Beneficial Effects of an Aged Black Garlic Extract in the Metabolic and Vascular Alterations Induced by a High Fat/Sucrose Diet in Male Rats

 ,
,
Abstract
1. Introduction
2. Material and Methods
2.1. Materials
2.2. Physical-Chemical Characterization of ABG10+® Extracts
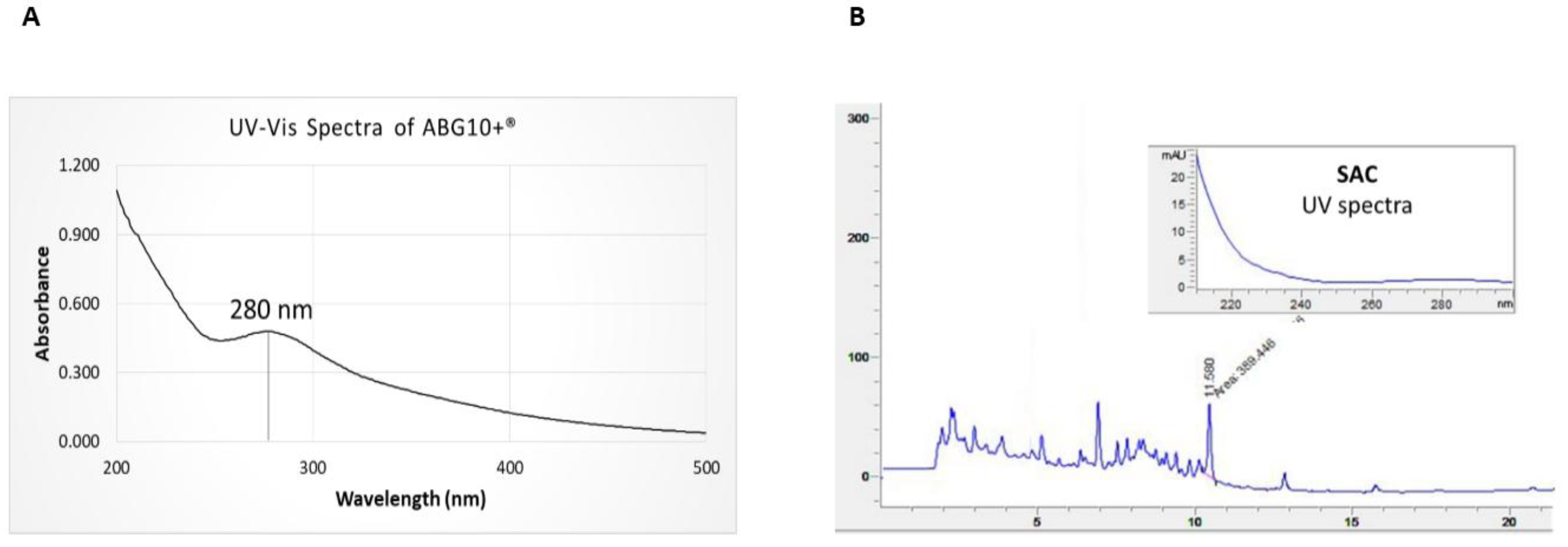
2.2.1. Absorption Spectra of ABG10+®
2.2.2. Analysis of Organosulfur Aminoacids by HPLC-PAD/MSD
2.3. In Vivo Experiments
2.3.1 Animals
2.3.2. Oral Glucose Tolerance Test (OGTT) and Homeostatic Model Assessment of Insulin Resistance (HOMA-IR)
2.3.3. Serums Measurements
2.3.4. RNA Extraction and Quantitative Real-Time PCR
2.3.5. Vascular Reactivity Experiments
2.4. Statistical Analysis
3. Results
3.1. Physicochemical Characterization of ABG10+® Extract
3.2. In Vivo Experiments in Rats
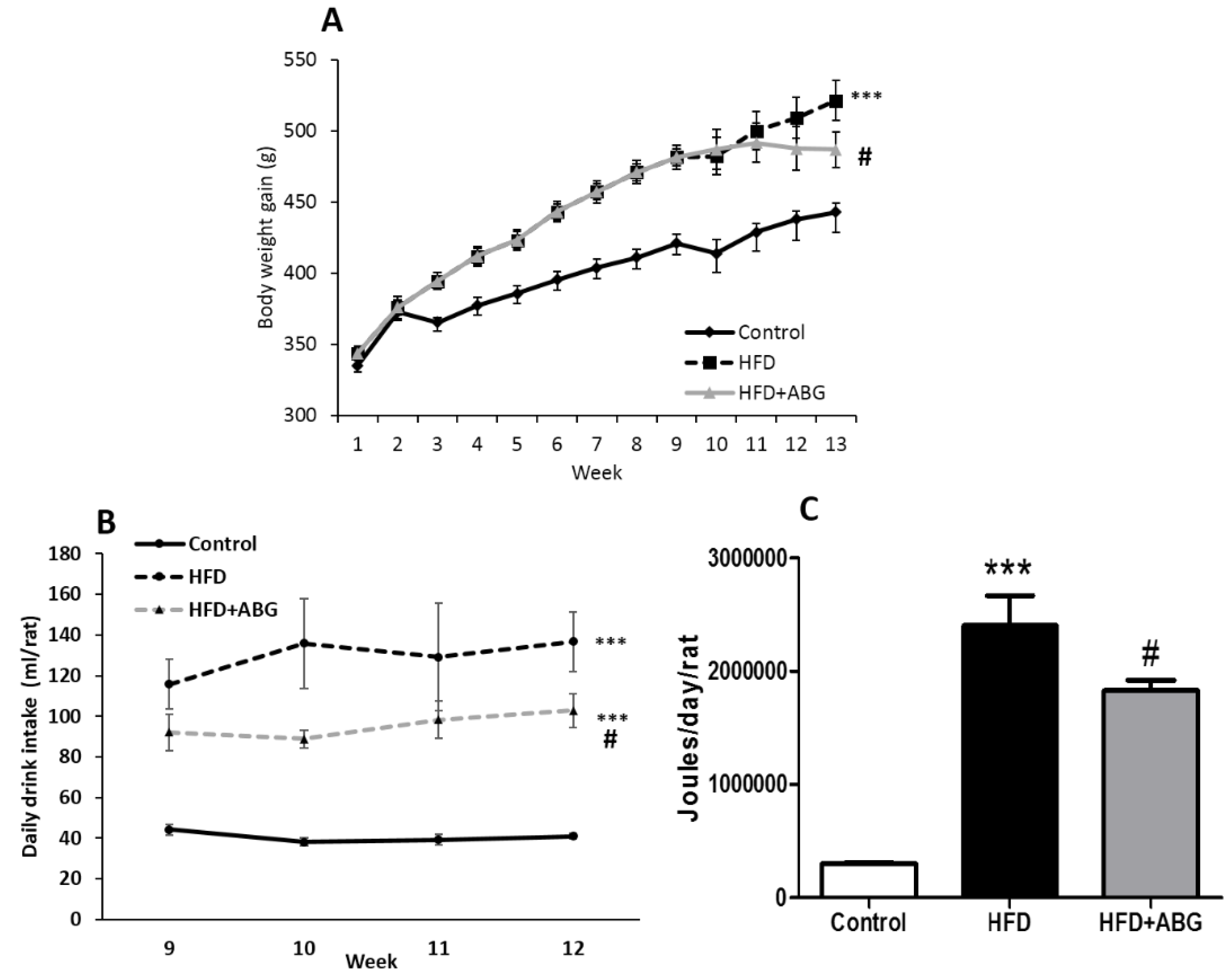
3.2.1. Body and Organ Weights
3.2.2. Daily Caloric Intake
3.2.3. Triglycerides, Total Cholesterol, LDL Cholesterol and HDL Cholesterol
3.2.4. Glycaemia, Insulin, Leptin and Adiponectin Serum Concentrations
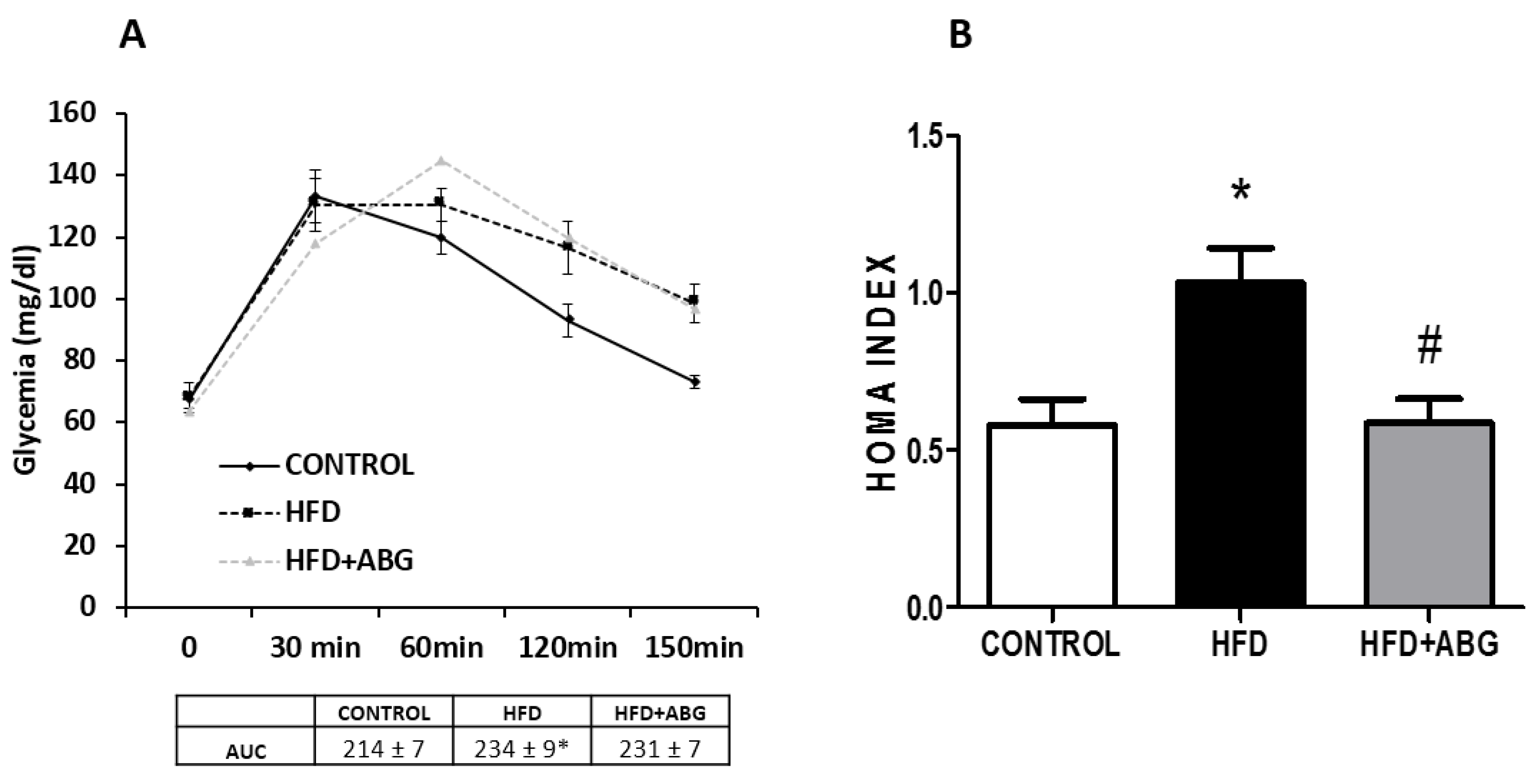
3.2.5. Oral Glucose Tolerance Test (OGTT) and HOMA Index
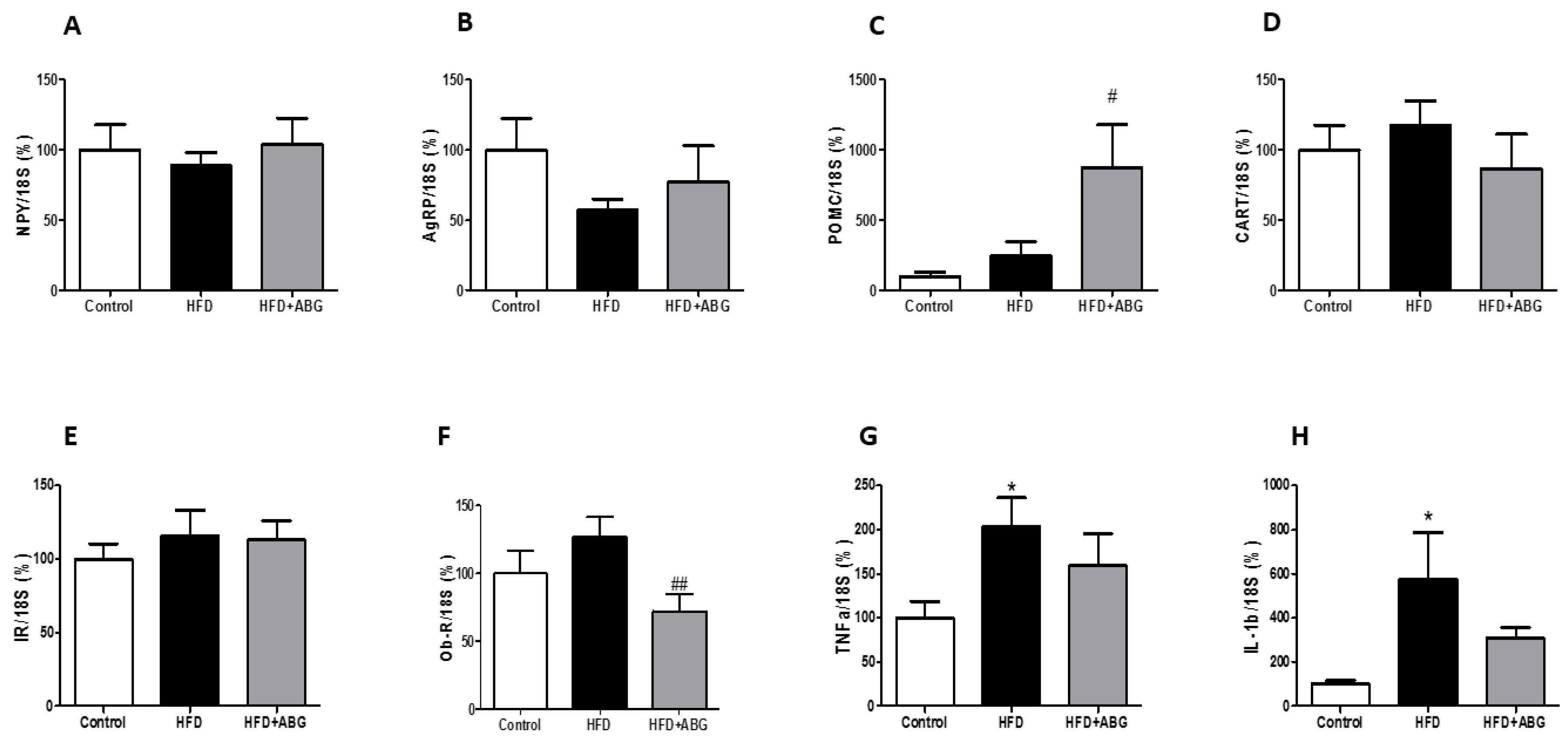
3.2.6. NPY, AgRP, POMC, CART, InsR, ObR mRNA, TNF-α and IL-1β Levels in the Hypothalamus
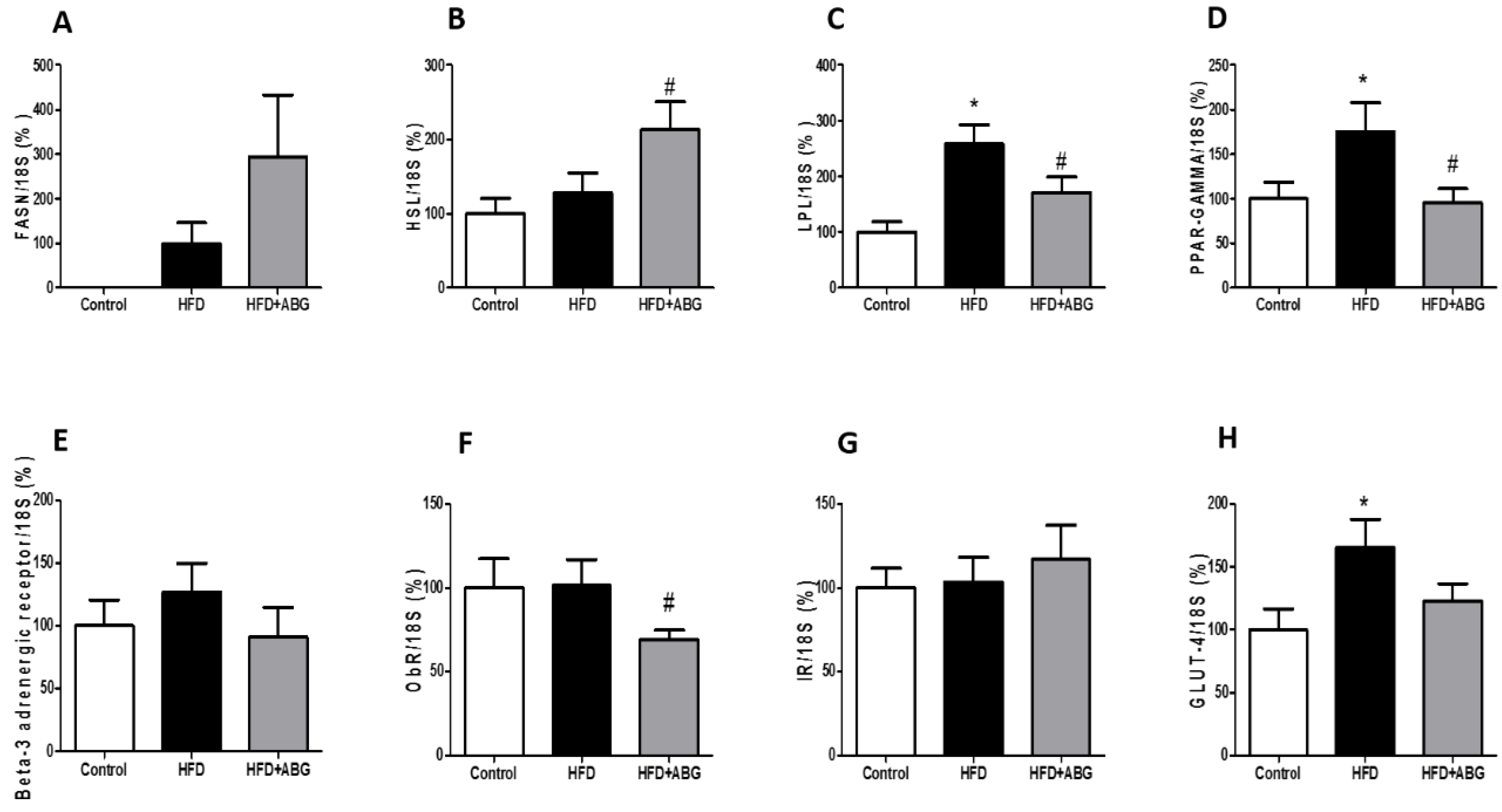
3.2.7. FASN, HSL, LPL, PPAR-γ, ObR, InsR, GLUT-4, β3-ADR, IL-1β, IL-6, TNF-α and NOX-4 mRNA Levels in Visceral and Subcutaneous Adipose Tissue
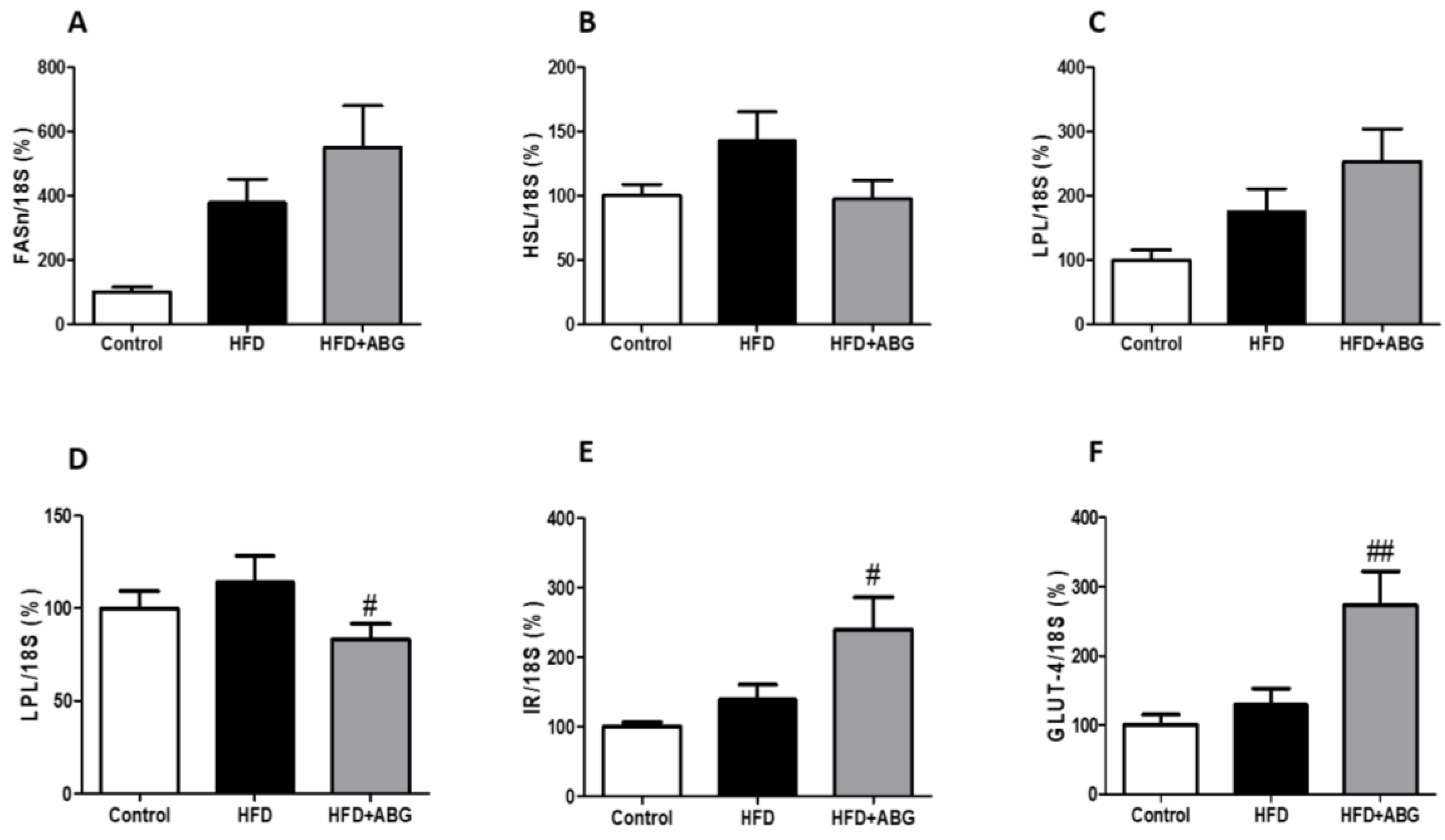
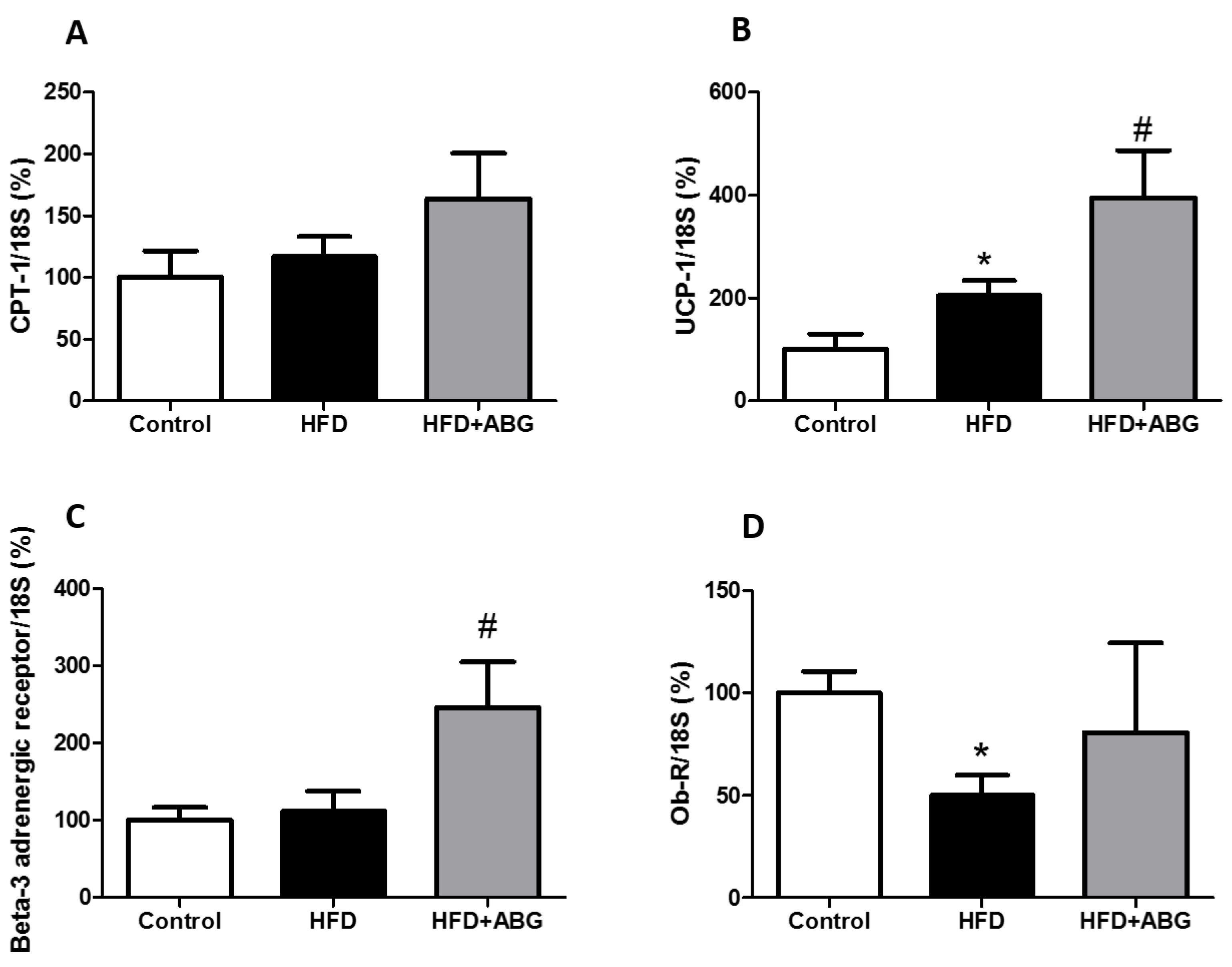
3.2.8. FASN, HSL, LPL, PPAR-γ, β3-ADR, ObR, InsR, GLUT-4, CPT-1 and UCP-1 mRNA Levels in Brown Adipose Tissue
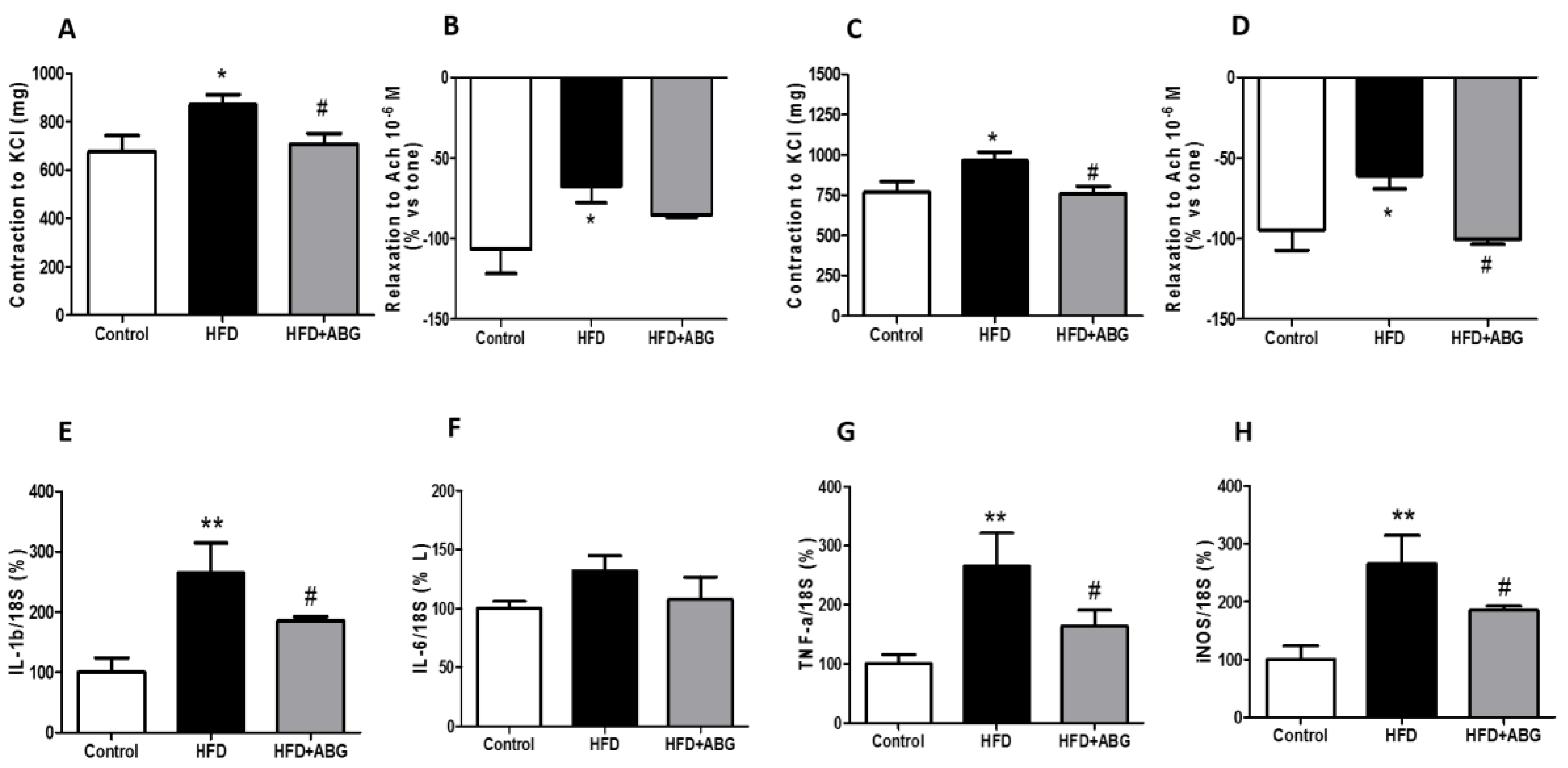
3.2.9. Vascular Response of Aortic Rings to Potassium Chloride and Acetylcholine
3.2.10. mRNA levels of IL-1β, IL-6, TNF-α and iNOS in Perivascular Adipose Tissue
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Ferrante, A.W., Jr. Macrophages, fat, and the emergence of immunometabolism. J. Clin. Investig. 2013, 123, 4992–4993. [Google Scholar] [CrossRef] [PubMed]
- Pirola, L.; Ferraz, J.C. Role of pro- and anti-inflammatory phenomena in the physiopathology of type 2 diabetes and obesity. World J. Biol. Chem. 2017, 8, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.G.; Park, C.Y. Anti-Obesity Drugs: A Review about Their Effects and Safety. Diabetes Metab. J. 2012, 36, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Tung, Y.C.; Pan, M.H.; Su, N.W.; Lai, Y.J.; Cheng, K.C. Black garlic: A critical review of its production, bioactivity, and application. J. Food Drug Anal. 2017, 25, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.H.; Kang, D. Physicochemical Properties, Biological Activity, Health Benefits, and General Limitations of Aged Black Garlic: A Review. Molecules 2017, 22, 919. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.E.; Cho, S.Y.; Won, Y.D.; Lee, S.H.; Park, H.J. A comparative study of the different analytical methods for analysis of S-allyl cysteine in black garlic by HPLC. LWT-Food Sci. Technol. 2012, 46, 532–535. [Google Scholar] [CrossRef]
- Lindenmeier, M.; Faist, V.; Hofmann, T. Structural and functional characterization of pronyl-lysine, a novel protein modification in bread crust melanoidins showing in vitro antioxidative and phase I/II enzyme modulating activity. J. Agric. Food Chem. 2002, 50, 6997–7006. [Google Scholar] [CrossRef]
- Moreira, A.S.; Nunes, F.M.; Domingues, M.R.; Coimbra, M.A. Coffee melanoidins: Structures, mechanisms of formation and potential health impacts. Food Funct. 2012, 3, 903–915. [Google Scholar] [CrossRef]
- Borrelli, R.C.; Visconti, A.; Mennella, C.; Anese, M.; Fogliano, V. Chemical characterization and antioxidant properties of coffee melanoidins. J. Agric. Food Chem. 2002, 50, 6527–6533. [Google Scholar] [CrossRef]
- Wang, Y.; Ho, C.T. Polyphenolic chemistry of tea and coffee: A century of progress. J. Agric. Food Chem. 2009, 57, 8109–8114. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.G.; Kang, M.J.; Hong, S.S.; Choi, Y.H.; Shin, J.H. Antiinflammatory Effects of Functionally Active Compounds Isolated from Aged Black Garlic. Phytother. Res. 2017, 31, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Nillert, N.; Pannangrong, W.; Welbat, J.U.; Chaijaroonkhanarak, W.; Sripanidkulchai, K.; Sripanidkulchai, B. Neuroprotective Effects of Aged Garlic Extract on Cognitive Dysfunction and Neuroinflammation Induced by beta-Amyloid in Rats. Nutrients 2017, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Jikihara, H.; Qi, G.; Nozoe, K.; Hirokawa, M.; Sato, H.; Sugihara, Y.; Shimamoto, F. Aged garlic extract inhibits 1,2-dimethylhydrazine-induced colon tumor development by suppressing cell proliferation. Oncol. Rep. 2015, 33, 1131–1140. [Google Scholar] [CrossRef] [PubMed]
- Perez-Torres, I.; Torres-Narvaez, J.C.; Pedraza-Chaverri, J.; Rubio-Ruiz, M.E.; Diaz-Diaz, E.; Del Valle-Mondragon, L.; Martinez-Memije, R.; Varela Lopez, E.; Guarner-Lans, V. Effect of the Aged Garlic Extract on Cardiovascular Function in Metabolic Syndrome Rats. Molecules 2016, 21, 1425. [Google Scholar] [CrossRef]
- Baluchnejadmojarad, T.; Kiasalari, Z.; Afshin-Majd, S.; Ghasemi, Z.; Roghani, M. S-allyl cysteine ameliorates cognitive deficits in streptozotocin-diabetic rats via suppression of oxidative stress, inflammation, and acetylcholinesterase. Eur. J. Pharmacol. 2017, 794, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.M.; Sok, D.E.; Kim, M.R. Anti-allergic action of aged black garlic extract in RBL-2H3 cells and passive cutaneous anaphylaxis reaction in mice. J. Med. Food. 2014, 17, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Villalón, A.L.; Amor, S.; Monge, L.; Fernández, N.; Prodanov, M.; Muñoz, M.; Inarejos-García, A.M.; Granado, M. In vitro studies of an aged black garlic extract enriched in S-allylcysteine and polyphenols with cardioprotective effects. J. Funct. Foods 2016, 27, 11. [Google Scholar] [CrossRef]
- Shin, J.H.; Lee, C.W.; Oh, S.J.; Yun, J.; Kang, M.R.; Han, S.B.; Park, H.; Jung, J.C.; Chung, Y.H.; Kang, J.S. Hepatoprotective effect of aged black garlic extract in rodents. Toxicol. Res. 2014, 30, 49–54. [Google Scholar] [CrossRef]
- Ha, A.W.; Ying, T.; Kim, W.K. The effects of black garlic (Allium satvium) extracts on lipid metabolism in rats fed a high fat diet. Nutr. Res. Pract. 2015, 9, 30–36. [Google Scholar] [CrossRef]
- Kim, J.H.; Yu, S.H.; Cho, Y.J.; Pan, J.H.; Cho, H.T.; Kim, J.H.; Bong, H.; Lee, Y.; Chang, M.H.; Jeong, Y.J.; et al. Preparation of S-Allylcysteine-Enriched Black Garlic Juice and Its Antidiabetic Effects in Streptozotocin-Induced Insulin-Deficient Mice. J. Agric. Food Chem. 2017, 65, 358–363. [Google Scholar] [CrossRef]
- Lee, Y.M.; Gweon, O.C.; Seo, Y.J.; Im, J.; Kang, M.J.; Kim, M.J.; Kim, J.I. Antioxidant effect of garlic and aged black garlic in animal model of type 2 diabetes mellitus. Nutr. Res. Pract. 2009, 3, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.S.; Park, S.H.; Choi, E.K.; Ryu, B.H.; Park, B.H.; Kim, D.S.; Kim, Y.G.; Chae, S.W. Reduction of blood lipid parameters by a 12-wk supplementation of aged black garlic: A randomized controlled trial. Nutrition 2014, 30, 1034–1039. [Google Scholar] [CrossRef] [PubMed]
- Kang, O.J. Evaluation of Melanoidins Formed from Black Garlic after Different Thermal Processing Steps. Prev. Nutr. Food Sci. 2016, 21, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Haffner, S.M.; Stern, M.P.; Hazuda, H.P.; Pugh, J.A.; Patterson, J.K. Hyperinsulinemia in a population at high risk for non-insulin-dependent diabetes mellitus. N. Engl. J. Med. 1986, 315, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Roza, N.A.; Possignolo, L.F.; Palanch, A.C.; Gontijo, J.A. Effect of long-term high-fat diet intake on peripheral insulin sensibility, blood pressure, and renal function in female rats. Food Nutr. Res. 2016, 60, 28536. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P. A reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. Biotechniques 1993, 15, 532–534, 536–537. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Colin-Gonzalez, A.L.; Santana, R.A.; Silva-Islas, C.A.; Chanez-Cardenas, M.E.; Santamaria, A.; Maldonado, P.D. The antioxidant mechanisms underlying the aged garlic extract- and S-allylcysteine-induced protection. Oxid. Med. Cell Longev. 2012, 2012, 907162. [Google Scholar] [CrossRef] [PubMed]
- Ilkun, O.; Boudina, S. Cardiac dysfunction and oxidative stress in the metabolic syndrome: An update on antioxidant therapies. Curr. Pharm. Des. 2013, 19, 4806–4817. [Google Scholar] [CrossRef]
- Xia, X.; Weng, J. Targeting metabolic syndrome: Candidate natural agents. J. Diabetes 2010, 2, 243–249. [Google Scholar] [CrossRef]
- Zhai, B.; Zhang, C.; Sheng, Y.; Zhao, C.; He, X.; Xu, W.; Huang, K.; Luo, Y. Hypoglycemic and hypolipidemic effect of S-allyl-cysteine sulfoxide (alliin) in DIO mice. Sci. Rep. 2018, 8, 3527. [Google Scholar] [CrossRef] [PubMed]
- Lembede, B.W.; Erlwanger, K.H.; Nkomozepi, P.; Chivandi, E. Effect of neonatal orally administered S-allyl cysteine in high-fructose diet fed Wistar rats. J. Dev. Orig. Health Dis. 2018, 9, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Seo, D.Y.; Lee, S.; Figueroa, A.; Kwak, Y.S.; Kim, N.; Rhee, B.D.; Ko, K.S.; Bang, H.S.; Baek, Y.H.; Han, J. Aged garlic extract enhances exercise-mediated improvement of metabolic parameters in high fat diet-induced obese rats. Nutr. Res. Pract. 2012, 6, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.M.; Lee, S.H.; Lee, D.S.; You, M.J.; Chung, I.K.; Cheon, W.H.; Kwon, Y.S.; Lee, Y.J.; Ku, S.K. Fermented garlic protects diabetic, obese mice when fed a high-fat diet by antioxidant effects. Nutr. Res. 2011, 31, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Atkin, M.; Laight, D.; Cummings, M.H. The effects of garlic extract upon endothelial function, vascular inflammation, oxidative stress and insulin resistance in adults with type 2 diabetes at high cardiovascular risk. A pilot double blind randomized placebo controlled trial. J. Diabetes Complic. 2016, 30, 723–727. [Google Scholar] [CrossRef]
- Gomez-Arbelaez, D.; Lahera, V.; Oubina, P.; Valero-Munoz, M.; de Las Heras, N.; Rodriguez, Y.; Garcia, R.G.; Camacho, P.A.; Lopez-Jaramillo, P. Aged garlic extract improves adiponectin levels in subjects with metabolic syndrome: A double-blind, placebo-controlled, randomized, crossover study. Mediat. Inflamm. 2013, 2013, 285795. [Google Scholar] [CrossRef] [PubMed]
- Seo, D.Y.; Kwak, H.B.; Lee, S.R.; Cho, Y.S.; Song, I.S.; Kim, N.; Bang, H.S.; Rhee, B.D.; Ko, K.S.; Park, B.J.; et al. Effects of aged garlic extract and endurance exercise on skeletal muscle FNDC-5 and circulating irisin in high-fat-diet rat models. Nutr. Res. Pract. 2014, 8, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Miki, S.; Inokuma, K.I.; Takashima, M.; Nishida, M.; Sasaki, Y.; Ushijima, M.; Suzuki, J.I.; Morihara, N. Aged garlic extract suppresses the increase of plasma glycated albumin level and enhances the AMP-activated protein kinase in adipose tissue in TSOD mice. Mol. Nutr. Food Res. 2017, 61, 1600797. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Bielohuby, M.; Fleming, T.; Grabner, G.F.; Foppen, E.; Bernhard, W.; Guzman-Ruiz, M.; Layritz, C.; Legutko, B.; Zinser, E.; et al. Dietary sugars, not lipids, drive hypothalamic inflammation. Mol. Metab. 2017, 6, 897–908. [Google Scholar] [CrossRef]
- Ibars, M.; Ardid-Ruiz, A.; Suarez, M.; Muguerza, B.; Blade, C.; Aragones, G. Proanthocyanidins potentiate hypothalamic leptin/STAT3 signalling and Pomc gene expression in rats with diet-induced obesity. Int. J. Obes. 2017, 41, 129–136. [Google Scholar] [CrossRef]
- Ishino, S.; Sugita, T.; Kondo, Y.; Okai, M.; Tsuchimori, K.; Watanabe, M.; Mori, I.; Hosoya, M.; Horiguchi, T.; Kamiguchi, H. Glucose uptake of the muscle and adipose tissues in diabetes and obesity disease models: Evaluation of insulin and beta3-adrenergic receptor agonist effects by (18)F-FDG. Ann. Nucl. Med. 2017, 31, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Mark, A.L.; Rahmouni, K.; Correia, M.; Haynes, W.G. A leptin-sympathetic-leptin feedback loop: Potential implications for regulation of arterial pressure and body fat. Acta Physiol. Scand. 2003, 177, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Pirzgalska, R.M.; Pereira, M.M.; Kubasova, N.; Barateiro, A.; Seixas, E.; Lu, Y.H.; Kozlova, A.; Voss, H.; Martins, G.G.; et al. Sympathetic neuro-adipose connections mediate leptin-driven lipolysis. Cell 2015, 163, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Portillo, M.P.; Simon, E.; Garcia-Calonge, M.A.; Del Barrio, A.S. Effect of high-fat diet on lypolisis in isolated adipocytes from visceral and subcutaneous WAT. Eur. J. Nutr. 1999, 38, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Zhang, F.; Zhao, M.X.; Ren, X.S.; Chen, Q.; Li, Y.H.; Kang, Y.M.; Zhu, G.Q. Reduced lipolysis response to adipose afferent reflex involved in impaired activation of adrenoceptor-cAMP-PKA-hormone sensitive lipase pathway in obesity. Sci. Rep. 2016, 6, 34374. [Google Scholar] [CrossRef]
- Fu, J.; Zeng, C.; Zeng, Z.; Wang, B.; Wen, X.; Yu, P.; Gong, D. Cinnamomum camphora Seed Kernel Oil Improves Lipid Metabolism and Enhances beta3-Adrenergic Receptor Expression in Diet-Induced Obese Rats. Lipids 2016, 51, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Lu, K.; Wen, X.Y.; Liu, H.; Chen, A.P.; Xu, M.W.; Zhang, H.; Yu, J. Jueming Prescription reduces body weight by increasing the mRNA expressions of beta3-adrenergic receptor and uncoupling protein-2 in adipose tissue of diet-induced obese rats. Chin. J. Integr. Med. 2012, 18, 775–781. [Google Scholar] [CrossRef]
- Mead, J.R.; Irvine, S.A.; Ramji, D.P. Lipoprotein lipase: Structure, function, regulation, and role in disease. J. Mol. Med. 2002, 80, 753–769. [Google Scholar] [CrossRef]
- Park, J.-A. Inhibition of Adipocyte Differentiation and Adipogenesis by Aged Black Garlic Extracts in 3T3-L1 Preadipocytes. J. Life Sci. 2011, 21, 720–728. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, H.K. Effect of garlic on high fat induced obesity. Acta Biol. Hung 2011, 62, 244–254. [Google Scholar] [CrossRef]
- Ahmadi, N.; Nabavi, V.; Hajsadeghi, F.; Zeb, I.; Flores, F.; Ebrahimi, R.; Budoff, M. Aged garlic extract with supplement is associated with increase in brown adipose, decrease in white adipose tissue and predict lack of progression in coronary atherosclerosis. Int. J. Cardiol. 2013, 168, 2310–2314. [Google Scholar] [CrossRef] [PubMed]
- Bao, B.; Chen, Y.G.; Zhang, L.; Na Xu, Y.L.; Wang, X.; Liu, J.; Qu, W. Momordica charantia (Bitter Melon) reduces obesity-associated macrophage and mast cell infiltration as well as inflammatory cytokine expression in adipose tissues. PLoS ONE 2013, 8, e84075. [Google Scholar] [CrossRef] [PubMed]
- Sampey, B.P.; Vanhoose, A.M.; Winfield, H.M.; Freemerman, A.J.; Muehlbauer, M.J.; Fueger, P.T.; Newgard, C.B.; Makowski, L. Cafeteria diet is a robust model of human metabolic syndrome with liver and adipose inflammation: Comparison to high-fat diet. Obesity 2011, 19, 1109–1117. [Google Scholar] [CrossRef] [PubMed]
- Alarcon, G.; Roco, J.; Medina, M.; Medina, A.; Peral, M.; Jerez, S. High fat diet-induced metabolically obese and normal weight rabbit model shows early vascular dysfunction: Mechanisms involved. Int. J. Obes. 2018, 42, 1535–1543. [Google Scholar] [CrossRef] [PubMed]
- Nosalski, R.; Guzik, T.J. Perivascular adipose tissue inflammation in vascular disease. Br. J. Pharmacol. 2017, 174, 3496–3513. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TR (min) | Compound | ESI + (m/z) | MM (Da) | Chemical Structure | Concentration (mg/g) |
|---|---|---|---|---|---|
| 3.9 | γ-L-glutamyl-S-methyl-L-cysteine (GSMC) | 265 | 264 |  | 0.00 ± 0.00 |
| 4.5 | (+)-S-methyl-L-cysteine sulfoxide (methiin) | 104 | 103 |  | 0.00 ± 0.00 |
| 4.9 | (+)-S-allyl-L-cysteine sulfoxide (alliin) | 178 | 177 |  | 0.03 ± 0.00 |
| 5.2 | (+)-S-(trans-1-propenyl)-L-cysteine sulfoxide (iso-alliin) | 178 | 177 |  | 0.01 ± 0.00 |
| 7.41 | S-allyl cysteine (SAC) | 162 | 161 |  | 1.15 ± 0.02 |
| 8.8 | iso-S-allyl-L-cystein (iso-SAC) | 162 | 161 | 0.04 ± 0.00 | |
| 10.9 | γ-L-glutamyl-S-allyl-L-cysteine (GSAC) | 291 | 290 |  | 0.00 ± 0.00 |
| 11.6 | γ-L-glutamyl-S-(trans-1-propenyl)-L-cysteine (GSPC) | 291 | 290 |  | 0.00 ± 0.00 |
| Control | HFD | HFD + ABG | |
|---|---|---|---|
| Heart (g) | 1.58 ± 0.03 | 1.72 ±0.02 | 1.85 ± 0.07 |
| Kidneys (g) | 2.33 ± 0.08 | 2.36 ± 0.10 | 2.30 ± 0.05 |
| Adrenal glands (mg) | 48.5 ± 1.6 | 48.3 ± 1.7 | 51.6 ± 2.2 |
| Spleen (g) | 0.84 ± 0.04 | 0.82 ± 0.05 | 0.88 ± 0.04 |
| Liver (g) | 9.79 ± 0.17 | 10.32 ± 0.44 | 10.27 ± 0.47 |
| Epidydimal visceral fat (g) | 5.31 ± 0.28 | 10.24 ± 0.97 *** | 10.00 ± 0.75 |
| Lumbar subcutaneous fat (g) | 2.41 ±0.13 | 4.62 ± 0.33 *** | 3.84 ± 0.21 # |
| Brown adipose tissue (g) | 0.67 ± 0.06 | 1.20 ± 0.05 *** | 0.88 ± 0.03 ### |
| Periaortic adipose tissue (g) | 0.057 ± 0.006 | 0.113 ± 0.015 ** | 0.111 ± 0.016 |
| Soleus (g) | 0.18 ± 0.04 | 0.20 ± 0.07 | 0.21 ± 0.01 |
| Gastrocnemius (g) | 2.28 ± 0.05 | 2.52 ± 0.07 | 2.50 ± 0.07 |
| Glycemia (mg/dL) | 67 ± 2 | 63.5 ± 2 | 63.6 ± 3 |
| Total Cholesterol (mg/dL) | 161 ± 8 | 187 ± 10 * | 180 ± 9 |
| LDL-cholesterol (mg/dL) | 86 ± 3 | 94 ± 3 * | 79 ± 6 # |
| HDL-cholesterol (mg/dL) | 111 ± 4 | 74 ± 6 ** | 108 ± 7 ## |
| Tryglycerides (mg/dL) | 105 ± 9 | 162 ± 16 ** | 127 ± 12 # |
| Insulin (ng/mL) | 3.5 ± 0.5 | 5.5 ± 0.6 * | 3.5 ± 0.4 # |
| Leptin (ng/mL) | 6.4 ± 3.2 | 19.4 ± 3 ** | 9.4 ± 1.6 # |
| Adiponectin (mg/dL) | 104 ± 5 | 109 ± 9 | 151 ± 13 # |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amor, S.; González-Hedström, D.; Martín-Carro, B.; Inarejos-García, A.M.; Almodóvar, P.; Prodanov, M.; García-Villalón, A.L.; Granado, M. Beneficial Effects of an Aged Black Garlic Extract in the Metabolic and Vascular Alterations Induced by a High Fat/Sucrose Diet in Male Rats. Nutrients 2019, 11, 153. https://doi.org/10.3390/nu11010153
Amor S, González-Hedström D, Martín-Carro B, Inarejos-García AM, Almodóvar P, Prodanov M, García-Villalón AL, Granado M. Beneficial Effects of an Aged Black Garlic Extract in the Metabolic and Vascular Alterations Induced by a High Fat/Sucrose Diet in Male Rats. Nutrients. 2019; 11(1):153. https://doi.org/10.3390/nu11010153
Chicago/Turabian StyleAmor, Sara, Daniel González-Hedström, Beatriz Martín-Carro, Antonio Manuel Inarejos-García, Paula Almodóvar, Marin Prodanov, Angel Luis García-Villalón, and Miriam Granado. 2019. "Beneficial Effects of an Aged Black Garlic Extract in the Metabolic and Vascular Alterations Induced by a High Fat/Sucrose Diet in Male Rats" Nutrients 11, no. 1: 153. https://doi.org/10.3390/nu11010153
APA StyleAmor, S., González-Hedström, D., Martín-Carro, B., Inarejos-García, A. M., Almodóvar, P., Prodanov, M., García-Villalón, A. L., & Granado, M. (2019). Beneficial Effects of an Aged Black Garlic Extract in the Metabolic and Vascular Alterations Induced by a High Fat/Sucrose Diet in Male Rats. Nutrients, 11(1), 153. https://doi.org/10.3390/nu11010153