Human Breast Milk Promotes the Secretion of Potentially Beneficial Metabolites by Probiotic Lactobacillus reuteri DSM 17938



Abstract
1. Introduction
2. Materials and Methods
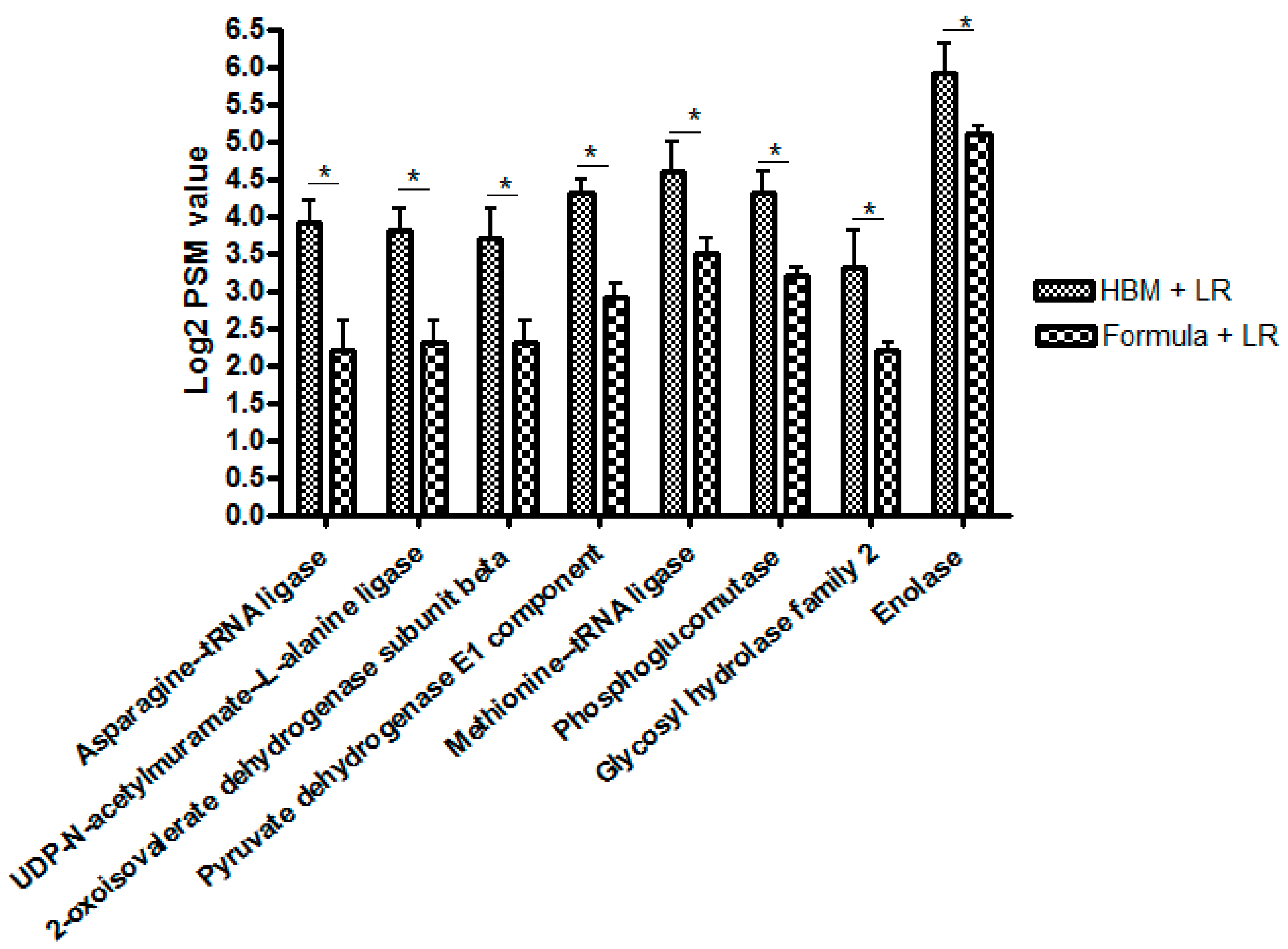
Proteomic Studies
3. Results
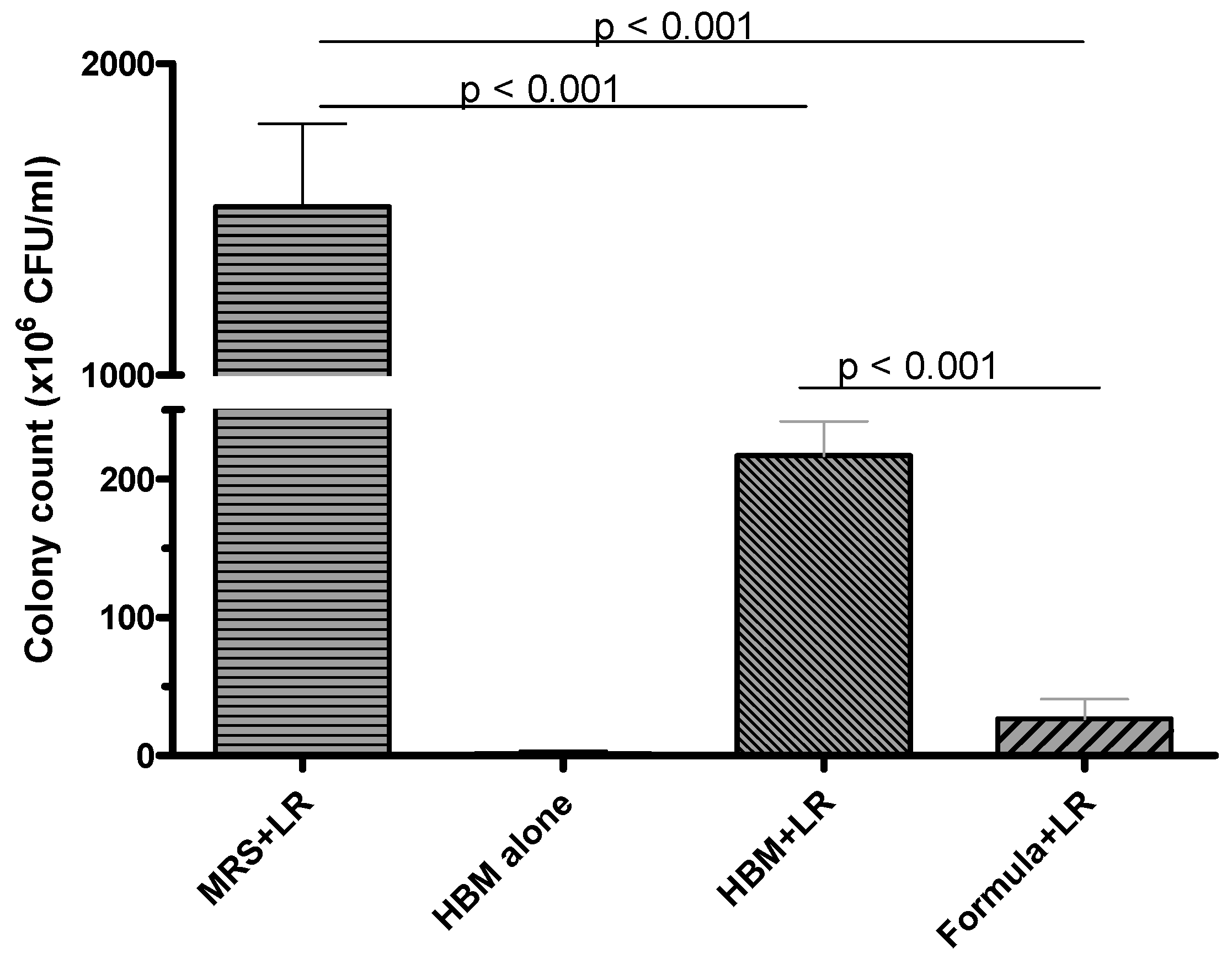
3.1. LR 17938 Growth was Higher in HBM as Compared to Its Growth in Formula
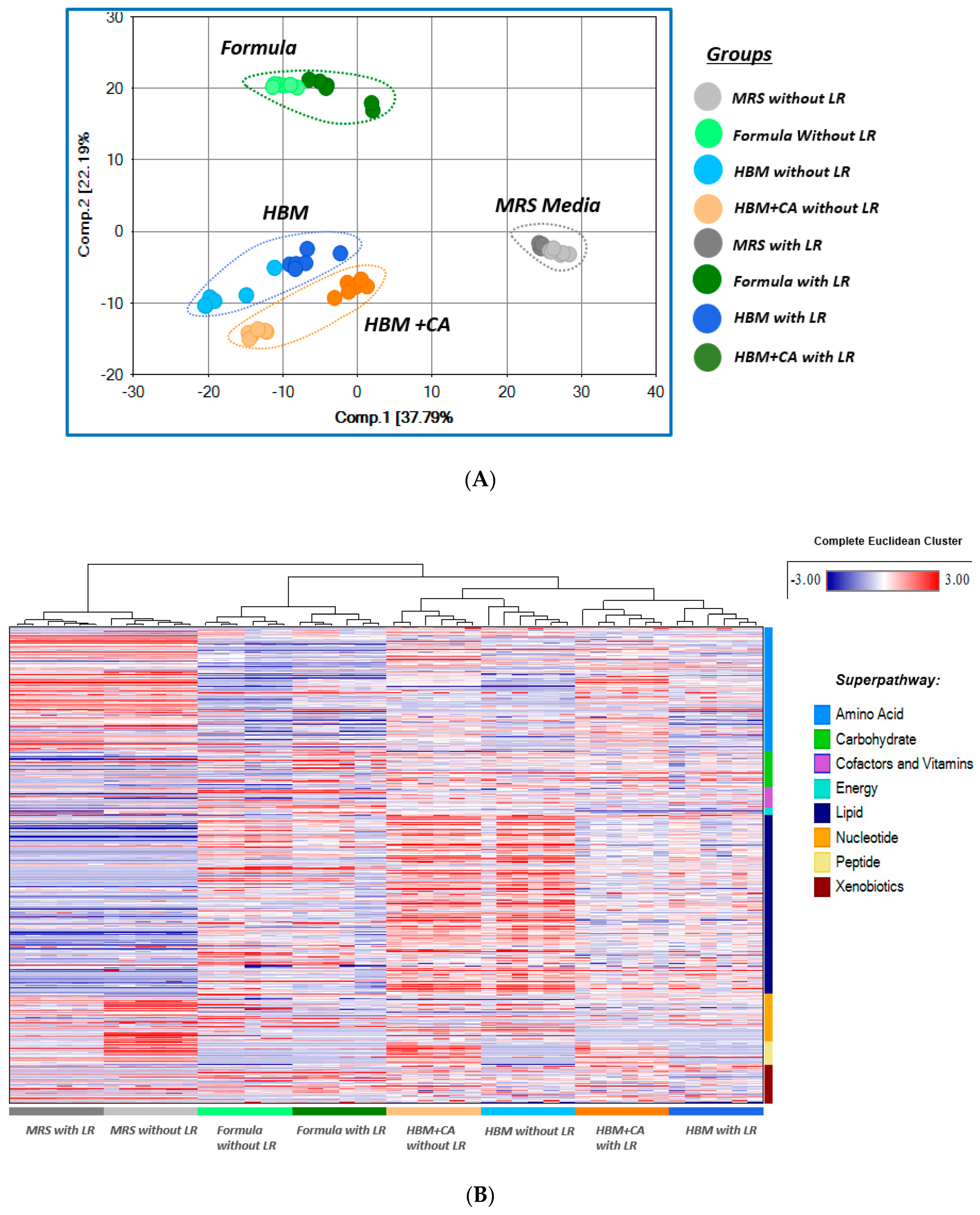
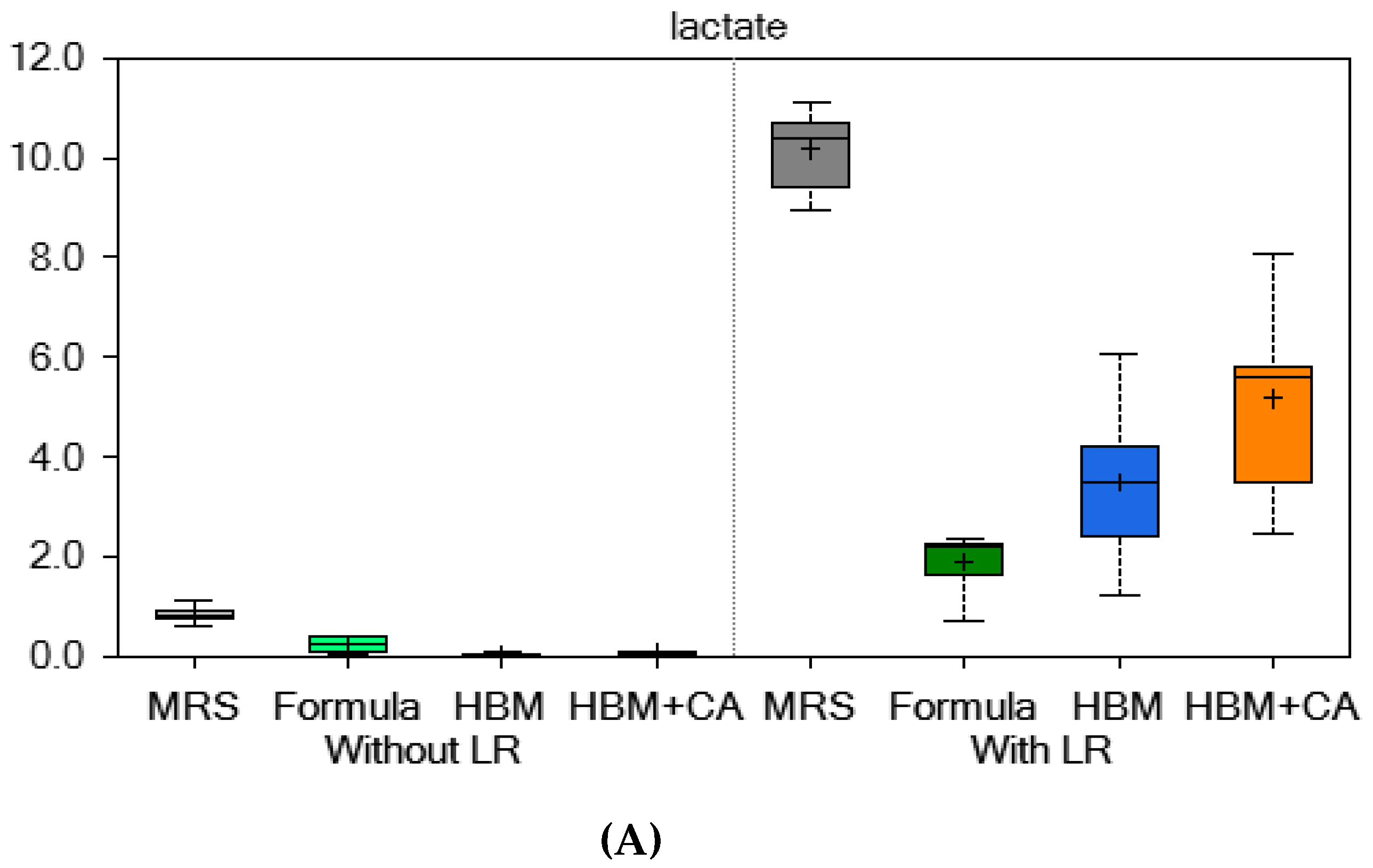
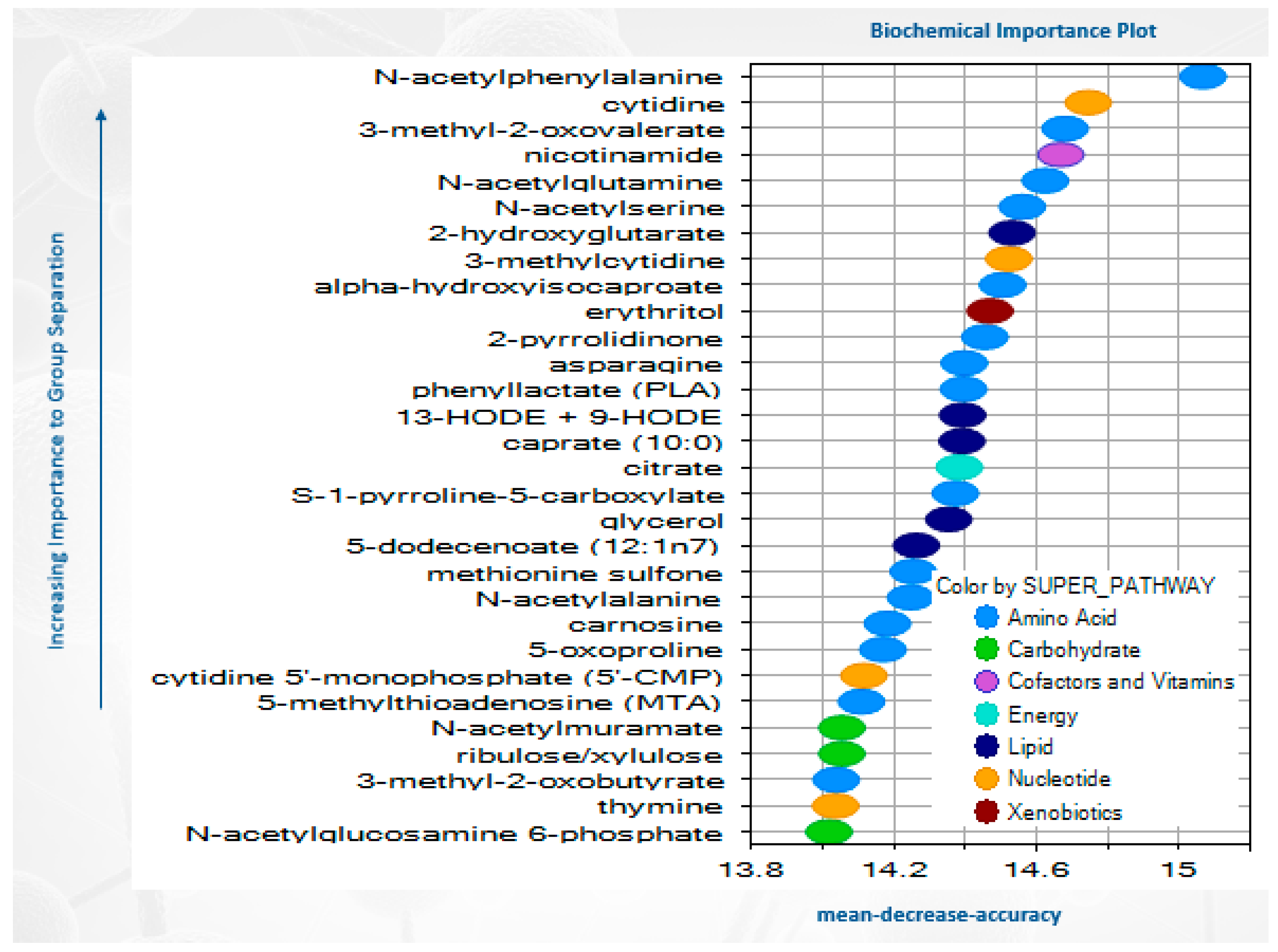
3.2. Metabolomics Data
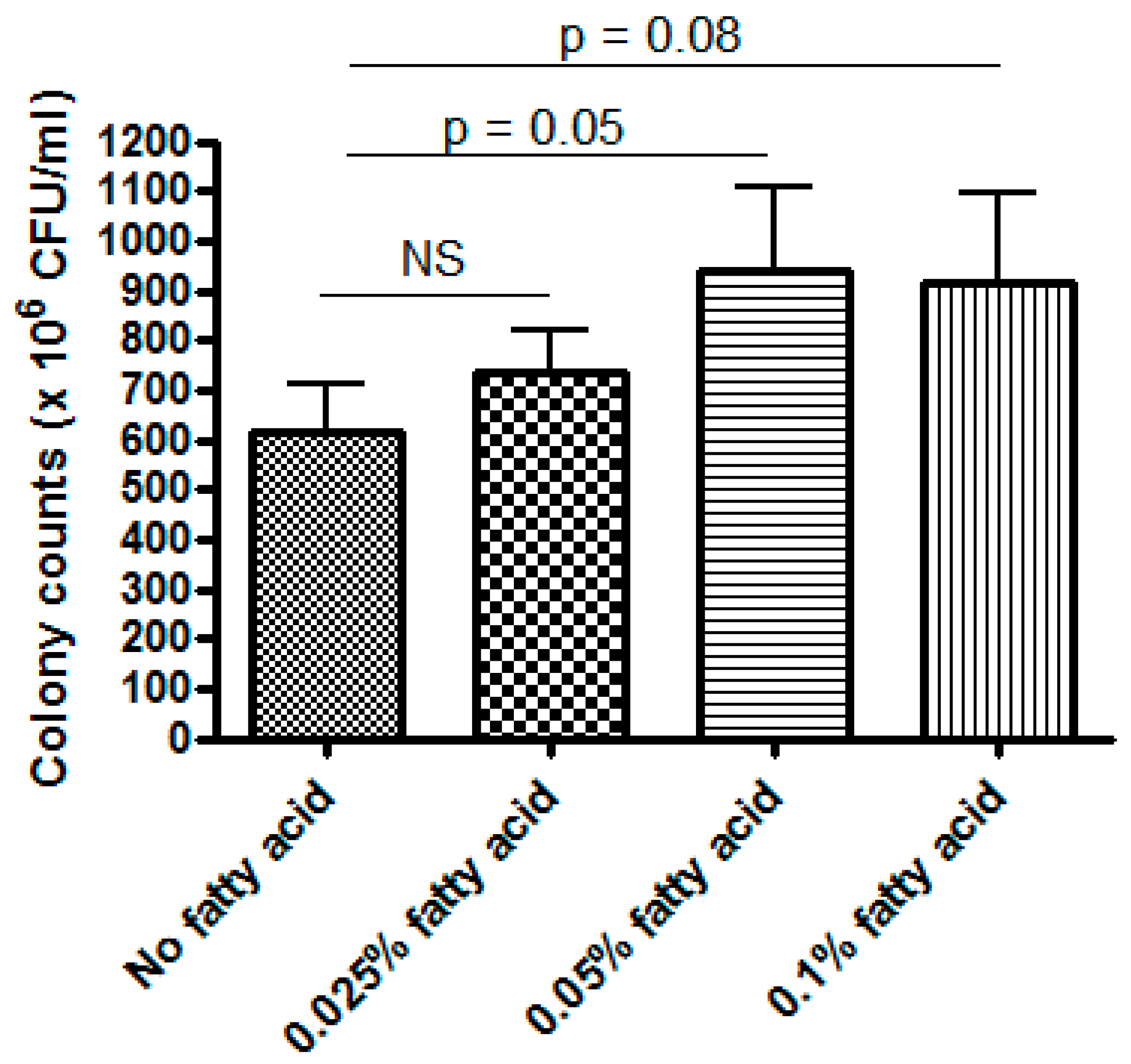
3.3. LR 17938 Growth was Promoted by 0.05% Fatty Acids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alfaleh, K.; Bassler, D. Probiotics for prevention of necrotizing enterocolitis in preterm infants. Cochrane Database Syst. Rev. 2008, 16, CD005496. [Google Scholar]
- Ford, A.C.; Quigley, E.M.; Lacy, B.E.; Lembo, A.J.; Saito, Y.A.; Schiller, L.R.; Soffer, E.E.; Spiegel, B.M.; Moayyedi, P. Efficacy of prebiotics, probiotics, and synbiotics in irritable bowel syndrome and chronic idiopathic constipation: Systematic review and meta-analysis. Am. J. Gastroenterol. 2014, 109, 1547–1561. [Google Scholar] [CrossRef] [PubMed]
- Ganji-Arjenaki, M.; Rafieian-Kopaei, M. Probiotics are a good choice in remission of inflammatory bowel diseases: A meta analysis and systematic review. J. Cell. Physiol. 2018, 233, 2091–2103. [Google Scholar] [CrossRef] [PubMed]
- Pattani, R.; Palda, V.A.; Hwang, S.W.; Shah, P.S. Probiotics for the prevention of antibiotic-associated diarrhea and Clostridium difficile infection among hospitalized patients: Systematic review and meta-analysis. Open. Med. 2013, 7, e56–e67. [Google Scholar] [PubMed]
- Salari, P.; Nikfar, S.; Abdollahi, M. A meta-analysis and systematic review on the effect of probiotics in acute diarrhea. Inflamm. Allergy Drug Targets 2012, 11, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Sung, V.; D’Amico, F.; Cabana, M.D.; Chau, K.; Koren, G.; Savino, F.; Szajewska, H.; Deshpande, G.; Dupont, C.; Indrio, F.; et al. Lactobacillus reuteri to Treat Infant Colic: A Meta-analysis. Pediatrics 2018, 141, e20171811. [Google Scholar] [CrossRef] [PubMed]
- Rosander, A.; Connolly, E.; Roos, S. Removal of antibiotic resistance gene-carrying plasmids from Lactobacillus reuteri ATCC 55730 and characterization of the resulting daughter strain, L. reuteri DSM 17938. Appl. Environ. Microbiol. 2008, 74, 6032–6040. [Google Scholar] [CrossRef]
- Liu, Y.; Fatheree, N.Y.; Mangalat, N.; Rhoads, J.M. Lactobacillus reuteri strains reduce incidence and severity of experimental necrotizing enterocolitis via modulation of TLR4 and NF-kappaB signaling in the intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G608–G617. [Google Scholar] [CrossRef]
- Liu, Y.; Fatheree, N.Y.; Dingle, B.M.; Tran, D.Q.; Rhoads, J.M. Lactobacillus reuteri DSM 17938 changes the frequency of Foxp3+ regulatory T cells in the intestine and mesenteric lymph node in experimental necrotizing enterocolitis. PLoS ONE 2013, 8, e56547. [Google Scholar] [CrossRef]
- Liu, Y.; Tran, D.Q.; Fatheree, N.Y.; Rhoads, J.M. Lactobacillus reuteri DSM 17938 differentially modulates effector memory T cells and Foxp3+ regulatory T cells in a mouse model of necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G177–G186. [Google Scholar] [CrossRef]
- Mai, T.; Liu, Y.; Hoang, T.K.; He, B.; Tran, D.Q.; Rhoads, J.M. Human breast milk promotes microbial proliferation and improves the immunomodulatory properties of Lactobacillus reuteri DSM 17938. Gastroenterology 2018, 154, S177. [Google Scholar] [CrossRef]
- Claud, E.C.; Walker, W.A. Bacterial colonization, probiotics, and necrotizing enterocolitis. J. Clin. Gastroenterol. 2008, 42, S46–S52. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, B.; Halpern, M.D.; Holubec, H.; Williams, C.S.; McWilliam, D.L.; Dominguez, J.A.; Stepankova, R.; Payne, C.M.; McCuskey, R.S. Epidermal growth factor reduces the development of necrotizing enterocolitis in a neonatal rat model. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 282, G156–G164. [Google Scholar] [CrossRef] [PubMed]
- Gronlund, M.M.; Gueimonde, M.; Laitinen, K.; Kociubinski, G.; Gronroos, T.; Salminen, S.; Isolauri, E. Maternal breast-milk and intestinal bifidobacteria guide the compositional development of the Bifidobacterium microbiota in infants at risk of allergic disease. Clin. Exp. Allergy 2007, 37, 1764–1772. [Google Scholar] [CrossRef] [PubMed]
- Gueimonde, M.; Laitinen, K.; Salminen, S.; Isolauri, E. Breast milk: A source of bifidobacteria for infant gut development and maturation? Neonatology 2007, 92, 64–66. [Google Scholar] [CrossRef]
- Lee, J.S.; Polin, R.A. Treatment and prevention of necrotizing enterocolitis. Semin. Neonatol. 2003, 8, 449–459. [Google Scholar] [CrossRef]
- Kulinich, A.; Liu, L. Human milk oligosaccharides: The role in the fine-tuning of innate immune responses. Carbohydr. Res. 2016, 432, 62–70. [Google Scholar] [CrossRef]
- Goehring, K.C.; Marriage, B.J.; Oliver, J.S.; Wilder, J.A.; Barrett, E.G.; Buck, R.H. Similar to Those Who Are Breastfed, Infants Fed a Formula Containing 2′-Fucosyllactose Have Lower Inflammatory Cytokines in a Randomized Controlled Trial. J. Nutr. 2016, 146, 2559–2566. [Google Scholar] [CrossRef]
- Weichert, S.; Koromyslova, A.; Singh, B.K.; Hansman, S.; Jennewein, S.; Schroten, H.; Hansman, G.S. Structural Basis for Norovirus Inhibition by Human Milk Oligosaccharides. J. Virol. 2016, 90, 4843–4848. [Google Scholar] [CrossRef]
- Karlsson, M. Evaluation of Lactobacillus reuteri DSM 17938 as Starter in Cheese Production. 2013. Available online: http://stud.epsilon.slu.se/5988/ (accessed on 7 July 2019).
- Kankaanpaa, P.; Yang, B.; Kallio, H.; Isolauri, E.; Salminen, S. Effects of polyunsaturated fatty acids in growth medium on lipid composition and on physicochemical surface properties of lactobacilli. Appl. Environ. Microbiol. 2004, 70, 129–136. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, Q.; Ren, Y.; Ruan, Z. Effect of probiotic Lactobacillus on lipid profile: A systematic review and meta-analysis of randomized, controlled trials. PLoS ONE 2017, 12, e0178868. [Google Scholar] [CrossRef] [PubMed]
- Achamrah, N.; Dechelotte, P.; Coeffier, M. Glutamine and the regulation of intestinal permeability: From bench to bedside. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.I.; Kang, K.S. N-acetylcysteine modulates lipopolysaccharide-induced intestinal dysfunction. Sci. Rep. 2019, 9, 1004. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.C.; Liu, Y.; Zhu, L.; Rhoads, J.M. Arginine and citrulline protect intestinal cell monolayer tight junctions from hypoxia-induced injury in piglets. Pediatr. Res. 2012, 72, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, J.M.; Plunkett, E.; Galanko, J.; Lichtman, S.; Taylor, L.; Maynor, A.; Weiner, T.; Freeman, K.; Guarisco, J.L.; Wu, G.Y. Serum citrulline levels correlate with enteral tolerance and bowel length in infants with short bowel syndrome. J. Pediatr. 2005, 146, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Timmons, J.; Chang, E.T.; Wang, J.Y.; Rao, J.N. Polyamines and Gut Mucosal Homeostasis. J. Gastrointest. Dig. Syst. 2012, 2, 001. [Google Scholar] [CrossRef]
- Tofalo, R.; Cocchi, S.; Suzzi, G. Polyamines and Gut Microbiota. Front. Nutr. 2019, 6, 16. [Google Scholar] [CrossRef]
- Qin, J.; Zhou, Y.J.; Krivoruchko, A.; Huang, M.; Liu, L.; Khoomrung, S.; Siewers, V.; Jiang, B.; Nielsen, J. Modular pathway rewiring of Saccharomyces cerevisiae enables high-level production of l-ornithine. Nat. Commun. 2015, 6, 8224. [Google Scholar] [CrossRef]
- Tachihara, K.; Uemura, T.; Kashiwagi, K.; Igarashi, K. Excretion of putrescine and spermidine by the protein encoded by YKL174c (TPO5) in Saccharomyces cerevisiae. J. Biol. Chem. 2005, 280, 12637–12642. [Google Scholar] [CrossRef]
- Iraporda, C.; Romanin, D.E.; Bengoa, A.A.; Errea, A.J.; Cayet, D.; Foligne, B.; Sirard, J.C.; Garrote, G.L.; Abraham, A.G.; Rumbo, M. Local Treatment with Lactate Prevents Intestinal Inflammation in the TNBS-Induced Colitis Model. Front. Immunol. 2016, 7, 651. [Google Scholar] [CrossRef]
- D’Abusco, A.S.; Politi, L.; Giordano, C.; Scandurra, R. A peptidyl-glucosamine derivative affects IKKalpha kinase activity in human chondrocytes. Arthritis Res. Ther. 2010, 12, R18. [Google Scholar]
- Veronesi, F.; Giavaresi, G.; Maglio, M.; d’Abusco, A.S.; Politi, L.; Scandurra, R.; Olivotto, E.; Grigolo, B.; Borzi, R.M.; Fini, M. Chondroprotective activity of N-acetyl phenylalanine glucosamine derivative on knee joint structure and inflammation in a murine model of osteoarthritis. Osteoarthr. Cartil. 2017, 25, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Hirvonen, J.; Salli, K.; Putaala, H.; Tiihonen, K.; Maukonen, J.; Ouwehand, A. Selective Utilization of Human Milk Oligosaccharides 2′-FL and 3-FL by Probiotic Bacteria Resulting in Different Metabolite Production by These Bacteria (P20-012-19). Curr. Dev. Nutr. 2019, 3, 1771. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Cycle | HBM + LR/Formula + LR |
|---|---|---|
| Glutamine | Glutamate Metabolism | 439 fold |
| Spermidine | Polyamine Metabolism | 44 fold |
| N-Acetylcysteine (NAC) (=glutathione precursor) | Methionine, Cysteine, SAM and Taurine Metabolism | 12 fold |
| Citrulline | Urea Cycle; Arginine and Proline Metabolism | 9 fold |
| Succinate | Glyoxylate Cycle | 5 fold |
| Subpathway | Number of Molecules Involved |
|---|---|
| Sphingomyelins | 16 |
| Monoacylglycerol | 15 |
| Long Chain Fatty Acid | 14 |
| Polyunsaturated Fatty Acid (n3 and n6) | 13 |
| Lysophospholipid | 13 |
| Plasmalogen | 10 |
| Fatty Acid, Monohydroxy | 7 |
| Medium Chain Fatty Acid | 6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mai, T.T.; Tran, D.Q.; Roos, S.; Rhoads, J.M.; Liu, Y. Human Breast Milk Promotes the Secretion of Potentially Beneficial Metabolites by Probiotic Lactobacillus reuteri DSM 17938. Nutrients 2019, 11, 1548. https://doi.org/10.3390/nu11071548
Mai TT, Tran DQ, Roos S, Rhoads JM, Liu Y. Human Breast Milk Promotes the Secretion of Potentially Beneficial Metabolites by Probiotic Lactobacillus reuteri DSM 17938. Nutrients. 2019; 11(7):1548. https://doi.org/10.3390/nu11071548
Chicago/Turabian StyleMai, Tu T., Dat Q. Tran, Stefan Roos, J. Marc Rhoads, and Yuying Liu. 2019. "Human Breast Milk Promotes the Secretion of Potentially Beneficial Metabolites by Probiotic Lactobacillus reuteri DSM 17938" Nutrients 11, no. 7: 1548. https://doi.org/10.3390/nu11071548
APA StyleMai, T. T., Tran, D. Q., Roos, S., Rhoads, J. M., & Liu, Y. (2019). Human Breast Milk Promotes the Secretion of Potentially Beneficial Metabolites by Probiotic Lactobacillus reuteri DSM 17938. Nutrients, 11(7), 1548. https://doi.org/10.3390/nu11071548