The Potential Impact of Probiotics on the Gut Microbiome of Athletes


Abstract
1. A Brief Introduction to Exercise and the Microbiome
2. Traditional Probiotics
2.1. Escherichia Coli
2.2. Lactobacillus
2.3. Bifidobacterium
2.4. Saccharomyces

{kind=link}
{kind=link}
| Probiotic Genus * | Found in the Body | Dietary Source: | Potential Benefits Attributed to Specific Strains | References |
|---|---|---|---|---|
| Lactobacillus | Colon, gut and vagina | Yoghurt, fermented foods, bread, sauerkraut, wine etc. | Gastroenteritis, easing lactose intolerance, immune system modulation, alleviating inflammation, lowering cholesterol, cancer protection, modulating brain activity, preventing pathogen colonisation, bile resistant. | [60,61,62,68,82,83,84,85,86,87] |
| Bifidobacterium | Colon, oral cavity, breast milk and vagina | Yoghurt, kombucha, sauerkraut, kefir etc. | Bile resistant, easing lactose intolerance, antibiotic-associated diarrhoea, eczema, immune system modulation, cholesterol lowering abilities | [68,71,88,89,90] |
| Saccharomyces | Colon, decaying fruit, plants, soil, insects | Wine, yoghurt, kombucha, sauerkraut etc. | Travellers’ diarrhoea, antibiotic-associated diarrhoea, preventing recurring Clostridium difficile infections, irritable bowel syndrome | [77,78,79] |
| Escherichia coli | Colon | Capsules | Antagonistic properties against a variety of pathogens, production of defensin, tight-junction protein modification, irritable bowel disorder, constipation, pro-inflammatory properties and colon cancer | [50,51,52,53,54,91] |
| Bacteroidetes | Colon | - | Immune system modulation, intestinal homeostasis | [92,93,94] |
| Akkermansia | Colon | - | Gut barrier function, fat mass storage, glucose homeostasis, immune system stimulation, production of Vitamin B12 | [95,96,97,98,99,100,101] |
| Faecalibacterium | Colon | - | Immune system modulation, ease inflammation | [102,103] |
| Eubacterium | Colon | - | Improve insulin sensitivity, increase energy production, produce Vitamin B12, maintain intestinal homoestasis, colon detoxification, reducing the symptoms of colitis | [104,105,106,107] |
3. Next-Generation Probiotics
4. Designer Probiotics
5. Safety Considerations
6. Probiotics in Treating the Overtraining Syndrome
6.1. Antioxidant Boosting
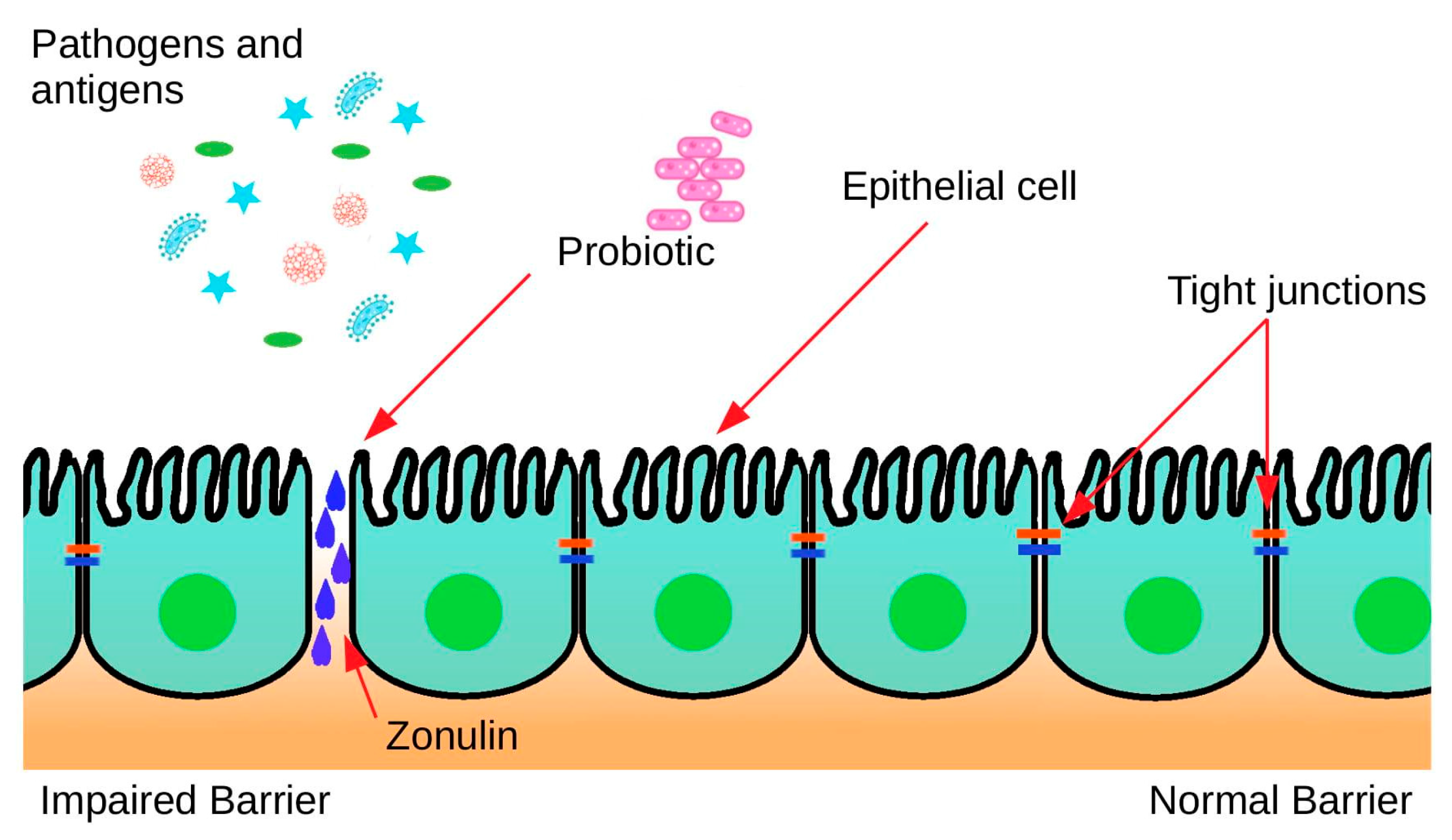
6.2. Tight-Junction Protein Modification
6.3. Immune System Modulation
6.4. Infection
6.5. Mental Health
7. Conclusion and Future Prospects
Funding
Conflicts of Interest
References
- Janssens, Y.; Nielandt, J.; Bronselaer, A.; Debunne, N.; Verbeke, F.; Wynendaele, E.; Van Immerseel, F.; Vandewynckel, Y.-P.; De Tré, G.; De Spiegeleer, B. Disbiome database: Linking the microbiome to disease. BMC Microbiol. 2018, 18, 50. [Google Scholar] [CrossRef]
- Gill, S.R.; Pop, M.; DeBoy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic analysis of the human distal gut microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef]
- Nash, A.K.; Auchtung, T.A.; Wong, M.C.; Smith, D.P.; Gesell, J.R.; Ross, M.C.; Stewart, C.J.; Metcalf, G.A.; Muzny, D.M.; Gibbs, R.A.; et al. The gut mycobiome of the Human Microbiome Project healthy cohort. Microbiome 2017, 5, 153. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef]
- O’Donovan, C.M.; O’Sullivan, O.; Cotter, P.D. Gut Microbiol—A Relatively Unexplored Domain; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Jandhyala, S.M. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787. [Google Scholar] [CrossRef] [PubMed]
- Manichanh, C.; Rigottier-Gois, L.; Bonnaud, E.; Gloux, K.; Pelletier, E.; Frangeul, L.; Nalin, R.; Jarrin, C.; Chardon, P.; Marteau, P. Reduced diversity of faecal microbiota in Crohn’s disease revealed by a metagenomic approach. Gut 2006, 55, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.C.; Perez-Chanona, E.; Mühlbauer, M.; Tomkovich, S.; Uronis, J.M.; Fan, T.-J.; Campbell, B.J.; Abujamel, T.; Dogan, B.; Rogers, A.B. Intestinal inflammation targets cancer-inducing activity of the microbiota. Science 2012, 338, 120–123. [Google Scholar] [CrossRef]
- De Goffau, M.C.; Luopajarvi, K.; Knip, M.; Ilonen, J.; Ruohtula, T.; Harkonen, T.; Orivuori, L.; Hakala, S.; Welling, G.W.; Harmsen, H.J.; et al. Fecal microbiota composition differs between children with beta-cell autoimmunity and those without. Diabetes 2013, 62, 1238–1244. [Google Scholar] [CrossRef]
- Mach, N.; Fuster-Botella, D. Endurance exercise and gut microbiota: A review. J. Sport Health Sci. 2017, 6, 179–197. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Shade, A. Diversity is the question, not the answer. ISME J. 2017, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Barton, W.; Penney, N.C.; Cronin, O.; Garcia-Perez, I.; Molloy, M.G.; Holmes, E.; Shanahan, F.; Cotter, P.D.; O’Sullivan, O. The microbiome of professional athletes differs from that of more sedentary subjects in composition and particularly at the functional metabolic level. Gut 2017, 67, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Clarke, S.F.; Murphy, E.F.; O’Sullivan, O.; Lucey, A.J.; Humphreys, M.; Hogan, A.; Hayes, P.; O’Reilly, M.; Jeffery, I.B.; Wood-Martin, R.; et al. Exercise and associated dietary extremes impact on gut microbial diversity. Gut 2014, 63, 1913–1920. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.M.; Bautista, E.J.; Nguyen, H.; Hanson, B.M.; Chen, L.; Lek, S.H.; Sodergren, E.; Weinstock, G.M. Community characteristics of the gut microbiomes of competitive cyclists. Microbiome 2017, 5, 98. [Google Scholar] [CrossRef] [PubMed]
- Cronin, O.; Barton, W.; Skuse, P.; Penney, N.C.; Garcia-Perez, I.; Murphy, E.F.; Woods, T.; Nugent, H.; Fanning, A.; Melgar, S.; et al. A Prospective Metagenomic and Metabolomic Analysis of the Impact of Exercise and/or Whey Protein Supplementation on the Gut Microbiome of Sedentary Adults. MSystems 2018, 3, e00044. [Google Scholar] [CrossRef] [PubMed]
- Jang, L.-G.; Choi, G.; Kim, S.-W.; Kim, B.-Y.; Lee, S.; Park, H. The combination of sport and sport-specific diet is associated with characteristics of gut microbiota: An observational study. J. Int. Soc. Sports Nutr. 2019, 16, 21. [Google Scholar] [CrossRef] [PubMed]
- Scheiman, J.; Luber, J.M.; Chavkin, T.A.; MacDonald, T.; Tung, A.; Pham, L.-D.; Wibowo, M.C.; Wurth, R.C.; Punthambaker, S.; Tierney, B.T. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nat. Med. 2019, 25, 1104–1109. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.D.; Koltyn, K.F.; Stegner, A.J.; Kim, J.-S.; Cook, D.B. Influence of exercise intensity for improving depressed mood in depression: A dose-response study. Behav. Ther. 2016, 47, 527–537. [Google Scholar] [CrossRef]
- Duscha, B.D.; Slentz, C.A.; Johnson, J.L.; Houmard, J.A.; Bensimhon, D.R.; Knetzger, K.J.; Kraus, W.E. Effects of exercise training amount and intensity on peak oxygen consumption in middle-age men and women at risk for cardiovascular disease. Chest 2005, 128, 2788–2793. [Google Scholar] [CrossRef]
- Puetz, T.W.; Flowers, S.S.; O’Connor, P.J. A randomized controlled trial of the effect of aerobic exercise training on feelings of energy and fatigue in sedentary young adults with persistent fatigue. Psychother. Psychosom. 2008, 77, 167–174. [Google Scholar] [CrossRef]
- Meeusen, R.; Duclos, M.; Foster, C.; Fry, A.; Gleeson, M.; Nieman, D.; Raglin, J.; Rietjens, G.; Steinacker, J.; Urhausen, A. Prevention, diagnosis and treatment of the overtraining syndrome: Joint consensus statement of the European College of Sport Science (ECSS) and the American College of Sports Medicine (ACSM). Eur. J. Sport Sci. 2013, 13, 1–24. [Google Scholar] [CrossRef]
- Allen, J.M.; Berg Miller, M.E.; Pence, B.D.; Whitlock, K.; Nehra, V.; Gaskins, H.R.; White, B.A.; Fryer, J.D.; Woods, J.A. Voluntary and forced exercise differentially alters the gut microbiome in C57BL/6J mice. J. Appl. Physiol. 2015, 118, 1059–1066. [Google Scholar] [CrossRef] [PubMed]
- Karl, J.P.; Margolis, L.M.; Madslien, E.H.; Murphy, N.E.; Castellani, J.W.; Gundersen, Y.; Hoke, A.V.; Levangie, M.W.; Kumar, R.; Chakraborty, N. Changes in intestinal microbiota composition and metabolism coincide with increased intestinal permeability in young adults under prolonged physiological stress. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G559–G571. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Xu, S.; Huang, H.; Liang, J.; Wu, Y.; Li, C.; Yuan, H.; Zhao, X.; Lai, X.; Hou, S. Influence of excessive exercise on immunity, metabolism, and gut microbial diversity in an overtraining mice model. Scand. J. Med. Sci. Sports 2018, 28, 1541–1551. [Google Scholar] [CrossRef] [PubMed]
- Walsh, N.P.; Gleeson, M.; Shephard, R.J.; Gleeson, M.; Woods, J.A.; Bishop, N.; Fleshner, M.; Green, C.; Pedersen, B.K.; Hoffman-Goete, L. Position Statement Part One: Immune Function and Exercise; Loughborough University: Loughborough, UK, 2011. [Google Scholar]
- Sina, C.; Gavrilova, O.; Förster, M.; Till, A.; Derer, S.; Hildebrand, F.; Raabe, B.; Chalaris, A.; Scheller, J.; Rehmann, A.G. Protein-coupled receptor 43 is essential for neutrophil recruitment during intestinal inflammation. J. Immunol. 2009, 183, 7514–7522. [Google Scholar] [CrossRef]
- Shouval, D.S.; Biswas, A.; Goettel, J.A.; McCann, K.; Conaway, E.; Redhu, N.S.; Mascanfroni, I.D.; Al Adham, Z.; Lavoie, S.; Ibourk, M. Interleukin-10 receptor signaling in innate immune cells regulates mucosal immune tolerance and anti-inflammatory macrophage function. Immunity 2014, 40, 706–719. [Google Scholar] [CrossRef] [PubMed]
- Sutterwala, F.S.; Noel, G.J.; Clynes, R.; Mosser, D.M. Selective suppression of interleukin-12 induction after macrophage receptor ligation. J. Exp. Med. 1997, 185, 1977–1985. [Google Scholar] [CrossRef]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef]
- Valles-Colomer, M.; Falony, G.; Darzi, Y.; Tigchelaar, E.F.; Wang, J.; Tito, R.Y.; Schiweck, C.; Kurilshikov, A.; Joossens, M.; Wijmenga, C. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat. Microbiol. 2019, 4, 623. [Google Scholar] [CrossRef]
- Naseribafrouei, A.; Hestad, K.; Avershina, E.; Sekelja, M.; Linløkken, A.; Wilson, R.; Rudi, K. Correlation between the human fecal microbiota and depression. Neurogastroenterol. Motil. 2014, 26, 1155–1162. [Google Scholar] [CrossRef]
- Maes, M.; Meltzer, H.Y.; Bosmans, E.; Bergmans, R.; Vandoolaeghe, E.; Ranjan, R.; Desnyder, R. Increased plasma concentrations of interleukin-6, soluble interleukin-6, soluble interleukin-2 and transferrin receptor in major depression. J. Affect. Disord. 1995, 34, 301–309. [Google Scholar] [CrossRef]
- Margonis, K.; Fatouros, I.G.; Jamurtas, A.Z.; Nikolaidis, M.G.; Douroudos, I.; Chatzinikolaou, A.; Mitrakou, A.; Mastorakos, G.; Papassotiriou, I.; Taxildaris, K. Oxidative stress biomarkers responses to physical overtraining: Implications for diagnosis. Free Radic. Biol. Med. 2007, 43, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Tanskanen, M.; Atalay, M.; Uusitalo, A. Altered oxidative stress in overtrained athletes. J. Sports Sci. 2010, 28, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.J.; Chiu, C.C.; Li, Y.P.; Huang, W.C.; Te Huang, Y.; Huang, C.C.; Chuang, H.L. Effect of intestinal microbiota on exercise performance in mice. J. Strength Cond. Res. 2015, 29, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Xu, C.; Chen, X.; Cai, X.; Yang, S.; Sheng, Y.; Wang, T. Regulation of an antioxidant blend on intestinal redox status and major microbiota in early weaned piglets. Nutr. 2014, 30, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Jeukendrup, A.; Vet-Joop, K.; Sturk, A.; Stegen, J.; Senden, J.; Saris, W.; Wagenmakers, A. Relationship between gastro-intestinal complaints and endotoxaemia, cytokine release and the acute-phase reaction during and after a long-distance triathlon in highly trained men. Clin. Sci. 2000, 98, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Holland, A.M.; Hyatt, H.W.; Smuder, A.J.; Sollanek, K.J.; Morton, A.B.; Roberts, M.D.; Kavazis, A.N. Influence of endurance exercise training on antioxidant enzymes, tight junction proteins, and inflammatory markers in the rat ileum. BMC Res. Notes 2015, 8, 514. [Google Scholar] [CrossRef]
- Schwellnus, M.P.; Derman, W.E.; Jordaan, E.; Page, T.; Lambert, M.I.; Readhead, C.; Roberts, C.; Kohler, R.; Collins, R.; Kara, S. Elite athletes travelling to international destinations > 5 time zone differences from their home country have a 2–3-fold increased risk of illness. Br. J. Sports Med. 2012, 46, 816–821. [Google Scholar] [CrossRef]
- Svendsen, I.S.; Taylor, I.M.; Tønnessen, E.; Bahr, R.; Gleeson, M. Training-related and competition-related risk factors for respiratory tract and gastrointestinal infections in elite cross-country skiers. Br. J. Sports Med. 2016, 50, 809–815. [Google Scholar] [CrossRef]
- Palleja, A.; Mikkelsen, K.H.; Forslund, S.K.; Kashani, A.; Allin, K.H.; Nielsen, T.; Hansen, T.H.; Liang, S.; Feng, Q.; Zhang, C. Recovery of gut microbiota of healthy adults following antibiotic exposure. Nat. Microbiol. 2018, 3, 1255. [Google Scholar] [CrossRef]
- Sode, J.; Obel, N.; Hallas, J.; Lassen, A. Use of fluroquinolone and risk of Achilles tendon rupture: A population-based cohort study. Eur. J. Clin. Pharmacol. 2007, 63, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Wiström, J.; Norrby, S.R.; Myhre, E.B.; Eriksson, S.; Granström, G.; Lagergren, L.; Englund, G.; Nord, C.E.; Svenungsson, B. Frequency of antibiotic-associated diarrhoea in 2462 antibiotic-treated hospitalized patients: A prospective study. J. Antimicrob. Chemother. 2001, 47, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Möller, G.B.; da Cunha Goulart, M.J.V.; Nicoletto, B.B.; Alves, F.D.; Schneider, C.D. Supplementation of Probiotics and Its Effects on Physically Active Individuals and Athletes: Systematic Review. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 481–492. [Google Scholar] [CrossRef]
- Parvez, S.; Malik, K.A.; Ah Kang, S.; Kim, H.Y. Probiotics and their fermented food products are beneficial for health. J. Appl. Microbiol. 2006, 100, 1171–1185. [Google Scholar] [CrossRef] [PubMed]
- PRNewswire Probiotics Market Will Register 7.5% CAGR to Exceed $67 Billion by 2024: Global Market Insights, Inc. Available online: https://www.prnewswire.com/news-releases/probiotics-market-will-register-75-cagr-to-exceed-67-billion-by-2024-global-market-insights-inc-681334981.html (accessed on 20 November 2018).
- Santosa, S.; Farnworth, E.; Jones, P.J. Probiotics and their potential health claims. Nutr. Rev. 2006, 64, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.L.; Thorsen, L.; Schwan, R.F.; Jespersen, L. Strain-specific probiotics properties of Lactobacillus fermentum, Lactobacillus plantarum and Lactobacillus brevis isolates from Brazilian food products. Food Microbiol. 2013, 36, 22–29. [Google Scholar] [CrossRef]
- Altenhoefer, A.; Oswald, S.; Sonnenborn, U.; Enders, C.; Schulze, J.; Hacker, J.; Oelschlaeger, T.A. The probioticEscherichia colistrain Nissle 1917 interferes with invasion of human intestinal epithelial cells by different enteroinvasive bacterial pathogens. FEMS Immunol. Med Microbiol. 2004, 40, 223–229. [Google Scholar] [CrossRef]
- Wehkamp, J.; Harder, J.; Wehkamp, K.; Wehkamp-von Meissner, B.; Schlee, M.; Enders, C.; Sonnenborn, U.; Nuding, S.; Bengmark, S.; Fellermann, K. NF-κB-and AP-1-mediated induction of human beta defensin-2 in intestinal epithelial cells by Escherichia coli Nissle 1917: A novel effect of a probiotic bacterium. Infect. Immun. 2004, 72, 5750–5758. [Google Scholar] [CrossRef]
- Ukena, S.N.; Singh, A.; Dringenberg, U.; Engelhardt, R.; Seidler, U.; Hansen, W.; Bleich, A.; Bruder, D.; Franzke, A.; Rogler, G.; et al. Probiotic Escherichia coli Nissle 1917 inhibits leaky gut by enhancing mucosal integrity. PLoS ONE 2007, 2, e1308. [Google Scholar] [CrossRef]
- Zyrek, A.A.; Cichon, C.; Helms, S.; Enders, C.; Sonnenborn, U.; Schmidt, M.A. Molecular mechanisms underlying the probiotic effects of Escherichia coli Nissle 1917 involve ZO-2 and PKCzeta redistribution resulting in tight junction and epithelial barrier repair. Cell. Microbiol. 2007, 9, 804–816. [Google Scholar] [CrossRef]
- Chmielewska, A.; Szajewska, H. Systematic review of randomised controlled trials: Probiotics for functional constipation. World J. Gastroenterol. 2010, 16, 69. [Google Scholar] [PubMed]
- Ukena, S.N.; Westendorf, A.M.; Hansen, W.; Rohde, M.; Geffers, R.; Coldewey, S.; Suerbaum, S.; Buer, J.; Gunzer, F. The host response to the probiotic Escherichia coli strain Nissle 1917: Specific up-regulation of the proinflammatory chemokine MCP-1. BMC Med Genet. 2005, 6, 43. [Google Scholar] [CrossRef] [PubMed]
- Henker, J.; Laass, M.; Blokhin, B.M.; Bolbot, Y.K.; Maydannik, V.G.; Elze, M.; Wolff, C.; Schulze, J. The probiotic Escherichia coli strain Nissle 1917 (EcN) stops acute diarrhoea in infants and toddlers. Eur. J. Pediatr. 2007, 166, 311–318. [Google Scholar] [CrossRef] [PubMed]
- De Smet, I.; Van Hoorde, L.; Vande Woestyne, M.; Christiaens, H.; Verstraete, W. Significance of bile salt hydrolytic activities of lactobacilli. J. Appl. Bacteriol. 1995, 79, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Pavlova, S.; Kilic, A.; Kilic, S.; So, J.S.; Nader-Macias, M.; Simoes, J.; Tao, L. Genetic diversity of vaginal lactobacilli from women in different countries based on 16S rRNA gene sequences. J. Appl. Microbiol. 2002, 92, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Isolauri, E.; Kaila, M.; Mykkänen, H.; Ling, W.H.; Salminen, S. Oral bacteriotherapy for viral gastroenteritis. Dig. Dis. Sci. 1994, 39, 2595–2600. [Google Scholar] [CrossRef]
- Vizoso Pinto, M.G.; Schuster, T.; Briviba, K.; Watzl, B.; Holzapfel, W.H.; Franz, C.M.A.P. Adhesive and Chemokine Stimulatory Properties of Potentially Probiotic Lactobacillus Strains. J. Food Prot. 2007, 70, 125–134. [Google Scholar] [CrossRef]
- Oh, N.S.; Joung, J.Y.; Lee, J.Y.; Kim, Y. Probiotic and anti-inflammatory potential of Lactobacillus rhamnosus 4B15 and Lactobacillus gasseri 4M13 isolated from infant feces. PLoS ONE 2018, 13, e0192021. [Google Scholar] [CrossRef]
- Reid, G. The scientific basis for probiotic strains ofLactobacillus. Appl. Environ. Microbiol. 1999, 65, 3763–3766. [Google Scholar]
- Giwercman, B.; Jensen, E.; Høiby, N.; Kharazmi, A.; Costerton, J. Induction of beta-lactamase production in Pseudomonas aeruginosa biofilm. Antimicrob. Agents Chemother. 1991, 35, 1008–1010. [Google Scholar] [CrossRef]
- Cox, A.J.; Pyne, D.B.; Saunders, P.U.; Fricker, P.A. Oral administration of the probiotic Lactobacillus fermentum VRI-003 and mucosal immunity in endurance athletes. Br. J. Sports Med. 2010, 44, 222–226. [Google Scholar] [CrossRef] [PubMed]
- West, N.P.; Pyne, D.B.; Cripps, A.W.; Hopkins, W.G.; Eskesen, D.C.; Jairath, A.; Christophersen, C.T.; Conlon, M.A.; Fricker, P.A. Lactobacillus fermentum (PCC®) supplementation and gastrointestinal and respiratory-tract illness symptoms: A randomised control trial in athletes. Nutr. J. 2011, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Shing, C.M.; Peake, J.M.; Lim, C.L.; Briskey, D.; Walsh, N.P.; Fortes, M.B.; Ahuja, K.D.K.; Vitetta, L. Effects of probiotics supplementation on gastrointestinal permeability, inflammation and exercise performance in the heat. Eur. J. Appl. Physiol. 2013, 114, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo-Cantabrana, C.; Delgado, S.; Ruiz, L.; Ruas-Madiedo, P.; Sánchez, B.; Margolles, A. Bifidobacteria and Their Health-Promoting Effects. Microbiol. Spectr. 2017, 5. [Google Scholar]
- Ruiz, L.; Margolles, A.; Sanchez, B. Bile resistance mechanisms in Lactobacillus and Bifidobacterium. Front. Microbiol. 2013, 4, 396. [Google Scholar] [CrossRef]
- Junick, J.; Blaut, M. Quantification of Human Fecal Bifidobacterium Species by Use of Quantitative Real-Time PCR Analysis Targeting thegroELGene. Appl. Environ. Microbiol. 2012, 78, 2613–2622. [Google Scholar] [CrossRef] [PubMed]
- Selinger, C.P.; Bell, A.; Cairns, A.; Lockett, M.; Sebastian, S.; Haslam, N. Probiotic VSL#3 prevents antibiotic-associated diarrhoea in a double-blind, randomized, placebo-controlled clinical trial. J. Hosp. Infect. 2013, 84, 159–165. [Google Scholar]
- Imaoka, A.; Shima, T.; Kato, K.; Mizuno, S.; Uehara, T.; Matsumoto, S.; Setoyama, H.; Hara, T.; Umesaki, Y. Anti-inflammatory activity of probiotic Bifidobacterium: Enhancement of IL-10 production in peripheral blood mononuclear cells from ulcerative colitis patients and inhibition of IL-8 secretion in HT-29 cells. World J. Gastroenterol. 2008, 14, 2511–2516. [Google Scholar] [CrossRef]
- West, N.P.; Horn, P.L.; Pyne, D.B.; Gebski, V.J.; Lahtinen, S.J.; Fricker, P.A.; Cripps, A.W. Probiotic supplementation for respiratory and gastrointestinal illness symptoms in healthy physically active individuals. Clin. Nutr. 2014, 33, 581–587. [Google Scholar] [CrossRef]
- Haywood, B.A.; Black, K.E.; Baker, D.; McGarvey, J.; Healey, P.; Brown, R.C. Probiotic supplementation reduces the duration and incidence of infections but not severity in elite rugby union players. J. Sci. Med. Sport 2014, 17, 356–360. [Google Scholar] [CrossRef]
- Jäger, R.; Purpura, M.; Stone, J.; Turner, S.; Anzalone, A.; Eimerbrink, M.; Pane, M.; Amoruso, A.; Rowlands, D.; Oliver, J. Probiotic Streptococcus thermophilus FP4 and Bifidobacterium breve BR03 supplementation attenuates performance and range-of-motion decrements following muscle damaging exercise. Nutrients 2016, 8, 642. [Google Scholar] [CrossRef] [PubMed]
- Pugh, J.N.; Sparks, A.S.; Doran, D.A.; Fleming, S.C.; Langan-Evans, C.; Kirk, B.; Fearn, R.; Morton, J.P.; Close, G.L. Four weeks of probiotic supplementation reduces GI symptoms during a marathon race. Eur. J. Appl. Physiol. 2019, 119, 1491–1501. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Naturer 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]
- McFarland, L.V. Meta-analysis of probiotics for the prevention of traveler’s diarrhea. Travel Med. Infect. Dis. 2007, 5, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Thygesen, J.B.; Glerup, H.; Tarp, B. Saccharomyces boulardii fungemia caused by treatment with a probioticum. BMJ Case Rep. 2012. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.H.; Jo, S.Y.; Park, H.J.; Chang, S.K.; Byeon, J.-S.; Myung, S.-J. A randomized, double-blind, placebo-controlled multicenter trial of Saccharomyces boulardii in irritable bowel syndrome: Effect on quality of life. J. Clin. Gastroenterol. 2011, 45, 679–683. [Google Scholar] [CrossRef] [PubMed]
- Kurugöl, Z.; Koturoğlu, G. Effects of Saccharomyces boulardii in children with acute diarrhoea. Acta Paediatr. 2005, 94, 44–47. [Google Scholar] [CrossRef]
- Billoo, A.; Memon, M.; Khaskheli, S.; Murtaza, G.; Iqbal, K.; Shekhani, M.S.; Siddiqi, A.Q. Role of a probiotic (Saccharomyces boulardii) in management and prevention of diarrhoea. World J. Gastroenterol. 2006, 12, 4557. [Google Scholar] [CrossRef]
- Walter, J. Ecological Role of Lactobacilli in the Gastrointestinal Tract: Implications for Fundamental and Biomedical Research. Appl. Environ. Microbiol. 2008, 74, 4985–4996. [Google Scholar] [CrossRef]
- Dugas, B.; Mercenier, A.; Lenoir-Wijnkoop, I.; Arnaud, C.; Dugas, N.; Postaire, E. Immunity and probiotics. Immunol. Today 1999, 20, 387–390. [Google Scholar] [CrossRef]
- Bezirtzoglou, E.; Stavropoulou, E. Immunology and probiotic impact of the newborn and young children intestinal microflora. Anaerobe 2011, 17, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Tulumoglu, S.; Yuksekdag, Z.N.; Beyatli, Y.; Simsek, O.; Cinar, B.; Yaşar, E. Probiotic properties of lactobacilli species isolated from children’s feces. Anaerobe 2013, 24, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Montes, R.; Bayless, T.M.; Saavedra, J.; Perman, J. Effect of milks inoculated with Lactobacillus acidophilus or a yogurt starter culture in lactose-maldigesting children. J. Dairy Sci. 1995, 78, 1657–1664. [Google Scholar] [CrossRef]
- Kim, Y.; Whang, J.Y.; Whang, K.Y.; Oh, S.; Kim, S.H. Characterization of the Cholesterol-Reducing Activity in a Cell-Free Supernatant ofLactobacillus acidophilusATCC 43121. Biosci. Biotechnol. Biochem. 2014, 72, 1483–1490. [Google Scholar] [CrossRef] [PubMed]
- Almeida, C.C.; Lorena, S.L.; Pavan, C.R.; Akasaka, H.M.; Mesquita, M.A. Beneficial effects of long-term consumption of a probiotic combination of Lactobacillus casei Shirota and Bifidobacterium breve Yakult may persist after suspension of therapy in lactose-intolerant patients. Nutr. Clin. Pract. 2012, 27, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kwon, J.H.; Ahn, S.H.; Lee, S.I.; Han, Y.S.; Choi, Y.O.; Lee, S.Y.; Ahn, K.M.; Ji, G.E. Effect of probiotic mix (Bifidobacterium bifidum, Bifidobacterium lactis, Lactobacillus acidophilus) in the primary prevention of eczema: A double-blind, randomized, placebo-controlled trial. Pediatric Allergy Immunol. 2010, 21, e386–e393. [Google Scholar] [CrossRef] [PubMed]
- Zanotti, I.; Turroni, F.; Piemontese, A.; Mancabelli, L.; Milani, C.; Viappiani, A.; Prevedini, G.; Sanchez, B.; Margolles, A.; Elviri, L.; et al. Evidence for cholesterol-lowering activity by Bifidobacterium bifidum PRL2010 through gut microbiota modulation. Appl. Microbiol. Biotechnol. 2015, 99, 6813–6829. [Google Scholar] [CrossRef] [PubMed]
- Behnsen, J.; Deriu, E.; Sassone-Corsi, M.; Raffatellu, M. Probiotics: Properties, examples, and specific applications. Cold Spring Harb. Perspect. Med. 2013, 3, a010074. [Google Scholar] [CrossRef]
- Troy, E.B.; Kasper, D.L. Beneficial effects of Bacteroides fragilis polysaccharides on the immune system. Front. Biosci. A J. Virtual Library 2010, 15, 25. [Google Scholar] [CrossRef]
- Curtis, M.M.; Hu, Z.; Klimko, C.; Narayanan, S.; Deberardinis, R.; Sperandio, V. The Gut Commensal Bacteroides thetaiotaomicron Exacerbates Enteric Infection through Modification of the Metabolic Landscape. Cell Host Microbe 2014, 16, 759–769. [Google Scholar] [CrossRef]
- Yanagibashi, T.; Hosono, A.; Oyama, A.; Tsuda, M.; Suzuki, A.; Hachimura, S.; Takahashi, Y.; Momose, Y.; Itoh, K.; Hirayama, K.; et al. IgA production in the large intestine is modulated by a different mechanism than in the small intestine: Bacteroides acidifaciens promotes IgA production in the large intestine by inducing germinal center formation and increasing the number of IgA + B cells. Immunobiology 2013, 218, 645–651. [Google Scholar] [PubMed]
- Kirmiz, N.; Galindo, K.; Cross, K.L.; Luna, E.; Rhoades, N.; Podar, M.; Flores, G.E. Comparative genomics guides elucidation of vitamin B12 biosynthesis in novel human associated Akkermansia. bioRxiv 2019. [Google Scholar] [CrossRef]
- Cani, P.D.; de Vos, W.M. Next-Generation Beneficial Microbes: The Case of Akkermansia muciniphila. Front. Microbiol. 2017, 8, 1765. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.-R.; Lee, J.-C.; Lee, H.-Y.; Kim, M.-S.; Whon, T.W.; Lee, M.-S.; Bae, J.-W. An increase in the Akkermansia spp. population induced by metformin treatment improves glucose homeostasis in diet-induced obese mice. Gut 2014, 63, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 2016, 23, 107. [Google Scholar] [CrossRef] [PubMed]
- De Vos, W.M.; Müller, M.; Norin, E.; Hooiveld, G.; Van Baarlen, P.; Derrien, M. Modulation of Mucosal Immune Response, Tolerance, and Proliferation in Mice Colonized by the Mucin-Degrader Akkermansia muciniphila. Front. Microbiol. 2011, 2, 166. [Google Scholar]
- Sanz, Y.; Ottman, N.; Reunanen, J.; Meijerink, M.; Pietilä, T.E.; Kainulainen, V.; Klievink, J.; Huuskonen, L.; Aalvink, S.; Skurnik, M.; et al. Pili-like proteins of Akkermansia muciniphila modulate host immune responses and gut barrier function. PLoS ONE 2017, 12, e0173004. [Google Scholar]
- Rossi, O.; van Berkel, L.A.; Chain, F.; Tanweer Khan, M.; Taverne, N.; Sokol, H.; Duncan, S.H.; Flint, H.J.; Harmsen, H.J.M.; Langella, P.; et al. Faecalibacterium prausnitzii A2-165 has a high capacity to induce IL-10 in human and murine dendritic cells and modulates T cell responses. Sci. Rep. 2016, 6, 18507. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Miquel, S.; Chain, F.; Natividad, J.M.; Jury, J.; Lu, J.; Sokol, H.; Theodorou, V.; Bercik, P.; Verdu, E.F.; et al. Faecalibacterium prausnitzii prevents physiological damages in a chronic low-grade inflammation murine model. BMC Microbiol. 2015, 15, 67. [Google Scholar] [CrossRef]
- Udayappan, S.; Manneras-Holm, L.; Chaplin-Scott, A.; Belzer, C.; Herrema, H.; Dallinga-Thie, G.M.; Duncan, S.H.; Stroes, E.S.; Groen, A.K.; Flint, H.J. Oral treatment with Eubacterium hallii improves insulin sensitivity in db/db mice. NPJ Biofilms Microbiomes 2016, 2, 16009. [Google Scholar] [CrossRef] [PubMed]
- Engels, C.; Ruscheweyh, H.-J.; Beerenwinkel, N.; Lacroix, C.; Schwab, C. The Common Gut Microbe Eubacterium hallii also Contributes to Intestinal Propionate Formation. Front. Microbiol. 2016, 7, 713. [Google Scholar] [CrossRef] [PubMed]
- Fekry, M.I.; Engels, C.; Zhang, J.; Schwab, C.; Lacroix, C.; Sturla, S.J.; Chassard, C. The strict anaerobic gut microbe Eubacterium hallii transforms the carcinogenic dietary heterocyclic amine 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP). Environ. Microbiol. Rep. 2016, 8, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Kanauchi, O.; Fukuda, M.; Matsumoto, Y.; Ishii, S.; Ozawa, T.; Shimizu, M.; Mitsuyama, K.; Andoh, A. Eubacterium limosum ameliorates experimental colitis and metabolite of microbe attenuates colonic inflammatory action with increase of mucosal integrity. World J. Gastroenterol. 2006, 12, 1071. [Google Scholar] [CrossRef] [PubMed]
- Brubaker, J.O.; Li, Q.; Tzianabos, A.O.; Kasper, D.L.; Finberg, R.W. Mitogenic activity of purified capsular polysaccharide A from Bacteroides fragilis: Differential stimulatory effect on mouse and rat lymphocytes in vitro. J. Immunol. 1999, 162, 2235–2242. [Google Scholar]
- Hack, V.; Weiss, C.; Friedmann, B.; Suttner, S.; Schykowski, M.; Erbe, N.; Benner, A.; Bartsch, P.; Droge, W. Decreased plasma glutamine level and CD4+ T cell number in response to 8 wk of anaerobic training. Am. J. Physiol. Endocrinol. Metab. 1997, 272, E788–E795. [Google Scholar] [CrossRef] [PubMed]
- Laing, S.; Gwynne, D.; Blackwell, J.; Williams, M.; Walters, R.; Walsh, N. Salivary IgA response to prolonged exercise in a hot environment in trained cyclists. Eur. J. Appl. Physiol. 2005, 93, 665–671. [Google Scholar] [CrossRef]
- Mackinnon, L.T.; Ginn, E.; Seymour, G.J. Decreased salivary immunoglobulin A secretion rate after intense interval exercise in elite kayakers. Eur. J. Appl. Physiol. Occup. Physiol. 1993, 67, 180–184. [Google Scholar] [CrossRef]
- Neville, V.; Gleeson, M.; Folland, J.P. Salivary IgA as a risk factor for upper respiratory infections in elite professional athletes. Med. Sci. Sports Exerc. 2008, 40, 1228–1236. [Google Scholar] [CrossRef]
- Pals, K.L.; Chang, R.-T.; Ryan, A.J.; Gisolfi, C.V. Effect of running intensity on intestinal permeability. J. Appl. Physiol. 1997, 82, 571–576. [Google Scholar] [CrossRef]
- Øktedalen, O.; Lunde, O.; Opstad, P.; Aabakken, L.; Kvernebo, K. Changes in the gastrointestinal mucosa after long-distance running. Scand. J. Gastroenterol. 1992, 27, 270–274. [Google Scholar] [CrossRef]
- Soman, V.R.; Koivisto, V.A.; Deibert, D.; Felig, P.; DeFronzo, R.A. Increased insulin sensitivity and insulin binding to monocytes after physical training. N. Engl. J. Med. 1979, 301, 1200–1204. [Google Scholar] [CrossRef] [PubMed]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humarán, L.G.; Gratadoux, J.-J.; Blugeon, S.; Bridonneau, C.; Furet, J.-P.; Corthier, G. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef] [PubMed]
- Chua, K.J.; Kwok, W.C.; Aggarwal, N.; Sun, T.; Chang, M.W. Designer probiotics for the prevention and treatment of human diseases. Curr. Opin. Chem. Biol. 2017, 40, 8–16. [Google Scholar] [CrossRef]
- Amalaradjou, M.A.; Bhunia, A.K. Bioengineered probiotics, a strategic approach to control enteric infections. Bioengineered 2013, 4, 379–387. [Google Scholar] [CrossRef]
- Doron, S.; Snydman, D.R. Risk and safety of probiotics. Clin. Infect. Dis. 2015, 60 (suppl. 2), S129–S134. [Google Scholar] [CrossRef] [PubMed]
- Panel, E.F. Guidance on the Characterisation Microorganisms 1 Used as Feed Additives or as Production Organisms; EFSA: Parma, Italy, 2017.
- Organisation, W.G. Probiotics and Prebiotic; WGO: Forest City, IA, USA, 2017. [Google Scholar]
- Health, N.I.O. Probiotics. Available online: https://ods.od.nih.gov/factsheets/Probiotics-HealthProfessional/ (accessed on 25 June 2019).
- Timmerman, H.; Koning, C.; Mulder, L.; Rombouts, F.; Beynen, A. Monostrain, multistrain and multispecies probiotics—A comparison of functionality and efficacy. Int. J. Food Microbiol. 2004, 96, 219–233. [Google Scholar] [CrossRef]
- Anadón, A.; Martínez-Larrañaga, M.R.; Arés, I.; Martínez, M.A. Prebiotics and Probiotics in Feed and Animal Health. In Nutraceuticals in Veterinary Medicine; Springer: Berlin/Heidelberg, Germany, 2016; pp. 3–23. [Google Scholar]
- Helenius, I.J.; Tikkanen, H.O.; Sarna, S.; Haahtela, T. Asthma and increased bronchial responsiveness in elite athletes: Atopy and sport event as risk factors. J. Allergy Clin. Immunol. 1998, 101, 646–652. [Google Scholar] [CrossRef]
- Nieman, D.C.; Johanssen, L.M.; Lee, J.W.; Arabatzis, K. Infectious episodes in runners before and after the Los Angeles Marathon. J Sports Med. Phys. Fit. 1990, 30, 316–328. [Google Scholar]
- Spence, L.; Brown, W.J.; Pyne, D.B.; Nissen, M.D.; Sloots, T.P.; McCormack, J.G.; Locke, A.S.; Fricker, P.A. Incidence, etiology, and symptomatology of upper respiratory illness in elite athletes. Med. Sci. Sports Exerc. 2007, 39, 577–586. [Google Scholar] [CrossRef]
- Ter Steege, R.W.; Van Der Palen, J.; Kolkman, J.J. Prevalence of gastrointestinal complaints in runners competing in a long-distance run: An internet-based observational study in 1281 subjects. Scand. J. Gastroenterol. 2008, 43, 1477–1482. [Google Scholar] [CrossRef] [PubMed]
- Gouttebarge, V.; Frings-Dresen, M.; Sluiter, J. Mental and psychosocial health among current and former professional footballers. Occup. Med. 2015, 65, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Martarelli, D.; Verdenelli, M.C.; Scuri, S.; Cocchioni, M.; Silvi, S.; Cecchini, C.; Pompei, P. Effect of a Probiotic Intake on Oxidant and Antioxidant Parameters in Plasma of Athletes During Intense Exercise Training. Curr. Microbiol. 2011, 62, 1689–1696. [Google Scholar] [CrossRef] [PubMed]
- Lamprecht, M.; Frauwallner, A. Exercise, intestinal barrier dysfunction and probiotic supplementation. Med. Sport Sci. 2012, 59, 47–56. [Google Scholar] [PubMed]
- Clancy, R.; Gleeson, M.; Cox, A.; Callister, R.; Dorrington, M.; D’este, C.; Pang, G.; Pyne, D.; Fricker, P.; Henriksson, A. Reversal in fatigued athletes of a defect in interferon γ secretion after administration of Lactobacillus acidophilus. Br. J. Sports Med. 2006, 40, 351–354. [Google Scholar] [CrossRef]
- Sashihara, T.; Nagata, M.; Mori, T.; Ikegami, S.; Gotoh, M.; Okubo, K.; Uchida, M.; Itoh, H. Effects of Lactobacillus gasseri OLL2809 and α-lactalbumin on university-student athletes: A randomized, double-blind, placebo-controlled clinical trial. Appl. Physiol. Nutr. Metab. 2013, 38, 1228–1235. [Google Scholar] [CrossRef] [PubMed]
- Strasser, B.; Geiger, D.; Schauer, M.; Gostner, J.M.; Gatterer, H.; Burtscher, M.; Fuchs, D. Probiotic Supplements Beneficially Affect Tryptophan-Kynurenine Metabolism and Reduce the Incidence of Upper Respiratory Tract Infections in Trained Athletes: A Randomized, Double-Blinded, Placebo-Controlled Trial. Nutrients 2016, 8, 752. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant Properties of Probiotic Bacteria. Nutrients 2017, 9, 521. [Google Scholar] [CrossRef]
- Saide, J.; Gilliland, S. Antioxidative activity of lactobacilli measured by oxygen radical absorbance capacity. J. Dairy Sci. 2005, 88, 1352–1357. [Google Scholar] [CrossRef]
- Brown, A.J.; Goldsworthy, S.M.; Barnes, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; Muir, A.I.; Wigglesworth, M.J.; Kinghorn, I.; Fraser, N.J. The Orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; Deroos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Zuhl, M.; Schneider, S.; Lanphere, K.; Conn, C.; Dokladny, K.; Moseley, P. Exercise regulation of intestinal tight junction proteins. Br. J. Sports Med. 2014, 48, 980–986. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.M.; Konstaninov, S.; Konings, I.; Karczewski, J. Effects of probiotic and commensals on epithelial barrier function. Int. J. Probiotics Prebiotics 2008, 3, 127–132. [Google Scholar]
- Lamprecht, M.; Bogner, S.; Schippinger, G.; Steinbauer, K.; Fankhauser, F.; Hallstroem, S.; Schuetz, B.; Greilberger, J.F. Probiotic supplementation affects markers of intestinal barrier, oxidation, and inflammation in trained men; a randomized, double-blinded, placebo-controlled trial. J. Int. Soc. Sports Nutr. 2012, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Hackney, A.C.; Koltun, K.J. The immune system and overtraining in athletes: Clinical implications. Acta Clin. Croat. 2012, 51, 633–641. [Google Scholar]
- West, N.; Pyne, D.; Peake, J.; Cripps, A. Probiotics, immunity and exercise: A review. Exerc. Immunol. Rev. 2009, 15, e26. [Google Scholar]
- Mack, D.R.; Michail, S.; Wei, S.; McDougall, L.; Hollingsworth, M.A. Probiotics inhibit enteropathogenic E. coli adherence in vitro by inducing intestinal mucin gene expression. Am. J. Physiol. Gastrointest. Liver Physiol. 1999, 276, G941–G950. [Google Scholar] [CrossRef]
- Snyder, J.D.; Walker, A. Structure and Function of Intestinal Mucin: Developmental Aspects. Int. Arch. Allergy Immunol. 1987, 82, 351–356. [Google Scholar] [CrossRef]
- Möndel, M.; Schroeder, B.; Zimmermann, K.; Huber, H.; Nuding, S.; Beisner, J.; Fellermann, K.; Stange, E.; Wehkamp, J. Probiotic E. coli treatment mediates antimicrobial human β-defensin synthesis and fecal excretion in humans. Mucosal Immunol. 2009, 2, 166. [Google Scholar] [CrossRef]
- Kekkonen, R.A.; Vasankari, T.J.; Vuorimaa, T.; Haahtela, T.; Julkunen, I.; Korpela, R. The effect of probiotics on respiratory infections and gastrointestinal symptoms during training in marathon runners. Int. J. Sport Nutr. Exerc. Metab. 2007, 17, 352–363. [Google Scholar] [CrossRef] [PubMed]
- Peters, E.M.; Bateman, E. Ultramarathon running and upper respiratory tract infections. An epidemiological survey. S. Afr. Med J. 1983, 64, 582–584. [Google Scholar] [PubMed]
- Romano, R.; Lu, D.; Holtom, P. Outbreak of community-acquired methicillin-resistant Staphylococcus aureus skin infections among a collegiate football team. J. Athl. Train. 2006, 41, 141. [Google Scholar] [PubMed]
- Suskovic, J. Antimicrobial Activity—The Most Important Property of probiotic and starter lactic acid bacteria. Food Technol. Biotechnol. 2010, 48, 296–307. [Google Scholar]
- Shokryazdan, P.; Sieo, C.C.; Kalavathy, R.; Liang, J.B.; Alitheen, N.B.; Faseleh Jahromi, M.; Ho, Y.W. Probiotic potential of Lactobacillus strains with antimicrobial activity against some human pathogenic strains. BioMed Res. Int. 2014, 2014, 927268. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.D.; Hill, C.; Ross, R.P. Food microbiology: Bacteriocins: Developing innate immunity for food. Nature Rev. Microbiol. 2005, 3, 777. [Google Scholar] [CrossRef] [PubMed]
- Hegarty, J.W.; Guinane, C.M.; Ross, R.P.; Hill, C.; Cotter, P.D. Bacteriocin production: A relatively unharnessed probiotic trait? F1000Research 2016, 5, 2587. [Google Scholar] [CrossRef]
- Dobson, A.; Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocin Production: A Probiotic Trait? Appl. Environ. Microbiol. 2012, 78, 1–6. [Google Scholar] [CrossRef]
- Hegarty, J.W.; Guinane, C.M.; Ross, R.P.; Hill, C.; Cotter, P.D. Lack of Heterogeneity in Bacteriocin Production Across a Selection of Commercial Probiotic Products. Probiotics Antimicrob. Proteins 2017, 9, 459–465. [Google Scholar] [CrossRef]
- Reardon, C.L.; Hainline, B.; Aron, C.M.; Baron, D.; Baum, A.L.; Bindra, A.; Budgett, R.; Campriani, N.; Castaldelli-Maia, J.M.; Currie, A. Mental health in elite athletes: International Olympic Committee consensus statement. Br. J. Sports Med. 2019, 53, 667–699. [Google Scholar] [CrossRef]
- Dougherty, D.M.; Marsh-Richard, D.M.; Mathias, C.W.; Hood, A.J.; Addicott, M.A.; Moeller, F.G.; Morgan, C.J.; Badawy, A.A.-B. Comparison of 50-and 100-g L-tryptophan depletion and loading formulations for altering 5-HT synthesis: Pharmacokinetics, side effects, and mood states. Psychopharmacology 2008, 198, 431. [Google Scholar] [CrossRef]


| Subject Group | Microbiome Change | Key Findings | Study Reference |
|---|---|---|---|
| Rugby players | ↑Akkermansia, Prevotella, ↓Ruminococcaceae Bacteroides Lactobacillus | Akkermanisa was associated with better immunity and gut barrier function while Prevotella was correlated to biosynthesis of branched-chain amino acid (BCAA) pathways which help with muscle recovery | [14] |
| Professional male athletes | ↑Akkermansia metabolic pathways/higher short-chain fatty acids (SCFA) metabolic pathways | Rugby players had a higher abundance of health-promoting Akkermansia genus, which has been associated with an improved gut barrier function and immune function stimulation. | [13] |
| Cyclists | ↑Prevotella, Methanobrevibacter Smithii | Prevotella was correlated to biosynthesis of BCAA pathways which help with muscle recovery, M. smithii has been associated with degradation of H2 which is used to make ATP/SCFA resulting in a more energetically efficient body | [15] |
| Sedentary adults challenged to eight week exercise regime | ↓in Archaea species and an ↑ in microbial diversity | Microbial diversity has been linked to an overall better health | [16] |
| Marathon runners | ↑Veillonella | Veillonella has been shown to metabolize lactate to SCFA, lower inflammation and increase performance in murine models | [18] |
| Bodybuilders and Distance runners | ↑Faecalibacterium, Sutterella, Clostridium, Haemophilus, Eisenbergiella ↓Bifidobacterium, Parasutterella and Eubacterium | Different sports and their sport specific diets can affect the gut microbiome in different ways | [17] |
| Subject Group | Intervention | Result | Limitations of the Study | References |
|---|---|---|---|---|
| 20 male elite distance runners. Randomized double-blinded, placebo controlled trial, | Lb. fermentum VRI-003 | ↓ risk and severity of respiratory systems ↑ INFƴ | A small sample size | [64] |
| 99 male and female competitive cyclists. Randomized, double-blinded, placebo trial | Lb. fermentum (PCC®) | ↓ severity of GIT symptoms ↓ severity/duration of lower respiratory illness ↓ cytokine imbalance | Inclusion criteria for antibiotics was only four weeks, study relied on self-reported illness, reported a higher rate of lower respiratory illness in females. | [65] |
| 10 male runners. Randomized, double-blinded, placebo trial | Lb. acidophilus, Lb. rhamnosus, Lb. casei, Lb. plantarum, Lb. fermentum, B. lactis, B. breve, B.bifidum and Streptococcus thermophilus | ↑ increased run time to fatigue, small to moderate improvement in gut permeability | Study only investigated males, sample size was too small, short study duration of 4 weeks. | [66] |
| 465 physically active males and females. Randomized double blind placebo controlled trial | B. animalis subsp. lactis BI-04 (BI-04), Lb. acidophilus NCFM and B.animalis subsp. lactis BI-04 (NCFM and BI-04) | ↓ the risk of URTIs by 27% | No separation between recreational and professional athletes, study relied on self-reported illness data, inclusion criteria for antibiotic use was only four weeks. | [72] |
| 30 male elite male rugby players. Randomized, double-blinded, Placebo trial. | B. bifidum B. longum Lb. gasseri | ↓ in the incidence of URTI/GIT | Relatively small sample size, study only looked at males, short study duration of 4 weeks, relied on self-reported illness data as opposed to measuring immune system markers | [73] |
| 15 resistance-trained men. Randomized, double-blinded, placebo trial. | B. breve BR03 and S. thermophilus FP4 | positive effect on the reduced performance and range of motion followed by intense muscle damaging exercise | Small sample size, looked at males only, didn’t include antibiotic use in inclusion criteria, short study duration of 3 weeks | [74] |
| 24 amateur athletes | Lb. acidophilus (CUL60/CUL21), B. bifidum (CUL20), B. animalis subsp. Lactis (CUL34) | ↓ incidence and severity of GIT symptoms, both during training and a marathon race | Small sample size, ratio of males to females was skewed, athletic levels weren’t standardized, not double-blinded. | [75] |
| 24 amateur male athletes | Lb. rhamnosus IMC 501 and Lb. paracasei IMC 502 | ↓antioxidant levels followed by exercise | Small sample size, short study duration of 4 weeks, studied males only, study did not include placebo in control group | [130] |
| 23 endurance male athletes Randomized, double-blind, placebo controlled trial. | EcologicWPerformance or OMNi-BiOTiCWPOWER, | ↓ zonulin ↓TNF-alpha | Small sample size, only looked at men, looked at only one marker or impaired intestinal permeability | [131] |
| 27 trained amateur athletes | Lb. acidophilus LAFTI® | ↓fatigue | Small sample size, sample size of control group was significantly lower than test group, the ratio of male to females was skewed, not randomized or double blinded. | [132] |
| 44 university student athletes. Randomized, double-blind, placebo controlled trial. | Lb. gasseri OLL2809 combined with alpha-lactalbumin | prevents the exercise induced drops in Natural Killer cells positive effect on minor fatigue ↑mood from a depressive state | Study only looked at males, all participants were at a university level, relied on self-reported illness, short study duration of 4 weeks | [133] |
| 33 highly trained individuals. Randomized, double-blinded, placebo controlled trial. | B. bifidum W23, B. W51, Enterococcus faecium W54, Lb. acidophilus W22, Lb. brevis W63, and Lactococcus lactis W58 | ↓drops of tryptophan levels caused by intense exercise ↓incidence of URTI’s | Relatively small sample size, women were overrepresented; the severity of illness could not be calculated due to lack of replies from participants. | [134] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wosinska, L.; Cotter, P.D.; O’Sullivan, O.; Guinane, C. The Potential Impact of Probiotics on the Gut Microbiome of Athletes. Nutrients 2019, 11, 2270. https://doi.org/10.3390/nu11102270
Wosinska L, Cotter PD, O’Sullivan O, Guinane C. The Potential Impact of Probiotics on the Gut Microbiome of Athletes. Nutrients. 2019; 11(10):2270. https://doi.org/10.3390/nu11102270
Chicago/Turabian StyleWosinska, Laura, Paul D. Cotter, Orla O’Sullivan, and Caitriona Guinane. 2019. "The Potential Impact of Probiotics on the Gut Microbiome of Athletes" Nutrients 11, no. 10: 2270. https://doi.org/10.3390/nu11102270
APA StyleWosinska, L., Cotter, P. D., O’Sullivan, O., & Guinane, C. (2019). The Potential Impact of Probiotics on the Gut Microbiome of Athletes. Nutrients, 11(10), 2270. https://doi.org/10.3390/nu11102270