Can Lycopene Impact the Androgen Axis in Prostate Cancer?: A Systematic Review of Cell Culture and Animal Studies


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Selection Criteria
2.2. Literature Search
2.3. Data Extraction and Quality Assessment
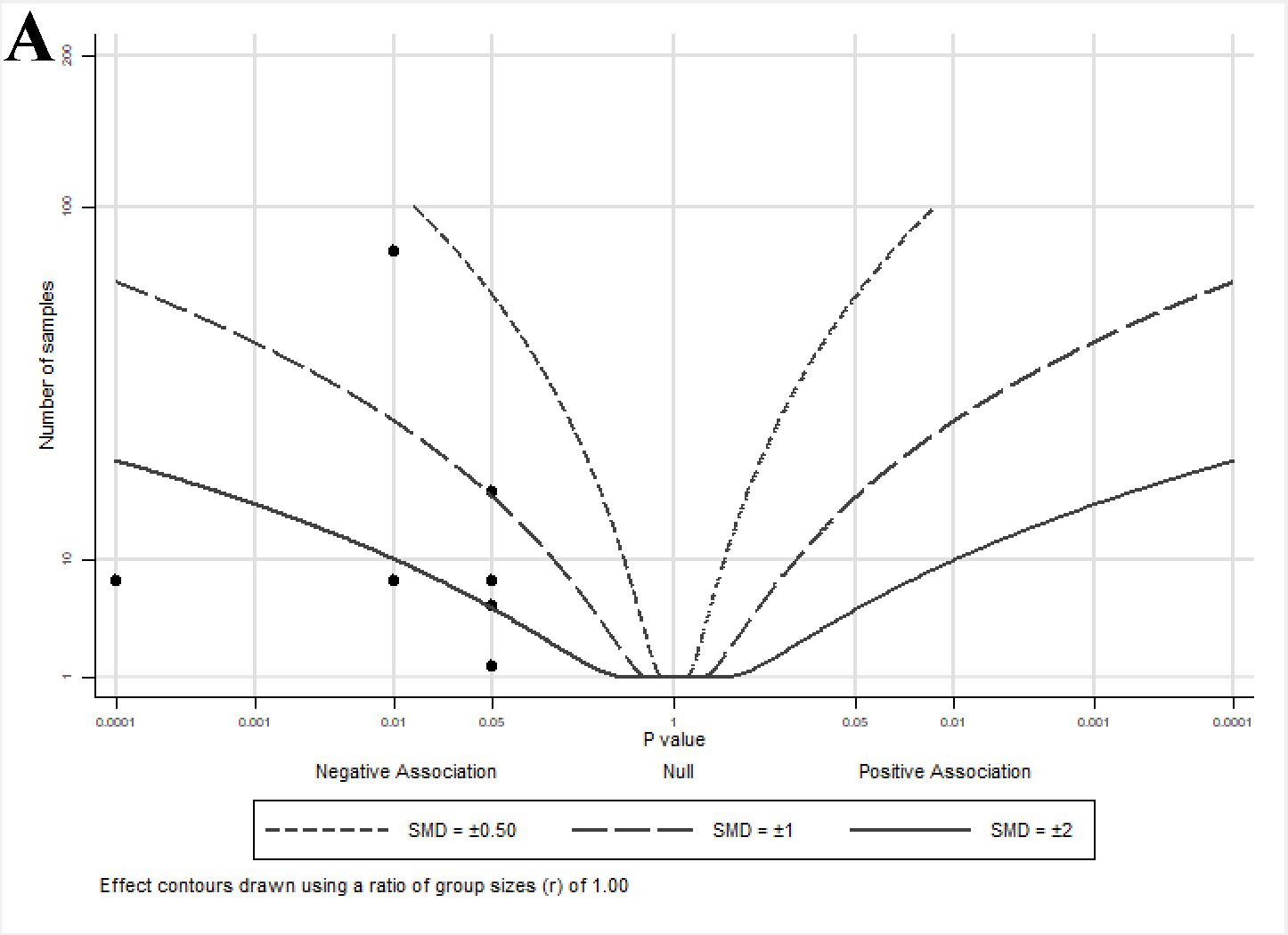
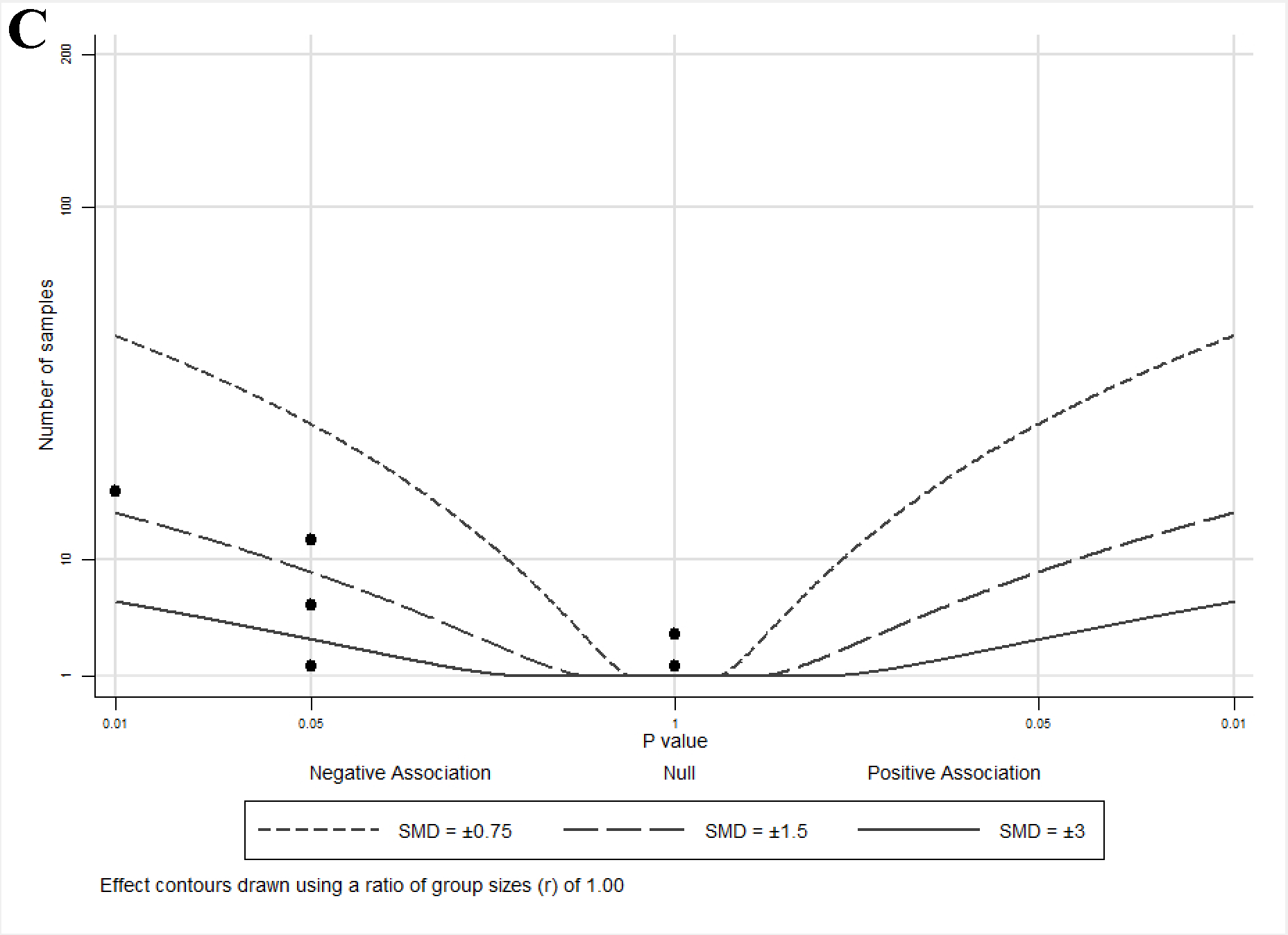
2.4. Albatross Plot Generation
3. Results
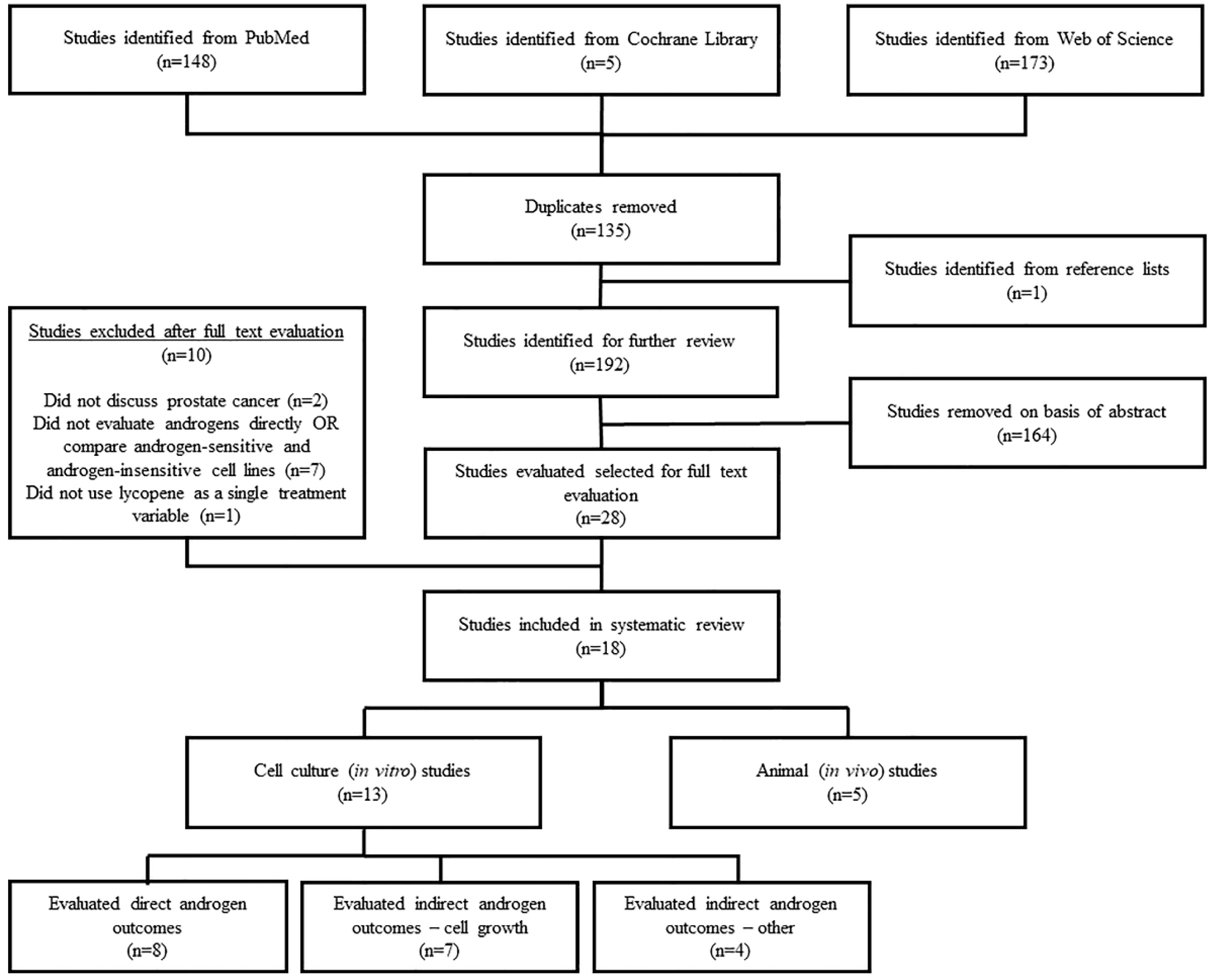
3.1. Literature Search
3.2. Study Characteristics
3.3. Animal Studies
3.4. Cell Culture Studies
3.4.1. Influence of Lycopene on Indirect Androgen Outcomes: Comparison of Androgen-Sensitive vs. Androgen-Insensitive Cell Lines
3.4.2. Influence of Lycopene on Direct Androgen Outcomes
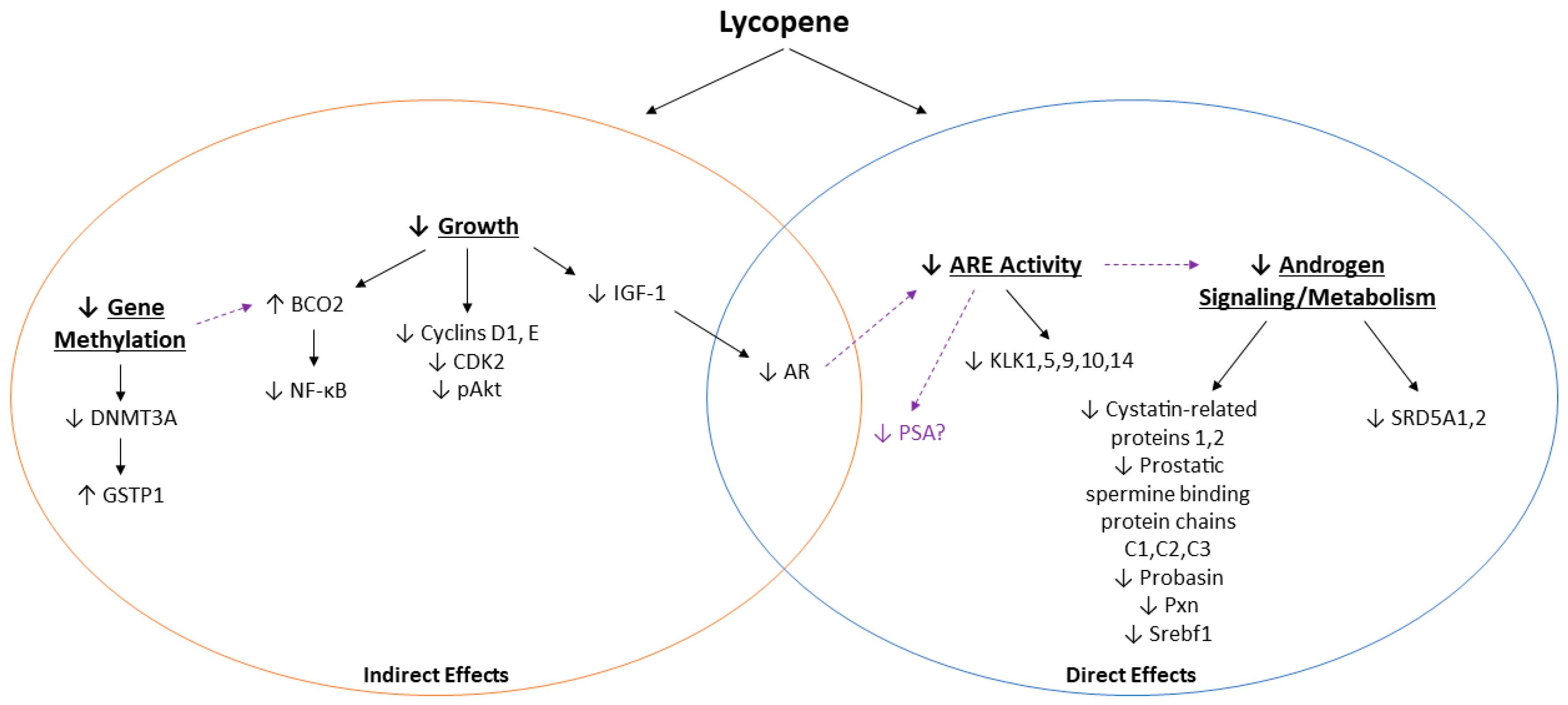
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author, Year | Selection Bias | Performance Bias | Detection Bias | Attrition Bias 8 | Reporting Bias 9 | Other Bias 10 | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sequence Generation 1 | Baseline 2 | Allocation Concealment 3 | Random Housing 4 | Blinding 5 | Random Outcome Assessment 6 | Blinding 7 | ||||
| Canene-Adams, 2009 [27] | Unclear | Low | Unclear | Low | Unclear | Unclear | Unclear | Low | Low | Low |
| Limpens, 2006 [28] | Low | Low | Unclear | Low | Unclear | Unclear | Low | Low | Low | Low |
| Lindshield, 2010 [29] | Unclear | Low | Unclear | Low | Unclear | Unclear | Unclear | Low | Low | Low |
| Siler, 2004 [30] | Low | Low | Unclear | Low | Unclear | Unclear | Unclear | Low | Low | Low |
| Wan, 2014 [31] | Low | Low | Unclear | Low | Unclear | Unclear | Unclear | Low | Low | Low |
| Author, Year | Source 1 | Experimental Conditions | Selective Reporting 6 | Total | |||
|---|---|---|---|---|---|---|---|
| Culture Conditions 2 | Replicates 3 | Controls 4 | Multiple Cell Lines 5 | ||||
| Fu, 2014 [44] | 1 | 1 | 0 | 1 | 1 | 1 | 5 |
| Gong, 2016 [40] | 1 | 1 | 1 | 1 | 1 | 1 | 6 |
| Gunasekera, 2007 [41] | 1 | 1 | 1 | 1 | 1 | 1 | 6 |
| Ivanov, 2007 [32] | 1 | 1 | 0 | 1 | 1 | 0 | 4 |
| Linnewiel-Hermoni, 2015 [33] | 1 | 1 | 1 | 1 | 1 | 1 | 6 |
| Liu, 2006 [34] | 1 | 1 | 1 | 0 | 1 | 1 | 5 |
| Liu, 2008 [35] | 1 | 1 | 1 | 1 | 1 | 1 | 6 |
| Peternac, 2008 [36] | 1 | 1 | 1 | 1 | 1 | 1 | 6 |
| Rafi, 2013 [37] | 1 | 1 | 1 | 1 | 0 | 1 | 5 |
| Richards, 2003 [38] | 1 | 0 | 1 | 1 | 0 | 0 | 3 |
| Tang, 2005 [42] | 1 | 1 | 1 | 1 | 1 | 1 | 6 |
| Tang, 2011 [43] | 1 | 1 | 1 | 1 | 1 | 0 | 5 |
| Zhang, 2010 [39] | 1 | 1 | 0 | 1 | 0 | 0 | 3 |
| Cell Line | Source | Androgen Receptor Expression | Androgen Sensitivity (AS or AI) | Other Characteristics |
|---|---|---|---|---|
| LNCaP [52] | Human PCa left supraclavicular lymph node metastasis | AR+ | AS | Broad AR ligand-specificity; PTEN mutation [52] |
| C4-2 [53] | Human PCa (subline of LNCaP cells derived from prostate epithelial cells of mouse xenograft cultured from osteosarcoma of mouse xenograft) | AR+ † | AI | High EGFR expression (5–10-fold higher) when compared to parental LNCaP cell line |
| LAPC-4 [54,55] | Human PCa (subline of LNCaP cells derived from mouse xenograft) | AR+ | AS | |
| 22Rv1 [56] | Human PCa (derived from prostate epithelial cells of muse xenograft after castration-induced regression and relapse of the parental, androgen-dependent CWR22 xenograft) | AR+ | AS | Express AR splice variants, making this line potentially AI |
| PC-3 [57] | Human PCa bone metastasis | AR− † | AI | Low SRD5A activities; PTEN deletion [52] |
| DU145 [57] | Human PCa brain metastasis | AR- | AI | High IGF-I expression [58] |
| 6S [35] | Primary human PCa stromal cells | AR+ | AS | |
| DTE [59] | Rat PCa (Dunning-R3327 tumors) | AR+ | AS | |
| AT-3 [59] | Rat PCa (derived from castration selection of parental DTE Dunning-R3327 tumors) | AR- | AI | Low SRD5A activities |
| Order of AR expression * | LNCaP > LAPC-4 > ? 22Rv1 > ? C4-2 > PC-3 > DU145 | |||
| Order of lycopene uptake [34,40] * | LNCaP > PC-3 > DU145 > C4-2 | |||
References
- Negoita, S.; Feuer, E.J.; Mariotto, A.; Cronin, K.A.; Petkov, V.I.; Hussey, S.K.; Benard, V.; Henley, S.J.; Anderson, R.N.; Fedewa, S.; et al. Annual Report to the Nation on the Status of Cancer, part II: Recent changes in prostate cancer trends and disease characteristics. Cancer 2018, 124, 2801–2814. [Google Scholar] [CrossRef] [PubMed]
- Pezaro, C.; Woo, H.H.; Davis, I.D. Prostate cancer: Measuring PSA. Intern. Med. J. 2014, 44, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Huggins, C.; Stevens, R.E., Jr.; Hodges, C.V. Studies on prostatic cancer: II. the effects of castration on advanced carcinoma of the prostate gland. Arch. Surg. 1941, 43, 209–223. [Google Scholar] [CrossRef]
- Tilki, D.; Evans, C.P. The changing landscape of advanced and castration resistant prostate cancer: Latest science and revised definitions. Can. J. Urol. 2014, 21, 7–13. [Google Scholar] [PubMed]
- Rowles, J.L., 3rd; Ranard, K.M.; Applegate, C.C.; Jeon, S.; An, R.; Erdman, J.W., Jr. Processed and raw tomato consumption and risk of prostate cancer: A systematic review and dose-response meta-analysis. Prostate Cancer Prostatic. Dis. 2018, 21, 319–336. [Google Scholar] [CrossRef] [PubMed]
- Rowles, J.L., 3rd; Ranard, K.M.; Smith, J.W.; An, R.; Erdman, J.W., Jr. Increased dietary and circulating lycopene are associated with reduced prostate cancer risk: A systematic review and meta-analysis. Prostate Cancer Prostatic. Dis. 2017, 20, 361–377. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, J.; Wang, X.; Wang, S.; Meng, S.; Zhu, Y.; Liang, Z.; Zheng, X.; Xie, L. Tomato consumption and prostate cancer risk: A systematic review and meta-analysis. Sci. Rep. 2016, 6, 37091. [Google Scholar] [CrossRef] [PubMed]
- Zu, K.; Mucci, L.; Rosner, B.A.; Clinton, S.K.; Loda, M.; Stampfer, M.J.; Giovannucci, E. Dietary lycopene, angiogenesis, and prostate cancer: A prospective study in the prostate-specific antigen era. J. Natl. Cancer Inst. 2014, 106, djt430. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cui, R.; Xiao, Y.; Fang, J.; Xu, Q. Effect of carotene and lycopene on the risk of prostate cancer: A systematic review and dose-response meta-analysis of observational studies. PLoS ONE 2015, 10, e0137427. [Google Scholar] [CrossRef]
- Paur, I.; Lilleby, W.; Bohn, S.K.; Hulander, E.; Klein, W.; Vlatkovic, L.; Axcrona, K.; Bolstad, N.; Bjoro, T.; Laake, P.; et al. Tomato-based randomized controlled trial in prostate cancer patients: Effect on PSA. Clin. Nutr. 2017, 36, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, Y.; Wang, Q. Lycopene can reduce prostate-specific antigen velocity in a phase II clinical study in Chinese population. Chin. Med. J. 2014, 127, 2143–2146. [Google Scholar] [PubMed]
- Wang, Y.; Jacobs, E.J.; Newton, C.C.; McCullough, M.L. Lycopene, tomato products and prostate cancer-specific mortality among men diagnosed with nonmetastatic prostate cancer in the Cancer Prevention Study II Nutrition Cohort. Int. J. Cancer 2016, 138, 2846–2855. [Google Scholar] [CrossRef] [PubMed]
- Key, T.J.; Appelby, P.N.; Travis, R.C.; Albanes, D.; Alberg, A.J.; Barricarte, A.; Black, A.; Boeing, H.; Bueno-de-Mesquita, H.B.; Chan, J.M.; et al. Carotenoids, retinol, tocopherols, and prostate cancer risk: A pooled analysis of 15 studies. Am. J. Clin. Nutr. 2015, 102, 1142–1157. [Google Scholar] [CrossRef] [PubMed]
- Schrecengost, R.; Knudsen, K.E. Molecular pathogenesis and progression of prostate cancer. Semin. Oncol. 2013, 40, 244–258. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Balk, S.P. Intratumoral androgen biosynthesis in prostate cancer pathogenesis and response to therapy. Endocr. Relat. Cancer 2011, 18, R175–R182. [Google Scholar] [CrossRef] [PubMed]
- Boileau, T.W.M.; Clinton, S.K.; Erdman, J.W., Jr. Tissue lycopene concentrations and isomer patterns are affected by androgen status and dietary lycopene concentration in male F344 rats. J. Nutr. 2000, 130, 1613–1618. [Google Scholar] [CrossRef] [PubMed]
- Boileau, T.W.M.; Clinton, S.K.; Zaripheh, S.; Monaco, M.H.; Donovan, S.M.; Erdman, J.W., Jr. Testosterone and food restriction modulate hepatic lycopene isomer concentrations in male F344 rats. J. Nutr. 2001, 131, 1746–1752. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.K.; Stroud, C.K.; Nakamura, M.T.; Lila, M.A.; Erdman, J.W., Jr. Serum testosterone is reduced following short-term phytofluene, lycopene, or tomato powder consumption in F344 rats. J. Nutr. 2006, 136, 2813–2819. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.-K.; Berk, M.; Chung, Y.-M.; Willard, B.; Bareja, R.; Rubin, M.; Sboner, A.; Sharifi, N. Loss of an androgen-inactivating and isoform-specific HSD17B4 splice form enables emergence of castration-resistant prostate cancer. Cell Rep. 2018, 22, 809–819. [Google Scholar] [CrossRef] [PubMed]
- Herzog, A.; Siler, U.; Spitzer, V.; Seifert, N.; Denelavas, A.; Hunziker, P.B.; Hunziker, W.; Goralczyk, R.; Wertz, K. Lycopene reduced gene expression of steroid targets and inflammatory markers in normal rat prostate. FASEB J. 2005, 19, 272–274. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Yuan, Y.; Vaishnav, A.; Tessel, M.A.; Nonn, L.; van Breemen, R.B. Effects of lycopene on protein expression in human primary prostatic epithelial cells. Cancer Prev. Res. 2013, 6, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.J.; Gardner, M.; Higgins, J.; Holly, J.M.P.; Gaunt, T.R.; Perks, C.M.; Turner, S.D.; Rinaldi, S.; Thomas, S.; Harrison, S.; et al. Developing the WCRF International/University of Bristol methodology for identifying and carrying out systematic reviews of mechanisms of exposure-cancer associations. Cancer Epidemiol. Biomark. Prev. 2017, 26, 1667–1675. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. J. Clin. Epidemiol. 2009, 62, 1006–1012. [Google Scholar] [CrossRef] [PubMed]
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.B.M.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.T.; Altman, D.G.; Gøtzsche, P.C.; Jüni, P.; Moher, D.; Oxman, A.D.; Savović, J.; Schulz, K.F.; Weeks, L.; Sterne, J.A.C. The Cochrane Collaboration’s tool for assessing risk of bias in randomised trials. BMJ 2011, 343, d5928. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.; Jones, H.E.; Martin, R.M.; Lewis, S.J.; Higgins, J.P.T. The albatross plot: A novel graphical tool for presenting results of diversely reported studies in a systematic review. Res. Synth. Methods 2017, 8, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Canene-Adams, K.; Lindshield, B.L.; Wang, S.; Jeffery, E.H.; Clinton, S.K.; Erdman, J.W., Jr. Combinations of tomato and broccoli enhance antitumor activity in dunning r3327-h prostate adenocarcinomas. Cancer Res. 2007, 67, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Limpens, J.; Schroder, F.H.; de Ridder, C.M.A.; Bolder, C.A.; Wildhagen, M.F.; Obermuller-Jevic, U.C.; Kramer, K.; van Weerden, W.M. Combined lycopene and vitamin E treatment suppresses the growth of PC-346C human prostate cancer cells in nude mice. J. Nutr. 2006, 136, 1287–1293. [Google Scholar] [CrossRef] [PubMed]
- Lindshield, B.L.; Ford, N.A.; Canene-Adams, K.; Diamond, A.M.; Wallig, M.A.; Erdman, J.W., Jr. Selenium, but not lycopene or vitamin E, decreases growth of transplantable dunning R3327-H rat prostate tumors. PLoS ONE 2010, 5, e10423. [Google Scholar] [CrossRef] [PubMed]
- Siler, U.; Barella, L.; Spitzer, V.; Schnorr, J.; Lein, M.; Goralczyk, R.; Wertz, K. Lycopene and Vitamin E interfere with autocrine/paracrine loops in the Dunning prostate cancer model. FASEB J. 2004, 18, 1019–1021. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.; Tan, H.L.; Thomas-Ahner, J.M.; Pearl, D.K.; Erdman, J.W., Jr.; Moran, N.E.; Clinton, S.K. Dietary tomato and lycopene impact androgen signaling and carcinogenesis-related gene expression during early TRAMP prostate carcinogenesis. Cancer Prev. Res. 2014, 7, 1228–1239. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, N.I.; Cowell, S.P.; Brown, P.; Rennie, P.S.; Guns, E.S.; Cox, M.E. Lycopene differentially induces quiescence and apoptosis in androgen-responsive and -independent prostate cancer cell lines. Clin. Nutr. 2007, 26, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Linnewiel-Hermoni, K.; Khanin, M.; Danilenko, M.; Zango, G.; Amosi, Y.; Levy, J.; Sharoni, Y. The anti-cancer effects of carotenoids and other phytonutrients resides in their combined activity. Arch. Biochem. Biophys. 2015, 572, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Pajkovic, N.; Pang, Y.; Zhu, D.; Calamini, B.; Mesecar, A.L.; van Breemen, R.B. Absorption and subcellular localization of lycopene in human prostate cancer cells. Mol. Cancer Ther. 2006, 5, 2879–2885. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.X.; Allen, J.D.; Arnold, J.T.; Blackman, M.R. Lycopene inhibits IGF-I signal transduction and growth in normal prostate epithelial cells by decreasing DHT-modulated IGF-I production in co-cultured reactive stromal cells. Carcinogenesis 2008, 29, 816–823. [Google Scholar] [CrossRef] [PubMed]
- Peternac, D.; Klima, I.; Cecchini, M.G.; Schwaninger, R.; Studer, U.E.; Thalmann, G.N. Agents used for chemoprevention of prostate cancer may influence PSA secretion independently of cell growth in the LNCaP model of human prostate cancer progression. Prostate 2008, 68, 1307–1318. [Google Scholar] [CrossRef] [PubMed]
- Rafi, M.M.; Kanakasabai, S.; Reyes, M.D.; Bright, J.J. Lycopene modulates growth and survival associated genes in prostate cancer. J. Nutr. Biochem. 2013, 24, 1724–1734. [Google Scholar] [CrossRef] [PubMed]
- Richards, L.T.R.; Benghuzzi, H.; Tucci, M.; Hughes, J. The synergistic effect of conventional and sustained delivery of antioxidants on LNCaP prostate cancer cell line. Biomed. Sci. Instrum. 2003, 39, 402–407. [Google Scholar] [PubMed]
- Zhang, X.; Wang, Q.; Neil, B.; Chen, X.A. Effect of lycopene on androgen receptor and prostate-specific antigen velocity. Chin. Med. J. 2010, 123, 2231–2236. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Marisiddaiah, R.; Zaripheh, S.; Wiener, D.; Rubin, L.P. Mitochondrial beta-carotene 9′,10′ oxygenase modulates prostate cancer growth via NF-kappaB inhibition: A lycopene-independent function. Mol. Cancer Res. 2016, 14, 966–975. [Google Scholar] [CrossRef] [PubMed]
- Gunasekera, R.S.; Sewgobind, K.; Desai, S.; Dunn, L.; Black, H.S.; McKeehan, W.L.; Patil, B. Lycopene and lutein inhibit proliferation in rat prostate carcinoma cells. Nutr. Cancer 2007, 58, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.L.; Jin, T.Y.; Zeng, X.B.; Wang, J.S. Lycopene inhibits the growth of human androgen-independent prostate cancer cells in vitro and in BALB/c nude mice. J. Nutr. 2005, 135, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Parmakhtiar, B.; Simoneau, A.R.; Xie, J.; Fruehauf, J.; Lilly, M.; Zi, X. Lycopene enhances docetaxel’s effect in castration-resistant prostate cancer associated with insulin-like growth factor I receptor levels. Neoplasia 2011, 13, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.J.; Ding, Y.B.; Wu, L.X.; Wen, C.J.; Qu, Q.; Zhang, X.; Zhou, H.H. The effects of lycopene on the methylation of the GSTP1 promoter and global methylation in prostatic cancer cell lines PC3 and LNCaP. Int. J. Endocrinol. 2014, 2014, 620125. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Huang, C.S.; Hu, M.L. The use of fetal bovine serum as delivery vehicle to improve the uptake and stability of lycopene in cell culture studies. Br. J. Nutr. 2007, 98, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.B.; Stanczyk, F.Z.; Wood, S.N.; Pfeiffer, R.M.; Hafi, M.; Veneroso, C.C.; Lynch, B.; Falk, R.T.; Zhou, C.K.; Niwa, S.; et al. Relationships between circulating and intraprostatic sex steroid hormone concentrations. Cancer Epidemiol. Biomark. Prev. 2017, 26, 1660–1666. [Google Scholar] [CrossRef] [PubMed]
- Izumi, K.; Shigehara, K.; Nohara, T.; Narimoto, K.; Kadono, Y.; Mizokami, A. Both high and low serum total testosterone levels indicate poor prognosis in patients with prostate cancer. Anticancer Res. 2017, 37, 5559–5564. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Chen, X.; Bird, V.Y.; Gerke, T.A.; Manini, T.M.; Prosperi, M. Association between age-related reductions in testosterone and risk of prostate cancer-an analysis of patients’ data with prostatic diseases. Int. J. Cancer 2017, 141, 1783–1793. [Google Scholar] [CrossRef] [PubMed]
- Shu, Q.; Cai, T.; Chen, X.; Zhu, H.H.; Xue, P.; Zhu, N.; Xie, Z.; Wei, S.; Zhang, Q.; Niu, L.; et al. Proteomic comparison and MRM-based comparative analysis of metabolites reveal metabolic shift in human prostate cancer cell lines. J. Proteome Res. 2015, 14, 3390–3402. [Google Scholar] [CrossRef] [PubMed]
- Jerónimo, C.; Bastian, P.J.; Bjartell, A.; Carbone, G.M.; Catto, J.W.F.; Clark, S.J.; Henrique, R.; Nelson, W.G.; Shariat, S.F. Epigenetics in prostate cancer: Biologic and clinical relevance. Eur. Urol. 2011, 60, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Tascilar, M.; Lee, W.-H.; Vles, W.J.; Lee, B.H.; Veeraswamy, R.; Asgari, K.; Freije, D.; van Rees, B.; Gage, W.R.; et al. GSTP1 CpG island hypermethylation is responsible for the absence of GSTP1 expression in human prostate cancer cells. Am. J. Pathol. 2001, 159, 1815–1826. [Google Scholar] [CrossRef]
- Russell, P.J.; Kingsley, E.A. Human prostate cancer cell lines. Methods Mol. Med. 2003, 81, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.C.; Hsieh, J.T.; Gleave, M.E.; Brown, N.M.; Pathak, S.; Chung, L.W. Derivation of androgen-independent human LNCaP prostatic cancer cell sublines: Role of bone stromal cells. Int. J. Cancer 1994, 57, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Klein, K.A.; Reiter, R.E.; Redula, J.; Moradi, H.; Zhu, X.L.; Brothman, A.R.; Lamb, D.J.; Marcelli, M.; Belldegrun, A.; Witte, O.N.; et al. Progression of metastatic human prostate cancer to androgen independence in immunodeficient SCID mice. Nat. Med. 1997, 3, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Hoefer, J.; Akbor, M.; Handle, F.; Ofer, P.; Puhr, M.; Parson, W.; Culig, Z.; Klocker, H.; Heidegger, I. Critical role of androgen receptor level in prostate cancer cell resistance to new generation antiandrogen enzalutamide. Oncotarget 2016, 7, 59781–59794. [Google Scholar] [CrossRef] [PubMed]
- Sramkoski, R.M.; Pretlow, T.G., II; Giaconia, J.M.; Pretlow, T.P.; Schwartz, S.; Sy, M.-S.; Marengo, S.R.; Rhim, J.S.; Zhang, D.; Jacobberger, J.W. A new human prostate carcinoma cell line, 22Rv1. In Vitro Cell Dev. Biol. Anim. 1999, 35, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, T.; Shigemura, K.; Yamamichi, F.; Fujisawa, M.; Kawabata, M.; Shirakawa, T. Detection of tumor markers in prostate cancer and comparison of sensitivity between real time and nested PCR. Kobe J. Med. Sci. 2012, 58, E51–E59. [Google Scholar] [PubMed]
- Pietrzkowski, Z.; Mulholland, G.; Gomella, L.; Jameson, B.A.; Wernicke, D.; Baserga, R. Inhibition of growth of prostatic cancer cell lines by peptide analogues of insulin-like growth factor 1. Cancer Res. 1993, 53, 1102–1106. [Google Scholar] [PubMed]
- Isaacs, J.T.; Isaacs, W.B.; Feitz, W.F.; Scheres, J. Establishment and characterization of seven dunning rat prostatic cancer cell lines and their use in developing methods for predicting metastatic abilities of prostatic cancers. Prostate 1986, 9, 261–281. [Google Scholar] [CrossRef] [PubMed]






| Author, Year | Animal Model | Baseline Diet(s) | Dietary Tomato and/or Lycopene Content * | Length of Intervention | Primary Findings |
|---|---|---|---|---|---|
| Canene-Adams, 2009 [27] | Dunning R3327-H transplantable tumors (Copenhagen rats) | AIN-93G-based diets fed; ad libitum | 10% TP (providing 13 nmol lycopene/g diet and resulting in 511 nmol/g serum lycopene), 23 nmol/g diet supplemental lycopene beadlet (252 nmol/g serum lycopene), or 224 nmol/g diet supplemental lycopene beadlet (884 nmol/g serum lycopene) | 18 weeks after tumor transplantation | No differences in serum testosterone or DHT between rats fed tomato, lycopene, or control diets |
| Limpens, 2006 [28] | Xenograft using PC-346C cells (athymic mice) | 821077 CRM(P) low (but adequate) vitamin E rodent diet; ad libitum | 5 or 50 mg/kg BW lycopene (from LycoVit) oral gavage = 0.5–1.5 mg lyco/day | 42 days after tumor inoculation | No differences in plasma PSA between mice given control or lycopene gavages (PSA levels were proportional to tumor size regardless of intervention) |
| Lindshield, 2010 [29] | Dunning R3327-H transplantable tumors (Copenhagen rats) | AIN-93G-based diets; ad libitum | 250 mg/kg diet supplemental lycopene beadlet = 5 mg lyco/day | 18 weeks after tumor transplantation | No differences in serum testosterone or DHT between rats fed lycopene or control diets |
| Siler, 2004 [30] | MayLyLu Dunning transplantable tumors (Copenhagen rats) | Kliba #2019 with added coconut fat (6%), <5 ppm vitamin E, reduced (but adequate) vitamin A, and devoid of phytosterols; did not indicate if ad libitum | 200 ppm lycopene (1.02 µM plasma lycopene) | 4 weeks on diet prior to tumor transplantation, then 18 additional days | Lycopene supplementation reduced tumor expression levels of SRD5A1 and androgen-target genes (cystatin related proteins 1 and 2; prostatic spermine binding protein; prostatic steroid-binding protein C1, C2, and C3; probasin) (only fold reductions reported—no other statistical values reported) |
| Wan, 2014 [31] | Transgenic (TRAMP mice) | AIN-93G -based diets; did not indicate if ad libitum | 10% TP (providing 384 mg lycopene/kg diet) or 462 mg/kg diet supplemental lycopene beadlet (0.36 µM plasma lycopene) = 3–4 mg lyco/day | 4 weeks diet prior to surgery, then 12 additional days | Reduced prostatic expression of genes related to androgen metabolism by tomato feeding (SRD5A2 (p = 0.04), Pxn (p = 0.04), and Srebf1 (p = 0.05)) and lycopene supplementation (SRD5A1, p = 0.03) |
| Author, Year | Cell Line(s) | Culture Conditions *† | Lycopene Dose(s) δ | Direct Androgen Outcomes: Primary Findings | Indirect Androgen Outcomes: Primary Findings | |
|---|---|---|---|---|---|---|
| Growth | Other | |||||
| Fu, 2014 [44] | LNCaP, PC-3 | RPMI1640, 10% FBS | 0, 5, 10, 20, 40 µM | 10 µM lycopene inhibited GSTP1 methylation (p < 0.05), increased GSTP1 gene expression (p < 0.05), and reduced DNMT3A gene expression (p < 0.01) in PC-3 but not LNCaP cells | ||
| Gong, 2016 [40] | LNCaP, C4-2, PC-3, DU145 | RPMI1640, 10% FBS | 1 µM | 1 µM lycopene inhibited growth of LNCaP (p < 0.05) but not in C4-2, PC-3, or DU145 cells | 1 µM lycopene induced BCO2 gene expression in LNCaP (p < 0.05) but not DU145 cells | |
| Gunasekera, 2007 [41] | AT3, DTE | RD (50% RPMI1640 + 50% DMEM), 2% FBS | 0.02, 0.2, 5, 10, 20 µM | 0.2 µM lycopene inhibited growth of AT3 (p < 0.0001) but not DTE cells | ||
| Ivanov, 2007 [32] | LNCaP, PC-3 | RPMI1640 or DMEM, 10% FBS | 0.01–10 µM (cell proliferation) 0.2, 0.4 µM (protein expression) 0–100 µM (androgen responsiveness) | Lycopene did not inhibit reporter activity of ARE-Luc transfected LNCaP cells at any concentration (no statistical values reported) | 0.2–0.8 µM lycopene inhibited growth of LNCaP and PC-3 cells (p < 0.05) | 0.2–0.8 µM lycopene inhibited Akt phosphorylation, cyclins D1 and E, and CDK2 in LNCaP and PC-3 cells (no statistical values reported) |
| Linnewiel-Hermoni, 2015 [33] | LNCaP, PC-3, DU145 | RPMI1640 or DMEM, 10% FCS, 10−9 DHT (for growth, stripped of steroid hormones prior to treatment) | 1–5 µM (cell proliferation) 8 µM (ARE-Luc) 2.5 µM (PSA) | 8 µM lycopene inhibited DHT-induced reporter activity of ARE-Luc transfected LNCaP cells (p < 0.01); non-significant decrease of DHT-induced PSA secretion by LNCaP cells treated with 2.5 µM lycopene | 1–5 µM lycopene inhibited DHT-induced growth of LNCaP cells (p < 0.01) | |
| Liu, 2006 [34] | LNCaP, PC-3, DU145 | RPMI1640 or Ham’s F12K or EMEM, 10% FBS | 1–1.48 µM | 1.48 µM lycopene did not directly bind to the AR (no statistical values reported) | Uptake is highest in LNCaP (p < 0.001) with 1.48 µM lycopene compared to PC-3 or DU145 cells | |
| Liu, 2008 [35] | 6S, 6S + NPE | DMEM, 5% FBS | 0.3, 1 µM | 0.3, 1 µM lycopene increased CM-mediated cell death and reduced IGF-I gene expression of 6S + NPE cells in the presence of DHT (p < 0.01); lycopene reduced DHT-induced total (p < 0.05) and nuclear (p < 0.01) AR protein expression in 6S cells | ||
| Peternac, 2008 [36] | LNCaP, C4-2 | T medium (80% DMEM + 20% Ham’s F12K) + 10% FCS | 0.04, 0.4, 4 µg/mL (equivalent to 0.075, 0.75, and 7.5 µM) | Lycopene did not reduce PSA gene or protein expression in LNCaP or C4-2 cells at any concentration | Lycopene inhibited growth of LNCaP (0.04, 0.4, 4 µg/mL) and C4-2 (0.4, 4 µg/mL) cells (p < 0.05) | |
| Rafi, 2013 [37] | PC-3 | RPMI1640, 10% FBS | 25 µM | 25 µM lycopene led to fold reductions of kallikrein peptidase family proteins gene expression in PC-3 cells (only fold reductions reported—no other statistical values reported) | ||
| Richards, 2003 [38] | LNCaP | Did not report cell culture conditions | 1, 10 µM | 1, 10 µM lycopene appeared to reduce PSA in LNCaP cells (no statistical values reported) | ||
| Tang, 2005 [42] | LNCaP, PC-3, DU145 | DMEM + Ham’s F12K, 10% FBS | Up to 50 µM | 10–50 µM lycopene more potently inhibited growth of PC-3 and DU145 cells (p < 0.01) compared to LNCaP cells | ||
| Tang, 2011 [43] | LNCaP, LAPC-4, PC-3, 22Rv1, DU145 | RPMI1640, 10% FBS | 1 µM | 1 µM lycopene appeared to reduce growth of all cell lines (no statistical values reported) | 1 µM lycopene more potently reduced Akt phosphorylation in DU145 (by 60%) than LNCaP (by 20%) cells (no statistical values reported) | |
| Zhang, 2010 [39] | LNCaP | RPMI1640, no other conditions reported | 0.5, 5, 10, 15 µM | 0.5–15 µM lycopene appeared to reduce reporter activity and ARE protein expression in ARE-Luc transfected LNCaP cells (no statistical values reported) | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Applegate, C.C.; Rowles, J.L., III; Erdman, J.W., Jr. Can Lycopene Impact the Androgen Axis in Prostate Cancer?: A Systematic Review of Cell Culture and Animal Studies. Nutrients 2019, 11, 633. https://doi.org/10.3390/nu11030633
Applegate CC, Rowles JL III, Erdman JW Jr. Can Lycopene Impact the Androgen Axis in Prostate Cancer?: A Systematic Review of Cell Culture and Animal Studies. Nutrients. 2019; 11(3):633. https://doi.org/10.3390/nu11030633
Chicago/Turabian StyleApplegate, Catherine C., Joe L. Rowles, III, and John W. Erdman, Jr. 2019. "Can Lycopene Impact the Androgen Axis in Prostate Cancer?: A Systematic Review of Cell Culture and Animal Studies" Nutrients 11, no. 3: 633. https://doi.org/10.3390/nu11030633
APA StyleApplegate, C. C., Rowles, J. L., III, & Erdman, J. W., Jr. (2019). Can Lycopene Impact the Androgen Axis in Prostate Cancer?: A Systematic Review of Cell Culture and Animal Studies. Nutrients, 11(3), 633. https://doi.org/10.3390/nu11030633