Protective Effect of Aplysin Supplementation on Intestinal Permeability and Microbiota in Rats Treated with Ethanol and Iron

Abstract
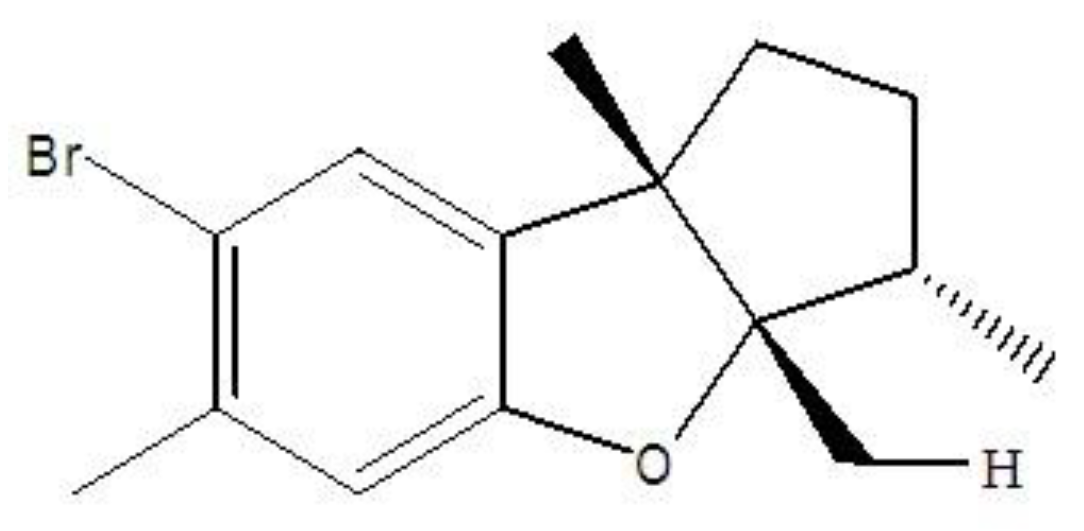
:1. Introduction
2. Materials and Methods
2.1. Animal and Ethics Statement
2.2. Experimental Designs and Animal Treatment
2.3. Histopathological Analysis
2.4. Estimation of Serum Ferritin, Hepcidin and Iron-Related Protein Expression
2.5. Measurement of Endotoxin Assay and Intestinal Mucosal Barrier Assay
2.6. Gut Microbiota Analysis
2.7. Statistical Analysis
3. Results
3.1. Effects of Aplysin on Food Intake and Growth Performance
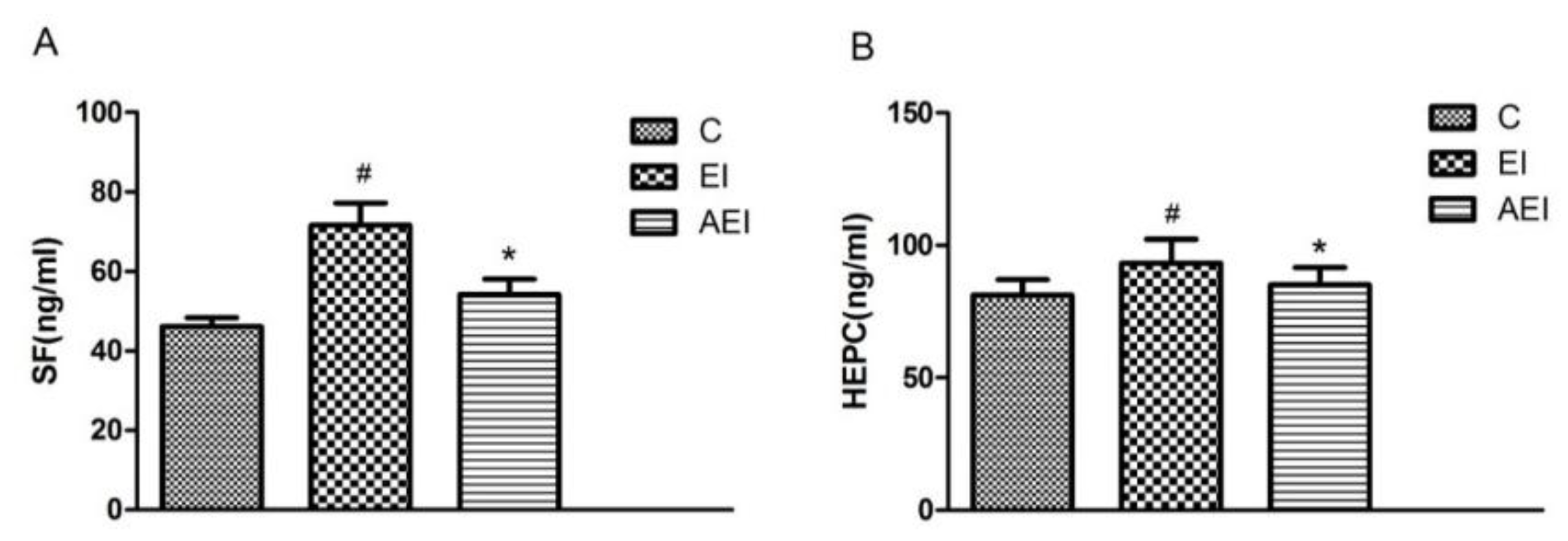
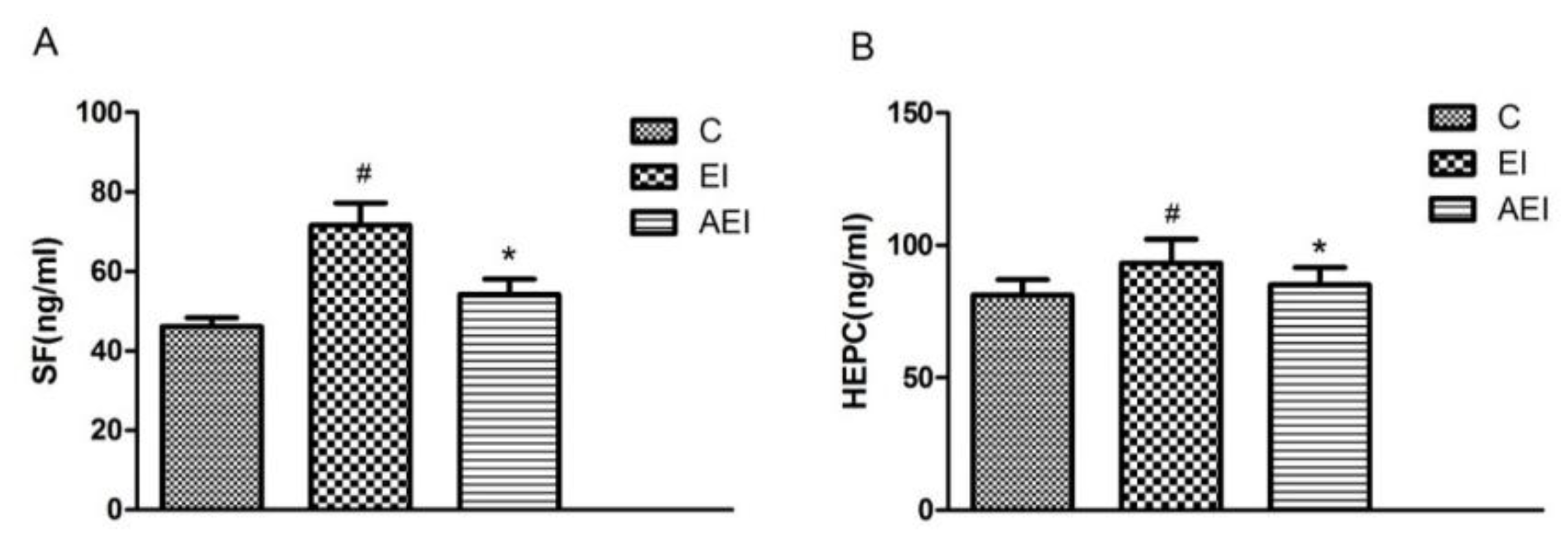
3.2. Effects of Aplysin on Serum Ferritin and Hepcidin
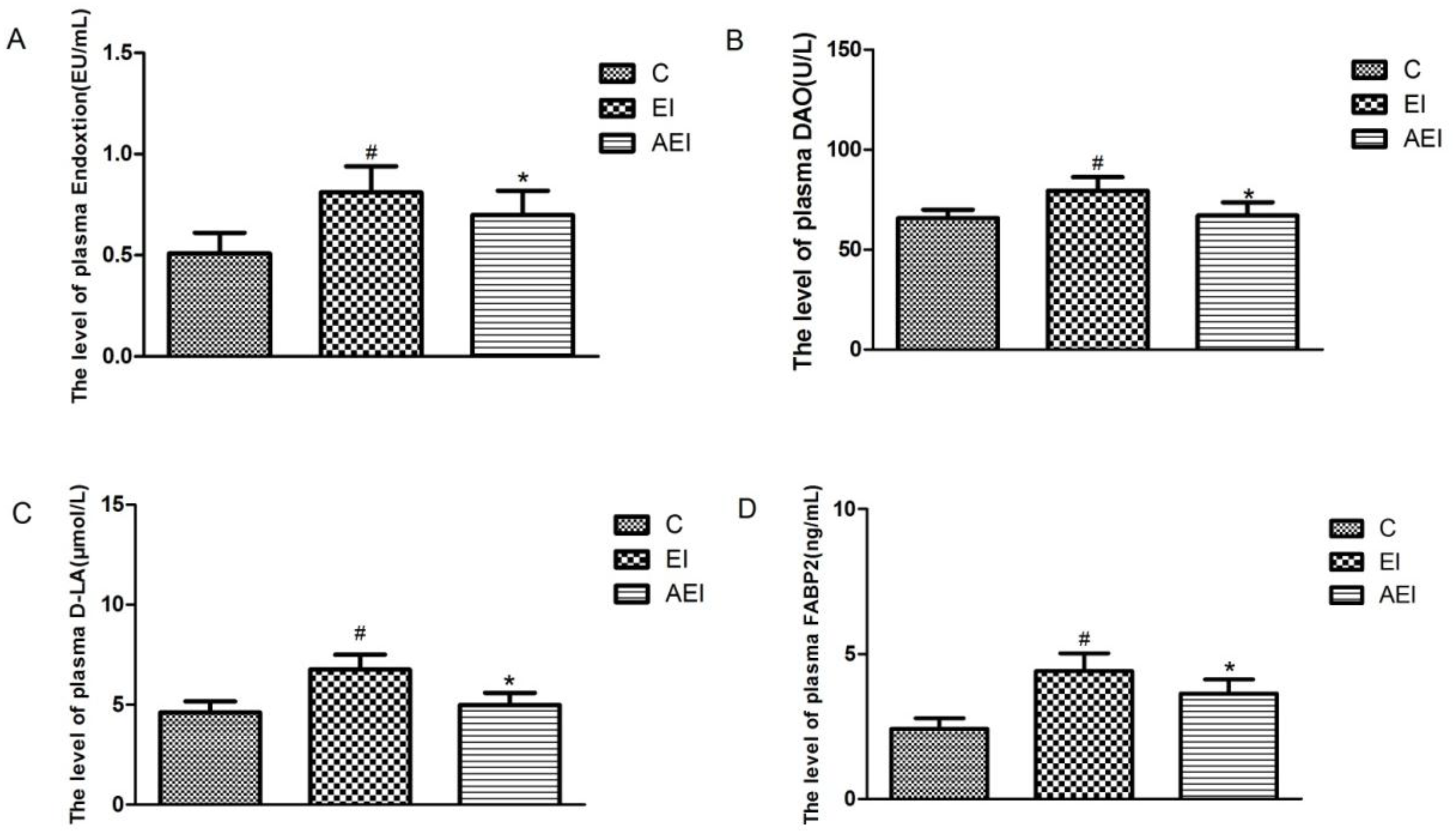
3.3. Effects of Aplysin on Intestinal Permeability
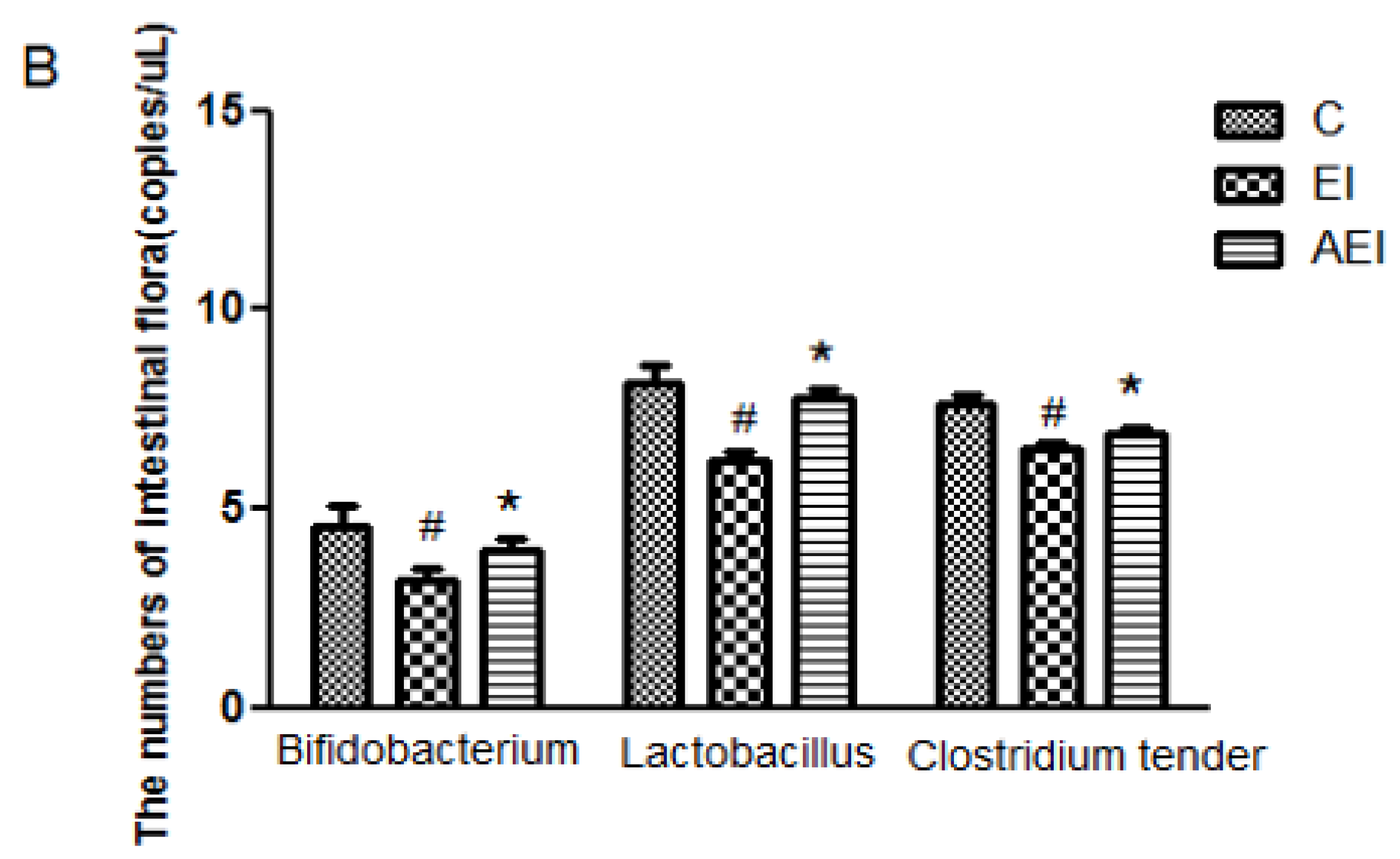
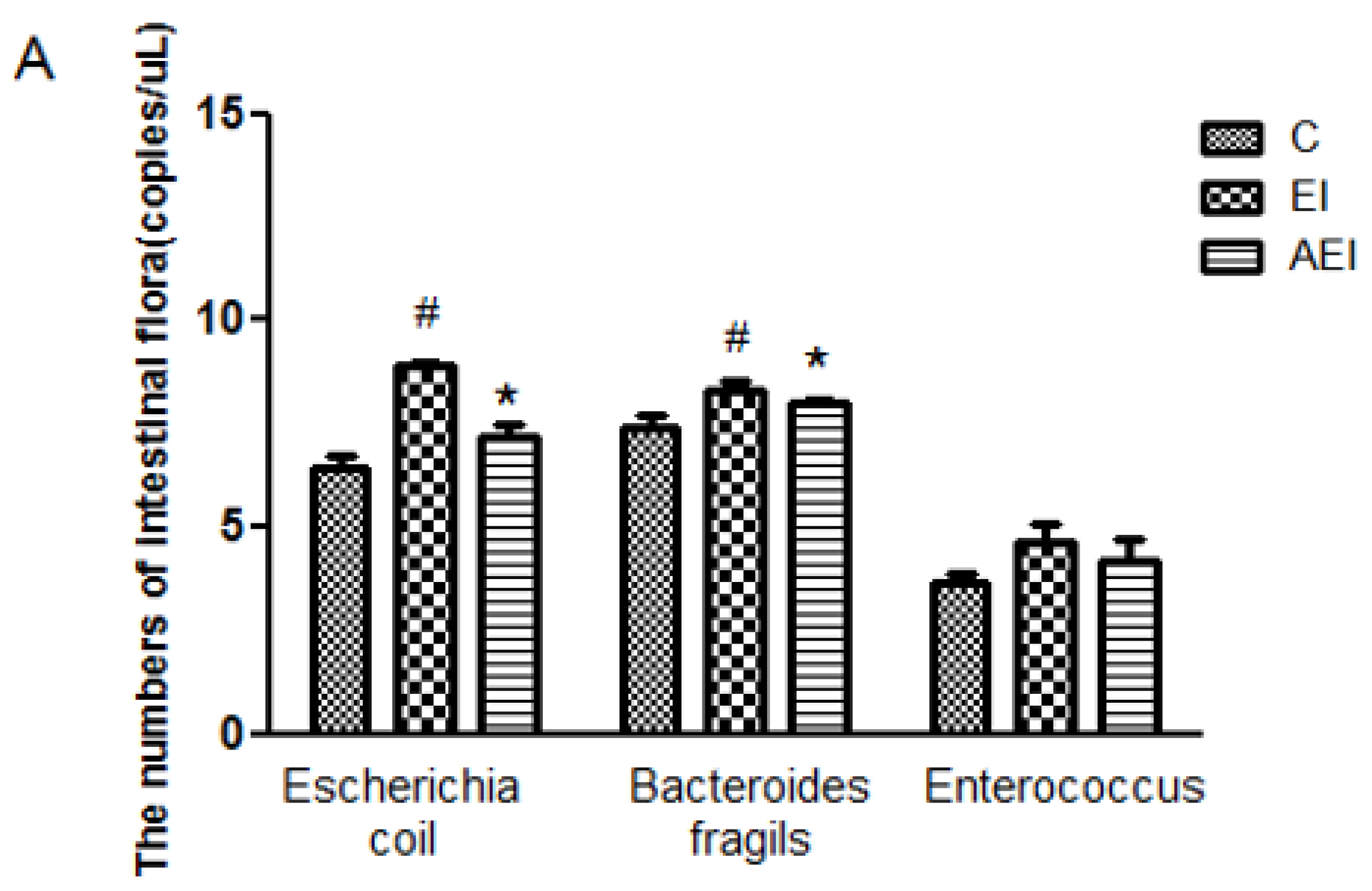
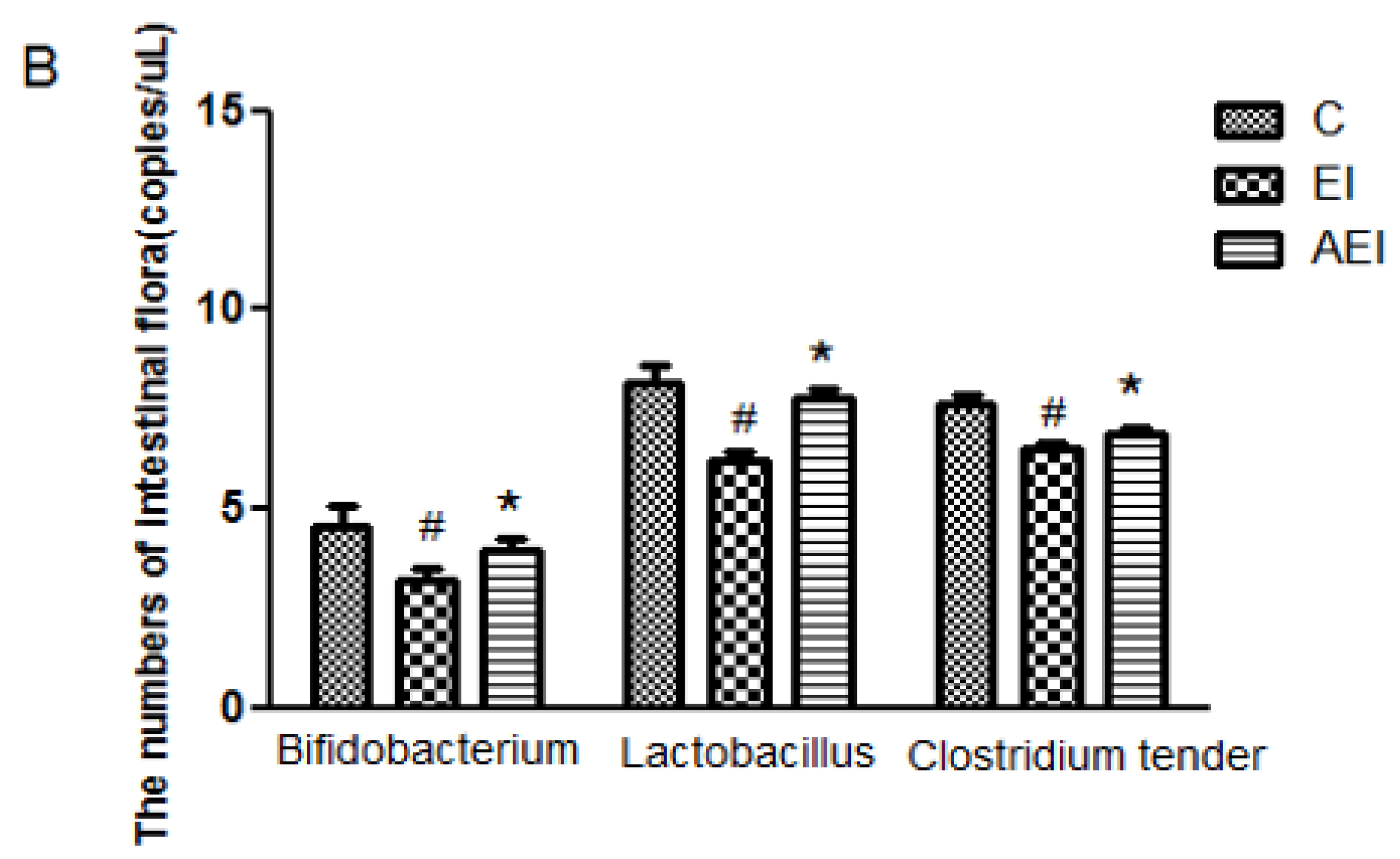
3.4. Effects of Aplysin on Intestinal Microbiota
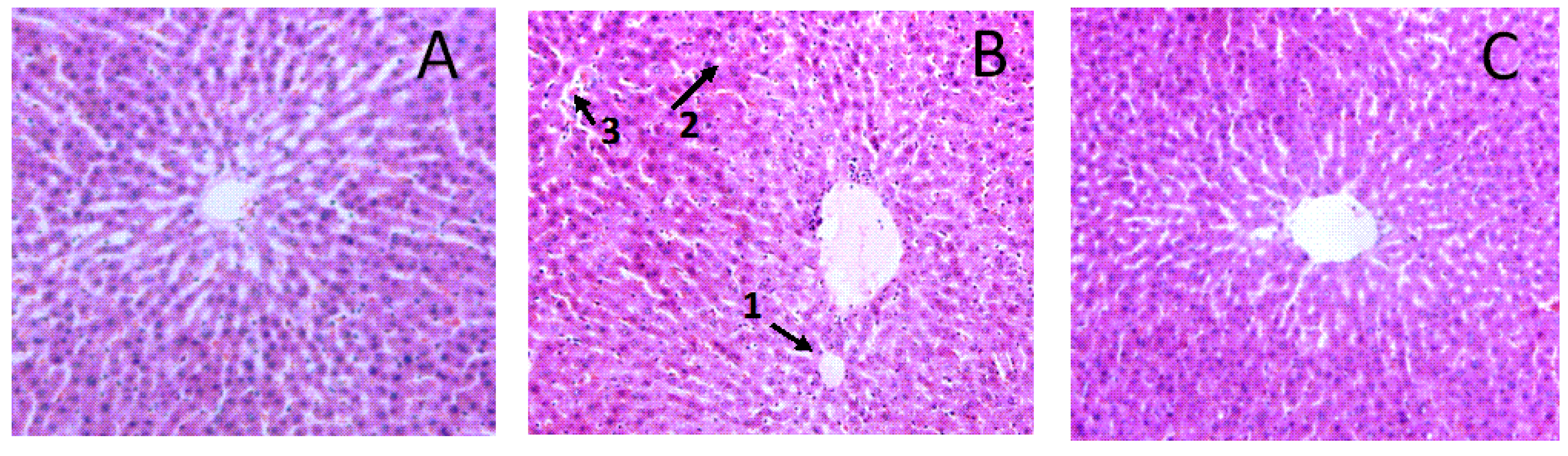
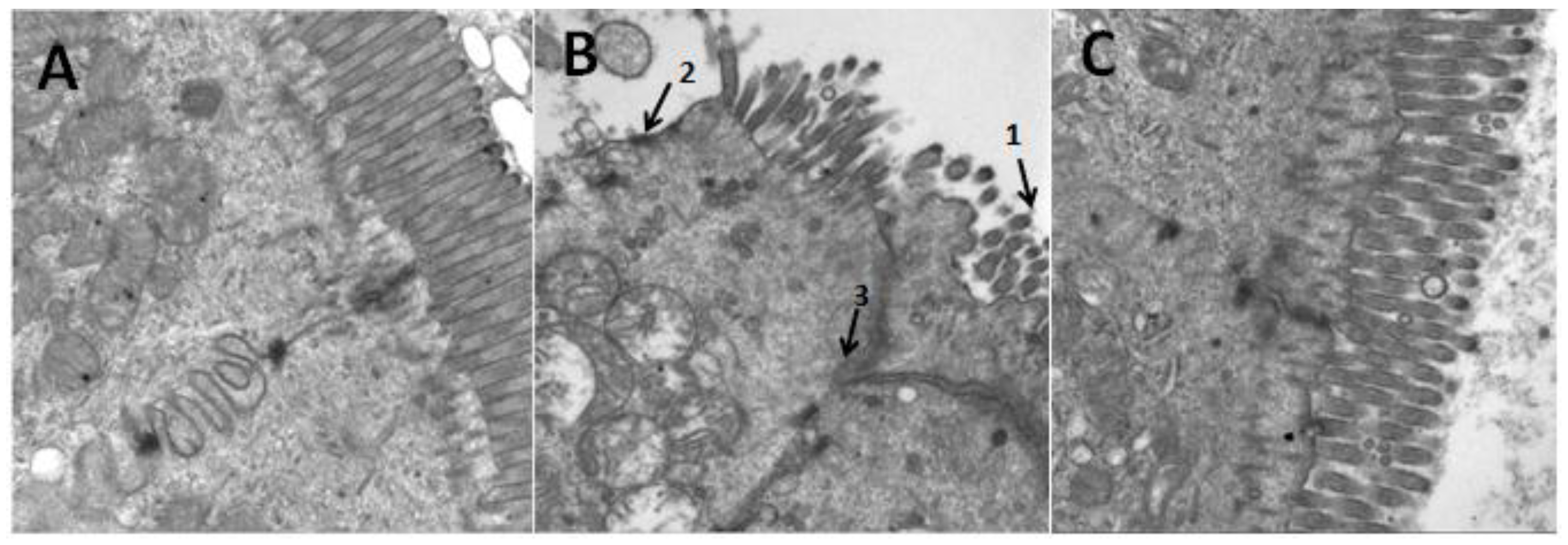
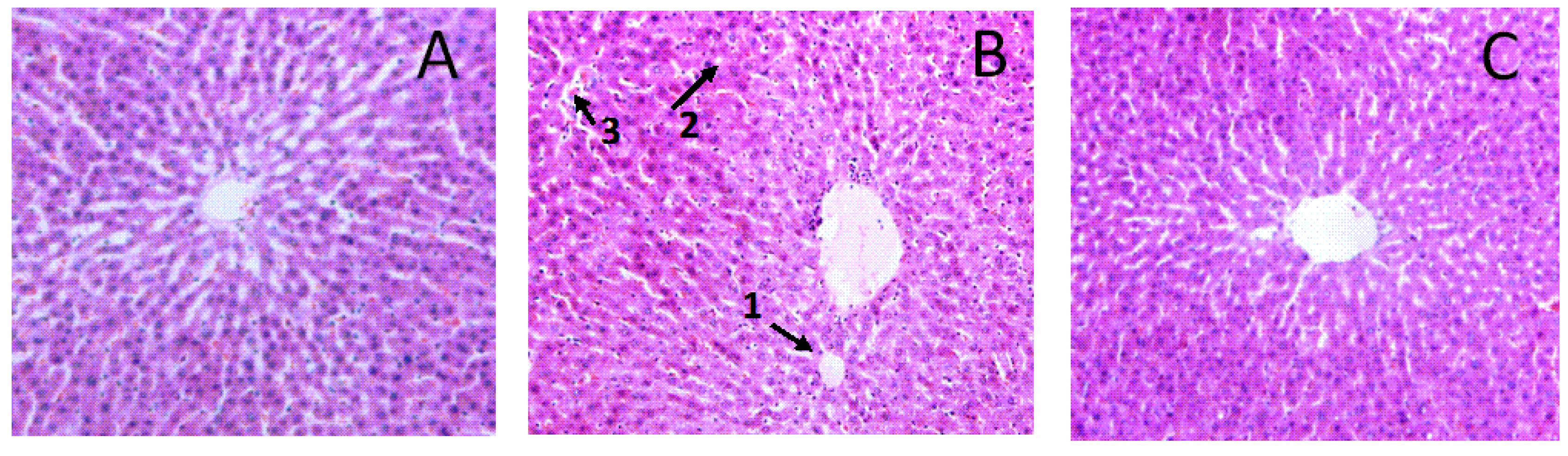
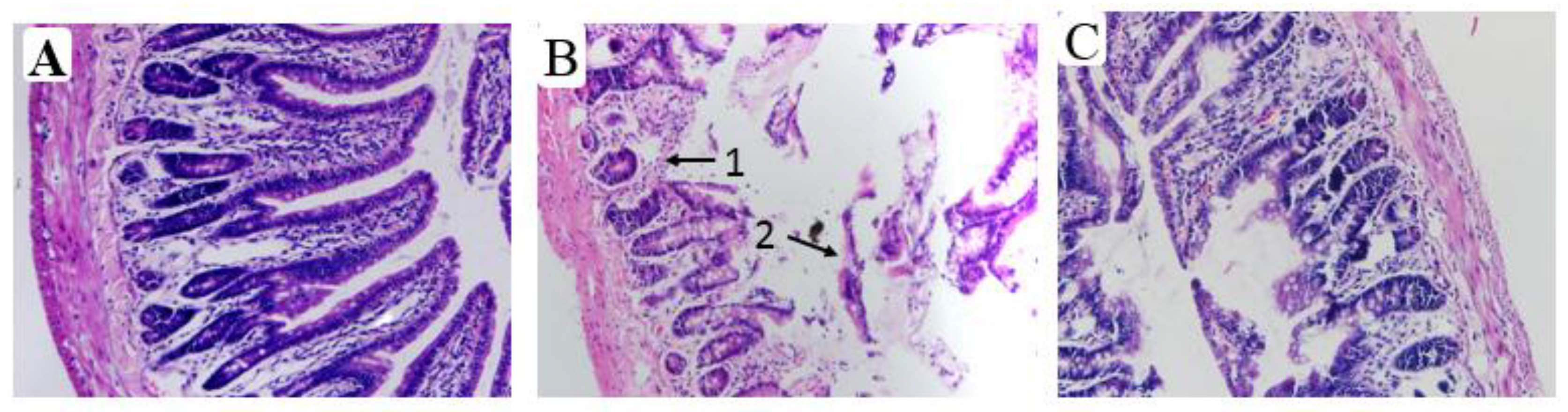
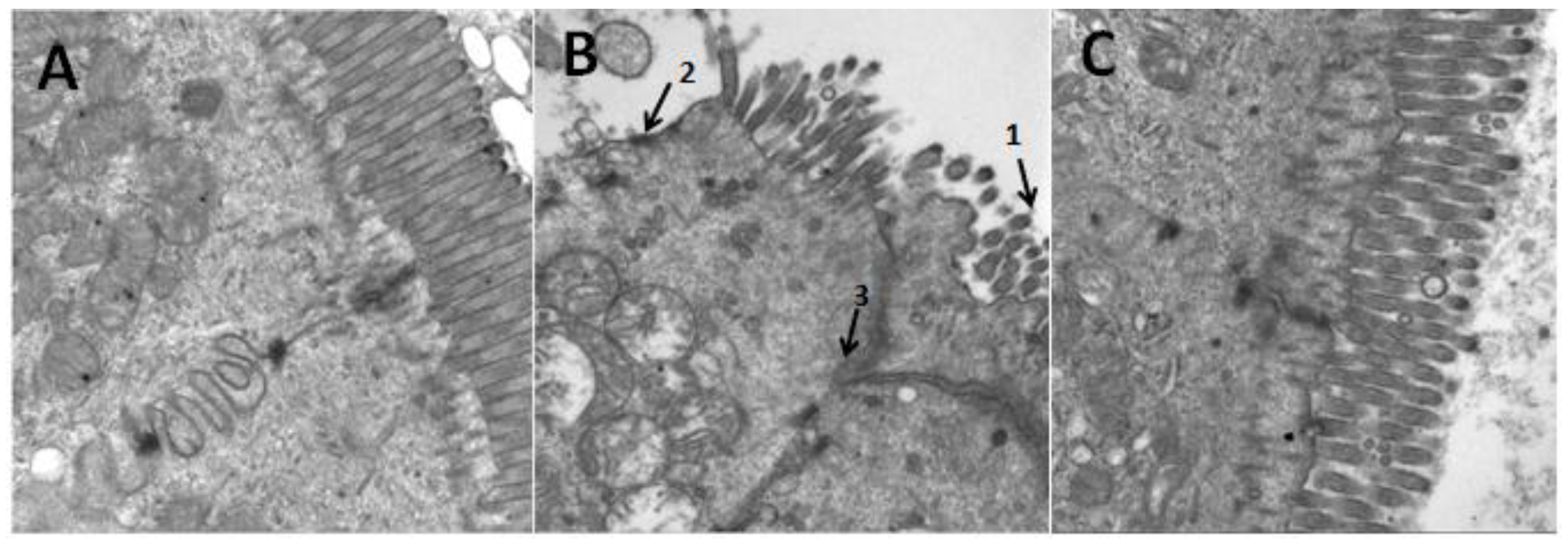
3.5. Effects of Aplysin on Pathological Changes of Liver and Intestine
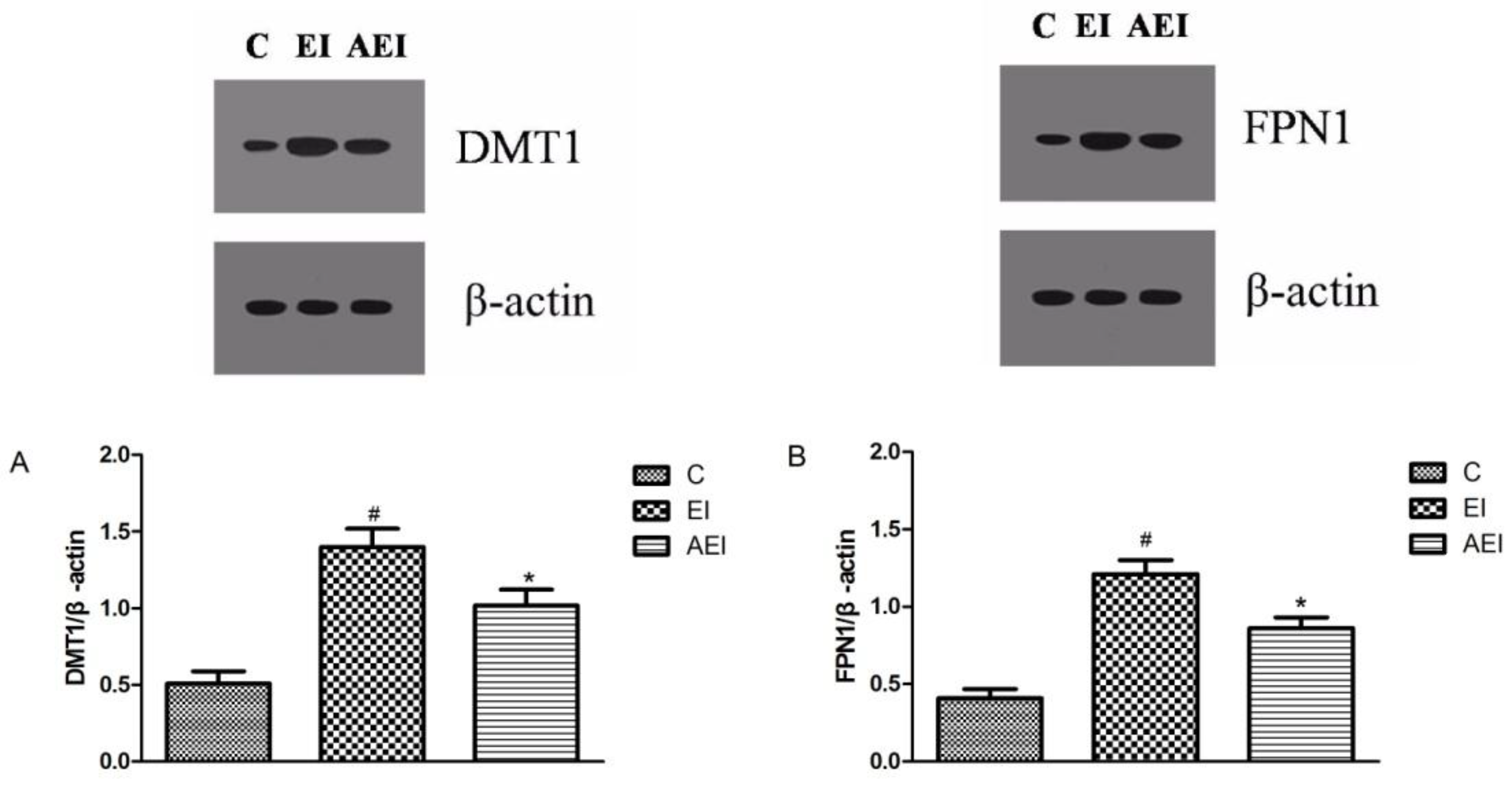
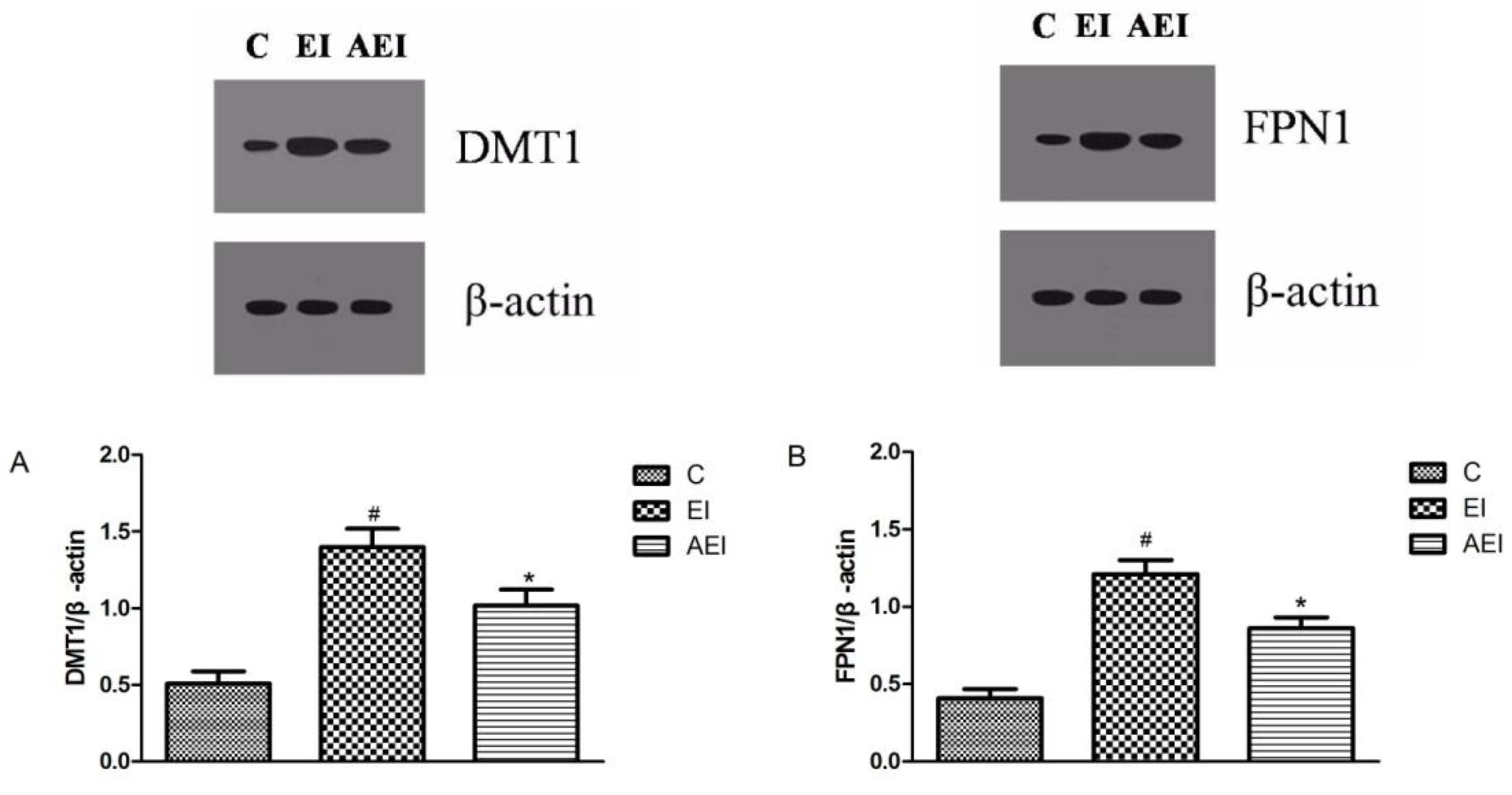
3.6. Effects of Aplysin on Expressions of Iron-Related Proteins in the Intestine
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Galaris, D.; Pantopoulos, K. Oxidative stress and iron homeostasis: Mechanistic and health aspects. Crit. Rev. Clin. Lab. Sci. 2008, 45, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, H.; Ebata, Y.; Sakabe, T.; Hama, S.; Kogure, K.; Shiota, G. High-fat, high-fructose diet induces hepatic iron overload via a hepcidin-independent mechanism prior to the onset of liver steatosis and insulin resistance in mice. Metabolism 2013, 62, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Pascoe, A.; Kerlin, P.; Steadman, C.; Clouston, A.; Jones, D.; Powell, L.; Jazwinska, E.; Lynch, S.; Strong, R. Spur cell anaemia and hepatic iron stores in patients with alcoholic liver disease undergoing orthotopic liver transplantation. Gut 1999, 45, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, L.M.; Halliday, J.W.; Powell, L.W. Interrelationships of alcohol and iron in liver disease with particular reference to the iron-binding proteins, ferritin and transferrin. J. Gastroenterol. Hepatol. 1999, 14, 202–214. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.W.; Fouts, D.E.; Brandl, J.; Starkel, P.; Torralba, M.; Schott, E.; Tsukamoto, H.; Nelson, K.E.; Brenner, D.A.; Schnabl, B. Enteric dysbiosis associated with a mouse model of alcoholic liver disease. Hepatology 2011, 53, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, F.; Lu, H.; Wang, B.; Chen, Y.; Lei, D.; Wang, Y.; Zhu, B.; Li, L. Characterization of fecal microbial communities in patients with liver cirrhosis. Hepatology 2011, 54, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Deitch, E.A. Dissolution of lipids from mucus: A possible mechanism for prompt disruption of gut barrier function by alcohol. Toxicol. Lett. 2015, 232, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Kirpich, I.A.; Feng, W.; Wang, Y.; Liu, Y.; Beier, J.I.; Arteel, G.E.; Falkner, K.C.; Barve, S.S.; McClain, C.J. Ethanol and dietary unsaturated fat (corn oil/linoleic acid enriched) cause intestinal inflammation and impaired intestinal barrier defense in mice chronically fed alcohol. Alcohol 2013, 47, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Kishikawa, H.; Miura, S.; Nishida, J.; Nakano, M.; Hirano, E.; Sudo, N.; Morishita, T.; Ishii, H. Ethanol-induced CXC-chemokine synthesis and barrier dysfunction in intestinal epithelial cells. Alcohol. Clin. Exp. Res. 2005, 29, 2116–2122. [Google Scholar] [CrossRef] [PubMed]
- Elamin, E.; Masclee, A.; Dekker, J.; Jonkers, D. Ethanol disrupts intestinal epithelial tight junction integrity through intracellular calcium-mediated Rho/ROCK activation. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G677–G685. [Google Scholar] [CrossRef] [PubMed]
- Summa, K.C.; Voigt, R.M.; Forsyth, C.B.; Shaikh, M.; Cavanaugh, K.; Tang, Y.; Vitaterna, M.H.; Song, S.; Turek, F.W.; Keshavarzian, A. Disruption of the Circadian Clock in Mice Increases Intestinal Permeability and Promotes Alcohol-Induced Hepatic Pathology and Inflammation. PLoS ONE 2013, 8, e67102. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, L.M.; Powell, L.W. Hemochromatosis and alcoholic liver disease. Alcohol 2003, 30, 131–136. [Google Scholar] [CrossRef]
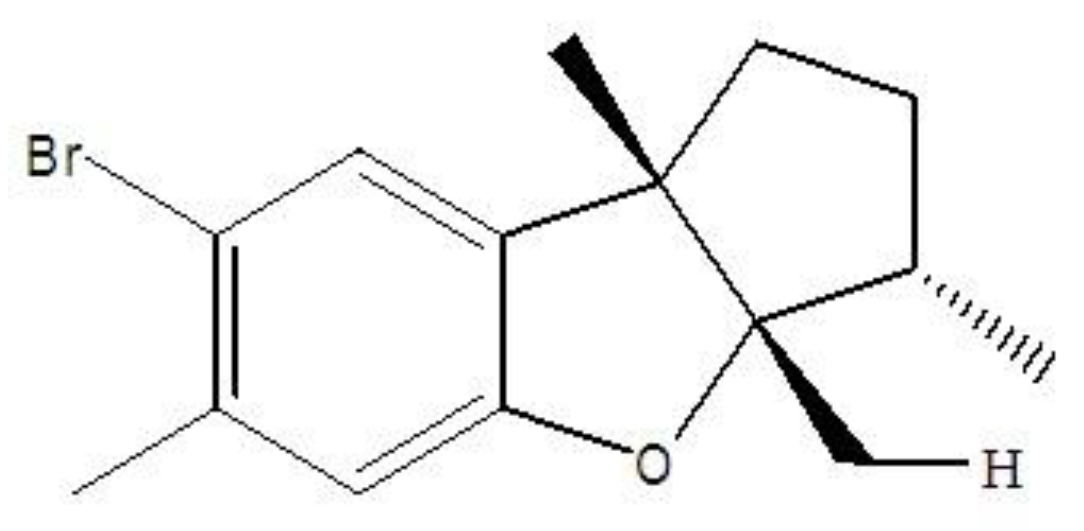
- Ge, N.; Liang, H.; Liu, Y.; Ma, A.G.; Han, L. Protective effect of Aplysin on hepatic injury in ethanol-treated rats. Food Chem. Toxicol. 2013, 62, 361–372. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Liang, H.; Li, Y.; Shi, D.Y.; Ma, A.G. Antioxidant effect of Aplysin on aged mice exposed to D-galatose. Chin. J. Public Health 2009, 25, 1122–1123. [Google Scholar]
- Xue, M.; Liu, Y.; Lyu, R.; Ge, N.; Liu, M.; Ma, Y.; Liang, H. Protective effect of aplysin on liver tissue and the gut microbiota in alcohol-fed rats. PLoS ONE 2017, 12, e0178684. [Google Scholar] [CrossRef] [PubMed]
- Li, R.Y.; Ma, Y.; Liu, Y.; Liu, M.; Liang, H. Protective effects of Aplysin on lipid metabolism and intestinal mucosal damage of rats exposed by excessive iron. Chin. J. Mar. Drugs 2017, 36, 54–60. [Google Scholar]
- Lieber, C.S. Hepatic, metabolic and toxic effects of ethanol: 1991 update. Alcohol. Clin. Exp. Res. 1991, 15, 573–592. [Google Scholar] [CrossRef] [PubMed]
- Rao, R. Endotoxemia and gut barrier dysfunction in alcoholic liver disease. Hepatology 2009, 50, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Eng, R.H.; Campos, J.M.; Chmel, H. D-lactic acid measurements in the diagnosis of bacterial infections. J. Clin. Microbiol. 1989, 27, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Qiu, P.; Wang, J.; Niu, C.; Pan, S. Effects of compound Ginkgo biloba on intestinal permeability in rats with alcohol-induced liver injury. Food Funct. 2015, 6, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.; Verma, A.K.; Ahuja, V.; Paul, J. Real-time analysis of mucosal flora in patients with inflammatory bowel disease in India. J. Clin. Microbiol. 2010, 48, 4279–4282. [Google Scholar] [CrossRef] [PubMed]
- Jarvelainen, H.A.; Fang, C.; Ingelman-Sundberg, M.; Lindros, K.O. Effect of chronic coadministration of endotoxin and ethanol on rat liver pathology and proinflammatory and anti-inflammatory cytokines. Hepatology 1999, 29, 1503–1510. [Google Scholar] [CrossRef] [PubMed]
- Sobhani, I.; Tap, J.; Roudot-Thoraval, F.; Roperch, J.P.; Letulle, S.; Langella, P.; Corthier, G.; Tran Van Nhieu, J.; Furet, J.P. Microbial dysbiosis in colorectal cancer (CRC) patients. PLoS ONE 2011, 6, e16393. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K.; et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Blum, S.; Haller, D.; Pfeifer, A.; Schiffrin, E.J. Probiotics and immune response. Clin. Rev. Allergy Immunol. 2002, 22, 287–309. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, Y.; Kunitoh-Asari, A.; Hayakawa, K.; Imai, S.; Kasuya, K.; Abe, K.; Adachi, Y.; Fukudome, S.; Takahashi, Y.; Hachimura, S. Oral administration of Lactobacillus plantarum strain AYA enhances IgA secretion and provides survival protection against influenza virus infection in mice. PLoS ONE 2014, 9, e86416. [Google Scholar] [CrossRef] [PubMed]
- Chiu, W.C.; Huang, Y.L.; Chen, Y.L.; Peng, H.C.; Liao, W.H.; Chuang, H.L.; Chen, J.R.; Yang, S.C. Synbiotics reduce ethanol-induced hepatic steatosis and inflammation by improving intestinal permeability and microbiota in rats. Food Funct. 2015, 6, 1692–1700. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, C.B.; Farhadi, A.; Jakate, S.M.; Tang, Y.; Shaikh, M.; Keshavarzian, A. Lactobacillus GG treatment ameliorates alcohol-induced intestinal oxidative stress, gut leakiness, and liver injury in a rat model of alcoholic steatohepatitis. Alcohol 2009, 43, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Dostal, A.; Fehlbaum, S.; Chassard, C.; Zimmermann, M.B.; Lacroix, C. Low iron availability in continuous in vitro colonic fermentations induces strong dysbiosis of the child gut microbial consortium and a decrease in main metabolites. FEMS Microbiol. Ecol. 2013, 83, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.B.; Chassard, C.; Rohner, F.; N’Goran, E.K.; Nindjin, C.; Dostal, A.; Utzinger, J.; Ghattas, H.; Lacroix, C.; Hurrell, R.F. The effects of iron fortification on the gut microbiota in African children: A randomized controlled trial in Cote d'Ivoire. Am. J. Clin. Nutr. 2010, 92, 1406–1415. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Li, Y.; Yu, H.; Gao, C.; Liu, L.; Chen, S.; Xing, M.; Liu, L.; Yao, P. Quercetin prevents ethanol-induced iron overload by regulating hepcidin through the BMP6/SMAD4 signaling pathway. J. Nutr. Biochem. 2014, 25, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, G.N.; Dominitz, J.A.; Weiss, N.S.; Heagerty, P.J.; Kowdley, K.V. The effect of alcohol consumption on the prevalence of iron overload, iron deficiency, and iron deficiency anemia. Gastroenterology 2004, 126, 1293–1301. [Google Scholar] [CrossRef] [PubMed]
- Duane, P.; Raja, K.B.; Simpson, R.J.; Peters, T.J. Intestinal iron absorption in chronic alcoholics. Alcohol Alcohol. 1992, 27, 539–544. [Google Scholar] [PubMed]
- Han, W.; Wang, C.; Su, C.; Xu, X. Effect of higher iron in diet on iron levels and hepcidin mRNA expression levels in rats. Wei Sheng Yan Jiu 2008, 37, 474–476. [Google Scholar] [PubMed]
- Sumida, Y.; Nakashima, T.; Yoh, T.; Kakisaka, Y.; Nakajima, Y.; Ishikawa, H.; Mitsuyoshi, H.; Okanoue, T.; Nakamura, H.; Yodoi, J. Serum thioredoxin elucidates the significance of serum ferritin as a marker of oxidative stress in chronic liver diseases. Liver 2001, 21, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Zoller, H.; Koch, R.O.; Theurl, I.; Obrist, P.; Pietrangelo, A.; Montosi, G.; Haile, D.J.; Vogel, W.; Weiss, G. Expression of the duodenal iron transporters divalent-metal transporter 1 and ferroportin 1 in iron deficiency and iron overload. Gastroenterology 2001, 120, 1412–1419. [Google Scholar] [CrossRef] [PubMed]
- Harrison-Findik, D.D. Is the iron regulatory hormone hepcidin a risk factor for alcoholic liver disease? World J. Gastroenterol. 2009, 15, 1186–1193. [Google Scholar] [CrossRef] [PubMed]
- Harrison-Findik, D.D.; Schafer, D.; Klein, E.; Timchenko, N.A.; Kulaksiz, H.; Clemens, D.; Fein, E.; Andriopoulos, B.; Pantopoulos, K.; Gollan, J. Alcohol metabolism-mediated oxidative stress down-regulates hepcidin transcription and leads to increased duodenal iron transporter expression. J. Biol. Chem. 2006, 281, 22974–22982. [Google Scholar] [CrossRef] [PubMed]
- Harrison-Findik, D.D.; Klein, E.; Crist, C.; Evans, J.; Timchenko, N.; Gollan, J. Iron-mediated regulation of liver hepcidin expression in rats and mice is abolished by alcohol. Hepatology 2007, 46, 1979–1985. [Google Scholar] [CrossRef] [PubMed]
- Dostalikova-Cimburova, M.; Balusikova, K.; Kratka, K.; Chmelikova, J.; Hejda, V.; Hnanicek, J.; Neubauerova, J.; Vranova, J.; Kovar, J.; Horak, J. Role of duodenal iron transporters and hepcidin in patients with alcoholic liver disease. J. Cell Mol. Med. 2014, 18, 1840–1850. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, G.; Roque, M.E. Iron overload induces changes of pancreatic and duodenal divalent metal transporter 1 and prohepcidin expression in mice. Acta Histochem. 2014, 116, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| The Composition of the Diets | (g/kg) |
|---|---|
| Sucrose | 500.0 |
| Casein | 200.0 |
| Corn starch | 150.0 |
| Cellulose | 50.0 |
| Corn oil | 50.0 |
| Mineral mix, AIN-76 (170,915) | 35.0 |
| Vitamin mix, AIN-76A (40,077) | 10.0 |
| DL-Methionine | 3.0 |
| Choline bitartrate | 2.0 |
| Ethoxyquin, antioxidant | 0.01 |
| Bacterial Species | Primer Sequences (5′3′) | Lengths (bp) | Annealing Temperature (°C) |
|---|---|---|---|
| Escherichia coil | F: 5′-GTTAATACCTTTGCTCATTGA-3′ | 340 | 51 |
| R: 5′-ACCAGGGTATCTTAATCCTGTT-3′ | |||
| Enterococcus | F: 5′-ACTCGTTGTACTTCCCATTGT-3′ | 144 | 52 |
| R:5′-CCCTTATTGTTAGTTGCCATCATT-3′ | |||
| Bifidobacterium | F: 5′-GGGTGGTAATGCCGGATG-3′ | 442 | 61 |
| R: 5′-TAAGCGATGGACTTTCACACC-3′ | |||
| Lactobacillus | F: 5′- AGCAGTAGGGAATCTTCCA-3′ | 341 | 55 |
| R: 5′-CACCGCTACACATGGAG-3′ | |||
| Bacteroides fragilis | F: 5′- CTGAACCAGCCAAGTAGCG-3′ | 230 | 62 |
| R:5′-CCGCAAACTTTCACAACTGACTTA-3′ | |||
| Clostridium tender | F: 5′-GCACAAGCAGTGGAGT-3′ | 246 | 58 |
| R: 5′-CTTCCTCCGTTTTGTCAA-3′ |
| Group (n = 10) | Food Intake (g) | Initial Body Weight (g) | Final Body Weight (g) |
|---|---|---|---|
| C | 35.93 + 1.76 | 233.40 + 12.35 | 398.00 + 12.75 |
| EI | 34.72 + 1.36 | 236.55 + 8.23 | 350.30 + 13.76 # |
| AEI | 35.50 + 1.80 | 238.00 + 12.14 | 379.10 + 14.91 #,* |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Li, R.; Liu, Y.; Liu, M.; Liang, H. Protective Effect of Aplysin Supplementation on Intestinal Permeability and Microbiota in Rats Treated with Ethanol and Iron. Nutrients 2018, 10, 681. https://doi.org/10.3390/nu10060681
Ma Y, Li R, Liu Y, Liu M, Liang H. Protective Effect of Aplysin Supplementation on Intestinal Permeability and Microbiota in Rats Treated with Ethanol and Iron. Nutrients. 2018; 10(6):681. https://doi.org/10.3390/nu10060681
Chicago/Turabian StyleMa, Yan, Ruiying Li, Ying Liu, Man Liu, and Hui Liang. 2018. "Protective Effect of Aplysin Supplementation on Intestinal Permeability and Microbiota in Rats Treated with Ethanol and Iron" Nutrients 10, no. 6: 681. https://doi.org/10.3390/nu10060681
APA StyleMa, Y., Li, R., Liu, Y., Liu, M., & Liang, H. (2018). Protective Effect of Aplysin Supplementation on Intestinal Permeability and Microbiota in Rats Treated with Ethanol and Iron. Nutrients, 10(6), 681. https://doi.org/10.3390/nu10060681