Use of an Extract of Annona muricata Linn to Prevent High-Fat Diet Induced Metabolic Disorders in C57BL/6 Mice †

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Extraction of Plant Material
2.2. Quantification of Total Phenols and Flavonoids
2.3. Quantification of Condensed Tannins
2.4. Assay of Antioxidant Activity Using the 2,2-Diphenyl-1-Picrylhydrazyl Free Radical (DPPH)
2.5. Isolation and Identification of Compounds
2.6. Ethics Statement
2.7. Acute Oral Toxicity
2.8. Animals and Experimental Design
2.9. Body Weight and Diet Intake
2.10. Biochemical Analysis
2.11. Oral Glucose Tolerance Test
2.12. Insulin Sensitivity Test
2.13. Quantification of Cytokines in the Adipose Tissue
2.14. Assessment of Body Fat and Liver Weight
2.15. Histopathological Analysis
2.16. Statistical Analyses
3. Results
3.1. Chemical Composition
3.2. Acute Oral Toxicity
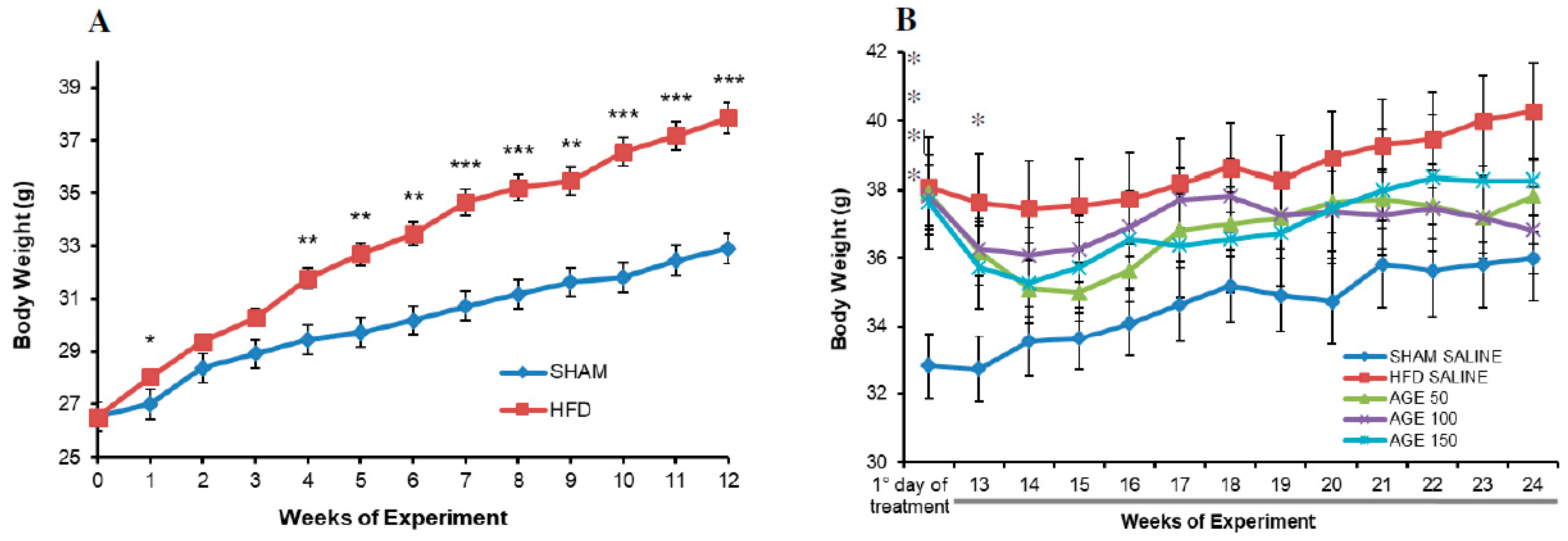
3.3. Effects of AGE on Body Weight and Food Intake
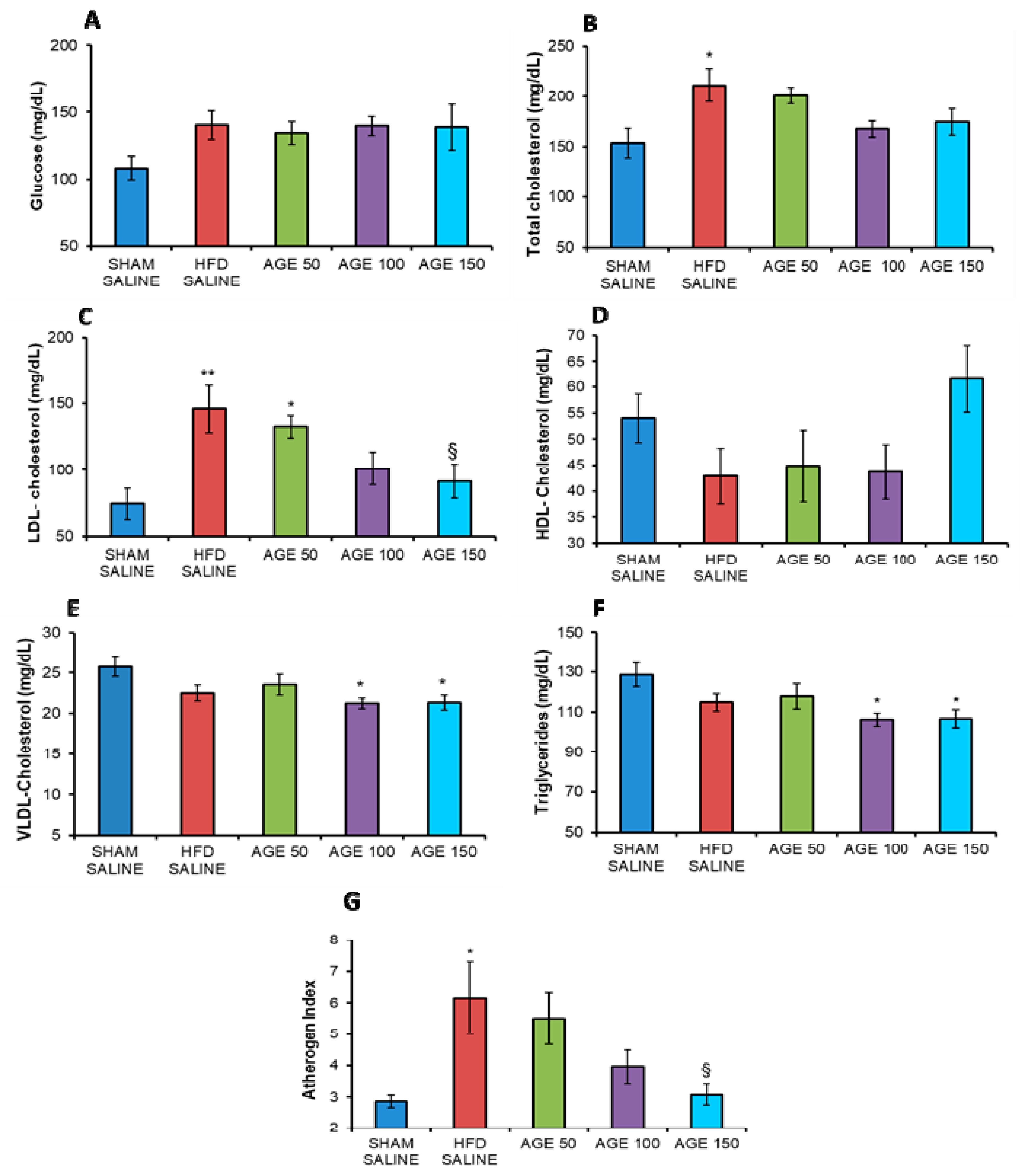
3.4. Effects of AGE on Serum Biochemical Parameters
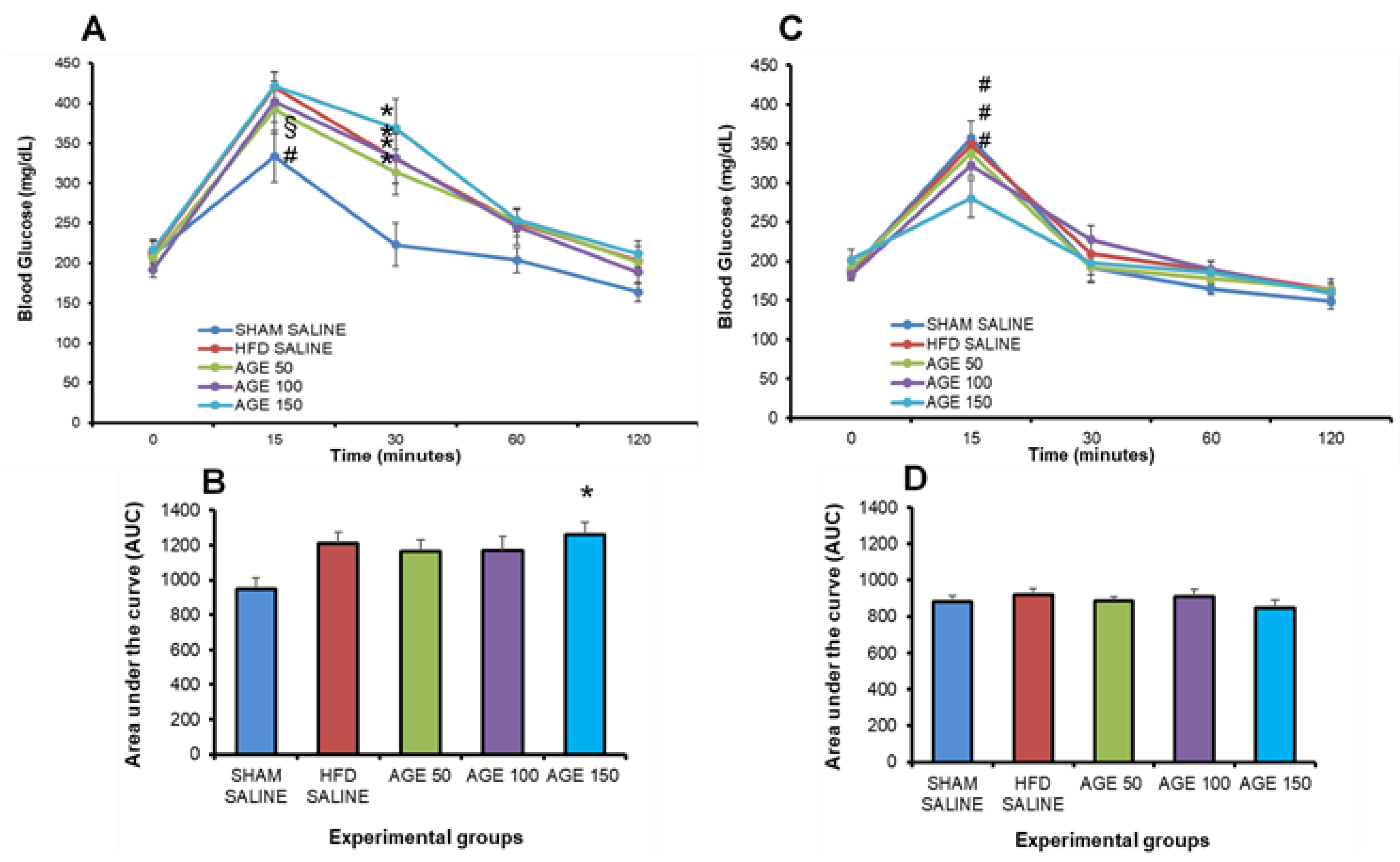
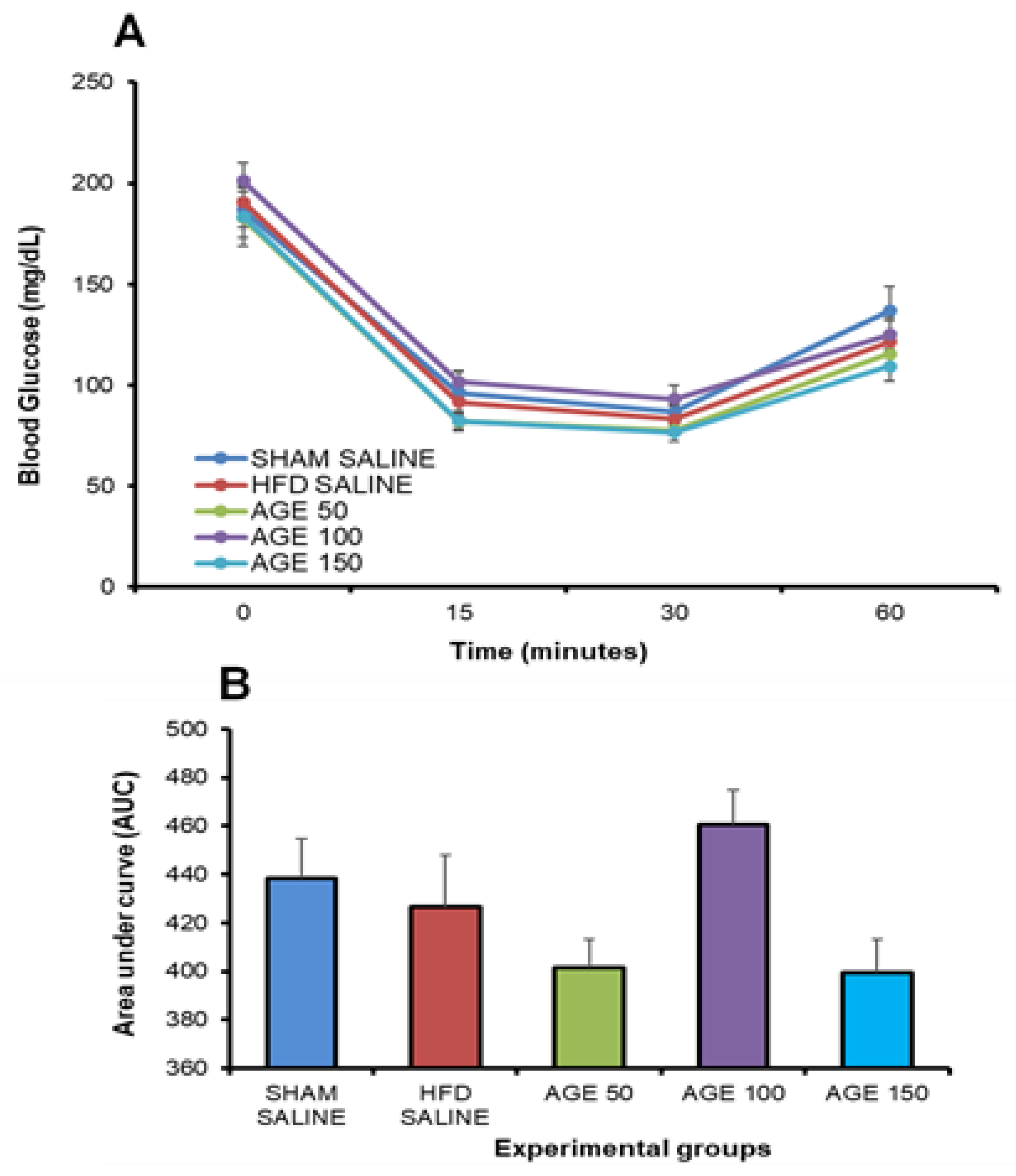
3.5. Effects of AGE on Insulin Sensitivity and Glucose Tolerance
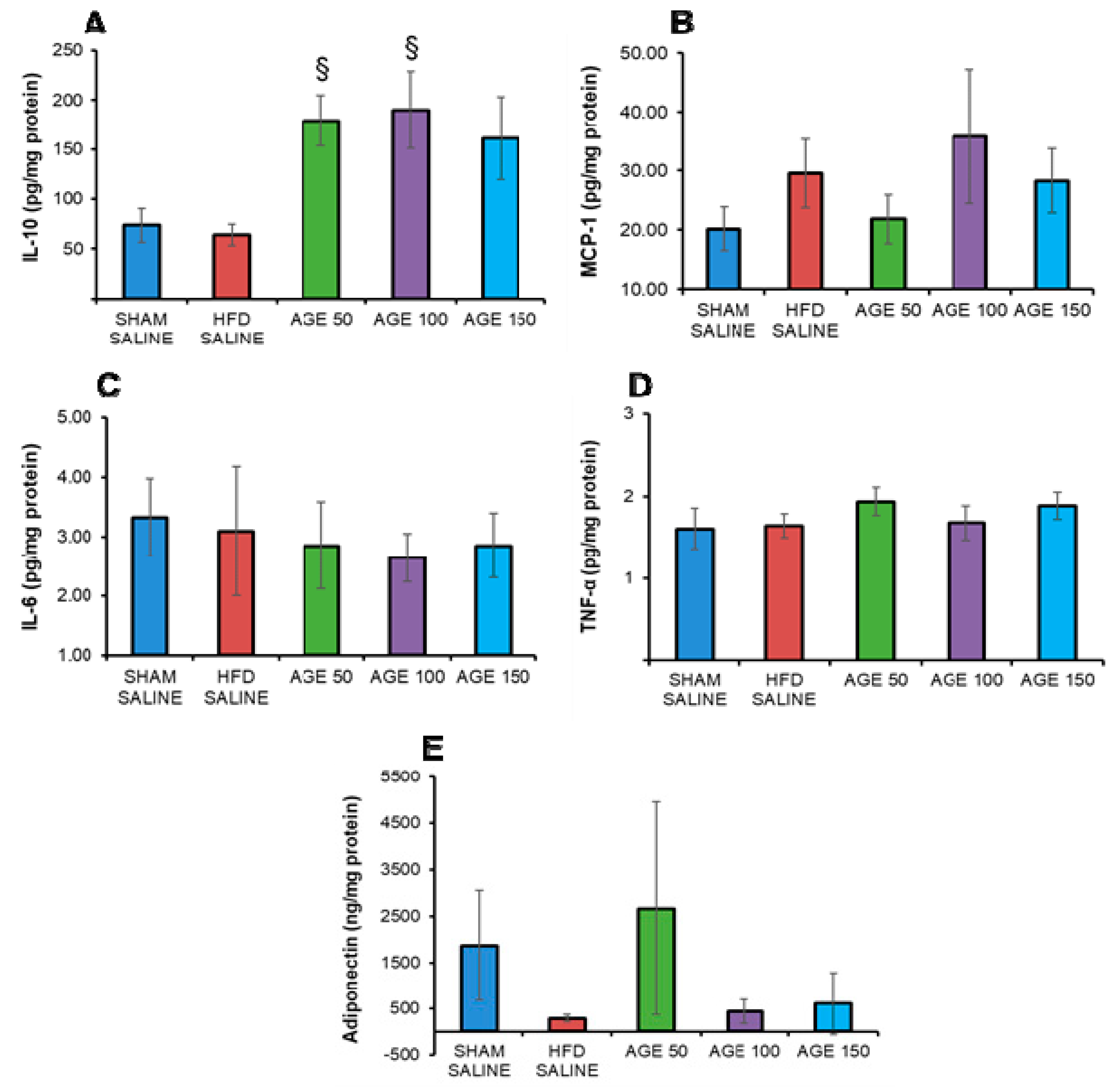
3.6. Effects of AGE on Anti- and Pro-inflammatory Cytokines, Chemokines and Adiponectin
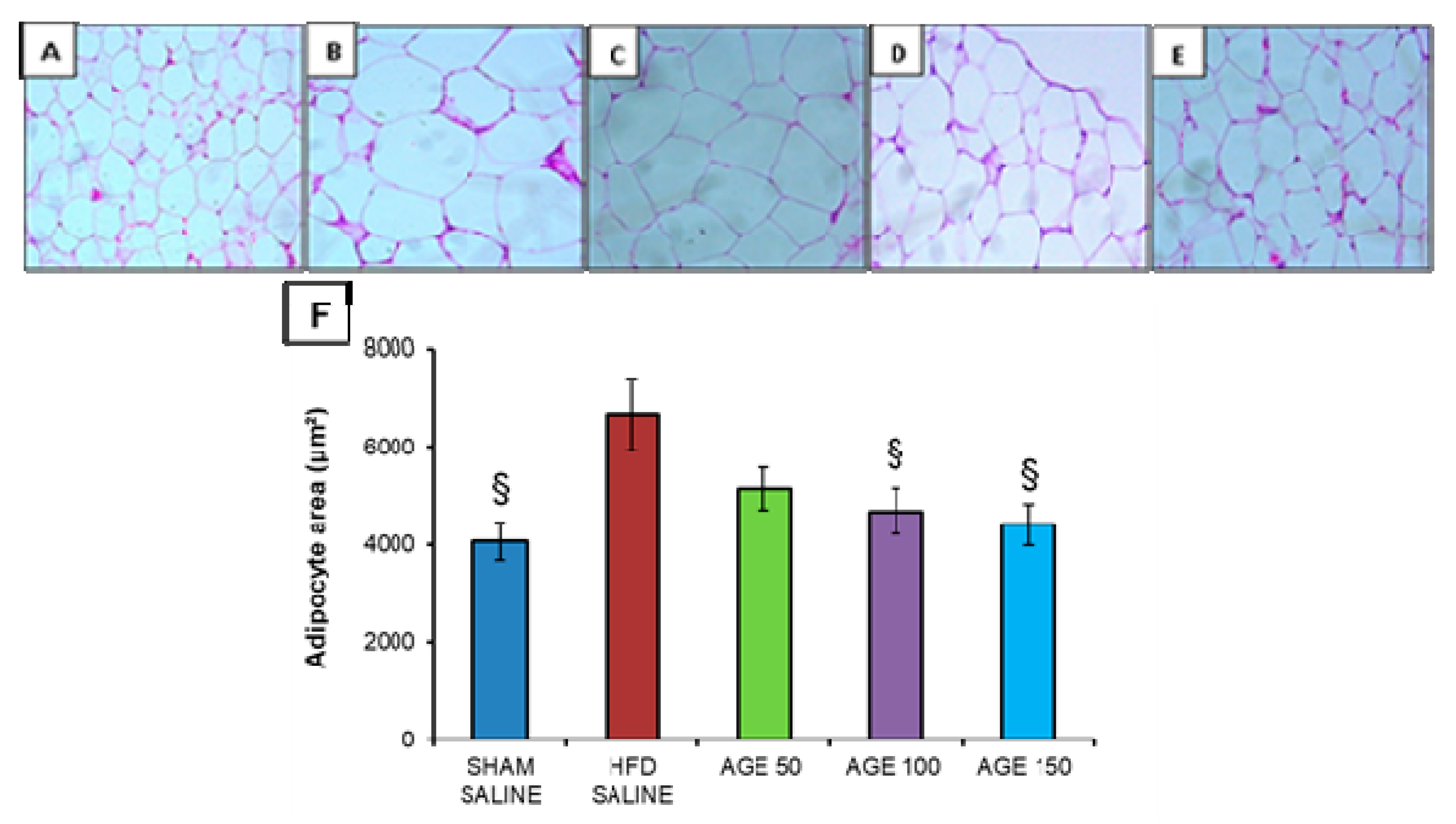
3.7. Effects of AGE on Fat Pads, Adiposity Index and Liver Weight
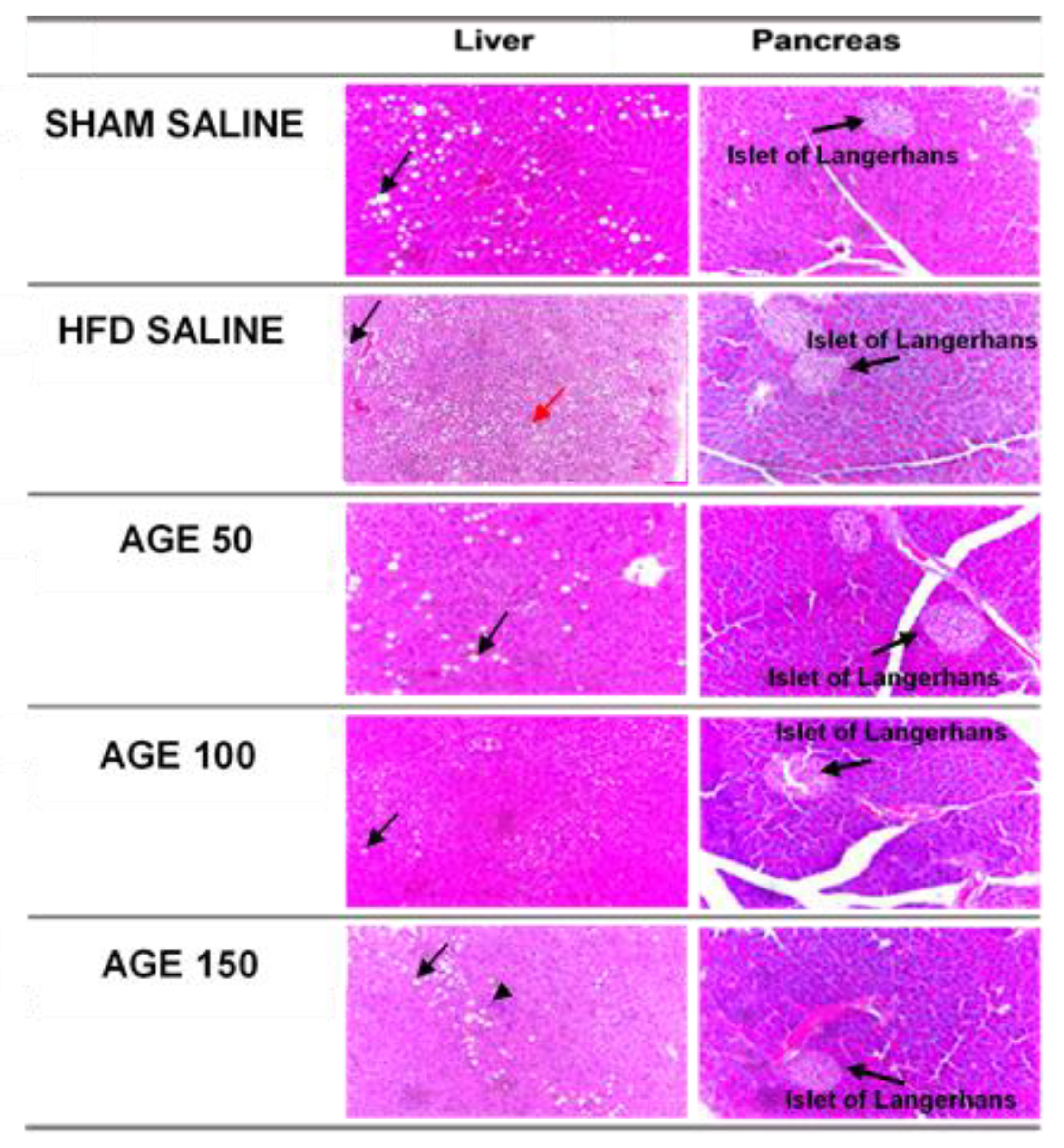
3.8. Effects of AGE on Liver, Pancreas, and Epididymal Adipose Tissue
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maki, C.; Funakoshi-Tago, M.; Aoyagi, R.; Ueda, F.; Kimura, M.; Kobata, K.; Tago, K.; Tamura, H. Coffee extract inhibits adipogenesis in 3T3-L1 preadipocyes by interrupting insulin signaling through the downregulation of IRS1. PLoS ONE 2017, 12, e0173264. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.Y.; Kim, D.S.; Kim, S.H.; Kim, H.K. Anti-obesity activity, acute toxicity, and chemical constituents of aqueous and ethanol Viola mandshurica extracts. BMC Complement. Altern. Med. 2017, 17, 297. [Google Scholar] [CrossRef] [PubMed]
- Han, T.S.; Lean, M.E. A clinical perspective of obesity, metabolic syndrome and cardiovascular disease. JRSM Cardiovasc. Dis. 2016, 5, 2048004016633371. [Google Scholar] [CrossRef] [PubMed]
- Vucenik, I.; Stains, J.P. Obesity and cancer risk: Evidence, mechanisms, and recommendations. Ann. N. Y. Acad. Sci. 2012, 1271, 37–43. [Google Scholar] [CrossRef]
- Hoving, L.R.; van der Zande, H.J.P.; Pronk, A.; Guigas, B.; Willems van Dijk, K.; van Harmelen, V. Dietary yeast-derived mannan oligosaccharides have immune-modulatory properties but do not improve high fat diet-induced obesity and glucose intolerance. PLoS ONE 2018, 13, e0196165. [Google Scholar] [CrossRef]
- Li, S.; Tan, H.Y.; Wang, N.; Zhang, Z.J.; Lao, L.; Wong, C.W.; Feng, Y. The Role of Oxidative Stress and Antioxidants in Liver Diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef]
- Showalter, M.R.; Nonnecke, E.B.; Linderholm, A.L.; Cajka, T.; Sa, M.R.; Lonnerdal, B.; Kenyon, N.J.; Fiehn, O. Obesogenic diets alter metabolism in mice. PLoS ONE 2018, 13, e0190632. [Google Scholar] [CrossRef]
- Zhang, C.; Ward, J.; Dauch, J.R.; Tanzi, R.E.; Cheng, H.T. Cytokine-mediated inflammation mediates painful neuropathy from metabolic syndrome. PLoS ONE 2018, 13, e0192333. [Google Scholar] [CrossRef]
- Gu, M.; Zhang, Y.; Fan, S.; Ding, X.; Ji, G.; Huang, C. Extracts of Rhizoma polygonati odorati prevent high-fat diet-induced metabolic disorders in C57BL/6 mice. PLoS ONE 2013, 8, e81724. [Google Scholar] [CrossRef]
- Tan, S.; Li, M.; Ding, X.; Fan, S.; Guo, L.; Gu, M.; Zhang, Y.; Feng, L.; Jiang, D.; Li, Y.; et al. Effects of Fortunella margarita fruit extract on metabolic disorders in high-fat diet-induced obese C57BL/6 mice. PLoS ONE 2014, 9, e93510. [Google Scholar] [CrossRef]
- Donado-Pestana, C.M.; Dos Santos-Donado, P.R.; Daza, L.D.; Belchior, T.; Festuccia, W.T.; Genovese, M.I. Cagaita fruit (Eugenia dysenterica DC.) and obesity: Role of polyphenols on already established obesity. Food Res. Int. 2018, 103, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.G.V.; Garcia-Diaz, D.F.; Jimenez, P.; Silva, P.I. Bioactive compounds and health benefits of exotic tropical red–black berries. J. Funct. Foods 2013, 5, 539–549. [Google Scholar] [CrossRef]
- Khan, W.; Parveen, R.; Chester, K.; Parveen, S.; Ahmad, S. Hypoglycemic Potential of Aqueous Extract of Moringa oleifera Leaf and In Vivo GC-MS Metabolomics. Front. Pharmacol. 2017, 8, 577. [Google Scholar] [CrossRef] [PubMed]
- Florence, N.T.; Benoit, M.Z.; Jonas, K.; Alexandra, T.; Desire, D.D.; Pierre, K.; Theophile, D. Antidiabetic and antioxidant effects of Annona muricata (Annonaceae), aqueous extract on streptozotocin-induced diabetic rats. J. Ethnopharmacol. 2014, 151, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Justino, A.B.; Miranda, N.C.; Franco, R.R.; Martins, M.M.; Silva, N.M.D.; Espindola, F.S. Annona muricata Linn. leaf as a source of antioxidant compounds with in vitro antidiabetic and inhibitory potential against alpha-amylase, alpha-glucosidase, lipase, non-enzymatic glycation and lipid peroxidation. Biomed. Pharmacother. 2018, 100, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Djeridane, A.; Yousfi, M.; Nadjemi, B.; Boutassouna, D.; Stocker, P.; Vidal, N. Antioxidant activity of some algerian medicinal plants extracts containing phenolic compounds. Food Chem. 2006, 97, 654–660. [Google Scholar] [CrossRef]
- Lin, J.-Y.; Tang, C.-Y. Determination of total phenolic and flavonoid contents in selected fruits and vegetables, as well as their stimulatory effects on mouse splenocyte proliferation. Food Chem. 2007, 101, 140–147. [Google Scholar] [CrossRef]
- Broadhurst, R.B.; Jones, W.T. Analysis of condensed tannins using acidified vanillin. J. Sci. Food Agric. 1978, 29, 788–794. [Google Scholar] [CrossRef]
- Kumaran, A.; Karunakaran, R.J. Nitric oxide radical scavenging active components from Phyllanthus emblica L. Plant Foods Hum. Nutr. 2006, 61, 1–5. [Google Scholar] [CrossRef]
- Berrondo, L.F.; Gabriel, F.T.; Fernandes, S.B.d.O.; Menezes, F.d.S.; Moreira, D.d.L. Dirhamnosyl flavonoid and other constituents from Brillantaisia palisatii. Química Nova 2003, 26, 922–923. [Google Scholar] [CrossRef]
- Harborne, J.B. The Flavonoids: Advances in Research Since 1986 (Harborne, J.B.). J. Chem. Educ. 1995, 72, 73. [Google Scholar]
- Agrawal, P.K. Carbon-13 NMR of Flavonoids, 1st ed.; Agrawal, P.K., Ed.; Elsevier Science: Amsterdam, The Netherlands, 1989; Volume 39, pp. 283–364. [Google Scholar]
- OECD. Test No. 425: Acute Oral Toxicity: Up-and-Down Procedure; OECD Publishing: Paris, France, 2008. [Google Scholar] [CrossRef]
- Malone, M.H.; Robichaud, R.C. A Hippocratic screen for pure or crude drug materials. Lloydia 1962, 25, 320–322. [Google Scholar]
- Da Silva, E.R.; Salmazzo, G.R.; da Silva Arrigo, J.; Oliveira, R.J.; Kassuya, C.A.; Cardoso, C.A. Anti-inflammatory Evaluation and Toxicological Analysis of Campomanesia xanthocarpa Berg. Inflammation 2016, 39, 1462–1468. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Hargrove, R.E.; Alford, J.A. Growth Rate and Feed Efficiency of Rats Fed Yogurt and Other Fermented Milks. J. Dairy Sci. 1978, 61, 11–19. [Google Scholar] [CrossRef]
- Santos, S.H.; Fernandes, L.R.; Mario, E.G.; Ferreira, A.V.; Porto, L.C.; Alvarez-Leite, J.I.; Botion, L.M.; Bader, M.; Alenina, N.; Santos, R.A. Mas deficiency in FVB/N mice produces marked changes in lipid and glycemic metabolism. Diabetes 2008, 57, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Donatto, F.F.; Neves, R.X.; Rosa, F.O.; Camargo, R.G.; Ribeiro, H.; Matos-Neto, E.M.; Seelaender, M. Resistance exercise modulates lipid plasma profile and cytokine content in the adipose tissue of tumour-bearing rats. Cytokine 2013, 61, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Lira, F.S.; Rosa, J.C.; Pimentel, G.D.; Tarini, V.A.; Arida, R.M.; Faloppa, F.; Alves, E.S.; do Nascimento, C.O.; Oyama, L.M.; Seelaender, M.; et al. Inflammation and adipose tissue: Effects of progressive load training in rats. Lipids Health Dis. 2010, 9, 109. [Google Scholar] [CrossRef] [PubMed]
- White, P.A.; Cercato, L.M.; Batista, V.S.; Camargo, E.A.; De Lucca, W.; Oliveira, A.S.; Silva, F.T.; Goes, T.C.; Oliveira, E.R.; Moraes, V.R.; et al. Aqueous extract of Chrysobalanus icaco leaves, in lower doses, prevent fat gain in obese high-fat fed mice. J. Ethnopharmacol. 2016, 179, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Chandran, R.; Parimelazhagan, T.; Shanmugam, S.; Thankarajan, S. Antidiabetic activity of Syzygium calophyllifolium in Streptozotocin-Nicotinamide induced Type-2 diabetic rats. Biomed. Pharmacother. 2016, 82, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xue, Y.; Wang, B.; Zhao, J.; Yan, X.; Huang, Y.; Du, M.; Zhu, M.J. Maternal obesity exacerbates insulitis and type 1 diabetes in non-obese diabetic mice. Reproduction 2014, 148, 73–79. [Google Scholar] [PubMed]
- Pereira, S.S.; Teixeira, L.G.; Aguilar, E.C.; Matoso, R.O.; Soares, F.L.; Ferreira, A.V.; Alvarez-Leite, J.I. Differences in adipose tissue inflammation and oxidative status in C57BL/6 and ApoE-/- mice fed high fat diet. Anim. Sci. J. 2012, 83, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Cercato, L.M.; White, P.A.; Nampo, F.K.; Santos, M.R.; Camargo, E.A. A systematic review of medicinal plants used for weight loss in Brazil: Is there potential for obesity treatment? J. Ethnopharmacol. 2015, 176, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Verma, N.; Shukla, S. Impact of various factors responsible for fluctuation in plant secondary metabolites. J. Appl. Res. Med. Aromat. Plants 2015, 2, 105–113. [Google Scholar] [CrossRef]
- George, V.C.; Kumar, D.R.; Suresh, P.K.; Kumar, R.A. Antioxidant, DNA protective efficacy and HPLC analysis of Annona muricata (soursop) extracts. J. Food Sci. Technol. 2015, 52, 2328–2335. [Google Scholar]
- Gavamukulya, Y.; Abou-Elella, F.; Wamunyokoli, F.; AEl-Shemy, H. Phytochemical screening, anti-oxidant activity and in vitro anticancer potential of ethanolic and water leaves extracts of Annona muricata (Graviola). Asian Pac. J. Trop. Med. 2014, 7, S355–S363. [Google Scholar] [CrossRef]
- Coria-Téllez, A.V.; Montalvo-Gónzalez, E.; Yahia, E.M.; Obledo-Vázquez, E.N. Annona muricata: A comprehensive review on its traditional medicinal uses, phytochemicals, pharmacological activities, mechanisms of action and toxicity. Arab. J. Chem. 2018, 11, 662–691. [Google Scholar] [CrossRef]
- Moghadamtousi, S.Z.; Fadaeinasab, M.; Nikzad, S.; Mohan, G.; Ali, H.M.; Kadir, H.A. Annona muricata (Annonaceae): A Review of Its Traditional Uses, Isolated Acetogenins and Biological Activities. Int. J. Mol. Sci. 2015, 16, 15625–15658. [Google Scholar] [CrossRef]
- Nawwar, M.; Ayoub, N.; Hussein, S.; Hashim, A.; El-Sharawy, R.; Wende, K.; Harms, M.; Lindequist, U. A flavonol triglycoside and investigation of the antioxidant and cell stimulating activities of Annona muricata Linn. Arch. Pharm. Res. 2012, 35, 761–767. [Google Scholar] [CrossRef]
- Panchal, S.K.; Brown, L. Rodent models for metabolic syndrome research. J. Biomed. Biotechnol. 2011, 2011, 351982. [Google Scholar] [PubMed]
- Della Vedova, M.C.; Muñoz, M.D.; Santillan, L.D.; Plateo-Pignatari, M.G.; Germanó, M.J.; Tosi, M.E.R.; Garcia, S.; Gomez, N.N.; Fornes, M.W.; Mejiba, S.E.G.; et al. A Mouse Model of Diet-Induced Obesity Resembling Most Features of Human Metabolic Syndrome. Nutr. Metab. Insights 2016, 9, NMI.S32907. [Google Scholar]
- Rupasinghe, H.P.; Sekhon-Loodu, S.; Mantso, T.; Panayiotidis, M.I. Phytochemicals in regulating fatty acid beta-oxidation: Potential underlying mechanisms and their involvement in obesity and weight loss. Pharmacol. Ther. 2016, 165, 153–163. [Google Scholar] [PubMed]
- Boadi, W.Y.; Lo, A. Effects of Quercetin, Kaempferol, and Exogenous Glutathione on Phospho- and Total-AKT in 3T3-L1 Preadipocytes. J. Diet Suppl. 2018, 15, 814–826. [Google Scholar] [PubMed]
- Adewole, S.O.; Ojewole, J.A. Protective effects of Annona muricata Linn. (Annonaceae) leaf aqueous extract on serum lipid profiles and oxidative stress in hepatocytes of streptozotocin-treated diabetic rats. Afr. J. Tradit. Complement. Altern. Med. 2008, 6, 30–41. [Google Scholar]
- Chernukha, I.M.; Fedulova, L.V.; Kotenkova, E.A.; Takeda, S.; Sakata, R. Hypolipidemic and anti-inflammatory effects of aorta and heart tissues of cattle and pigs in the atherosclerosis rat model. Anim. Sci. J. 2018, 89, 784–793. [Google Scholar] [PubMed]
- Mazaherioun, M.; Djalali, M.; Koohdani, F.; Javanbakht, M.H.; Zarei, M.; Beigy, M.; Ansari, S.; Rezvan, N.; Saedisomeolia, A. Beneficial Effects of n-3 Fatty Acids on Cardiometabolic and Inflammatory Markers in Type 2 Diabetes Mellitus: A Clinical Trial. Med. Princ. Pract. 2017, 26, 535–541. [Google Scholar]
- Adeyemi, D.O.; Komolafe, O.A.; Adewole, O.S.; Obuotor, E.M.; Adenowo, T.K. Anti hyperglycemic activities of Annona muricata (Linn). Afr. J. Tradit. Complement. Altern. Med. 2008, 6, 62–69. [Google Scholar]
- Ahalya, B.; Shankar, K.R.; Kiranmayi, G.V.N. Exploration of anti-hyperglycemic and hypolipidemic activities of ethanolic extract of Annona muricata bark in alloxan induced diabetic rats. Int. J. Pharm. Sci. Rev. Res. 2014, 25, 21–27. [Google Scholar]
- Cavalot, F. Do data in the literature indicate that glycaemic variability is a clinical problem? Glycaemic variability and vascular complications of diabetes. Diabetes Obes. Metab. 2013, 15 (Suppl. 2), 3–8. [Google Scholar] [CrossRef]
- Ceriello, A.; Esposito, K.; Piconi, L.; Ihnat, M.; Thorpe, J.; Testa, R.; Bonfigli, A.R.; Giugliano, D. Glucose “peak” and glucose “spike”: Impact on endothelial function and oxidative stress. Diabetes Re. Clin. Pract. 2008, 82, 262–267. [Google Scholar]
- Galan, A.; Mayer, I.; Rafaj, R.B.; Bendelja, K.; Susic, V.; Ceron, J.J.; Mrljak, V. MCP-1, KC-like and IL-8 as critical mediators of pathogenesis caused by Babesia canis. PLoS ONE 2018, 13, e0190474. [Google Scholar] [CrossRef] [PubMed]
- Veloso, C.C.; Oliveira, M.C.; Rodrigues, V.G.; Oliveira, C.C.; Duarte, L.P.; Teixeira, M.M.; Ferreira, A.V.M.; Perez, A.C. Evaluation of the effects of extracts of Maytenus imbricata (Celastraceae) on the treatment of inflammatory and metabolic dysfunction induced by high-refined carbohydrate diet. Inflammopharmacology 2018, 27, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, D.; Yin, C.; Wang, S.; Wang, M.; Xiao, Y. IL-10/STAT3 is reduced in childhood obesity with hypertriglyceridemia and is related to triglyceride level in diet-induced obese rats. BMC Endocr. Disord. 2018, 18, 39. [Google Scholar] [CrossRef] [PubMed]
- Scoditti, E.; Massaro, M.; Carluccio, M.A.; Pellegrino, M.; Wabitsch, M.; Calabriso, N.; Storelli, C.; De Caterina, R. Additive regulation of adiponectin expression by the mediterranean diet olive oil components oleic Acid and hydroxytyrosol in human adipocytes. PLoS ONE 2015, 10, e0128218. [Google Scholar]
- Wu, T.; Tang, Q.; Gao, Z.; Yu, Z.; Song, H.; Zheng, X.; Chen, W. Blueberry and mulberry juice prevent obesity development in C57BL/6 mice. PLoS ONE 2013, 8, e77585. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Groups | AIN-93M Diet | High-Fat Diet (HFD) |
|---|---|---|
| Composition (g/kg) | ||
| Cornstarch | 620.69 | 320.69 |
| Casein | 140.00 | 140.00 |
| Fat | - | 320.00 |
| Sucrose | 100.00 | 100.00 |
| Soybean oil | 40.00 | 20.00 |
| Fiber | 50.00 | 50.00 |
| Mineral mix | 35.00 | 35.00 |
| Vitamin mix | 10.00 | 10.00 |
| L-cystine | 1.80 | 1.80 |
| Choline bitartrate | 2.50 | 2.50 |
| Tert-butylhydroquinone | 0.008 | 0.008 |
| Energy (kcal/kg) | 3802.8 | 5302.8 |
| Carbohydrates (%) | 75.81 | 31.73 |
| Protein (%) | 14.73 | 10.56 |
| Lipids (%) | 9.47 | 57.71 |
| Calories/g of diet | 3.80 | 5.30 |
| Parameter | Experimental Group | |
|---|---|---|
| SHAM (n = 11) | HFD (n = 44) | |
| Initial weight (g) | 26.55 ± 0.55 | 26.52 ± 0.28 |
| Final weight (g) | 32.91 ± 0.99 | 37.87 ± 0.60 *** |
| Weight gain (g) | 6.36 ± 0.95 | 11.34 ± 0.68 ** |
| Food intake (g/day) | 3.47 ± 0.05 | 2.73 ± 0.03 *** |
| Food intake (kcal/day) | 13.18 ± 0.19 | 14.54 ± 0.16 *** |
| Feed efficiency index | 0.0216 ± 0.0031 | 0.0501 ± 0.0033 *** |
| Parameter | Experimental Group | ||||
|---|---|---|---|---|---|
| SHAM SALINE | HFD SALINE | AGE 50 | AGE 100 | AGE 150 | |
| Initial weight (g) | 32.89 ± 0.94 | 38.09 ± 1.42 * | 37.91 ± 1.09 * | 37.82 ± 0.91 * | 37.64 ± 1.43 * |
| Final weight (g) | 35.91 ± 1.26 | 40.27 ± 1.42 | 37.82 ± 1.05 | 36.82 ± 1.33 | 38.27 ± 1.94 |
| Weight gain (g) | 3.09 ± 0.60 | 2.18 ± 0.26 | −0.09 ± 0.53 * | −1.00 ± 1.14 **,§ | 0.64 ± 0.92 |
| Food intake (g/day) | 3.40 ± 0.43 | 2.62 ± 0.62 *** | 2.76 ± 0.05 *** | 2.70 ± 0.98 *** | 2.83 ± 0.07 *** |
| Food intake (kcal/day) | 12.89 ± 0.16 | 13.89 ± 0.33 | 14.65 ± 0.25 ** | 14.30 ± 0.52 * | 14.99 ± 0.37 *** |
| Feed efficiency index | 0.0109 ± 0.0021 | 0.010 ± 0.00128 | −0.0006 ± 0.0023 | −0.0036 ± 0.0050 *,§ | 0.0031 ± 0.0040 |
| Parameter | Experimental Group | ||||
|---|---|---|---|---|---|
| SHAM SALINE | HFD SALINE | AGE 50 | AGE 100 | AGE 150 | |
| Omental weight (g) | 0.028 ± 0.009 | 0.018 ± 0.005 | 0.014 ± 0.004 | 0.016 ± 0.004 | 0.012 ± 0.004 |
| Epididymal weight (g) | 1.068 ± 0.127 | 1.506 ± 0.112 | 1.543 ± 0.783 | 1.290 ± 0.149 | 1.413 ± 0.169 |
| Mesenteric weight (g) | 0.496 ± 0.634 | 0.707 ± 0.092 | 0.659 ± 0.049 | 0.556 ± 0.122 | 0.664 ± 0.136 |
| Retroperitoneal weight (g) | 0.360 ± 0.053 | 0.728 ± 0.087 * | 0.664 ± 0.053 | 0.502 ± 0.090 | 0.705 ± 0.125 |
| Perirenal weight (g) | 0.198 ± 0.039 | 0.255 ± 0.039 | 0.231 ± 0.026 | 0.275 ± 0.061 | 0.198 ± 0.036 |
| Adiposity index (%) | 6.080 ± 0.514 | 10.893 ± 0.481 *** | 11.620 ± 0.387 *** | 10.212 ± 0.797 *** | 10.713 ± 0.779 *** |
| Liver (g) | 1.225 ± 0.066 | 1.236 ± 0.056 | 1.213 ± 0.030 | 1.200 ± 0.058 | 1.185 ± 0.047 |
| Variable | Experimental Group | ||||
|---|---|---|---|---|---|
| SHAM SALINE | HFD SALINE | AGE 50 | AGE 100 | AGE 150 | |
| Changes in the pancreas | |||||
| Islet of Langerhans (p = 0.291) | |||||
| No change | 36.4 (4) | 45.5 (5) | 72.7 (8) | 80.0 (8) | 54.5 (6) |
| Discrete atrophy | 9.1 (1) | 0.0 (0) | 0.0 (0) | 0.0 (0) | 9.1 (1) |
| Atrophy | 18.2 (2) | 36.4 (4) | 9.1 (1) | 0.0 (0) | 0.0 (0) |
| Discrete hypertrophy | 18.2 (2) | 9.1 (1) | 0.0 (0) | 20.0 (2) | 27.3 (3) |
| Hypertrophy | 18.2 (2) | 9.1 (1) | 18.2 (2) | 0.0 (0) | 9.1 (1) |
| Pancreatic acini (p = 0.400) | |||||
| No change | 81.8 (9) | 72.7 (8) | 90.9 (10) | 100.0 (10) | 90.9 (10) |
| Necrosis/Atrophy | 18.2 (2) | 27.3 (3) | 9.1 (1) | 0.0 (0) | 9.1 (1) |
| Inflammatory cells (p = 0.458) | |||||
| No change | 90.9 (10) | 81.8 (9) | 90.9 (10) | 100.0 (10) | 100.0 (11) |
| Insulitis | 0.0 (0) | 0.0 (0) | 0.0 (0) | 0.0 (0) | 0.0 (0) |
| Perinsulitis | 9.1 (1) | 18.2 (2) | 9.1 (1) | 0.0 (0) | 0.0 (0) |
| Variable | Experimental Group | ||||
|---|---|---|---|---|---|
| SHAM SALINE | HFD SALINE | AGE 50 | AGE 100 | AGE 150 | |
| Liver Changes | |||||
| Steatosis (p = 0.881) | |||||
| < 5% | 54.5 (5) | 36.4 (4) | 54.5 (6) | 60.0 (6) | 54.5 (6) |
| 5 to 33% | 36.4 (4) | 36.4 (4) | 36.4 (4) | 10.0 (1) | 36.4 (4) |
| 34 to 66% | 9.1 (1) | 18.2 (2) | 9.1 (1) | 20.0 (2) | 9.1 (1) |
| >66% | 0.0 (0) | 9.1 (1) | 0.0 (0) | 10.0 (1) | 0.0 (0) |
| Microvesicular steatosis (p = 0.501) | |||||
| Absent | 45.5 (5) | 18.2 (2) | 54.5 (6) | 40.0 (4) | 36.4 (4) |
| Present | 54.5 (6) | 81.8 (9) | 45.5 (5) | 60.0 (6) | 63.6 (7) |
| Lobular inflammation (p = 0.919) | |||||
| Absent | 63.6 (7) | 72.7 (8) | 81.8 (9) | 70.7 (7) | 72.7 (8) |
| <1 focus/field | 36.4 (4) | 27.3 (3) | 18.2 (2) | 30.0 (3) | 27.3 (3) |
| 2–4 focuses/field | 0.0 (0) | 0.0 (0) | 0.0 (0) | 0.0 (0) | 0.0 (0) |
| > 4 focuses/field | 0.0 (0) | 0.0 (0) | 0.0 (0) | 0.0 (0) | 0.0 (0) |
| Ballooning (p = 0.91) | |||||
| Absent | 72.7 (8) | 36.4 (4) | 81.8 (9) | 70.7 (7) | 72.7 (8) |
| Few cells | 27.3 (3) | 63.6 (7) | 18.2 (2) | 30.0 (3) | 27.3 (3) |
| Many cells | 0.0 (0) | 0.0 (0) | 0.0 (0) | 0.0 (0) | 0.0 (0) |
| Mallory’s hyaline (p = 0.91) | |||||
| Absent | 100.0 (11) | 100.0 (11) | 100.0 (11) | 100.0 (10) | 90.9 (10) |
| Present | 0.0 (0) | 0.0 (0) | 0.0 (0) | 0.0 (0) | 9.1 (1) |
| Apoptosis | |||||
| Absent | 100.0 (11) | 100.0 (11) | 100.0 (11) | 100.0 (10) | 100.0 (11) |
| Present | 0.0 (0) | 0.0 (0) | 0.0 (0) | 0.0 (0) | 0.0 (0) |
| Glycogenate nucleus (p = 0.408) | |||||
| None/rare | 100.0 (11) | 100.0 (11) | 100.0 (11) | 100.0 (10) | 90.9 (10) |
| Some | 0.0 (0) | 0.0 (0) | 0.0 (0) | 0.0 (0) | 9.1 (1) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sasso, S.; Sampaio e Souza, P.C.; Santana, L.F.; Cardoso, C.A.L.; Alves, F.M.; Portugal, L.C.; de Faria, B.B.; da Silva, A.F.; Motta-Castro, A.R.C.; Soares, L.S.; et al. Use of an Extract of Annona muricata Linn to Prevent High-Fat Diet Induced Metabolic Disorders in C57BL/6 Mice. Nutrients 2019, 11, 1509. https://doi.org/10.3390/nu11071509
Sasso S, Sampaio e Souza PC, Santana LF, Cardoso CAL, Alves FM, Portugal LC, de Faria BB, da Silva AF, Motta-Castro ARC, Soares LS, et al. Use of an Extract of Annona muricata Linn to Prevent High-Fat Diet Induced Metabolic Disorders in C57BL/6 Mice. Nutrients. 2019; 11(7):1509. https://doi.org/10.3390/nu11071509
Chicago/Turabian StyleSasso, Sandramara, Priscilla Cristovam Sampaio e Souza, Lidiani Figueiredo Santana, Claudia Andréa Lima Cardoso, Flávio Macedo Alves, Luciane Candeloro Portugal, Bernardo Bacelar de Faria, Anderson Fernandes da Silva, Ana Rita Coimbra Motta-Castro, Luana Silva Soares, and et al. 2019. "Use of an Extract of Annona muricata Linn to Prevent High-Fat Diet Induced Metabolic Disorders in C57BL/6 Mice" Nutrients 11, no. 7: 1509. https://doi.org/10.3390/nu11071509
APA StyleSasso, S., Sampaio e Souza, P. C., Santana, L. F., Cardoso, C. A. L., Alves, F. M., Portugal, L. C., de Faria, B. B., da Silva, A. F., Motta-Castro, A. R. C., Soares, L. S., Bandeira, L. M., Guimarães, R. d. C. A., & Freitas, K. d. C. (2019). Use of an Extract of Annona muricata Linn to Prevent High-Fat Diet Induced Metabolic Disorders in C57BL/6 Mice. Nutrients, 11(7), 1509. https://doi.org/10.3390/nu11071509