Effect of the Strawberry Genotype, Cultivation and Processing on the Fra a 1 Allergen Content


Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material
2.3. Heterologous Expression and Purification of rFra a 1.02 Protein
2.4. Protein Determination
2.5. Production of Polyclonal Anti-Fra a 1.02 Antibody
2.6. Thermal Treatment of rFra a 1.02
2.7. Processing of Strawberry Fruits
2.8. Strawberry Extracts
2.9. Temperature Stability of Protein Extracts
2.10. Indirect Competitive ELISA
2.11. Statistical Analysis
3. Results
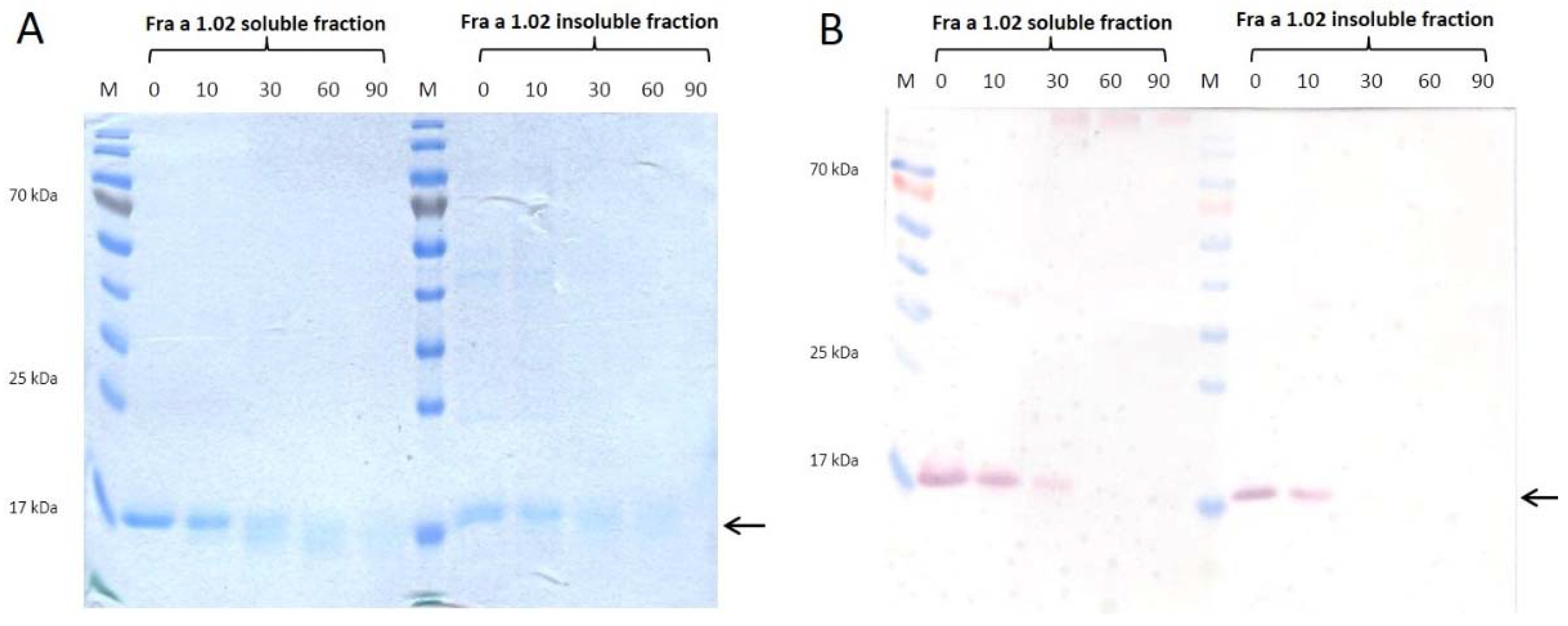
3.1. Purification and Thermal Stability of Recombinant Fra a 1.02 Protein from Soluble and Insoluble Fraction
3.2. Establishment and Validation of the ELISA
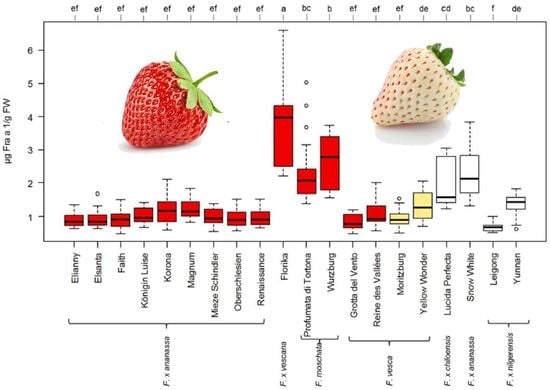
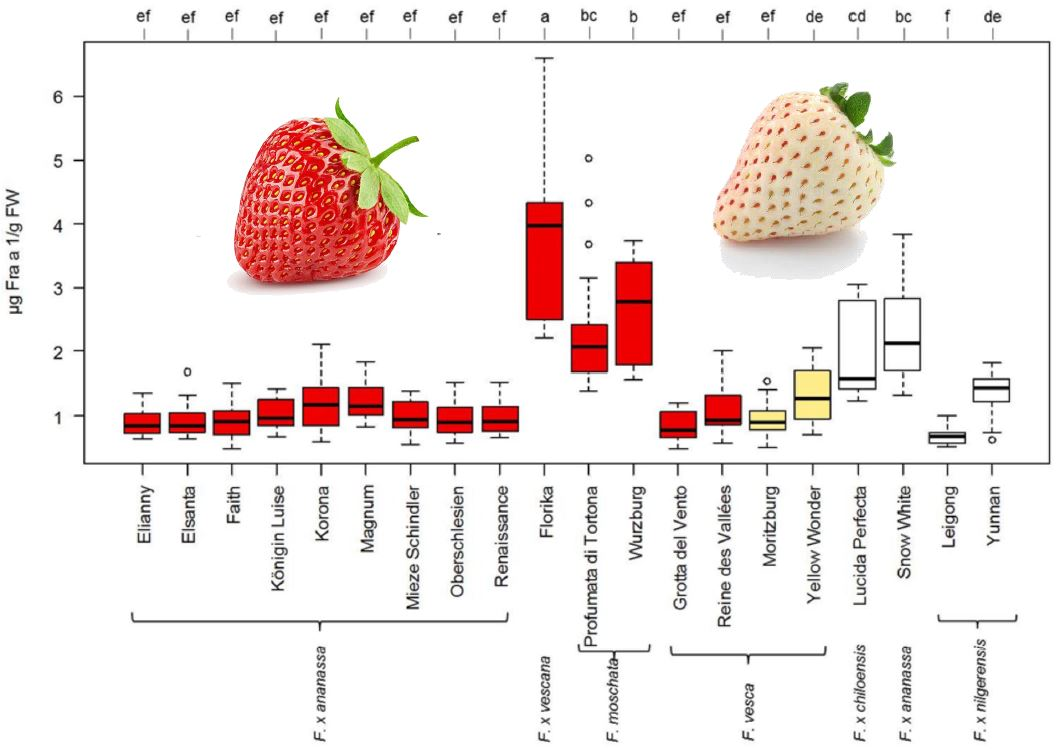
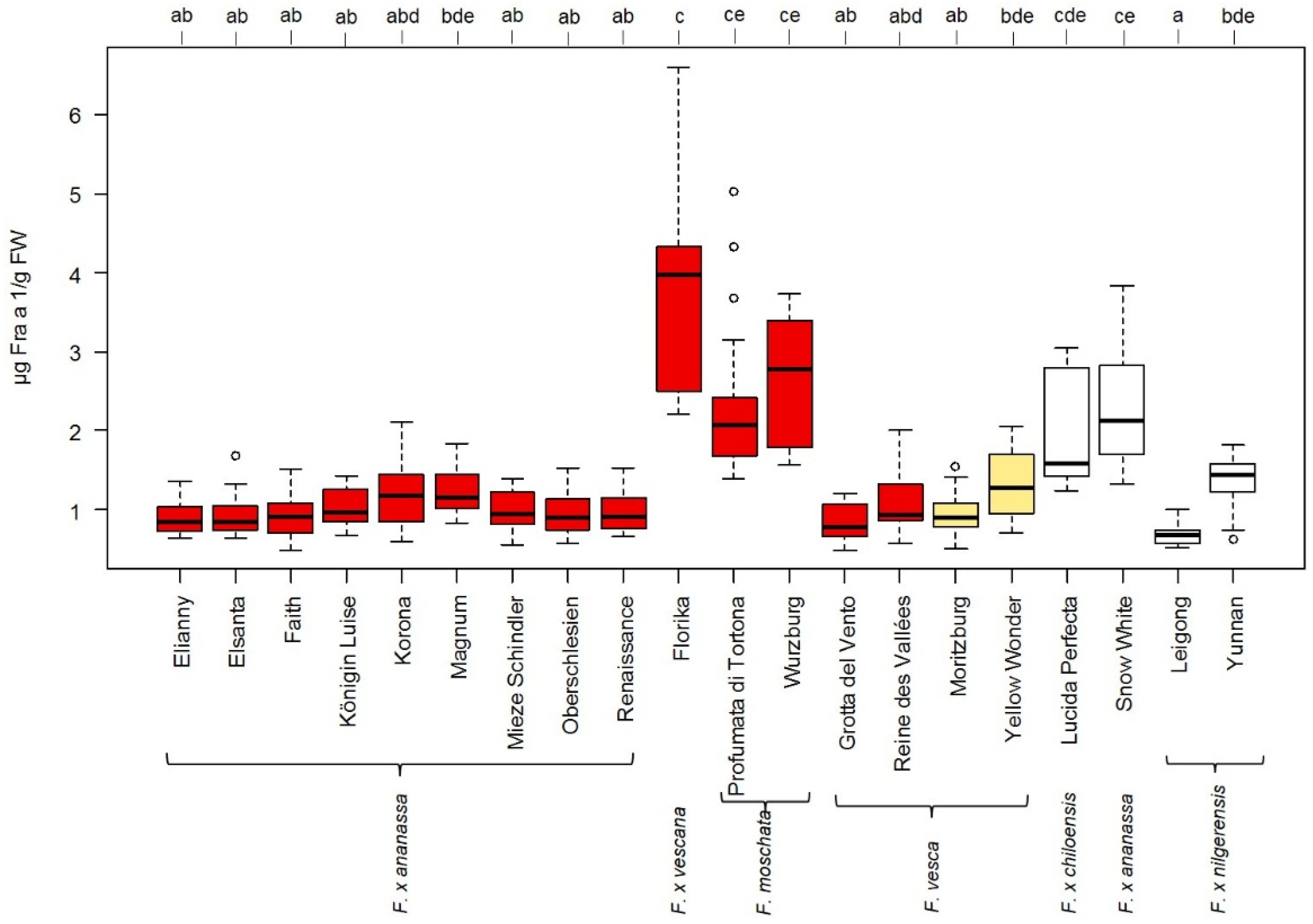
3.3. Influence of Strawberry Variety on Fra a 1 Content
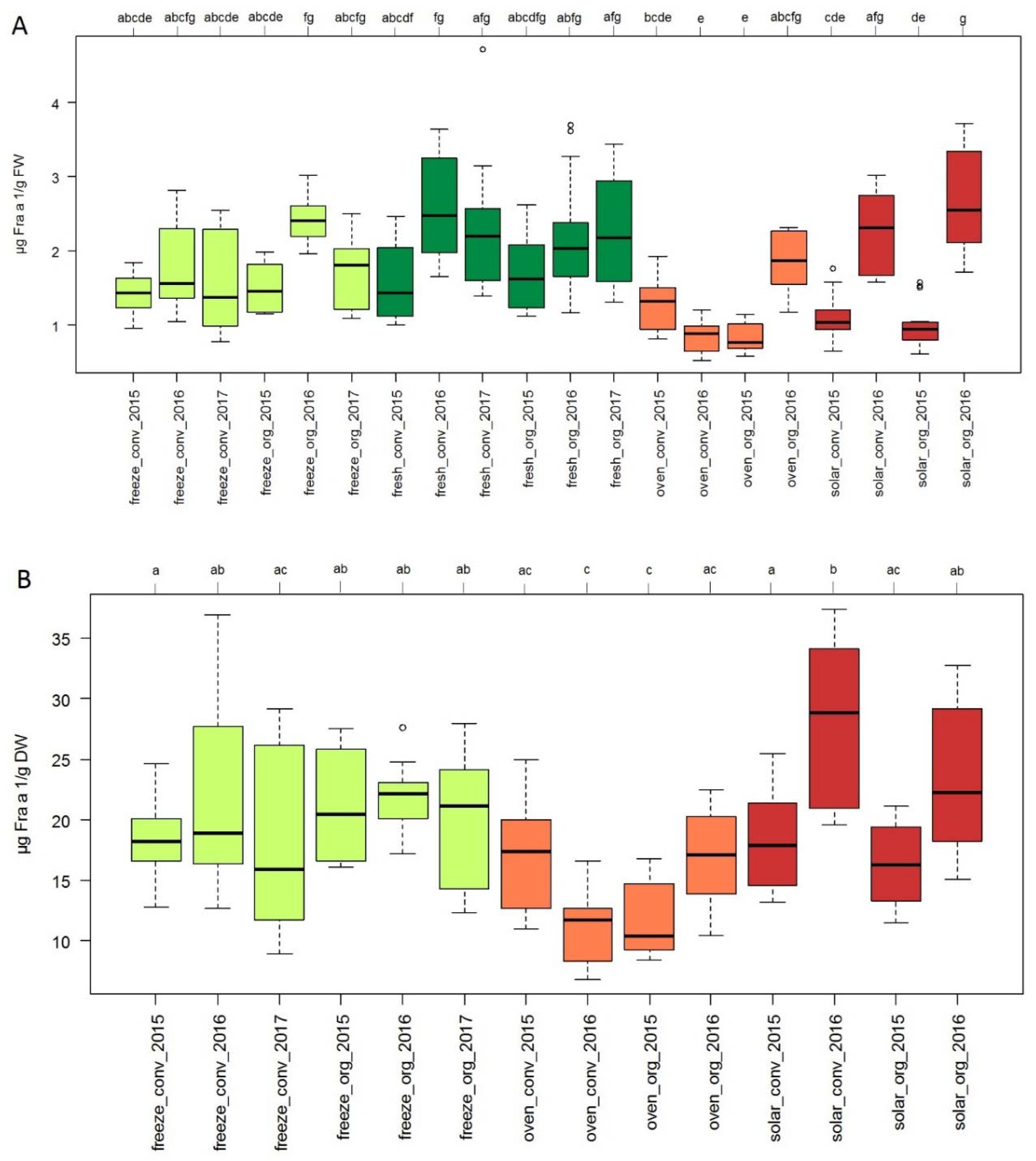
3.4. Effect of Cultivation and Processing Methods on Allergen Content in Strawberry Fruits
3.5. Temperature Stability of Native Fra a 1 in Strawberry Protein Extracts
4. Discussion
4.1. Biochemical Properties and Thermal Stability of Recombinant Fra a 1.02
4.2. Fra a 1 Content of Strawberry Fruits Is Genotype Dependent
4.3. Cultivation Condition and Drying Processing of Strawberries has Minor Effect on Fra a 1 Content
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Giampieri, F.; Tulipani, S.; Alvarez-Suarez, J.M.; Quiles, J.L.; Mezzetti, B.; Battino, M. The strawberry: Composition, nutritional quality, and impact on human health. Nutrition 2012, 28, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Afrin, S.; Gasparrini, M.; Forbes-Hernandez, T.Y.; Reboredo-Rodriguez, P.; Mezzetti, B.; Varela-López, A.; Giampieri, F.; Battino, M. Promising Health Benefits of the Strawberry: A Focus on Clinical Studies. J. Agric. Food Chem. 2016, 64, 4435–4449. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Rhone, M.; Lyons, T.J. Berries: Emerging impact on cardiovascular health. Nutr. Rev. 2010, 68, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Adams, L.S.; Zhang, Y.; Lee, R.; Sand, D.; Scheuller, H.S.; Heber, D. Blackberry, black raspberry, blueberry, cranberry, red raspberry, and strawberry extracts inhibit growth and stimulate apoptosis of human cancer cells in vitro. J. Agric. Food Chem. 2006, 54, 9329–9339. [Google Scholar] [CrossRef] [PubMed]
- Sicherer, S.H.; Sampson, H.A. Food allergy. J. Allergy Clin. Immunol. 2010, S116–S125. [Google Scholar] [CrossRef] [PubMed]
- Dölle, S.; Schwarz, D.; Lehmann, K.; Weckwerth, W.; George, E.; Worm, M.; Franken, P. Tomato allergy: impact of genotype and environmental factors on the biological response. J. Sci. Food Agric. 2011, 91, 2234–2240. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, A.L.; Alm, R.; Ekstrand, B.; Fjelkner-Modig, S.; Schiött, Å.; Bengtsson, U.; Björk, L.; Hjernø, K.; Roepstorff, P.; Emanuelsson, C.S. Bet v 1 homologues in strawberry identified as IgE-binding proteins and presumptive allergens. Allergy 2004, 59, 1277–1284. [Google Scholar] [CrossRef] [PubMed]
- Sinha, M.; Singh, R.P.; Kushwaha, G.S.; Iqbal, N.; Singh, A.; Kaushik, S.; Kaur, P.; Sharma, S.; Singh, T.P. Current Overview of Allergens of Plant Pathogenesis Related Protein Families. Sci. World J. 2014, 2014, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Ebner, C.; Hirschwehr, R.; Bauer, L.; Breiteneder, H.; Valenta, R.; Ebner, H.; Kraft, D.; Scheiner, O. Identification of allergens in fruits and vegetables: IgE cross-reactivities with the important birch pollen allergens Bet v 1 and Bet v 2 (birch profilin). J. Allergy Clin. Immunol. 1995, 96, 962–969. [Google Scholar] [CrossRef]
- Breiteneder, H.; Ebner, C. Molecular and biochemical classification of plant-derived food allergens. J. Allergy Clin. Immunol. 2000, 106, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann-Sommergruber, K. Plant Allergens and Pathogenesis-Related Proteins: What Do They Have in Common? Int. Arch. Allergy Immunol. 2000, 122, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Franz-Oberdorf, K.; Eberlein, B.; Edelmann, K.; Bleicher, P.; Kurze, E.; Helm, D.; Olbricht, K.; Darsow, U.; Ring, J.; Schwab, W. White-fruited strawberry genotypes are not per se hypoallergenic. Food Res. Int. 2017, 100, 748–756. [Google Scholar] [CrossRef] [PubMed]
- Darrow, G.M. The Strawberry: History, Breeding and Physiology; Holt, Rinehart and Winston: New York, NY, USA, 1966. [Google Scholar]
- Ulrich, D.; Komes, D.; Olbricht, K.; Hoberg, E. Diversity of aroma patterns in wild and cultivated Fragaria accessions. Genet. Resour. Crop Evol. 2007, 54, 1185–1196. [Google Scholar] [CrossRef]
- Zuidmeer, L.; Salentijn, E.; Rivas, M.F.; Mancebo, E.G.; Asero, R.; Matos, C.I.; Pelgrom, K.T.B.; Gilissen, L.J.W.J.; van Ree, R. The role of profilin and lipid transfer protein in strawberry allergy in the Mediterranean area. Clin. Exp. Allergy 2006, 36, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Franz-Oberdorf, K.; Eberlein, B.; Edelmann, K.; Hücherig, S.; Besbes, F.; Darsow, U.; Ring, J.; Schwab, W. Fra a 1.02 Is the Most Potent Isoform of the Bet v 1-like Allergen in Strawberry Fruit. J. Agric. Food Chem. 2016, 64, 3688–3696. [Google Scholar] [CrossRef] [PubMed]
- Seutter von Loetzen, C.; Schweimer, K.; Schwab, W.; Rösch, P.; Hartl-Spiegelhauer, O. Solution structure of the strawberry allergen Fra a 1. Biosci. Rep. 2012, 32, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Casañal, A.; Zander, U.; Muñoz, C.; Dupeux, F.; Luque, I.; Botella, M.A.; Schwab, W.; Valpuesta, V.; Marquez, J.A. The Strawberry Pathogenesis-related 10 (PR-10) Fra a Proteins Control Flavonoid Biosynthesis by Binding to Metabolic Intermediates. J. Biol. Chem. 2013, 288, 35322–35332. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, C.; Hoffmann, T.; Escobar, N.M.; Ludemann, F.; Botella, M.A.; Valpuesta, V.; Schwab, W. The Strawberry Fruit Fra a Allergen Functions in Flavonoid Biosynthesis. Mol. Plant 2010, 3, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Matthes, A.; Schmitz-Eiberger, M. Apple (Malus domestica L. Borkh.) Allergen Mal d 1: Effect of Cultivar, Cultivation System, and Storage Conditions. J. Agric. Food Chem. 2009, 57, 10548–10553. [Google Scholar] [CrossRef] [PubMed]
- Marzban, G.; Puehringer, H.; Dey, R.; Brynda, S.; Ma, Y.; Martinelli, A.; Zaccarini, M.; van der Weg, E.; Housley, Z.; Kolarich, D.; et al. Localisation and distribution of the major allergens in apple fruits. Plant Sci. 2005, 169, 387–394. [Google Scholar] [CrossRef]
- Franz-Oberdorf, K.; Langer, A.; Strasser, R.; Isono, E.; Ranftl, Q.L.; Wunschel, C.; Schwab, W. Physical interaction between the strawberry allergen Fra a 1 and an associated partner FaAP: Interaction of Fra a 1 proteins and FaAP. Proteins 2017, 85, 1891–1901. [Google Scholar] [CrossRef] [PubMed]
- Vassilopoulou, E.V.; Zuidmeer, L.; Akkerdaas, J.; Rigby, N.; Moreno, F.J.; Papadopoulos, N.G.; Saxoni-Papageorgiou, P.; Mills, C.; van Ree, R. Optimized techniques for the extraction of grape allergens appropriate for in vivo and in vitro testing and diagnosis. Mol. Nutr. Food Res. 2007, 51, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Vieths, S.; Scheurer, S.; Ballmer-Weber, B. Current Understanding of Cross-Reactivity of Food Allergens and Pollen. Ann. N. Y. Acad. Sci. 2002, 964, 47–68. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, R.; Mageney, V.; Schwefel, K.; Vollmers, D.; Krüger, A.; Horn, R. Genotyping of red and white fruited strawberry (Fragaria L.) accessions and hybrids based on microsatellite markers and on the genetic diversity in the allergen genes fra a 1 and fra a 3. Genet. Resour. Crop Evol. 2016, 63, 1203–1217. [Google Scholar] [CrossRef]
- Eriksson, N.E.; Möller, C.; Werner, S.; Magnusson, J.; Bengtsson, U.; Zolubas, M. Self-Reported Food Hypersensitivity in Sweden, Denmark, Estonia, Lithuania, and Russia. J. Investig. Allergol. Clin. Immunol. 2004, 14, 70–79. [Google Scholar] [PubMed]
- Musidlowska-Persson, A.; Alm, R.; Emanuelsson, C. Cloning and sequencing of the Bet v 1-homologous allergen Fra a 1 in strawberry (Fragaria ananassa) shows the presence of an intron and little variability in amino acid sequence. Mol. Immunol. 2007, 44, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Wurm, D.J.; Quehenberger, J.; Mildner, J.; Eggenreich, B.; Slouka, C.; Schwaighofer, A.; Wieland, K.; Lendl, B.; Rajamanickam, V.; Herwig, C. Teaching an old pET new tricks: tuning of inclusion body formation and properties by a mixed feed system in E. coli. Appl. Microbiol. Biotechnol. 2018, 102, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Karamloo, F.; Scheurer, S.; Wangorsch, A.; May, S.; Haustein, D.; Vieths, S. Pyr c 1, the major allergen from pear (Pyrus communis), is a new member of the Bet v 1 allergen family. J. Chromatogr. B Biomed. Sci. Appl. 2001, 756, 281–293. [Google Scholar] [CrossRef]
- Gruber, P.; Vieths, S.; Wangorsch, A.; Nerkamp, J.; Hoffmann, T. Maillard Reaction and Enzymatic Browning Affect the Allergenicity of Pru av 1, the Major Allergen from Cherry (Prunus avium). J. Agric. Food Chem. 2004, 52, 4002–4007. [Google Scholar] [CrossRef] [PubMed]
- Vieths, S.; Hoffmann, A.; Holzhauser, T.; Müller, U.; Reindl, J.; Haustein, D. Factors influencing the quality of food extracts for in vitro and in vivo diagnosis. Allergy 1998, 53, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Verhoeckx, K.C.M.; Vissers, Y.M.; Baumert, J.L.; Faludi, R.; Feys, M.; Flanagan, S.; Herouet-Guicheney, C.; Holzhauser, T.; Shimojo, R.; van der Bolt, N.; et al. Food processing and allergenicity. Food Chem. Toxicol. 2015, 80, 223–240. [Google Scholar] [CrossRef] [PubMed]
- Kamath, S.D.; Rahman, A.M.A.; Voskamp, A.; Komoda, T.; Rolland, J.M.; O’Hehir, R.E.; Lopata, A.L. Effect of heat processing on antibody reactivity to allergen variants and fragments of black tiger prawn: A comprehensive allergenomic approach. Mol. Nutr. Food Res. 2014, 58, 1144–1155. [Google Scholar] [CrossRef] [PubMed]
- Hjernø, K.; Alm, R.; Canbäck, B.; Matthiesen, R.; Trajkovski, K.; Björk, L.; Roepstroff, P.; Emanuelsson, C. Down-regulation of the strawberry Bet v 1-homologous allergen in concert with the flavonoid biosynthesis pathway in colorless strawberry mutant. Proteomics 2006, 6, 1574–1587. [Google Scholar] [CrossRef] [PubMed]
- Alm, R.; Ekefjärd, A.; Krogh, M.; Häkkinen, J.; Emanuelsson, C. Proteomic variation is as large within as between strawberry varieties. J. Proteome Res. 2007, 6, 3011–3020. [Google Scholar] [CrossRef] [PubMed]
- Dölle, S.; Lehmann, K.; Schwarz, D.; Weckwert, W.; Scheler, C.; George, E.; Franken, P.; Worm, M. Allergenic activity of different tomato cultivars in tomato allergic subjects. Clin. Exp. Allergy 2011, 41, 1643–1652. [Google Scholar] [CrossRef] [PubMed]
- Laimer, M.; Maghuly, F. Awareness and knowledge of allergens: A need and a challenge to assure a safe and healthy consumption of small fruits. J. Berry Res. 2010, 1, 61–71. [Google Scholar] [CrossRef]
- Asero, R.; Marzban, G.; Martinelli, A.; Zaccarini, M.; Machado, M.L. Search for low-allergenic apple cultivars for birch-pollen-allergic patients: is there a correlation between in vitro assays and patient response? Eur. Ann. Allergy Clin. Immunol. 2006, 38, 94–98. [Google Scholar] [PubMed]
- Besler, M.; Steinhart, H.; Paschke, A. Stability of food allergens and allergenicity of processed foods. J. Chromatogr. B Biomed. Sci. Appl. 2001, 756, 207–228. [Google Scholar] [CrossRef]
- Ortolani, C.; Ispano, M.; Pastorello, E.A.; Ansaloni, E.; Magri, G.C. Comparison of results of skin prick test (with fresh foods and commercial food extracts) and RAST in 100 patients with oral allergy syndrome. J. Allergy Clin. Immunol. 1989, 83, 683–690. [Google Scholar] [CrossRef]
- Tulipani, S.; Marzban, G.; Herndl, A.; Laimer, M.; Mezzetti, B.; Battino, M. Influence of environmental and genetic factors on health-related compounds in strawberry. Food Chem. 2011, 124, 906–913. [Google Scholar] [CrossRef]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurze, E.; Kock, V.; Lo Scalzo, R.; Olbricht, K.; Schwab, W. Effect of the Strawberry Genotype, Cultivation and Processing on the Fra a 1 Allergen Content. Nutrients 2018, 10, 857. https://doi.org/10.3390/nu10070857
Kurze E, Kock V, Lo Scalzo R, Olbricht K, Schwab W. Effect of the Strawberry Genotype, Cultivation and Processing on the Fra a 1 Allergen Content. Nutrients. 2018; 10(7):857. https://doi.org/10.3390/nu10070857
Chicago/Turabian StyleKurze, Elisabeth, Vanessa Kock, Roberto Lo Scalzo, Klaus Olbricht, and Wilfried Schwab. 2018. "Effect of the Strawberry Genotype, Cultivation and Processing on the Fra a 1 Allergen Content" Nutrients 10, no. 7: 857. https://doi.org/10.3390/nu10070857
APA StyleKurze, E., Kock, V., Lo Scalzo, R., Olbricht, K., & Schwab, W. (2018). Effect of the Strawberry Genotype, Cultivation and Processing on the Fra a 1 Allergen Content. Nutrients, 10(7), 857. https://doi.org/10.3390/nu10070857