Cholesterol-Lowering and Liver-Protective Effects of Cooked and Germinated Mung Beans (Vigna radiata L.)

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Samples
2.2. Animals, Diets and Experimental Procedure
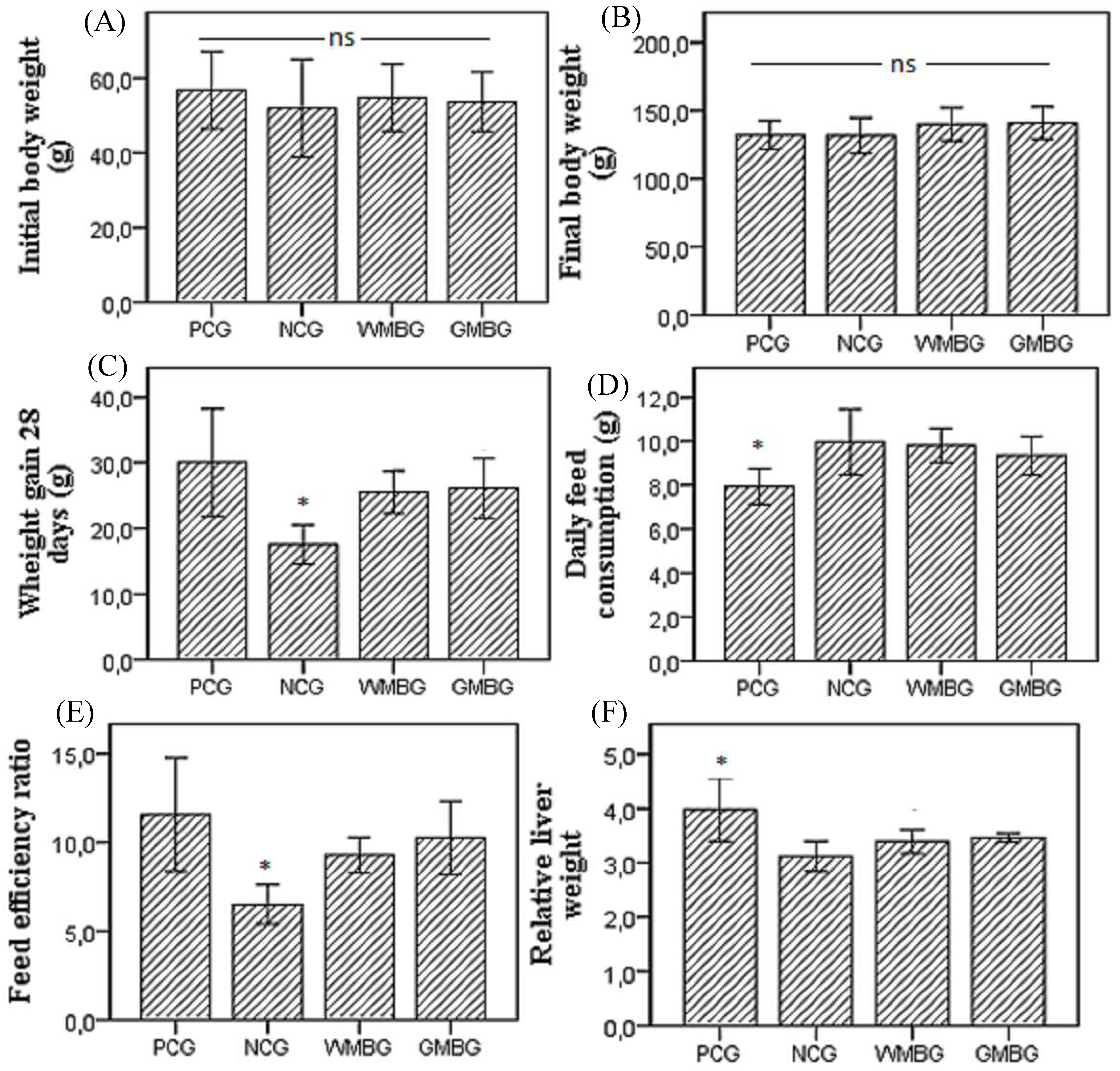
2.3. Parameters of Growth and Food Consumption
2.4. Sample Collection
2.5. Plasma Lipid and Lipoprotein Analyses
2.6. Analysis of Faecal Cholesterol
2.7. Liver Function and Liver Damage
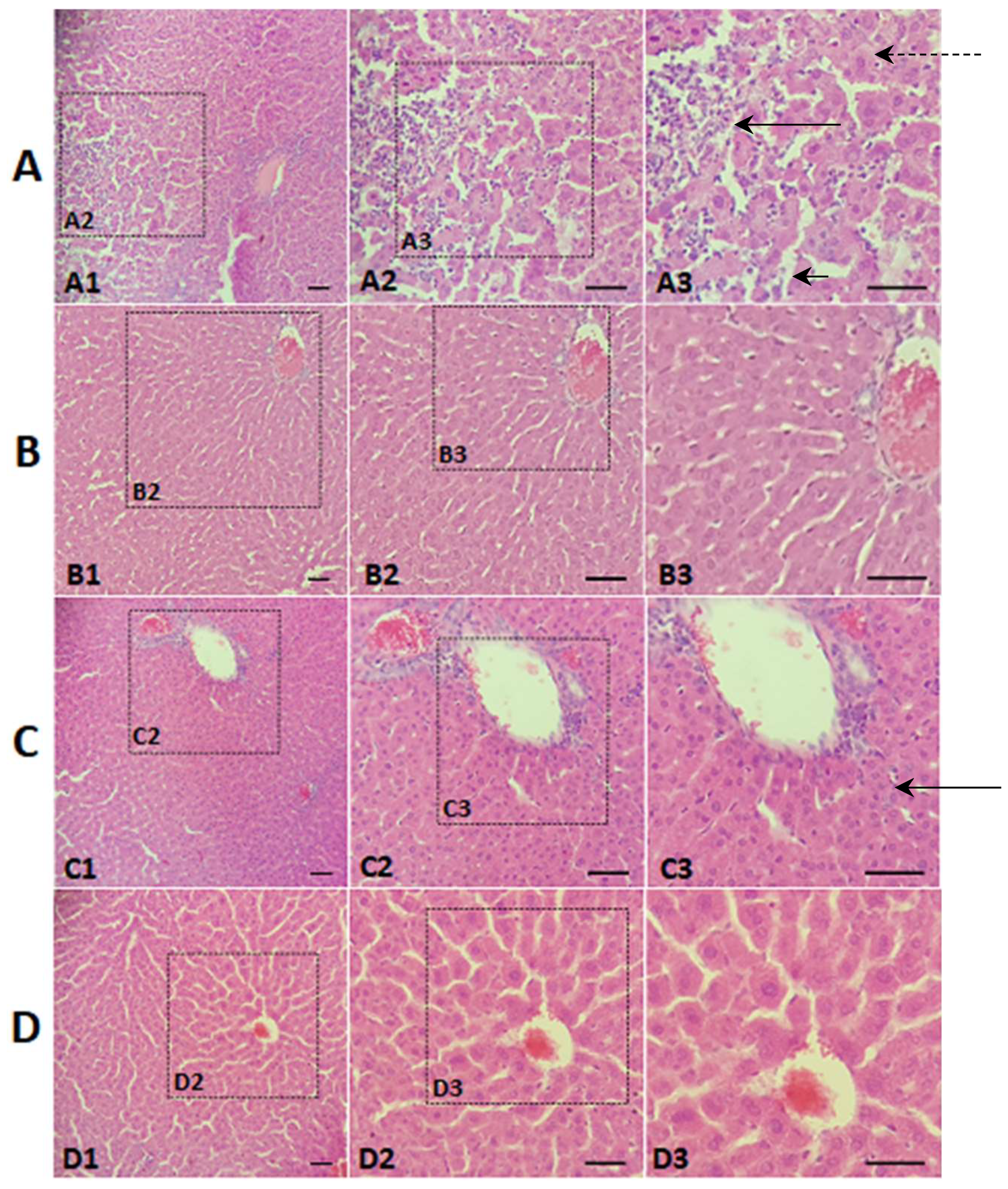
2.8. Histological Evaluation
2.9. Statistical Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). Cardiovascular Diseases. Available online: http://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 21 August 2017).
- World Health Organization (WHO). Mortality and Burden of Disease Attributable to Selected Major Risks: Results. Available online: http://www.who.int/iris/handle/10665/44203 (accessed on 21 August 2017).
- Mensink, R.P.; Zock, P.L.; Kester, A.D.M.; Katan, M.B. Effects of dietary fatty acids and carbohydrates on the ratio of serum total to HDL cholesterol and on serum lipids and apolipoproteins: A meta-analysis of 60 controlled trials. Am. J. Clin. Nutr. 2003, 77, 1146–1155. [Google Scholar] [CrossRef] [PubMed]
- Browning, J.D.; Horton, J.D. Molecular mediators of hepatic steatosis and liver injury. J. Clin. Investig. 2004, 114, 147. [Google Scholar] [CrossRef] [PubMed]
- Gaemers, I.; Groen, A. Novos esclarecimentos sobre a patogênese da doença gordurosa não-alcoólica do fígado. Curr. Opin. Lipidol. 2006, 17, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Barbana, C.; Boucher, A.; Boye, J. In vitro binding of bile salts by lentil flours, lentil protein concentrates and lentil protein hydrolysates. Food Res. Int. 2011, 44, 174–180. [Google Scholar] [CrossRef]
- Fontanari, G.G.; Batistuti, J.P.; Cruz, R.J.; Saldiva, P.H.N.; Arêas, J.A.G. Cholesterol-lowering effect of whole lupin (Lupinus albus) seed and its protein isolate. Food Chem. 2012, 132, 1521–1526. [Google Scholar] [CrossRef] [PubMed]
- Frota, K.M.G.; Mendonça, S.; Saldiva, P.H.N.; Cruz, R.J.; Arêas, J.A.G. Cholesterol-Lowering Properties of Whole Cowpea Seed and Its Protein Isolate in Hamsters. J. Food Sci. 2008, 73, H235–H240. [Google Scholar] [CrossRef] [PubMed]
- Frota, K.M.G.; Santos, R.D.; Ribeiro, V.Q.; Areas, J.A. Cowpea protein reduces LDL-cholesterol and apolipoprotein B concentrations, but does not improve biomarkers of inflammation or endothelial dysfunction in adults with moderate hypercholesterolemia. Nutr. Hosp. 2015, 31, 1611–1619. [Google Scholar] [CrossRef]
- Xue, Z.; Hou, X.; Yu, W.; Wen, H.; Zhang, Q.; Li, D.; Kou, X. Lipid metabolism potential and mechanism of CPe-III from chickpea (Cicer arietinum L.). Food Res. Int. 2017, 104, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, S.; Saldiva, P.H.; Cruz, R.J.; Arêas, J.A.G. Amaranth protein presents cholesterol-lowering effect. Food Chem. 2009, 116, 738–742. [Google Scholar] [CrossRef]
- Chavez-Santoscoy, R.A.; Gutierrez-Uribe, J.A.; Granados, O.; Torre-Villalvazo, I.; Serna-Saldivar, S.O.; Torres, N.; Palacios-Gonzalez, B.; Tovar, A.R. Flavonoids and saponins extracted from black bean (Phaseolus vulgaris L.) seed coats modulate lipid metabolism and biliary cholesterol secretion in C57BL/6 mice. Br. J. Nutr. 2014, 112, 886–899. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.R.; Fontanari, G.G.; Pimenta, D.C.; Soares-Freitas, R.M.; Arêas, J.A.G. Proteolytic hydrolysis of cowpea proteins is able to release peptides with hypocholesterolemic activity. Food Res. Int. 2015, 77, 43–48. [Google Scholar] [CrossRef]
- Soares, R.; Mendonça, S.; de Castro, L.Í.; Menezes, A.; Arêas, J. Major Peptides from Amaranth (Amaranthus cruentus) Protein Inhibit HMG-CoA Reductase Activity. Int. J. Mol. Sci. 2015, 16, 4150–4160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shang, P.; Qin, F.; Zhou, Q.; Gao, B.; Huang, H.; Yang, H.; Shi, H.; Lucy Yu, L. Chemical composition and antioxidative and anti-inflammatory properties of ten commercial mung bean samples. LWT Food Sci. Technol. 2013, 54, 171–178. [Google Scholar] [CrossRef]
- Vieira, R.F.; Oliveira, V.R.; Vieira, C. Cultivo do feijão-mungo-verde no verão em Viçosa e em Prudente de Morais. Hortic. Bras. 2003, 21, 37–43. [Google Scholar] [CrossRef]
- Barradas, C.A.A.; Sayão, F.A.D.; Duque, F.F. Feijão Mungo-Uma Alternativa Protéica na Alimentação; Empresa Brasileira de Pesquisa Agropecuária: Brasília, Brasil, 1989. [Google Scholar]
- Mamilla, R.K.; Mishra, V.K. Effect of germination on antioxidant and ACE inhibitory activities of legumes. LWT Food Sci. Technol. 2017, 75, 51–58. [Google Scholar] [CrossRef]
- Tang, D.; Dong, Y.; Guo, N.; Li, L.; Ren, H. Metabolomic analysis of the polyphenols in germinating mung beans (Vigna radiata) seeds and sprouts. J. Sci. Food Agric. 2014, 94, 1639–1647. [Google Scholar] [CrossRef] [PubMed]
- Wongsiri, S.; Ohshima, T.; Duangmal, K. Chemical Composition, Amino Acid Profile and Antioxidant Activities of Germinated Mung Beans (Vigna radiata). J. Food Process. Preserv. 2015, 39, 1956–1964. [Google Scholar] [CrossRef]
- Tachibana, N.; Wanezaki, S.; Nagata, M.; Motoyama, T. Intake of Mung Bean Protein Isolate Reduces Plasma Triglyceride Level in Rats. Funct. Foods Health Dis. 2013, 3, 365–376. [Google Scholar]
- Yao, Y.; Hao, L.; Shi, Z.; Wang, L.; Cheng, X.; Wang, S.; Ren, G. Mung Bean Decreases Plasma Cholesterol by Up-regulation of CYP7A1. Plant Foods Hum. Nutr. 2014, 69, 134–136. [Google Scholar] [CrossRef] [PubMed]
- Yeap, S.K.; Beh, B.K.; Ho, W.Y.; Mohd Yusof, H.; Mohamad, N.E.; Ali, N.M.; Jaganath, I.B.; Alitheen, N.B.; Koh, S.P.; Long, K. In vivo antioxidant and hypolipidemic effects of fermented mung bean on hypercholesterolemic mice. Evid.-Based Complement. Altern. Med. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.R.; Soares-Freitas, R.A.M.; Corrêa, C.A.C.; Siguemoto, É.S.; Fontanari, G.G.; Arêas, J.A.G. Peptides from cowpea present antioxidant activity, inhibit cholesterol synthesis and its solubilisation into micelles. Food Chem. 2015, 168, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, H.; Jiao, R.; Peng, C.; Wong, Y.M.; Yeung, V.S.Y.; Huang, Y.; Chen, Z.Y. Choosing hamsters but not rats as a model for studying plasma cholesterol-lowering activity of functional foods. Mol. Nutr. Food Res. 2009, 53, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Cai, W.; Xu, B. Kinetic changes of nutrients and antioxidant capacities of germinated soybean (glycine max l.) and mung bean (vigna radiata l.) with germination time. Food Chem. 2014, 143, 268–276. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed; National Academies Press: Washington, DC, USA, 2010. [Google Scholar]
- Association of Offccial Analytical Chemists (AOAC). Official Methods of Analysis; Association of Offccial Analytical Chemists: Washington, DC, USA, 2016. [Google Scholar]
- Food and Agriculture Organization of the United Nations (FAO)/World Health Organization (WHO). Protein Quality Evaluation: Report of the JOINT FAO/WHO Expert Consultation; FAO: Rome, Italy; WHO: Bethesda, MD, USA, 1991; ISBN 9251030979. [Google Scholar]
- Weingand, K.; Chemistry, B.D.-C. Quantification of high-density-lipoprotein cholesterol in plasma from hamsters by differential precipitation. Clin. Chem. 1990, 36, 575. [Google Scholar] [PubMed]
- Goulinet, S.; Chapman, M.J. Plasma lipoproteins in the golden Syrian hamster (Mesocricetus auratus): Heterogeneity of apoB- and apoA-I-containing particles. J. Lipid Res. 1993, 34, 943–959. [Google Scholar] [PubMed]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the Concentration of Low-Density Lipoprotein Cholesterol in Plasma, without Use of the Preparative Ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [PubMed]
- Saldanha, T.; Sawaya, A.C.H.F.; Eberlin, M.N.; Bragagnolo, N. HPLC Separation and Determination of 12 Cholesterol Oxidation Products in Fish: Comparative Study of RI, UV, and APCI-MS Detectors. J. Agric. Food Chem. 2006, 54, 4107–4113. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.C.; Lin, Y.L.; Kuo, C.F. Effect of high-fat diet on hepatic proteomics of hamsters. J. Agric. Food Chem. 2015, 63, 1869–1881. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, E.; Amaral, A. Hypocholesterolaemic effect of rat-administered oral doses of the isolated 7S globulins from cowpeas and adzuki beans. J. Nutr. Sci. 2015, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, N.; Iwaoka, Y.; Hirotsuka, M.; Horio, F.; Kohno, M. β-Conglycinin Lowers Very-Low-Density Lipoprotein-Triglyceride Levels by Increasing Adiponectin and Insulin Sensitivity in Rats. Biosci. Biotechnol. Biochem. 2010, 74, 1250–1255. [Google Scholar] [CrossRef] [PubMed]
- Higaki, N.; Sato, K.; Suda, H.; Suzuka, T. Evidence for the existence of a soybean resistant protein that captures bile acid and stimulates its fecal excretion. Bioscience 2006, 70, 2844–2852. [Google Scholar] [CrossRef] [PubMed]
- Muzquiz, M.; Varela, A.; Burbano, C.; Cuadrado, C.; Guillamón, E.; Pedrosa, M.M. Bioactive compounds in legumes: Pronutritive and antinutritive actions. implications for nutrition and health. Phytochem. Rev. 2012, 11, 227–244. [Google Scholar] [CrossRef]
- Talukder, S. Effect of dietary fiber on properties and acceptance of meat products: A review. Crit. Rev. Food Sci. Nutr. 2015, 55, 1005–1101. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.N.; Yusof, M.H.; Long, K.; Yeap, S.K.; Ho, W.Y.; Beh, B.K.; Koh, S.P.; Abdullah, M.P.; Alitheen, N.B. Antioxidant and hepatoprotective effect of aqueous extract of germinated and fermented mung bean on ethanol-mediated liver damage. BioMed Res. Int. 2012, 2013. [Google Scholar] [CrossRef]
- Bellassoued, K.; Ghrab, F.; Makni-Ayadi, F.; Van Pelt, J.; Elfeki, A.; Ammar, E. Protective effect of kombucha on rats fed a hypercholesterolemic diet is mediated by its antioxidant activity. Pharm. Biol. 2015, 53, 1699–1709. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.W.; Adams, L.A. Non-alcoholic fatty liver disease. Crit. Rev. Clin. Lab. 2011, 48, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Rawson, A.; Patras, A.; Tiwari, B.K.; Noci, F.; Koutchma, T.; Brunton, N. Effect of thermal and non thermal processing technologies on the bioactive content of exotic fruits and their products: Review of recent advances. Food Res. Int. 2011, 44, 1875–1887. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Component (%) | WMBF | GMBF |
|---|---|---|
| Moisture | 7.4 ± 0.6 b | 7.0 ± 0.3 a |
| Ash | 3.4 ± 0.1 a | 4.1 ± 0.1 b |
| Lipid | 1.3 ± 0.1 b | 0.6 ± 0.1 a |
| Proteín | 23.4 ± 0.2 a | 26.2 ± 0.4 b |
| Carbohydrate A | 72.0 ± 0.2 b | 69.0 ± 0.3 a |
| Planned Composition of the Diets (g/kg Dry Matter) | ||||
| Ingredients (g/kg) | CD A | CHD | WMBD | GMBD |
| Caseín B | - | 221 | - | - |
| Whole mung bean | - | - | 855 | - |
| Germinated mung bean | - | - | - | 769 |
| L-methionine | - | - | 3 | 3 |
| Sucrose | - | 50 | 50 | 50 |
| Corn starch | - | 427.5 | - | - |
| Microcrystalline cellulose | - | 100 | 60 | - |
| Soy oil | - | 20 | 8 | 14 |
| Coconut oil | - | 130 | 130 | 130 |
| Cholesterol | - | 1 | 1 | 1 |
| Choline bitartrate | - | 2.5 | 2.5 | 2.5 |
| Mineral mix AIN 93 C | - | 35 | 35 | 35 |
| Vitamin mix AIN 93 C | - | 10 | 10 | 10 |
| BHT | - | 0.024 | 0.024 | 0.024 |
| Determined Centesimal Composition of the Diets (g/100 g Dry Matter) | ||||
| Moisture | 10.2 ± 0.1 c | 6.8 ± 0.2 a | 7.9 ± 0.2 b | 7.3 ± 0.2 a |
| Ash | 7.8 ± 0.1 d | 4.8 ± 0.2 b | 5.4 ± 0.1 c | 4.1 ± 0.6 a |
| Lipid | 3.3 ± 0.1 c | 14.5 ± 0.1 b | 12.7 ± 0.1 a,b | 11.8 ± 0.2 a |
| Proteín (N × 6.25) | 20.9 ± 0.4 a | 19.1 ± 1.0 a | 20.3 ± 0.2 a | 18.9 ± 0.2 a |
| Carbohydrate D | 68.2 ± 0.3 c | 61.3 ± 0.8 a | 61.5 ± 0.6 a | 65.5 ± 0.4 b |
| TEV (KJ/g) | 16.36 | 19.0 | 18.6 | 18.7 |
| Lipid Parameters | NCG | PCG | WMBG | GMBG |
|---|---|---|---|---|
| Total cholesterol (mg/dL) | 93.1 ± 4.1 a | 191.0 ± 12.6 c | 151.4 ± 5.3 b | 145.3 ± 6.4 b |
| Triacylglycerols (mg/dL) | 31.9 ± 5.7 a | 88.8 ± 4.5 b | 68.7 ± 7.3 b | 78.0 ± 9.3 b |
| HDL-cholesterol (mg/dL) | 24.7 ± 1.2 a | 44.8 ± 3.8 b | 41.0 ± 3.6 b | 38.4 ± 1.1 b |
| Non-HDL cholesterol (mg/dL) A | 68.4 ± 3.6 a | 146.2 ± 12.5 c | 110.4 ± 5.5 b | 106.8 ± 5.7 b |
| Faecal cholesterol (μmol/d/BW) | 2.9 ± 0.4 a | 5.8 ± 0.7 b | 11.1 ± 0.3 c | 11.3 ± 0.4 c |
| Analysis | NCG | PCG | WMBG | GMBG |
|---|---|---|---|---|
| Total protein A (g/dL) | 5.6 ± 0.1 | 5.92 ± 0.1 | 5.77 ± 0.1 | 5.85 ± 0.2 |
| Albumin A (g/dL) | 2.1 ± 0.5 | 2.23 ± 0.2 | 2.15 ± 0.5 | 2.22 ± 0.5 |
| AST B (U/L) | 61.4 ± 5.7 * | 110.16 ± 6.9 | 74.90 ± 8.0 * | 56.75 ± 7.3 * |
| ALT B (U/L) | 73.7 ± 6.9 * | 105.16 ± 21.5 | 75.70 ± 4.1 * | 65.37 ± 8.6 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopes, L.A.R.; Martins, M.D.C.d.C.e.; Farias, L.M.d.; Brito, A.K.d.S.; Lima, G.D.M.; Carvalho, V.B.L.d.; Pereira, C.F.d.C.; Conde Júnior, A.M.; Saldanha, T.; Arêas, J.A.G.; et al. Cholesterol-Lowering and Liver-Protective Effects of Cooked and Germinated Mung Beans (Vigna radiata L.). Nutrients 2018, 10, 821. https://doi.org/10.3390/nu10070821
Lopes LAR, Martins MDCdCe, Farias LMd, Brito AKdS, Lima GDM, Carvalho VBLd, Pereira CFdC, Conde Júnior AM, Saldanha T, Arêas JAG, et al. Cholesterol-Lowering and Liver-Protective Effects of Cooked and Germinated Mung Beans (Vigna radiata L.). Nutrients. 2018; 10(7):821. https://doi.org/10.3390/nu10070821
Chicago/Turabian StyleLopes, Lays Arnaud Rosal, Maria Do Carmo de Carvalho e Martins, Luciana Melo de Farias, Ana Karolinne da Silva Brito, Geovanni De Morais Lima, Vanessa Brito Lira de Carvalho, Cristian Francisco de Carvalho Pereira, Aírton Mendes Conde Júnior, Tatiana Saldanha, José Alfredo Gomes Arêas, and et al. 2018. "Cholesterol-Lowering and Liver-Protective Effects of Cooked and Germinated Mung Beans (Vigna radiata L.)" Nutrients 10, no. 7: 821. https://doi.org/10.3390/nu10070821
APA StyleLopes, L. A. R., Martins, M. D. C. d. C. e., Farias, L. M. d., Brito, A. K. d. S., Lima, G. D. M., Carvalho, V. B. L. d., Pereira, C. F. d. C., Conde Júnior, A. M., Saldanha, T., Arêas, J. A. G., Silva, K. J. D. e., & Frota, K. D. M. G. (2018). Cholesterol-Lowering and Liver-Protective Effects of Cooked and Germinated Mung Beans (Vigna radiata L.). Nutrients, 10(7), 821. https://doi.org/10.3390/nu10070821