google Scholar] [CrossRef]
- McCurdy, P.R. Is there an anemia responsive to pantothenic acid? J. Am. Geriatr. Soc. 1973, 21, 88–91. [Google Scholar] [CrossRef]
- Rosenthal, T.C.; Majeroni, B.A.; Pretorius, R.; Malik, K. Fatigue: An overview. Am. Fam. Physician 2008, 78, 1173–1179. [Google Scholar]
- Hisano, M.; Suzuki, R.; Sago, H.; Murashima, A.; Yamaguchi, K. Vitamin B6 deficiency and anemia in pregnancy. Eur. J. Clin. Nutr. 2010, 64, 221–223. [Google Scholar] [CrossRef] [PubMed]
- Reinken, L.; Kurz, R. Activity studies of an iron-vitamin B6 preparation for euteral treatment of iron deficiency anemia. Int. J. Vitam. Nutr. Res. 1975, 45, 411–418. [Google Scholar] [PubMed]
- Lanier, J.B.; Park, J.J.; Callahan, R.C. Anemia in Older Adults. Am. Fam. Physician. 2018, 98, 437–442. [Google Scholar]
- Stabler, S.P. Clinical practice. Vitamin B12 deficiency. N. Engl. J. Med. 2013, 368, 149–160. [Google Scholar] [CrossRef]
- McCall, S.J.; Clark, A.B.; Luben, R.N.; Wareham, N.J.; Khaw, K.T.; Myint, P.K. Plasma Vitamin C Levels: Risk Factors for Deficiency and Association with Self-Reported Functional Health in the European Prospective Investigation into Cancer-Norfolk. Nutrients 2019, 11, 1552. [Google Scholar] [CrossRef]
- Myint, P.K.; Welch, A.A.; Bingham, S.A.; Surtees, P.G.; Wainwright, N.W.; Luben, R.N.; Wareham, N.J.; Smith, R.D.; Harvey, I.M.; Day, N.E.; et al. Fruit and vegetable consumption and self-reported functional health in men and women in the European Prospective Investigation into Cancer-Norfolk (EPIC-Norfolk): A population-based cross-sectional study. Public Health Nutr. 2007, 10, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Yokoi, K.; Konomi, A. Iron deficiency without anemia is a potential cause of fatigue: Meta-analyses of randomised controlled trials and cross-sectional studies. Br. J. Nutr. 2017, 117, 1422–1431. [Google Scholar] [CrossRef] [PubMed]
- Haas, J.D.; Brownlie, T.T. Iron deficiency and reduced work capacity: A critical review of the research to determine a causal relationship. J. Nutr. 2001, 131, 676S–688S, discussion 688S–690S. [Google Scholar] [CrossRef]
- Johnson, S. The multifaceted and widespread pathology of magnesium deficiency. Med. Hypotheses 2001, 56, 163–170. [Google Scholar] [CrossRef]
- Zhang, Y.; Xun, P.; Wang, R.; Mao, L.; He, K. Can Magnesium Enhance Exercise Performance? Nutrients 2017, 9, 946. [Google Scholar] [CrossRef]
- Santos, D.A.; Matias, C.N.; Monteiro, C.P.; Silva, A.M.; Rocha, P.M.; Minderico, C.S.; Bettencourt Sardinha, L.; Laires, M.J. Magnesium intake is associated with strength performance in elite basketball, handball and volleyball players. Magnes. Res. 2011, 24, 215–219. [Google Scholar] [CrossRef]
- Lukaski, H.C. Magnesium, zinc, and chromium nutriture and physical activity. Am. J. Clin. Nutr. 2000, 72, 585S–593S. [Google Scholar] [CrossRef]
- Suzuki, M.; Itokawa, Y. Effects of thiamine supplementation on exercise-induced fatigue. Metab. Brain Dis. 1996, 11, 95–106. [Google Scholar] [CrossRef]
- Powers, H.J.; Hill, M.H.; Mushtaq, S.; Dainty, J.R.; Majsak-Newman, G.; Williams, E.A. Correcting a marginal riboflavin deficiency improves hematologic status in young women in the United Kingdom (RIBOFEM). Am. J. Clin. Nutr. 2011, 93, 1274–1284. [Google Scholar] [CrossRef]
- Hoffman, M.D.; Valentino, T.R.; Stuempfle, K.J.; Hassid, B.V. A Placebo-Controlled Trial of Riboflavin for Enhancement of Ultramarathon Recovery. Sports Med. Open 2017, 3, 14. [Google Scholar] [CrossRef]
- Miller, J.; Rucker, R. Pantothenic acid. In Present Knowledge in Nutrition; Erdman, J., Macdonald, I., Zeisel, S., Eds.; Wiley-Blackwell: Washington, DC, USA, 2012; pp. 375–390. [Google Scholar]
- Tabei, S.M.; Mazloom, M.; Shahriari, M.; Zareifar, S.; Azimi, A.; Hadaegh, A.; Karimi, M. Determining and surveying the role of carnitine and folic acid to decrease fatigue in beta-thalassemia minor subjects. Pediatr. Hematol. Oncol. 2013, 30, 742–747. [Google Scholar] [CrossRef]
- Huck, C.J.; Johnston, C.S.; Beezhold, B.L.; Swan, P.D. Vitamin C status and perception of effort during exercise in obese adults adhering to a calorie-reduced diet. Nutrition 2013, 29, 42–45. [Google Scholar] [CrossRef]
- Yeom, H.; Jung, G.C.; Shin, S.; Kim, S.; Choi, J.; Lee, W.; Kang, J.; Song, K. Changes In Worker Fatigue After Vitamin C Administration. J. Orthomol. Med. 2008, 23, 205–209. [Google Scholar]
- Suh, S.Y.; Bae, W.K.; Ahn, H.Y.; Choi, S.E.; Jung, G.C.; Yeom, C.H. Intravenous vitamin C administration reduces fatigue in office workers: A double-blind randomized controlled trial. Nutr. J. 2012, 11, 7. [Google Scholar] [CrossRef]
- Carr, A.C.; Vissers, M.C.; Cook, J.S. The effect of intravenous vitamin C on cancer- and chemotherapy-related fatigue and quality of life. Front. Oncol. 2014, 4, 283. [Google Scholar] [CrossRef]
- World Health Organization. Nutritional Anaemias: Tools for Effective Prevention and Control; Licence: CC BY-NC-SA 3.0 IGO; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Houston, B.L.; Hurrie, D.; Graham, J.; Perija, B.; Rimmer, E.; Rabbani, R.; Bernstein, C.N.; Turgeon, A.F.; Fergusson, D.A.; Houston, D.S.; et al. Efficacy of iron supplementation on fatigue and physical capacity in non-anemic iron-deficient adults: A systematic review of randomised controlled trials. BMJ Open 2018, 8, e019240. [Google Scholar] [CrossRef]
- Vaucher, P.; Druais, P.L.; Waldvogel, S.; Favrat, B. Effect of iron supplementation on fatigue in nonanemic menstruating women with low ferritin: A randomized controlled trial. CMAJ 2012, 184, 1247–1254. [Google Scholar] [CrossRef]
- Favrat, B.; Balck, K.; Breymann, C.; Hedenus, M.; Keller, T.; Mezzacasa, A.; Gasche, C. Evaluation of a single dose of ferric carboxymaltose in fatigued, iron-deficient women--PREFER a randomized, placebo-controlled study. PLoS ONE 2014, 9, e94217. [Google Scholar] [CrossRef]
- Burden, R.J.; Morton, K.; Richards, T.; Whyte, G.P.; Pedlar, C.R. Is iron treatment beneficial in, iron-deficient but non-anemic (IDNA) endurance athletes? A systematic review and meta-analysis. Br. J. Sports Med. 2015, 49, 1389–1397. [Google Scholar] [CrossRef]
- Pasricha, S.R.; Low, M.; Thompson, J.; Farrell, A.; De-Regil, L.M. Iron supplementation benefits physical performance in women of reproductive age: A systematic review and meta-analysis. J. Nutr. 2014, 144, 906–914. [Google Scholar] [CrossRef]
- Pompano, L.M.; Haas, J.D. Increasing Iron Status through Dietary Supplementation in Iron-Depleted, Sedentary Women Increases Endurance Performance at Both Near-Maximal and Submaximal Exercise Intensities. J. Nutr. 2019, 149, 231–239. [Google Scholar] [CrossRef]
- Park, H.; Parker, G.L.; Boardman, C.H.; Morris, M.M.; Smith, T.J. A pilot phase II trial of magnesium supplements to reduce menopausal hot flashes in breast cancer patients. Support. Care Cancer 2011, 19, 859–863. [Google Scholar] [CrossRef]
- Brilla, L.R.; Gunther, K.B. Effect of magnesium supplementation on exercise time to exhaustion. Med. Exerc. Nutr. Health 1995, 4, 230. [Google Scholar]
- Brilla, L.R.; Haley, T.F. Effect of magnesium supplementation on strength training in humans. J. Am. Coll. Nutr. 1992, 11, 326–329. [Google Scholar] [CrossRef]
- Kass, L.S.; Poeira, F. The effect of acute vs chronic magnesium supplementation on exercise and recovery on resistance exercise, blood pressure and total peripheral resistance on normotensive adults. J. Int. Soc. Sports Nutr. 2015, 12, 19. [Google Scholar] [CrossRef]
- Terblanche, S.; Noakes, T.D.; Dennis, S.C.; Marais, D.; Eckert, M. Failure of magnesium supplementation to influence marathon running performance or recovery in magnesium-replete subjects. Int. J. Sport Nutr. 1992, 2, 154–164. [Google Scholar] [CrossRef]
- Cinar, V.; Nizamlioglu, M.; Mogulkoc, R. The effect of magnesium supplementation on lactate levels of sportsmen and sedanter. Acta Physiol. Hung. 2006, 93, 137–144. [Google Scholar] [CrossRef]
- Mosnier, E.; Niemetzky, F.; Stroot, J.; Pommier de Santi, V.; Brousse, P.; Guarmit, B.; Blanchet, D.; Ville, M.; Abboud, P.; Djossou, F.; et al. A Large Outbreak of Thiamine Deficiency Among Illegal Gold Miners in French Guiana. Am. J. Trop. Med. Hyg. 2017, 96, 1248–1252. [Google Scholar] [CrossRef][Green Version]
- Isenberg-Grzeda, E.; Kutner, H.E.; Nicolson, S.E. Wernicke-Korsakoff-syndrome: Under-recognized and under-treated. Psychosomatics 2012, 53, 507–516. [Google Scholar] [CrossRef]
- Gibson, G.E.; Hirsch, J.A.; Fonzetti, P.; Jordan, B.D.; Cirio, R.T.; Elder, J. Vitamin B1 (thiamine) and dementia. Ann. N. Y. Acad. Sci. 2016, 1367, 21–30. [Google Scholar] [CrossRef]
- Oldham, M.A.; Ivkovic, A. Pellagrous encephalopathy presenting as alcohol withdrawal delirium: A case series and literature review. Addict. Sci. Clin. Pract. 2012, 7, 12. [Google Scholar] [CrossRef]
- Noda, S.; Haratake, J.; Sasaki, A.; Ishii, N.; Umezaki, H.; Horie, A. Acute encephalopathy with hepatic steatosis induced by pantothenic acid antagonist, calcium hopantenate, in dogs. Liver 1991, 11, 134–142. [Google Scholar] [CrossRef]
- Gregory, A.; Polster, B.J.; Hayflick, S.J. Clinical and genetic delineation of neurodegeneration with brain iron accumulation. J. Med. Genet. 2009, 46, 73–80. [Google Scholar] [CrossRef]
- Gul-Mert, G.; Incecik, F.; Herguner, M.O.; Ceylaner, S.; Altunbasak, S. Pyridoxine-dependent epilepsy in two Turkish patients in Turkey and review of the literature. Turk. J. Pediatr. 2015, 57, 394–397. [Google Scholar]
- O’Leary, F.; Samman, S. Vitamin B(12) in Health and Disease. Nutrients 2010, 2, 299–316. [Google Scholar] [CrossRef]
- Hunt, A.; Harrington, D.; Robinson, S. Vitamin B12 deficiency. BMJ 2014, 349, g5226. [Google Scholar] [CrossRef] [PubMed]
- Salminen, L.E.; Paul, R.H. Oxidative stress and genetic markers of suboptimal antioxidant defense in the aging brain: A theoretical review. Rev. Neurosci. 2014, 25, 805–819. [Google Scholar] [CrossRef]
- Grantham-McGregor, S.; Ani, C. A review of studies on the effect of iron deficiency on cognitive development in children. J. Nutr. 2001, 131, 649S–666S. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, G.; Jang, W.; Kim, S.Y.; Chang, N. Association between intake of B vitamins and cognitive function in elderly Koreans with cognitive impairment. Nutr. J. 2014, 13, 118. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Evans, D.A.; Bienias, J.L.; Scherr, P.A.; Tangney, C.C.; Hebert, L.E.; Bennett, D.A.; Wilson, R.S.; Aggarwal, N. Dietary niacin and the risk of incident Alzheimer’s disease and of cognitive decline. J. Neurol. Neurosurg. Psychiatry 2004, 75, 1093–1099. [Google Scholar] [CrossRef]
- Qin, B.; Xun, P.; Jacobs, D.R., Jr.; Zhu, N.; Daviglus, M.L.; Reis, J.P.; Steffen, L.M.; Van Horn, L.; Sidney, S.; He, K. Intake of niacin, folate, vitamin B-6, and vitamin B-12 through young adulthood and cognitive function in midlife: The Coronary Artery Risk Development in Young Adults (CARDIA) study. Am. J. Clin. Nutr. 2017, 106, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kang, S.W. Relationships between Dietary Intake and Cognitive Function in Healthy Korean Children and Adolescents. J. Lifestyle Med. 2017, 7, 10–17. [Google Scholar] [CrossRef]
- Davison, K.M.; Kaplan, B.J. Nutrient intakes are correlated with overall psychiatric functioning in adults with mood disorders. Can. J. Psychiatry 2012, 57, 85–92. [Google Scholar] [CrossRef]
- Rubio-López, N.; Morales-Suárez-Varela, M.; Pico, Y.; Livianos-Aldana, L.; Llopis-González, A. Nutrient Intake and Depression Symptoms in Spanish Children: The ANIVA Study. Int. J. Environ. Res. Public Health 2016, 13, 352. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Tsujiguchi, H.; Kambayashi, Y.; Hara, A.; Miyagi, S.; Yamada, Y.; Nakamura, H.; Shimizu, Y.; Hori, D.; Suzuki, F.; et al. Relationship between Vitamin Intake and Depressive Symptoms in Elderly Japanese Individuals: Differences with Gender and Body Mass Index. Nutrients 2017, 9, 1319. [Google Scholar] [CrossRef]
- Miyaki, K.; Song, Y.; Htun, N.C.; Tsutsumi, A.; Hashimoto, H.; Kawakami, N.; Takahashi, M.; Shimazu, A.; Inoue, A.; Kurioka, S.; et al. Folate intake and depressive symptoms in Japanese workers considering SES and job stress factors: J-HOPE study. BMC Psychiatry 2012, 12, 33. [Google Scholar] [CrossRef] [PubMed]
- Yary, T. The association between dietary intake of folate and physical activity with psychological dimensions of depressive symptoms among students from Iran. Biomed. Res. Int. 2013, 2013, 582693. [Google Scholar] [CrossRef] [PubMed]
- Doets, E.L.; Ueland, P.M.; Tell, G.S.; Vollset, S.E.; Nygard, O.K.; Van’t Veer, P.; de Groot, L.C.; Nurk, E.; Refsum, H.; Smith, A.D.; et al. Interactions between plasma concentrations of folate and markers of vitamin B(12) status with cognitive performance in elderly people not exposed to folic acid fortification: The Hordaland Health Study. Br. J. Nutr. 2014, 111, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Bender, A.; Hagan, K.E.; Kingston, N. The association of folate and depression: A meta-analysis. J. Psychiatr. Res. 2017, 95, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Petridou, E.T.; Kousoulis, A.A.; Michelakos, T.; Papathoma, P.; Dessypris, N.; Papadopoulos, F.C.; Stefanadis, C. Folate and B12 serum levels in association with depression in the aged: A systematic review and meta-analysis. Aging Ment. Health 2016, 20, 965–973. [Google Scholar] [CrossRef]
- Agnew-Blais, J.C.; Wassertheil-Smoller, S.; Kang, J.H.; Hogan, P.E.; Coker, L.H.; Snetselaar, L.G.; Smoller, J.W. Folate, vitamin B-6, and vitamin B-12 intake and mild cognitive impairment and probable dementia in the Women’s Health Initiative Memory Study. J. Acad. Nutr. Diet. 2015, 115, 231–241. [Google Scholar] [CrossRef]
- Venkatramanan, S.; Armata, I.E.; Strupp, B.J.; Finkelstein, J.L. Vitamin B-12 and Cognition in Children. Adv. Nutr. 2016, 7, 879–888. [Google Scholar] [CrossRef]
- Doets, E.L.; van Wijngaarden, J.P.; Szczecinska, A.; Dullemeijer, C.; Souverein, O.W.; Dhonukshe-Rutten, R.A.; Cavelaars, A.E.; van ‘t Veer, P.; Brzozowska, A.; de Groot, L.C. Vitamin B12 intake and status and cognitive function in elderly people. Epidemiol. Rev. 2013, 35, 2–21. [Google Scholar] [CrossRef]
- O’Leary, F.; Allman-Farinelli, M.; Samman, S. Vitamin B(1)(2) status, cognitive decline and dementia: A systematic review of prospective cohort studies. Br. J. Nutr. 2012, 108, 1948–1961. [Google Scholar] [CrossRef]
- Kvestad, I.; Hysing, M.; Shrestha, M.; Ulak, M.; Thorne-Lyman, A.L.; Henjum, S.; Ueland, P.M.; Midttun, O.; Fawzi, W.; Chandyo, R.K.; et al. Vitamin B-12 status in infancy is positively associated with development and cognitive functioning 5 y later in Nepalese children. Am. J. Clin. Nutr. 2017, 105, 1122–1131. [Google Scholar] [CrossRef]
- Louwman, M.W.; van Dusseldorp, M.; van de Vijver, F.J.; Thomas, C.M.; Schneede, J.; Ueland, P.M.; Refsum, H.; van Staveren, W.A. Signs of impaired cognitive function in adolescents with marginal cobalamin status. Am. J. Clin. Nutr. 2000, 72, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Hooshmand, B.; Solomon, A.; Kareholt, I.; Leiviska, J.; Rusanen, M.; Ahtiluoto, S.; Winblad, B.; Laatikainen, T.; Soininen, H.; Kivipelto, M. Homocysteine and holotranscobalamin and the risk of Alzheimer disease: A longitudinal study. Neurology 2010, 75, 1408–1414. [Google Scholar] [CrossRef] [PubMed]
- Tiemeier, H.; van Tuijl, H.R.; Hofman, A.; Meijer, J.; Kiliaan, A.J.; Breteler, M.M. Vitamin B12, folate, and homocysteine in depression: The Rotterdam Study. Am. J. Psychiatry 2002, 159, 2099–2101. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Cui, N.; Liu, J. Neurocognitive Function Is Associated With Serum Iron Status in Early Adolescents. Biol. Res. Nurs. 2017, 19, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Moretti, R.; Caruso, P. The Controversial Role of Homocysteine in Neurology: From Labs to Clinical Practice. Int. J. Mol. Sci. 2019, 20, 231. [Google Scholar] [CrossRef]
- Gariballa, S. Poor vitamin C status is associated with increased depression symptoms following acute illness in older people. Int. J. Vitam. Nutr. Res. 2014, 84, 12–17. [Google Scholar] [CrossRef]
- Pullar, J.M.; Carr, A.C.; Bozonet, S.M.; Vissers, M.C.M. High Vitamin C Status Is Associated with Elevated Mood in Male Tertiary Students. Antioxidants 2018, 7, 91. [Google Scholar] [CrossRef]
- Payne, M.E.; Steck, S.E.; George, R.R.; Steffens, D.C. Fruit, vegetable, and antioxidant intakes are lower in older adults with depression. J. Acad. Nutr. Diet. 2012, 112, 2022–2027. [Google Scholar] [CrossRef]
- Radlowski, E.C.; Johnson, R.W. Perinatal iron deficiency and neurocognitive development. Front. Hum. Neurosci. 2013, 7, 585. [Google Scholar] [CrossRef]
- Khedr, E.; Hamed, S.A.; Elbeih, E.; El-Shereef, H.; Ahmad, Y.; Ahmed, S. Iron states and cognitive abilities in young adults: Neuropsychological and neurophysiological assessment. Eur. Arch. Psychiatry Clin. Neurosci. 2008, 258, 489–496. [Google Scholar] [CrossRef]
- Murray-Kolb, L.E.; Beard, J.L. Iron treatment normalizes cognitive functioning in young women. Am. J. Clin. Nutr. 2007, 85, 778–787. [Google Scholar] [CrossRef] [PubMed]
- Abeyawardhane, D.L.; Lucas, H.R. Iron Redox Chemistry and Implications in the Parkinson’s Disease Brain. Oxid. Med. Cell. Longev. 2019, 2019, 4609702. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xu, S.F.; Fan, Y.G.; Li, L.B.; Guo, C. Iron Pathophysiology in Alzheimer’s Diseases. Adv. Exp. Med. Biol. 2019, 1173, 67–104. [Google Scholar] [CrossRef] [PubMed]
- Kieboom, B.C.T.; Licher, S.; Wolters, F.J.; Ikram, M.K.; Hoorn, E.J.; Zietse, R.; Stricker, B.H.; Ikram, M.A. Serum magnesium is associated with the risk of dementia. Neurology 2017, 89, 1716–1722. [Google Scholar] [CrossRef] [PubMed]
- Derom, M.L.; Sayon-Orea, C.; Martinez-Ortega, J.M.; Martinez-Gonzalez, M.A. Magnesium and depression: A systematic review. Nutr. Neurosci. 2013, 16, 191–206. [Google Scholar] [CrossRef]
- Li, B.; Lv, J.; Wang, W.; Zhang, D. Dietary magnesium and calcium intake and risk of depression in the general population: A meta-analysis. Aust. N. Z J. Psychiatry 2017, 51, 219–229. [Google Scholar] [CrossRef]
- Cheungpasitporn, W.; Thongprayoon, C.; Mao, M.A.; Srivali, N.; Ungprasert, P.; Varothai, N.; Sanguankeo, A.; Kittanamongkolchai, W.; Erickson, S.B. Hypomagnesaemia linked to depression: A systematic review and meta-analysis. Intern. Med. J. 2015, 45, 436–440. [Google Scholar] [CrossRef]
- Tarleton, E.K.; Kennedy, A.G.; Rose, G.L.; Crocker, A.; Littenberg, B. The Association between Serum Magnesium Levels and Depression in an Adult Primary Care Population. Nutrients 2019, 11, 1475. [Google Scholar] [CrossRef]
- Veronese, N.; Zurlo, A.; Solmi, M.; Luchini, C.; Trevisan, C.; Bano, G.; Manzato, E.; Sergi, G.; Rylander, R. Magnesium Status in Alzheimer’s Disease: A Systematic Review. Am. J. Alzheimers Dis. Other Demen. 2016, 31, 208–213. [Google Scholar] [CrossRef]
- Grochowski, C.; Blicharska, E.; Baj, J.; Mierzwinska, A.; Brzozowska, K.; Forma, A.; Maciejewski, R. Serum iron, Magnesium, Copper, and Manganese Levels in Alcoholism: A Systematic Review. Molecules 2019, 24, 1361. [Google Scholar] [CrossRef]
- Nechifor, M. Magnesium in drug abuse and addiction. In Magnesium in the Central Nervous System; Vink, R., Nechifor, M., Eds.; University of Adelaide Press: Adelaide, Australia, 2011; pp. 331–342. [Google Scholar]
- Swardfager, W.; Herrmann, N.; McIntyre, R.S.; Mazereeuw, G.; Goldberger, K.; Cha, D.S.; Schwartz, Y.; Lanctot, K.L. Potential roles of zinc in the pathophysiology and treatment of major depressive disorder. Neurosci. Biobehav. Rev. 2013, 37, 911–929. [Google Scholar] [CrossRef] [PubMed]
- Vashum, K.P.; McEvoy, M.; Milton, A.H.; McElduff, P.; Hure, A.; Byles, J.; Attia, J. Dietary zinc is associated with a lower incidence of depression: Findings from two Australian cohorts. J. Affect. Disord. 2014, 166, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Yary, T.; Aazami, S. Dietary intake of zinc was inversely associated with depression. Biol. Trace Elem. Res. 2012, 145, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, B.; Song, X.; Zhang, D. Dietary zinc and iron intake and risk of depression: A meta-analysis. Psychiatry Res. 2017, 251, 41–47. [Google Scholar] [CrossRef]
- Skalny, A.V.; Skalnaya, M.G.; Grabeklis, A.R.; Skalnaya, A.A.; Tinkov, A.A. Zinc deficiency as a mediator of toxic effects of alcohol abuse. Eur. J. Nutr. 2018, 57, 2313–2322. [Google Scholar] [CrossRef]
- Mezzaroba, L.; Alfieri, D.F.; Colado Simao, A.N.; Vissoci Reiche, E.M. The role of zinc, copper, manganese and iron in neurodegenerative diseases. Neurotoxicology 2019, 74, 230–241. [Google Scholar] [CrossRef]
- Chou, W.P.; Chang, Y.H.; Lin, H.C.; Chang, Y.H.; Chen, Y.Y.; Ko, C.H. Thiamine for preventing dementia development among patients with alcohol use disorder: A nationwide population-based cohort study. Clin. Nutr. 2018. [Google Scholar] [CrossRef]
- Pan, X.; Chen, Z.; Fei, G.; Pan, S.; Bao, W.; Ren, S.; Guan, Y.; Zhong, C. Long-Term Cognitive Improvement After Benfotiamine Administration in Patients with Alzheimer’s Disease. Neurosci. Bull. 2016, 32, 591–596. [Google Scholar] [CrossRef]
- Benton, D.; Griffiths, R.; Haller, J. Thiamine supplementation mood and cognitive functioning. Psychopharmacology (Berl.) 1997, 129, 66–71. [Google Scholar] [CrossRef]
- Durga, J.; van Boxtel, M.P.; Schouten, E.G.; Kok, F.J.; Jolles, J.; Katan, M.B.; Verhoef, P. Effect of 3-year folic acid supplementation on cognitive function in older adults in the FACIT trial: A randomised, double blind, controlled trial. Lancet 2007, 369, 208–216. [Google Scholar] [CrossRef]
- Fioravanti, M.; Ferrario, E.; Massaia, M.; Cappa, G.; Rivolta, G.; Grossi, E.; Buckley, A.E. Low folate levels in the cognitive decline of elderly patients and the efficacy of folate as a treatment for improving memory deficits. Arch. Gerontol. Geriatr. 1998, 26, 1–13. [Google Scholar] [CrossRef]
- Pathansali, R.; Mangoni, A.A.; Creagh-Brown, B.; Lan, Z.C.; Ngow, G.L.; Yuan, X.F.; Ouldred, E.L.; Sherwood, R.A.; Swift, C.G.; Jackson, S.H. Effects of folic acid supplementation on psychomotor performance and hemorheology in healthy elderly subjects. Arch. Gerontol. Geriatr. 2006, 43, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.G.; Batterham, P.J.; Mackinnon, A.J.; Jorm, A.F.; Hickie, I.; Fenech, M.; Kljakovic, M.; Crisp, D.; Christensen, H. Oral folic acid and vitamin B-12 supplementation to prevent cognitive decline in community-dwelling older adults with depressive symptoms--the Beyond Ageing Project: A randomized controlled trial. Am. J. Clin. Nutr. 2012, 95, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Li, Q.; Zhou, X.; Zhao, J.; Song, A.; Li, W.; Liu, H.; Xu, W.; Huang, G. Effects of folic acid supplementation on cognitive function and Abeta-related biomarkers in mild cognitive impairment: A randomized controlled trial. Eur. J. Nutr. 2019, 58, 345–356. [Google Scholar] [CrossRef]
- Gautam, M.; Agrawal, M.; Gautam, M.; Sharma, P.; Gautam, A.S.; Gautam, S. Role of antioxidants in generalized anxiety disorder and depression. Indian J. Psychiatry 2012, 54, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Lux-Battistelli, C.; Battistelli, D. Latent scurvy with tiredness and leg pain in alcoholics: An underestimated disease three case reports. Medicine (Baltim.) 2017, 96, e8861. [Google Scholar] [CrossRef]
- Low, M.; Farrell, A.; Biggs, B.A.; Pasricha, S.R. Effects of daily iron supplementation in primary-school-aged children: Systematic review and meta-analysis of randomized controlled trials. CMAJ 2013, 185, E791–802. [Google Scholar] [CrossRef]
- Scott, S.P.; Murray-Kolb, L.E.; Wenger, M.J.; Udipi, S.A.; Ghugre, P.S.; Boy, E.; Haas, J.D. Cognitive Performance in Indian School-Going Adolescents Is Positively Affected by Consumption of Iron-Biofortified Pearl Millet: A 6-Month Randomized Controlled Efficacy Trial. J. Nutr. 2018, 148, 1462–1471. [Google Scholar] [CrossRef]
- Ward, R.J.; Zucca, F.A.; Duyn, J.H.; Crichton, R.R.; Zecca, L. The role of iron in brain ageing and neurodegenerative disorders. Lancet Neurol. 2014, 13, 1045–1060. [Google Scholar] [CrossRef]
- Lomagno, K.A.; Hu, F.; Riddell, L.J.; Booth, A.O.; Szymlek-Gay, E.A.; Nowson, C.A.; Byrne, L.K. Increasing iron and zinc in pre-menopausal women and its effects on mood and cognition: A systematic review. Nutrients 2014, 6, 5117–5141. [Google Scholar] [CrossRef]
- Boyle, N.B.; Lawton, C.; Dye, L. The Effects of Magnesium Supplementation on Subjective Anxiety and Stress-A Systematic Review. Nutrients 2017, 9, 429. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, M. Rôle du Magne-B6 dans les Manifestations Anxieuses en Pratique Medicale Courante. Psychiatr. Pract. Med. 1987, 39, 18–22. [Google Scholar]
- Pouteau, E.; Kabir-Ahmadi, M.; Noah, L.; Mazur, A.; Dye, L.; Hellhammer, J.; Pickering, G.; Dubray, C. Superiority of magnesium and vitamin B6 over magnesium alone on severe stress in healthy adults with low magnesemia: A randomized, single-blind clinical trial. PLoS ONE 2018, 13, e0208454. [Google Scholar] [CrossRef] [PubMed]
- Rajizadeh, A.; Mozaffari-Khosravi, H.; Yassini-Ardakani, M.; Dehghani, A. Effect of magnesium supplementation on depression status in depressed patients with magnesium deficiency: A randomized, double-blind, placebo-controlled trial. Nutrition 2017, 35, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Warthon-Medina, M.; Moran, V.H.; Stammers, A.L.; Dillon, S.; Qualter, P.; Nissensohn, M.; Serra-Majem, L.; Lowe, N.M. Zinc intake, status and indices of cognitive function in adults and children: A systematic review and meta-analysis. Eur. J. Clin. Nutr. 2015, 69, 649–661. [Google Scholar] [CrossRef]
- Maylor, E.A.; Simpson, E.E.; Secker, D.L.; Meunier, N.; Andriollo-Sanchez, M.; Polito, A.; Stewart-Knox, B.; McConville, C.; O’Connor, J.M.; Coudray, C. Effects of zinc supplementation on cognitive function in healthy middle-aged and older adults: The ZENITH study. Br. J. Nutr. 2006, 96, 752–760. [Google Scholar]
- Lam, L.F.; Lawlis, T.R. Feeding the brain—The effects of micronutrient interventions on cognitive performance among school-aged children: A systematic review of randomized controlled trials. Clin. Nutr. 2017, 36, 1007–1014. [Google Scholar] [CrossRef]
- Eilander, A.; Gera, T.; Sachdev, H.S.; Transler, C.; van der Knaap, H.C.; Kok, F.J.; Osendarp, S.J. Multiple micronutrient supplementation for improving cognitive performance in children: Systematic review of randomized controlled trials. Am. J. Clin. Nutr. 2010, 91, 115–130. [Google Scholar] [CrossRef]
- Kennedy, D.O.; Haskell, C.F. Vitamins and cognition: What is the evidence? Drugs 2011, 71, 1957–1971. [Google Scholar] [CrossRef]
- Long, S.J.; Benton, D. Effects of vitamin and mineral supplementation on stress, mild psychiatric symptoms, and mood in nonclinical samples: A meta-analysis. Psychosom. Med. 2013, 75, 144–153. [Google Scholar] [CrossRef]
- Young, L.M.; Pipingas, A.; White, D.J.; Gauci, S.; Scholey, A. A Systematic Review and Meta-Analysis of B Vitamin Supplementation on Depressive Symptoms, Anxiety, and Stress: Effects on Healthy and ‘At-Risk’ Individuals. Nutrients 2019, 11, 2232. [Google Scholar] [CrossRef] [PubMed]
- White, D.J.; Cox, K.H.; Peters, R.; Pipingas, A.; Scholey, A.B. Effects of Four-Week Supplementation with a Multi-Vitamin/Mineral Preparation on Mood and Blood Biomarkers in Young Adults: A Randomised, Double-Blind, Placebo-Controlled Trial. Nutrients 2015, 7, 9005–9017. [Google Scholar] [CrossRef] [PubMed]
- White, D.J.; Cox, K.H.; Hughes, M.E.; Pipingas, A.; Peters, R.; Scholey, A.B. Functional Brain Activity Changes after 4 Weeks Supplementation with a Multi-Vitamin/Mineral Combination: A Randomized, Double-Blind, Placebo-Controlled Trial Exploring Functional Magnetic Resonance Imaging and Steady-State Visual Evoked Potentials during Working Memory. Front. Aging Neurosci. 2016, 8, 288. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Cook, N.; Manson, J.; Buring, J.E.; Albert, C.M.; Grodstein, F. A trial of B vitamins and cognitive function among women at high risk of cardiovascular disease. Am. J. Clin. Nutr. 2008, 88, 1602–1610. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Tolerable Upper Intake Levels for Vitamins and Minerals. 2006. Available online: http://www.efsa.europa.eu/sites/default/files/efsa_rep/blobserver_assets/ndatolerableuil.pdf (accessed on 9 January 2020).


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tardy, A.-L.; Pouteau, E.; Marquez, D.; Yilmaz, C.; Scholey, A. Vitamins and Minerals for Energy, Fatigue and Cognition: A Narrative Review of the Biochemical and Clinical Evidence. Nutrients 2020, 12, 228. https://doi.org/10.3390/nu12010228
Tardy A-L, Pouteau E, Marquez D, Yilmaz C, Scholey A. Vitamins and Minerals for Energy, Fatigue and Cognition: A Narrative Review of the Biochemical and Clinical Evidence. Nutrients. 2020; 12(1):228. https://doi.org/10.3390/nu12010228
Chicago/Turabian StyleTardy, Anne-Laure, Etienne Pouteau, Daniel Marquez, Cansu Yilmaz, and Andrew Scholey. 2020. "Vitamins and Minerals for Energy, Fatigue and Cognition: A Narrative Review of the Biochemical and Clinical Evidence" Nutrients 12, no. 1: 228. https://doi.org/10.3390/nu12010228
APA StyleTardy, A.-L., Pouteau, E., Marquez, D., Yilmaz, C., & Scholey, A. (2020). Vitamins and Minerals for Energy, Fatigue and Cognition: A Narrative Review of the Biochemical and Clinical Evidence. Nutrients, 12(1), 228. https://doi.org/10.3390/nu12010228