Comparative Nutrient Profiling of Retail Goat and Cow Milk

 , ,
, ,
Abstract

1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Milk Analysis
2.3. Statistical Analysis
2.4. Modelling of Nutrient Intakes by Consumers
3. Results
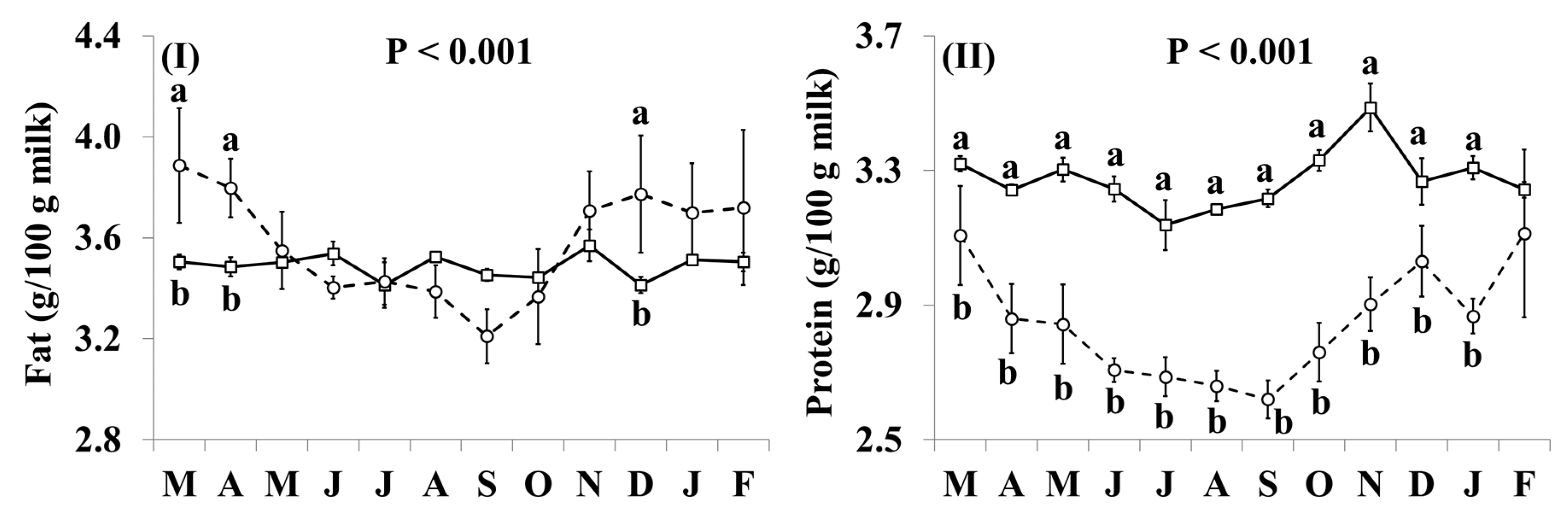
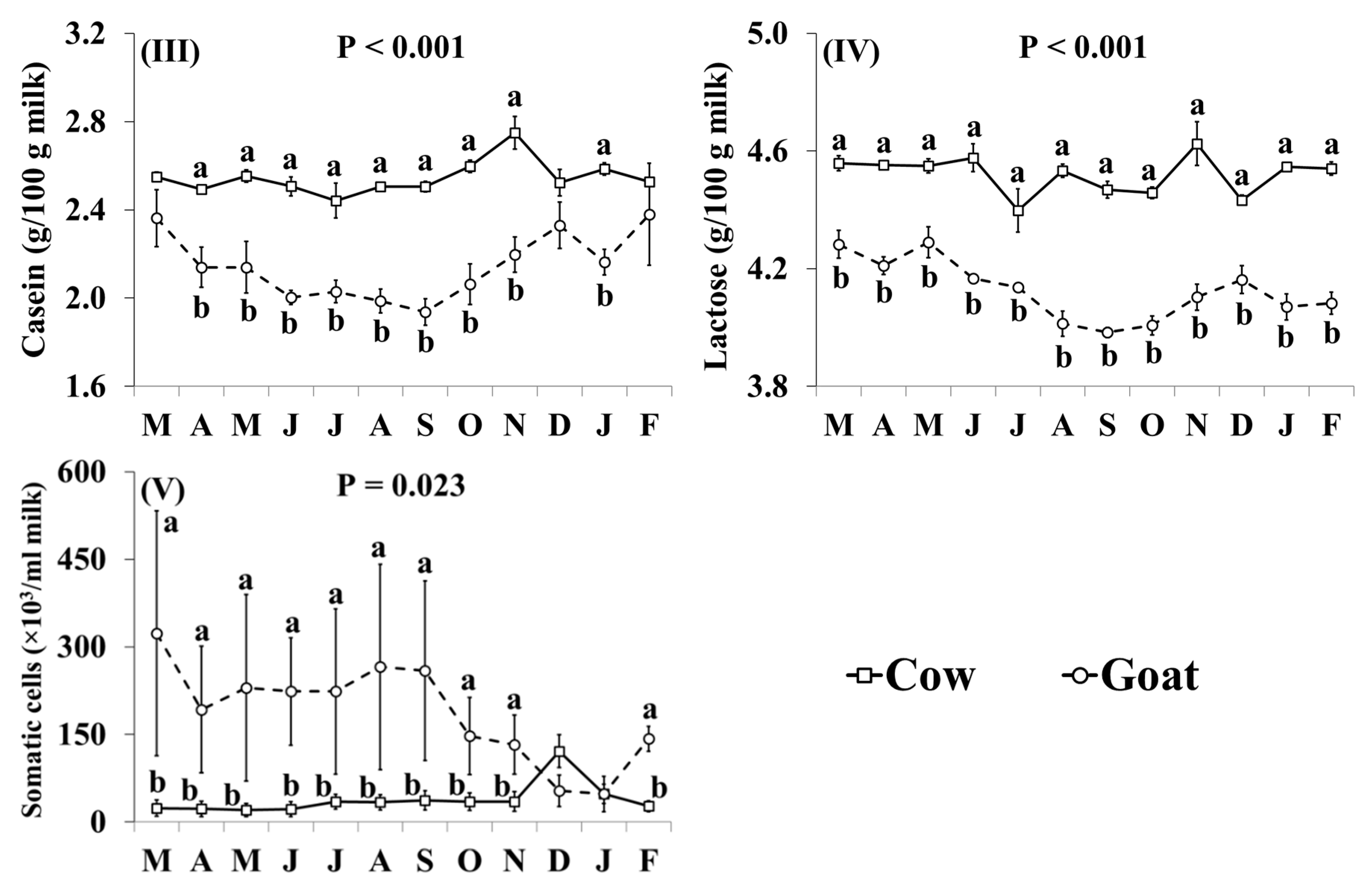
3.1. Basic Composition
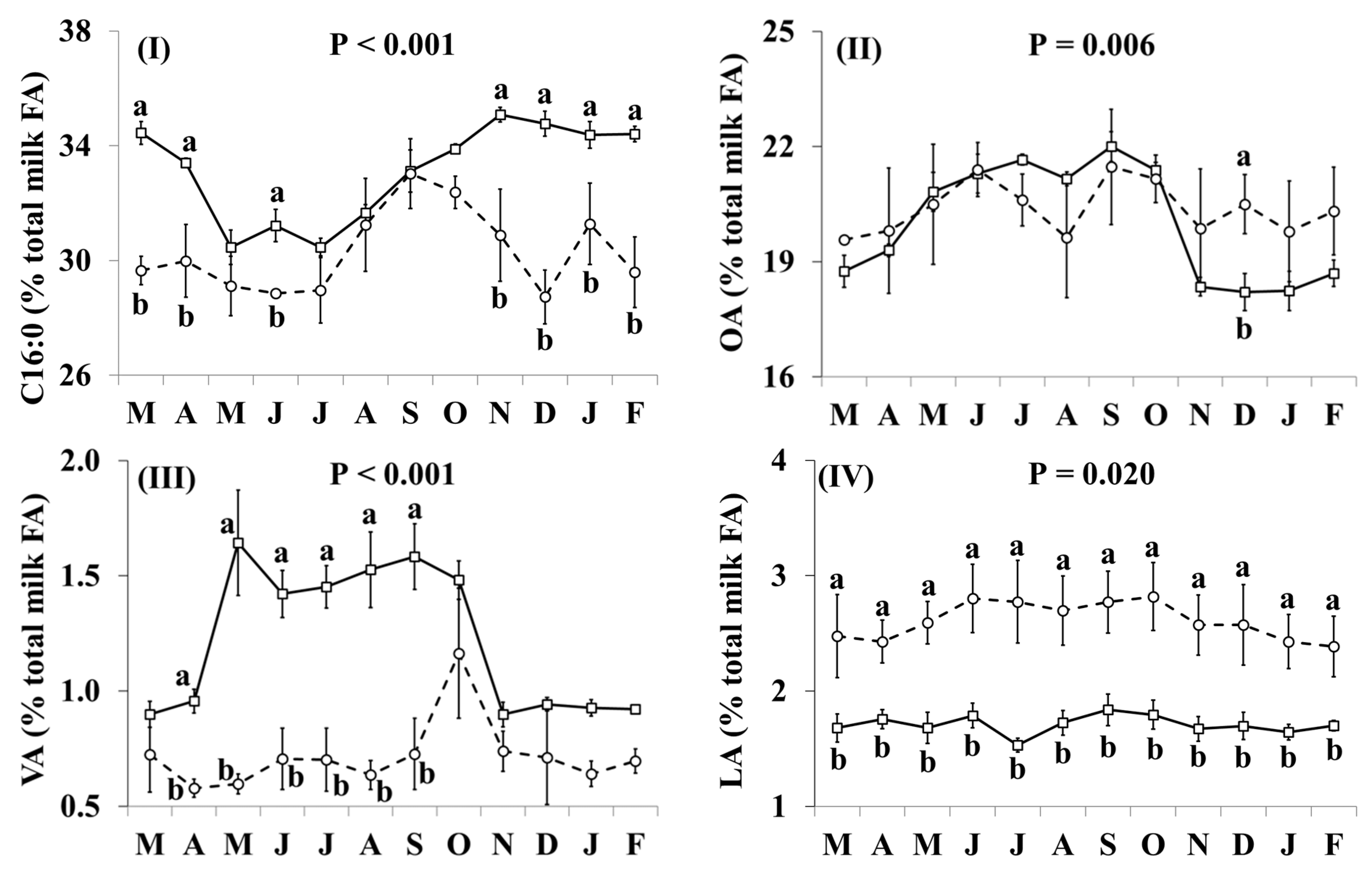
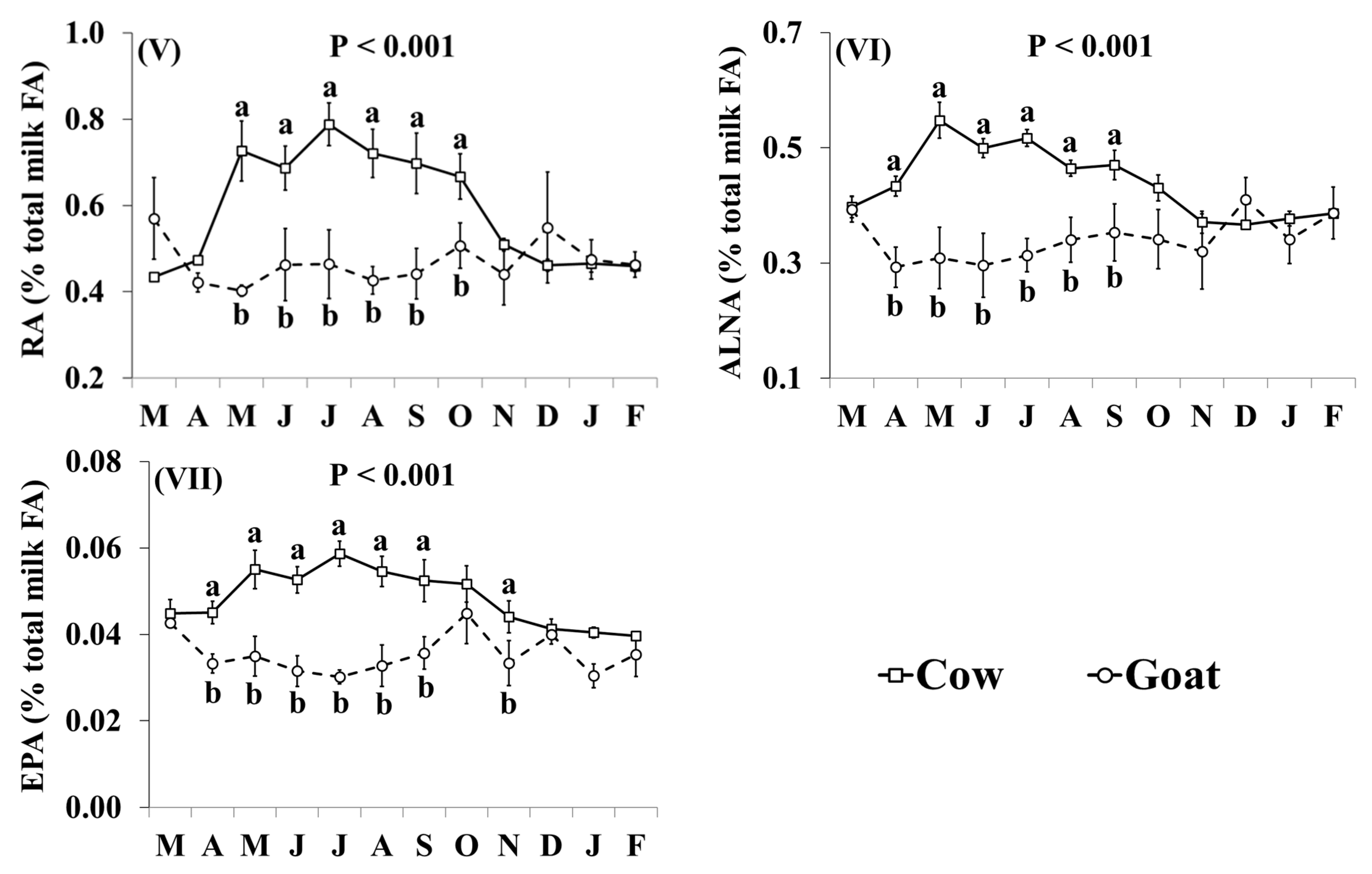
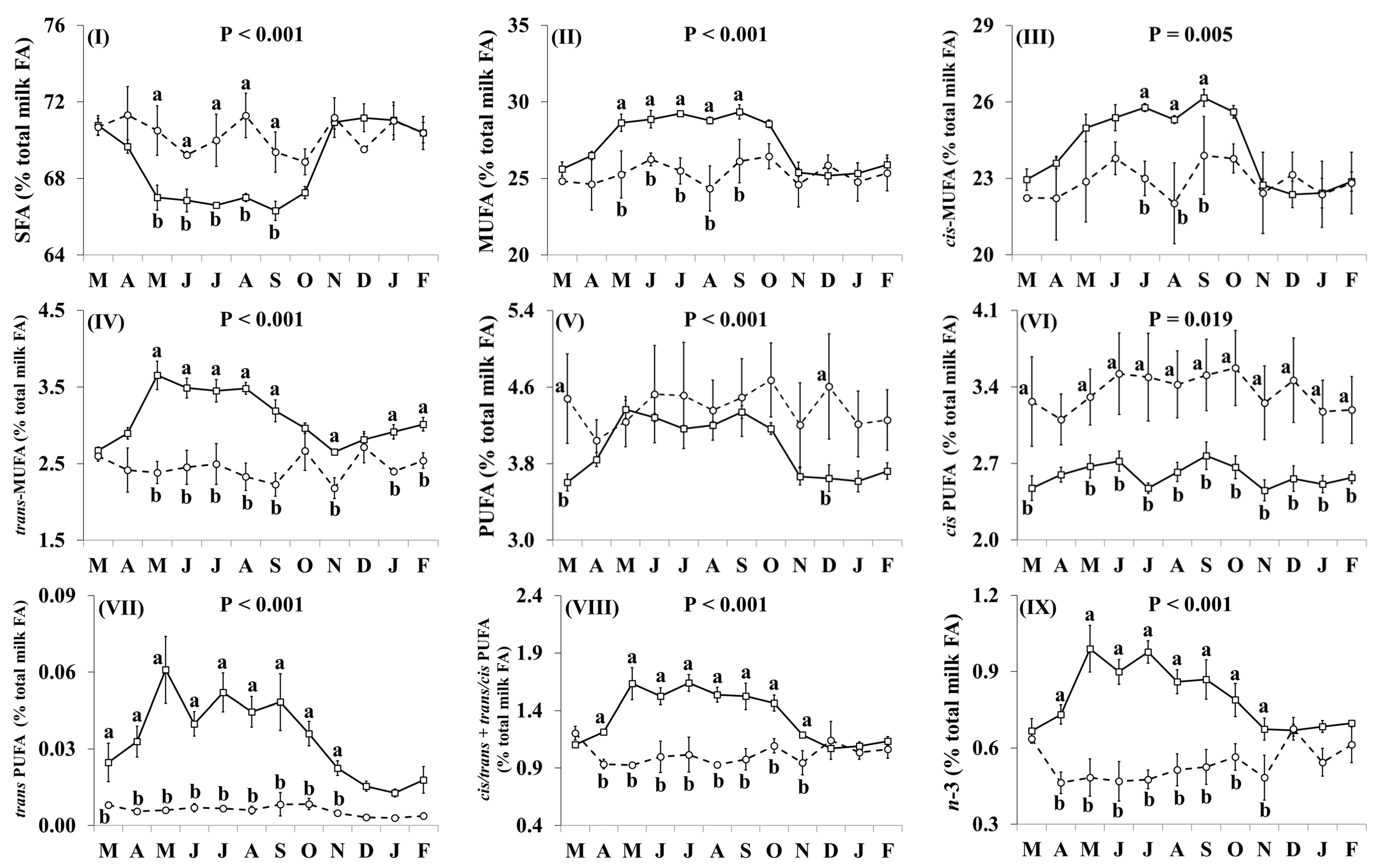
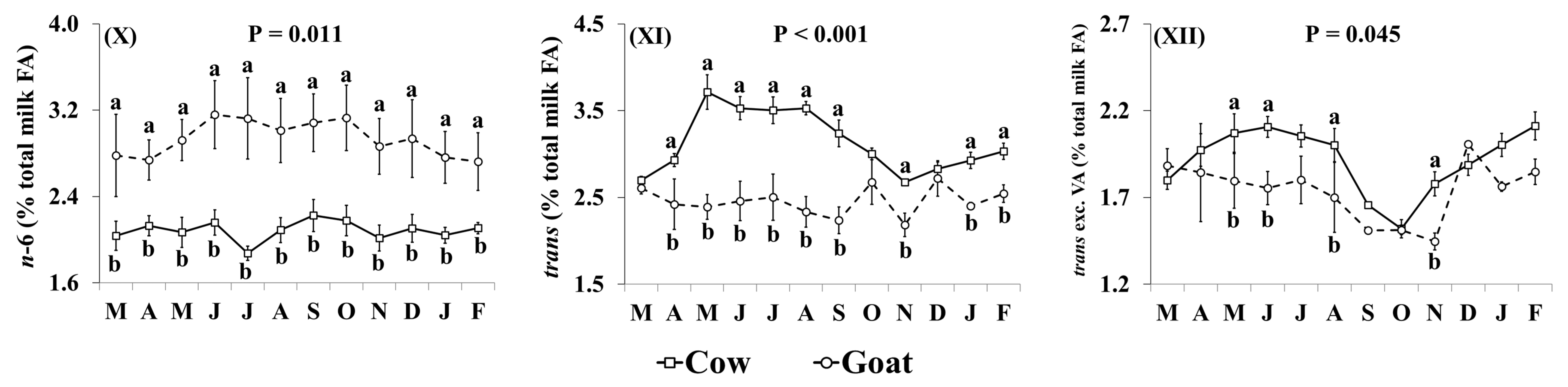
3.2. Fatty Acid Profile
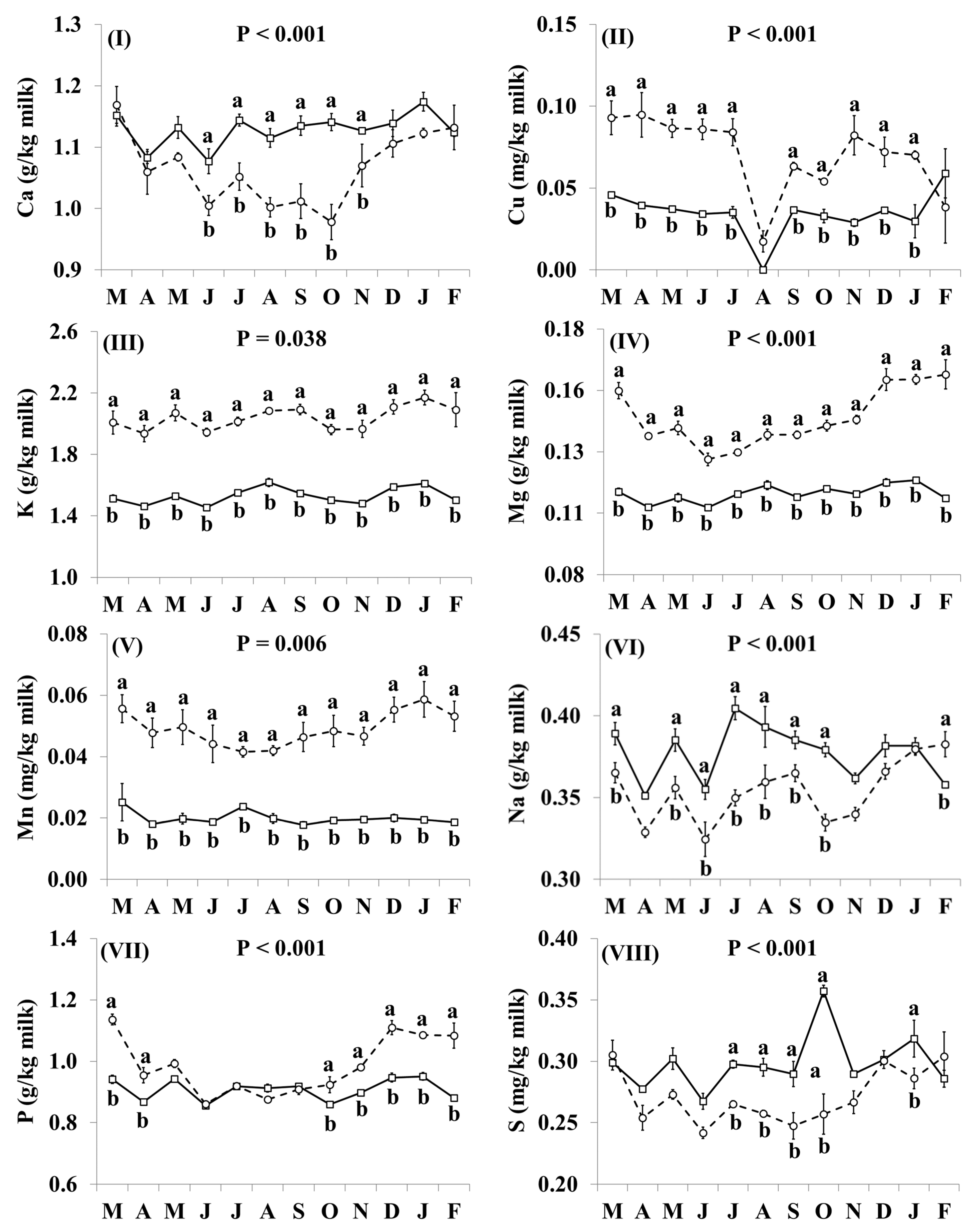
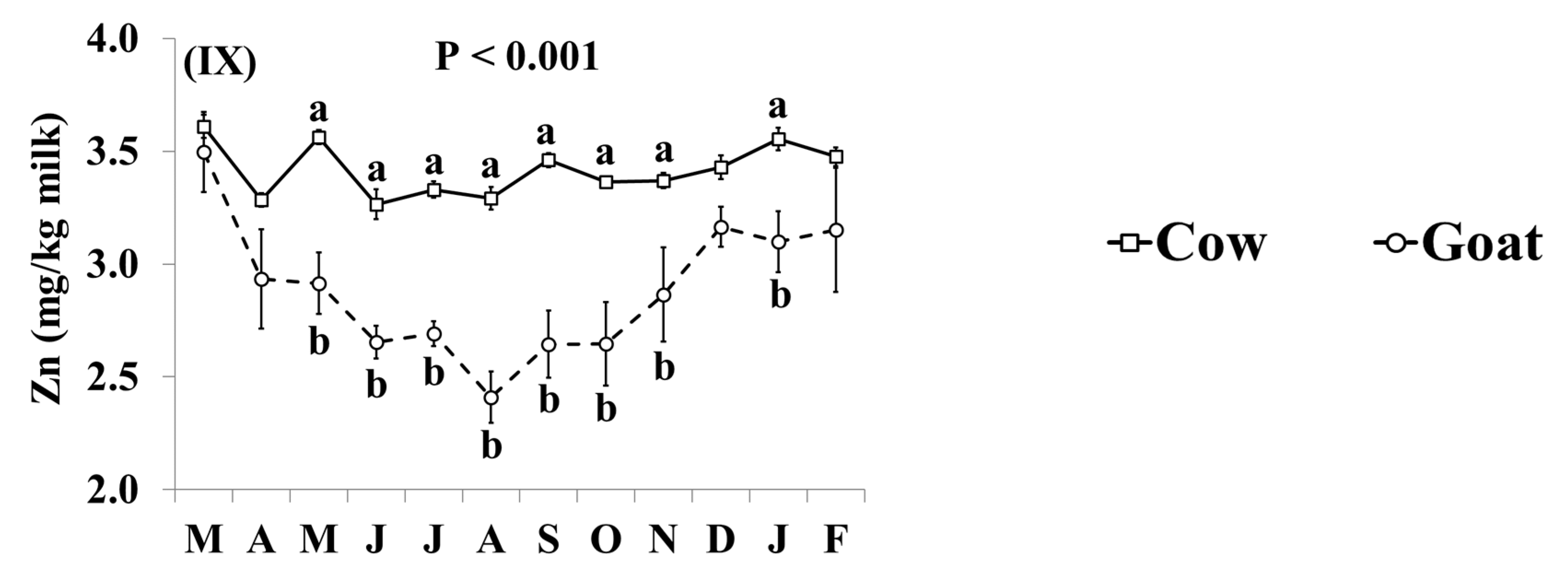
3.3. Mineral Concentrations
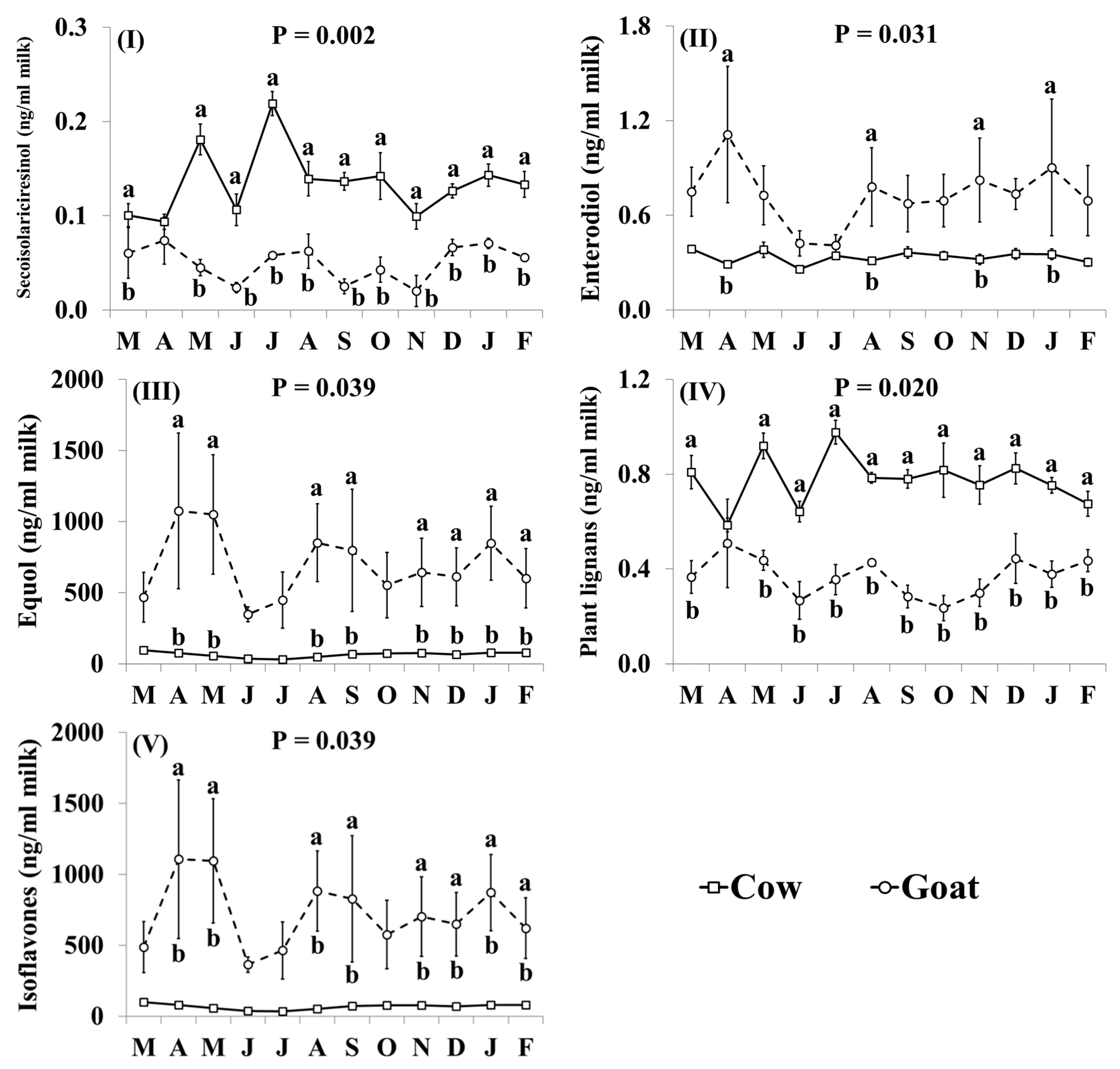
3.4. Phytoestrogen Concentrations
4. Discussion
4.1. Basic Composition of Retail Goat Milk
4.2. Fatty Acid Profile of Retail Goat Milk
4.3. Mineral Concentrations of Retail Goat Milk
4.4. Phytoestrogen Concentrations of Retail Goat Milk
4.5. Impact of Consuming Goat Milk on Nutrient Intakes of UK Consumers
4.5.1. Intakes of Fatty Acids
4.5.2. Intakes of Minerals
4.5.3. Intakes of Phytoestrogens
4.6. Strengths and Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Skapetas, B.; Bampidis, V. Goat Production in the World: Present Situation and Trends; Fundación CIPAV: Cali, Colombia, 2016; Volume 28, p. 200. [Google Scholar]
- Escareño, L.; Salinas-Gonzalez, H.; Wurzinger, M.; Iñiguez, L.; Sölkner, J.; Meza-Herrera, C. Dairy goat production systems: Status quo, perspectives and challenges. Trop. Anim. Health Prod. 2013, 45, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Milking Goat Association. Industry Statements: Herd Size and Welfare. Available online: https://www.milkinggoat.org.uk/herd-size-welfare (accessed on 18 June 2019).
- Haenlein, G.F.W. Goat milk in human nutrition. Small Rumin. Res. 2004, 51, 155–163. [Google Scholar] [CrossRef]
- Haenlein, G.F.W. Nutritional value of dairy products of ewe and goat milk. In Proceedings of the IDF/Greek National Commission IDF/Cerval Seminars, Crete, Greece, 19–21 October 1995; IDF Special Issue 9603. International Dairy Federation: Brussels, Belgium, 1996; pp. 159–178. [Google Scholar]
- Chilliard, Y.; Glasser, F.; Ferlay, A.; Bernard, L.; Rouel, J.; Doreau, M. Diet, rumen biohydrogenation and nutritional quality of cow and goat milk fat. Eur. J. Lipid Sci. Technol. 2007, 109, 828–855. [Google Scholar] [CrossRef]
- Stergiadis, S.; Berlitz, C.B.; Hunt, B.; Garg, S.; Ian Givens, D.; Kliem, K.E. An update to the fatty acid profiles of bovine retail milk in the United Kingdom: Implications for nutrition in different age and gender groups. Food Chem. 2019, 276, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Stergiadis, S.; Leifert, C.; Seal, C.J.; Eyre, M.D.; Nielsen, J.H.; Larsen, M.K.; Slots, T.; Steinshamn, H.; Butler, G. Effect of feeding intensity and milking system on nutritionally relevant milk components in dairy farming systems in the north east of England. J. Agric. Food Chem. 2012, 60, 7270–7281. [Google Scholar] [CrossRef]
- Stergiadis, S.; Seal, C.J.; Leifert, C.; Eyre, M.D.; Larsen, M.K.; Butler, G. Variation in nutritionally relevant components in retail Jersey and Guernsey whole milk. Food Chem. 2013, 139, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Currò, S.; De Marchi, M.; Claps, S.; Salzano, A.; De Palo, P.; Manuelian, C.L.; Neglia, G. Differences in the Detailed Milk Mineral Composition of Italian Local and Saanen Goat Breeds. Animals (Basel) 2019, 9, 412. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on Dietary Reference Values for fats, including saturated fatty acids, polyunsaturated fatty acids, monounsaturated fatty acids, trans fatty acids, and cholesterol. EFSA J. 2010, 8, 1461. [Google Scholar]
- FAO. Fats and Fatty Acids in Human Nutrition-Report of an Expert Consultation; Food and Agriculture Organization of the United Nations: Rome, Italy, 2010. [Google Scholar]
- Barcelo-Goblijn, G.; Murphy, E.J. Alpha-linolenic acid and its conversion to longer chain n-3 fatty acids: Benefits for human health and a role in maintaining tissue n-3 fatty acid levels. Prog. Lipid Res. 2009, 48, 355–374. [Google Scholar] [CrossRef]
- Dilzer, A.; Park, Y. Implication of Conjugated Linoleic Acid (CLA) in Human Health. Crit. Rev. Food Sci. Nutr. 2012, 52, 488–513. [Google Scholar] [CrossRef]
- Field, C.J.; Blewett, H.H.; Proctor, S.; Vine, D. Human health benefits of vaccenic acid. Appl. Physiol. Nutr. Metab. 2009, 34, 979–991. [Google Scholar] [CrossRef] [PubMed]
- Haug, A.; Hostmark, A.T.; Harstad, O.M. Bovine milk in human nutrition. Lipids Health Dis. 2007, 6, 25. [Google Scholar] [CrossRef] [PubMed]
- Swanson, D.; Block, R.; Mousa, S. Omega-3 fatty acids EPA and DHA: Health benefits throughout life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Strain, J.J.; Cashman, K.D. Minerals and trace elements. In Introduction to Human Nutrition, 2nd ed.; Gibney, M.J., Lanham-New, S.A., Cassidy, A., Vorster, H.H., Eds.; Wiley-Blackwell: Chichester, UK, 2009; pp. 188–237. [Google Scholar]
- Gaucheron, F. The minerals of milk. Reprod. Nutr. Dev. 2005, 45, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Dunshea, F.; Walker, G.; Williams, R.; Doyle, P. Mineral and citrate concentrations in milk are affected by seasons, stage of lactation and management practices. Agriculture 2019, 9, 25. [Google Scholar] [CrossRef]
- Park, Y.W.; Juárez, M.; Ramos, M.; Haenlein, G.F.W. Physico-chemical characteristics of goat and sheep milk. Small Rumin. Res. 2007, 68, 88–113. [Google Scholar] [CrossRef]
- Adlercreutz, H. Lignans and human health. Crit. Rev. Clin. Lab. Sci. 2007, 44, 483–525. [Google Scholar] [CrossRef]
- Fardet, A. New hypotheses for the health-protective mechanisms of whole-grain cereals: What is beyond fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef]
- Jungbauer, A.; Medjakovic, S. Phytoestrogens and the metabolic syndrome. J. Steroid Biochem. Mol. Biol. 2014, 139, 277–289. [Google Scholar] [CrossRef]
- Leitzmann, C. Characteristics and health benefits of phytochemicals. Forsch. Komplementarmed. 2016, 23, 69–74. [Google Scholar] [CrossRef]
- Wang, X.; Ouyang, Y.Y.; Liu, J.; Zhao, G. Flavonoid intake and risk of CVD: A systematic review and meta-analysis of prospective cohort studies. Br. J. Nutr. 2014, 111, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Turkmen, N. Chapter 35-The nutritional value and health benefits of goat milk components. In Nutrients in Dairy and Their Implications on Health and Disease; Watson, R.R., Collier, R.J., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 441–449. [Google Scholar] [CrossRef]
- McDonald, P.; Edwards, R.A.; Greenhalgh, J.F.D.; Morgan, C.A.; Sinclair, L.A.; Wilkinson, R.G. Animal Nutrition. 16. Lactation, 7th ed.; Pearson Education Limited: Harlow, UK, 2011; pp. 405–460. [Google Scholar]
- Stergiadis, S.; Bieber, A.; Chatzidimitriou, E.; Franceschin, E.; Isensee, A.; Rempelos, L.; Baranski, M.; Maurer, V.; Cozzi, G.; Bapst, B.; et al. Impact of US Brown Swiss genetics on milk quality from low-input herds in Switzerland: Interactions with season. Food Chem. 2018, 251, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, J.M.; Garnsworthy, P.C.; Huxley, J.N. Continuous Housing of Dairy Cows; British Society of Animal Science: Penicuik, UK, 2011; Available online: https://bsas.org.uk/articles/animal-briefs/continuous-housing-of-dairy-cows (accessed on 2 September 2019).
- Stergiadis, S.; Leifert, C.; Seal, C.J.; Eyre, M.D.; Larsen, M.K.; Slots, T.; Nielsen, J.H.; Butler, G. A 2-year study on milk quality from three pasture-based dairy systems of contrasting production intensities in Wales. J. Agric. Sci-Camb. 2015, 153, 708–731. [Google Scholar] [CrossRef]
- British Goat Society. Available online: https://www.britishgoatsociety.com/ (accessed on 14 June 2019).
- Anzuino, K.; Knowles, T.; Lee, M.R.F.; Grogono-Thomas, R. Survey of husbandry and health on UK commercial dairy goat farms. Vet. Rec. 2019, 185, 267. [Google Scholar] [CrossRef] [PubMed]
- Nørskov, N.P.; Givens, I.; Purup, S.; Stergiadis, S. Concentrations of phytoestrogens in conventional, organic and free-range retail milk in England. Food Chem. 2019, 295, 1–9. [Google Scholar] [CrossRef]
- Gilmour, A.R.; Thompson, R.; Cullis, B.R. Average information REML: An efficient algorithm for variance parameter estimation in linear-mixed models. Biometrics 1995, 51, 1440–1450. [Google Scholar] [CrossRef]
- Kay, J.K.; Mackle, T.R.; Auldist, M.J.; Thomson, N.A.; Bauman, D.E. Endogenous synthesis of cis-9, trans-11 conjugated linoleic acid in dairy cows fed fresh pasture. J. Dairy Sci. 2004, 87, 369–378. [Google Scholar] [CrossRef]
- Bates, B.; Lennox, A.; Prentice, A.; Bates, C.; Page, P.; Nicholson, S.; Swan, G. National Diet and Nutrition Survey. Results from Years 1-4 (Combined) of the Rolling Programme (2008/09–2011/12) (Revised February 2017); Public Health England: London, UK, 2014; p. 139. Available online: https://www.gov.uk/government/statistics/national-diet-and-nutrition-survey-results-from-years-1-to-4-combined-of-the-rolling-programme-for-2008-and-2009-to-2011-and-2012 (accessed on 22 September 2019).
- Glasser, F.; Doreau, M.; Ferlay, A.; Chilliard, Y. Technical note: Estimation of milk fatty acid yield from milk fat data. J. Dairy Sci. 2007, 90, 2302–2304. [Google Scholar] [CrossRef]
- Roberts, C.; Steer, T.; Maplethorpe, N.; Cox, L.; Meadows, S.; Nicholson, S.; Page, P.; Swan, G. National Diet and Nutrition Survey: Results from Year 7 and 8 (Combined) of the Rolling Programme (2014/2015 to 2015/2016); Public Health England: London, UK, 2018; p. 29. Available online: https://www.gov.uk/government/statistics/ndns-results-from-years-7-and-8-combined (accessed on 22 September 2019).
- SACN. Draft Report: Saturated Fats and Health; Scientific Advisory Committee in Nutrition: Crown Copyright: London, UK, 2018. [Google Scholar]
- AFSSA. Opinion of the French Food Safety Agency on the Update of French Population Reference Intakes (ANCs) for Fatty Acids; Food Safety Agency: Maisons-Alfort, France, 2010. [Google Scholar]
- SACN. Dietary Reference Values for Energy; Scientific Advisory Committee in Nutrition: Crown Copyright: London, UK, 2011. [Google Scholar]
- Srednicka-Tober, D.; Baranski, M.; Seal, C.; Sanderson, R.; Benbrook, C.; Steinshamn, H.; Gromadzka-Ostrowska, J.; Rembialkowska, E.; Skwarlo-Sonta, K.; Eyre, M.; et al. Composition differences between organic and conventional meat: A systematic literature review and meta-analysis. Br. J. Nutr. 2016, 115, 994–1011. [Google Scholar] [CrossRef]
- Toral, P.G.; Chilliard, Y.; Rouel, J.; Leskinen, H.; Shingfield, K.J.; Bernard, L. Comparison of the nutritional regulation of milk fat secretion and composition in cows and goats. J. Dairy Sci. 2015, 98, 7277–7297. [Google Scholar] [CrossRef]
- Fougere, H.; Delavaud, C.; Bernard, L. Diets supplemented with starch and corn oil, marine algae, or hydrogenated palm oil differentially modulate milk fat secretion and composition in cows and goats: A comparative study. J. Dairy Sci. 2018, 101, 8429–8445. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, L.S.; Morales, E.R.; de la Torre Adarve, G.; Castro, J.D.; Martínez, L.P.; Sampelayo, M.R.S. Composition of goat and cow milk produced under similar conditions and analyzed by identical methodology. J. Food Compos. Anal. 2009, 22, 322–329. [Google Scholar] [CrossRef]
- Amills, M.; Jordana, J.; Zidi, A.; Seradilla, J.M. Genetic factors that regulate milk protein and lipid composition in goats. In Milk Production-Advanced Genetic Traits, Cellular Mechanism, Animal Management and Health; Chaiyabutr, N., Ed.; IntechOpen: London, UK, 2012; p. 29. [Google Scholar]
- Costa, A.; Lopez-Villalobos, N.; Sneddon, N.W.; Shalloo, L.; Franzoi, M.; De Marchi, M.; Penasa, M. Invited review: Milk lactose—Current status and future challenges in dairy cattle. J. Dairy Sci. 2019, 102, 5883–5898. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zheng, N.; Wang, J.; Yang, Y. Comparative milk fatty acid analysis of different dairy species. Dairy Technol. 2017, 71, 544–550. [Google Scholar] [CrossRef]
- Póti, P.; Pajor, F.; Bodnár, Á.; Penksza, K.; Köles, P. Effect of micro-alga supplementation on goat and cow milk fatty acid composition. Chil. J. Agric. Res. 2015, 75, 259–263. [Google Scholar] [CrossRef]
- Fougere, H.; Bernard, L. Effect of diets supplemented with starch and corn oil, marine algae, or hydrogenated palm oil on mammary lipogenic gene expression in cows and goats: A comparative study. J. Dairy Sci. 2019, 102, 768–779. [Google Scholar] [CrossRef] [PubMed]
- Bernard, L.; Toral, P.G.; Chilliard, Y. Comparison of mammary lipid metabolism in dairy cows and goats fed diets supplemented with starch, plant oil, or fish oil. J. Dairy Sci. 2017, 100, 9338–9351. [Google Scholar] [CrossRef]
- Dewhurst, R.; Shingfield, K.; Lee, M.R.F.; Scollan, N. Increasing the concentrations of beneficial polyunsaturated fatty acids in milk produced by dairy cows in high forage systems. Anim. Feed Sci. Technol. 2006, 131, 168–206. [Google Scholar] [CrossRef]
- Rymer, C.; Givens, D.I. Dietary strategies for increasing docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) concentrations in bovine milk: A review. Nutr. Abstr. Rev. Ser. B Livest. Feed. Feed. 2003, 73, 9R–25R. [Google Scholar]
- Holt, C.; Jenness, R. Interrelationships of constituents and partition of salts in milk samples from eight species. Comp. Biochem. Physiol. A Comp. Physiol. 1984, 77, 275–282. [Google Scholar] [CrossRef]
- Yadav, A.; Singh, J.; Kumar Yadav, S. Composition, nutritional and therapeutic values of goat milk: A review. Asian J. Dairy Food Res. 2016, 35, 96–102. [Google Scholar] [CrossRef]
- Jenness, R. Composition and characteristics of goat milk: Review 1968−1979. J. Dairy Sci. 1980, 63, 1605–1630. [Google Scholar] [CrossRef]
- Trancoso, I.M.; Trancoso, M.A.; Martins, A.P.L.; Roseiro, L.B. Chemical composition and mineral content of goat milk from four indigenous Portuguese breeds in relation to one foreign breed. Int. J. Dairy Technol. 2010, 63, 516–522. [Google Scholar] [CrossRef]
- Kondyli, E.; Katsiari, M.C.; Voutsinas, L.P. Variations of vitamin and mineral contents in raw goat milk of the indigenous Greek breed during lactation. Food Chem. 2007, 100, 226–230. [Google Scholar] [CrossRef]
- Simos, E.; Voutsinas, L.P.; Pappas, C.P. Composition of milk of native Greek goats in the region of metsovo. Small Rumin. Res. 1991, 4, 47–60. [Google Scholar] [CrossRef]
- Voutsinas, L.; Pappas, C.; Katsiari, M. The composition of Alpine goats’ milk during lactation in Greece. J. Dairy Res. 1990, 57, 41–51. [Google Scholar] [CrossRef]
- StrzaŁkowska, N.; Bagnicka, E.; Jóźwik, A.; Krzyzewski, J. Macro- and micro-elements’ concentration in goat milk during lactation. J. Elem. 2008, 80, 404–411. [Google Scholar]
- Juniper, D.T.; Phipps, R.H.; Jones, A.K.; Bertin, G. Selenium supplementation of lactating dairy cows: Effect on selenium concentration in blood, milk, urine, and feces. J. Dairy Sci. 2006, 89, 3544–3551. [Google Scholar] [CrossRef]
- Lee, M.R.F.; Fleming, H.; Cogan, T.; Hodgson, C.; Davies, D.R. Assessing the ability of silage lactic acid bacteria to incorporate and transform inorganic selenium within laboratory scale silos. Anim. Feed Sci. Technol. 2019, 253, 125–134. [Google Scholar] [CrossRef]
- Niero, G.; Franzoi, M.; Vigolo, V.; Penasa, M.; Cassandro, M.; Boselli, C.; Giangolini, G.; De Marchi, M. Validation of a gold standard method for iodine quantification in raw and processed milk, and its variation in different dairy species. J. Dairy Sci. 2019, 102, 4808–4815. [Google Scholar] [CrossRef]
- Givens, D.I. Review: Dairy foods, red meat and processed meat in the diet: Implications for health at key life stages. Anim. Int. J. Anim. Biosci. 2018, 12, 1709–1721. [Google Scholar] [CrossRef] [PubMed]
- Anke, M.; Gunn, M.; Partshefeld, M. The essentiality of arsenic for animals. In Trace Substances in Environmental Health; Hemphill, D.D., Ed.; University of Missouri: Columbia, MO, USA, 1977. [Google Scholar]
- Kasparovska, J.; Pecinkova, M.; Dadakova, K.; Krizova, L.; Hadrova, S.; Lexa, M.; Lochman, J.; Kasparovsky, T. Effects of isoflavone-enriched feed on the rumen microbiota in dairy cows. PLoS ONE 2016, 11, e0154642. [Google Scholar] [CrossRef]
- Gagnon, N.; Cortes, C.; da Silva, D.; Kazama, R.; Benchaar, C.; dos Santos, G.; Zeoula, L.; Petit, H.V. Ruminal metabolism of flaxseed (Linum usitatissimum) lignans to the mammalian lignan enterolactone and its concentration in ruminal fluid, plasma, urine and milk of dairy cows. Br. J. Nutr. 2009, 102, 1015–1023. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, H.; Viala, D.; Ollier, A.; Combeau, A.; Besle, J.M. Isoflavones in several clover species and in milk from goats fed clovers. Biofactors 2004, 22, 237–239. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Han, Z.K.; Zhu, W.Y. The metabolism of linseed lignans in rumen and its impact on ruminal metabolism in male goats. J. Anim. Feed Sci. 2009, 18, 51–60. [Google Scholar] [CrossRef][Green Version]
- Wang, L.Z.; Xu, Q.; Kong, F.L.; Yang, Y.D.; Wu, D.; Mishra, S.; Li, Y. Exploring the goat rumen microbiome from seven days to two years. PLoS ONE 2016, 11, e0154354. [Google Scholar] [CrossRef]
- Yao, W.; Zhu, W.Y.; Han, Z.K.; Akkermans, A.D.L.; Williams, B.; Tamminga, S. Analysis of rumen bacterial diversity of goat by denaturing gradient gel electrophoresis and 16S rDNA sequencing. Sci. Agric. Sin. 2004, 37, 1374–1378. [Google Scholar]
- Kalac, P. Fresh and ensiled forages as a source of estrogenic equol in bovine milk: A review. Czech J. Anim. Sci. 2013, 58, 296–303. [Google Scholar] [CrossRef]
- Steinshamn, H.; Purup, S.; Thuen, E.; Hansen-Moller, J. Effects of clover-grass silages and concentrate supplementation on the content of phytoestrogens in dairy cow milk. J. Dairy Sci. 2008, 91, 2715–2725. [Google Scholar] [CrossRef]
- Kasparovska, J.; Krizova, L.; Lochman, J.; Dadakova, K.; Kasparovsky, T. Soybean-derived isoflavone determination in rumen fluid and milk by LC-MS-(TOF). J. Chromatogr. Sci. 2016, 54, 997–1003. [Google Scholar] [CrossRef]
- Schogor, A.L.B.; Huws, S.A.; Santos, G.T.D.; Scollan, N.D.; Hauck, B.D.; Winters, A.L.; Kim, E.J.; Petit, H.V. Ruminal Prevotella spp. may play an important role in the conversion of plant lignans into human health beneficial antioxidants. PLoS ONE 2014, 9, e87949. [Google Scholar] [CrossRef]
- Stevenson, D.M.; Weimer, P.J. Dominance of Prevotella and low abundance of classical ruminal bacterial species in the bovine rumen revealed by relative quantification real-time PCR. Appl. Microbiol. Biotechnol. 2007, 75, 165–174. [Google Scholar] [CrossRef]
- Simopoulos, A. The importance of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Kabu, M.; Uyarlar, C.; Żarczyńska, K.; Milewska, W.; Sobiech, P. The role of boron in animal health. J. Elem. 2015, 20, 535–541. [Google Scholar]
- Bath, S.C.; Steer, C.D.; Golding, J.; Emmett, P.; Rayman, M.P. Effect of inadequate iodine status in UK pregnant women on cognitive outcomes in their children: Results from the Avon Longitudinal Study of Parents and Children (ALSPAC). Lancet (London England) 2013, 382, 331–337. [Google Scholar] [CrossRef]
- Andersson, M.; de Benoist, B.; Darnton-Hill, I.; Delange, F. Iodine Deficiency in Europe: A Continuing Public Health Problem; World Health Organization (Published Jointly with UNICEF): Geneva, Switzerland, 2007. [Google Scholar]
- van der Reijden, O.L.; Zimmermann, M.B.; Galetti, V. Iodine in dairy milk: Sources, concentrations and importance to human health. Best Pract. Res. Clin. Endocrinol. Metab. 2017, 31, 385–395. [Google Scholar] [CrossRef]
- SFC. Opinion of the Scientific Committee on Food on the Tolerable Upper Intake Level of Iodine; SFC: Brussels, Belgium, 2002; p. 25. [Google Scholar]
- Perez, V.; Chang, E.T. Sodium-to-potassium ratio and blood pressure, hypertension, and related Factors. Adv. Nutr. 2014, 5, 712–741. [Google Scholar] [CrossRef]
- Meshref, A.M.S.; Moselhy, W.A.; Hassan, N.E.-H.Y. Heavy metals and trace elements levels in milk and milk products. J. Food Meas. Charact. 2014, 8, 381–388. [Google Scholar] [CrossRef]
- Iino, C.; Shimoyama, T.; Iino, K.; Yokoyama, Y.; Chinda, D.; Sakuraba, H.; Fukuda, S.; Nakaji, S. Daidzein intake is associated with equol producing status through an increase in the intestinal bacteria responsible for equol production. Nutrients 2019, 11, 433. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cow | Goat | ANOVA | ||
|---|---|---|---|---|
| Milk Solids | n = 48 | n = 36 | SE | p-Values a |
| Fat (g/100g milk) | 3.49 | 3.58 | 0.033 | ns |
| Protein (g/100g milk) | 3.27 | 2.85 | 0.027 | ** |
| Casein (g/100g milk) | 2.55 | 2.14 | 0.025 | ** |
| Lactose (g/100g milk) | 4.52 | 4.13 | 0.016 | *** |
| SCC (somatic cell count) b (×103/mL milk) | 38 | 187 | 18.7 | † |
| Cow | Goat | ANOVA | ||
|---|---|---|---|---|
| Individual FA and FA Groups | n = 48 | n = 36 | SE | p-Values a |
| SFA (% of total FA) b | ||||
| C12:0 | 3.33 | 4.16 | 0.079 | * |
| C14:0 | 11.1 | 10.3 | 0.09 | ** |
| C16:0 | 33.1 | 30.3 | 0.31 | * |
| C18:0 | 9.95 | 9.08 | 0.152 | ns |
| MUFA (% of total FA) c | ||||
| OA | 20.0 | 20.4 | 0.27 | ns |
| VA | 1.22 | 0.72 | 0.046 | ** |
| PUFA (% of total FA) d | ||||
| LA | 1.71 | 2.61 | 0.051 | * |
| RA | 0.591 | 0.469 | 0.0198 | † |
| ALNA | 0.439 | 0.342 | 0.0111 | † |
| EPA | 0.048 | 0.035 | 0.0012 | * |
| DPA | 0.079 | 0.075 | 0.0018 | ns |
| DHA | 0.007 | 0.015 | 0.0012 | ** |
| FA groups (% of total FA) | ||||
| SFA | 68.8 | 70.3 | 0.29 | * |
| MUFA | 27.3 | 25.3 | 0.29 | † |
| cis MUFA e | 24.2 | 22.9 | 0.27 | ns |
| trans MUFA f | 3.10 | 2.45 | 0.054 | *** |
| PUFA | 3.97 | 4.38 | 0.075 | ns |
| cis PUFA g | 2.59 | 3.36 | 0.056 | * |
| trans PUFA h | 0.034 | 0.006 | 0.0017 | ** |
| cis/trans + trans/cis PUFA i | 1.34 | 1.02 | 0.032 | ** |
| n-3 j | 0.792 | 0.538 | 0.0199 | * |
| n-6 k | 2.09 | 2.94 | 0.053 | * |
| trans FA l | 3.13 | 2.46 | 0.055 | *** |
| trans FA (exc. VA) | 1.91 | 1.74 | 0.036 | † |
| Indices | ||||
| Human health-related | ||||
| AI m | 2.60 | 2.56 | 0.044 | ns |
| TI n | 3.13 | 3.13 | 0.042 | ns |
| n-3/n-6 | 0.388 | 0.185 | 0.0105 | ** |
| Δ9-desaturase activity | ||||
| Δ9I o | 0.297 | 0.304 | 0.0032 | ns |
| C14:1/C14:0 | 0.084 | 0.015 | 0.0005 | *** |
| C16:1/C16:0 | 0.058 | 0.035 | 0.0005 | *** |
| OA/C18:0 | 2.01 | 2.27 | 0.026 | † |
| RA/VA | 0.491 | 0.678 | 0.0124 | *** |
| Cow | Goat | ANOVA | ||
|---|---|---|---|---|
| Minerals | n = 47 | n = 36 | SE | p-Values a |
| As (μg/kg) | 0.249 | 0.232 | 0.0237 | ns |
| B (mg/kg) | 0.176 | 0.263 | 0.0290 | * |
| Ca (g/kg) | 1.128 | 1.066 | 0.0084 | ** |
| Cd (μg/kg) | 0.047 | 0.044 | 0.0088 | ns |
| Co (μg/kg) | 0.335 | 0.299 | 0.0339 | ns |
| Cu (mg/kg) | 0.035 | 0.070 | 0.0034 | *** |
| Fe (mg/kg) | 0.214 | 0.216 | 0.0168 | ns |
| I (mg/kg) | 0.363 | 0.673 | 0.0346 | * |
| K (g/kg) | 1.528 | 2.037 | 0.0133 | *** |
| Mg (g/kg) | 0.113 | 0.144 | 0.0013 | *** |
| Mn (mg/kg) | 0.020 | 0.049 | 0.0010 | *** |
| Mo (mg/kg) | 0.039 | 0.024 | 0.0023 | ns |
| Na (g/kg) | 0.377 | 0.354 | 0.0032 | ** |
| Ni (μg/kg) | 1.151 | 0.826 | 0.2838 | ns |
| P (g/kg) | 0.908 | 0.986 | 0.0108 | *** |
| Pb (μg/kg) | 0.583 | 0.374 | 0.1390 | ns |
| S (mg/kg) | 0.299 | 0.272 | 0.0040 | * |
| Se (mg/kg) | 0.016 | 0.017 | 0.0004 | ns |
| Zn (mg/kg) | 3.416 | 2.889 | 0.0413 | ** |
| Cow | Goat | ANOVA | ||
|---|---|---|---|---|
| Phytoestrogens (ng/mL) | n = 48 | n = 36 | SE | p-Values a |
| Plant lignans | ||||
| Secoisolariciresinol | 0.135 | 0.050 | 0.005 | *** |
| Matairesinol | 0.123 | 0.061 | 0.008 | ** |
| Lariciresinol | 0.343 | 0.180 | 0.010 | *** |
| Hydroxymatairesinol | 0.176 | 0.077 | 0.009 | *** |
| Sum of plant lignans | 0.777 | 0.369 | 0.023 | *** |
| Mammalian lignans | ||||
| Enterolactone | 61.8 | 20.0 | 1.18 | *** |
| Enterodiol | 0.334 | 0.726 | 0.037 | † |
| Sum mammalian lignans | 62.2 | 20.7 | 1.18 | *** |
| Sum of lignans | 63.0 | 21.1 | 1.19 | *** |
| Plant isoflavones | ||||
| Daidzein | 0.952 | 8.066 | 0.559 | * |
| Genistein | 0.833 | 9.350 | 1.154 | † |
| Glycitein | 2.07 | 5.52 | 0.139 | *** |
| Formononetin | 0.082 | 6.479 | 0.770 | ns |
| Naringenin | 0.173 | 0.493 | 0.029 | ** |
| Sum of plant isoflavones | 4.11 | 29.91 | 2.489 | † |
| Mammalian isoflavones | ||||
| Equol | 63.6 | 690.6 | 42.20 | * |
| Sum of isoflavones | 67.7 | 720.5 | 43.78 | * |
| Plant coumestans | ||||
| Coumestrol | 0.096 | 0.367 | 0.046 | ns |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stergiadis, S.; Nørskov, N.P.; Purup, S.; Givens, I.; Lee, M.R.F. Comparative Nutrient Profiling of Retail Goat and Cow Milk. Nutrients 2019, 11, 2282. https://doi.org/10.3390/nu11102282
Stergiadis S, Nørskov NP, Purup S, Givens I, Lee MRF. Comparative Nutrient Profiling of Retail Goat and Cow Milk. Nutrients. 2019; 11(10):2282. https://doi.org/10.3390/nu11102282
Chicago/Turabian StyleStergiadis, Sokratis, Natalja P. Nørskov, Stig Purup, Ian Givens, and Michael R. F. Lee. 2019. "Comparative Nutrient Profiling of Retail Goat and Cow Milk" Nutrients 11, no. 10: 2282. https://doi.org/10.3390/nu11102282
APA StyleStergiadis, S., Nørskov, N. P., Purup, S., Givens, I., & Lee, M. R. F. (2019). Comparative Nutrient Profiling of Retail Goat and Cow Milk. Nutrients, 11(10), 2282. https://doi.org/10.3390/nu11102282