Pro-Apoptotic and Anti-Cancer Properties of Diosgenin: A Comprehensive and Critical Review


Abstract
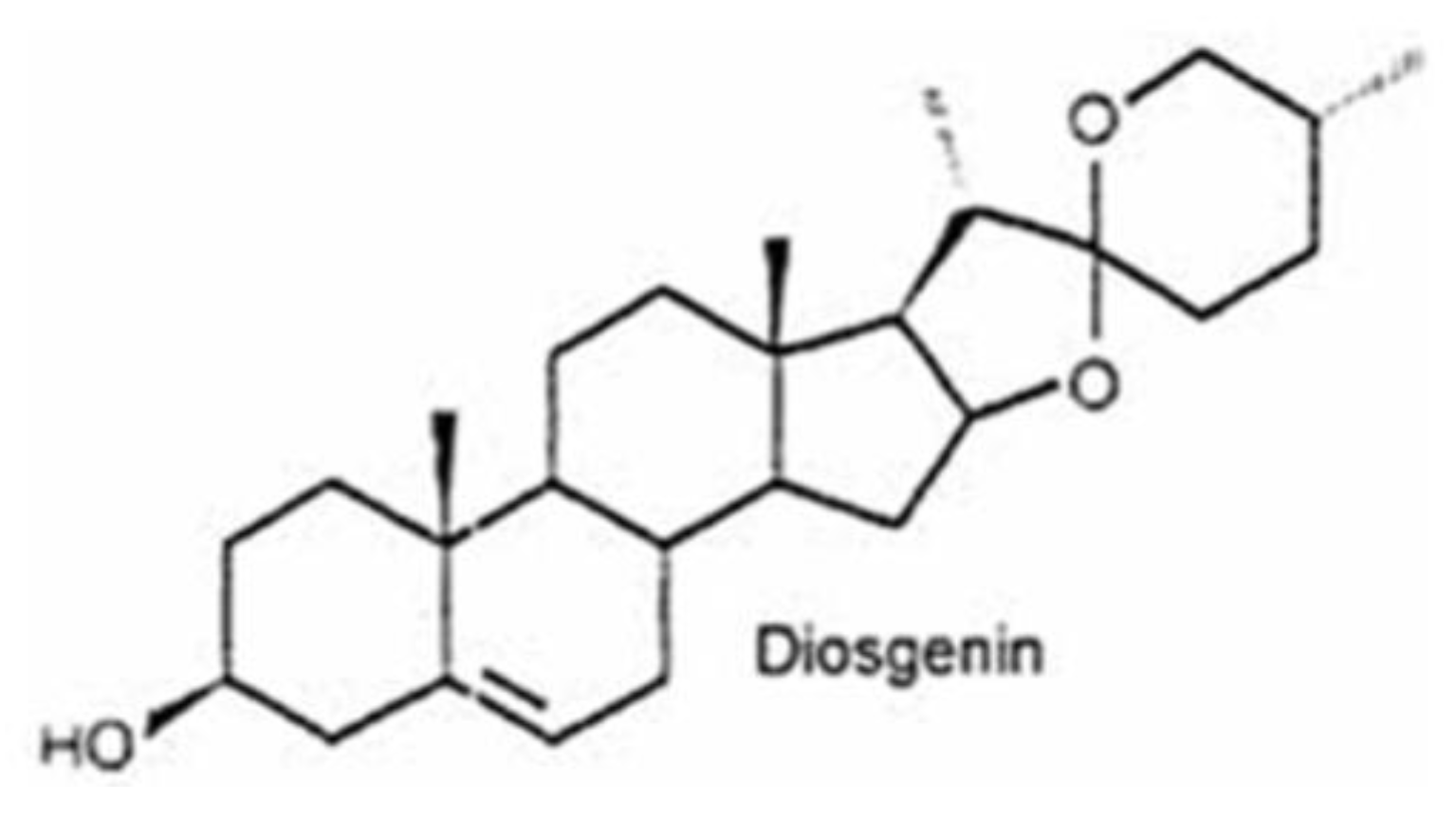
1. Introduction
2. Fenugreek Seed Bioactive Compounds
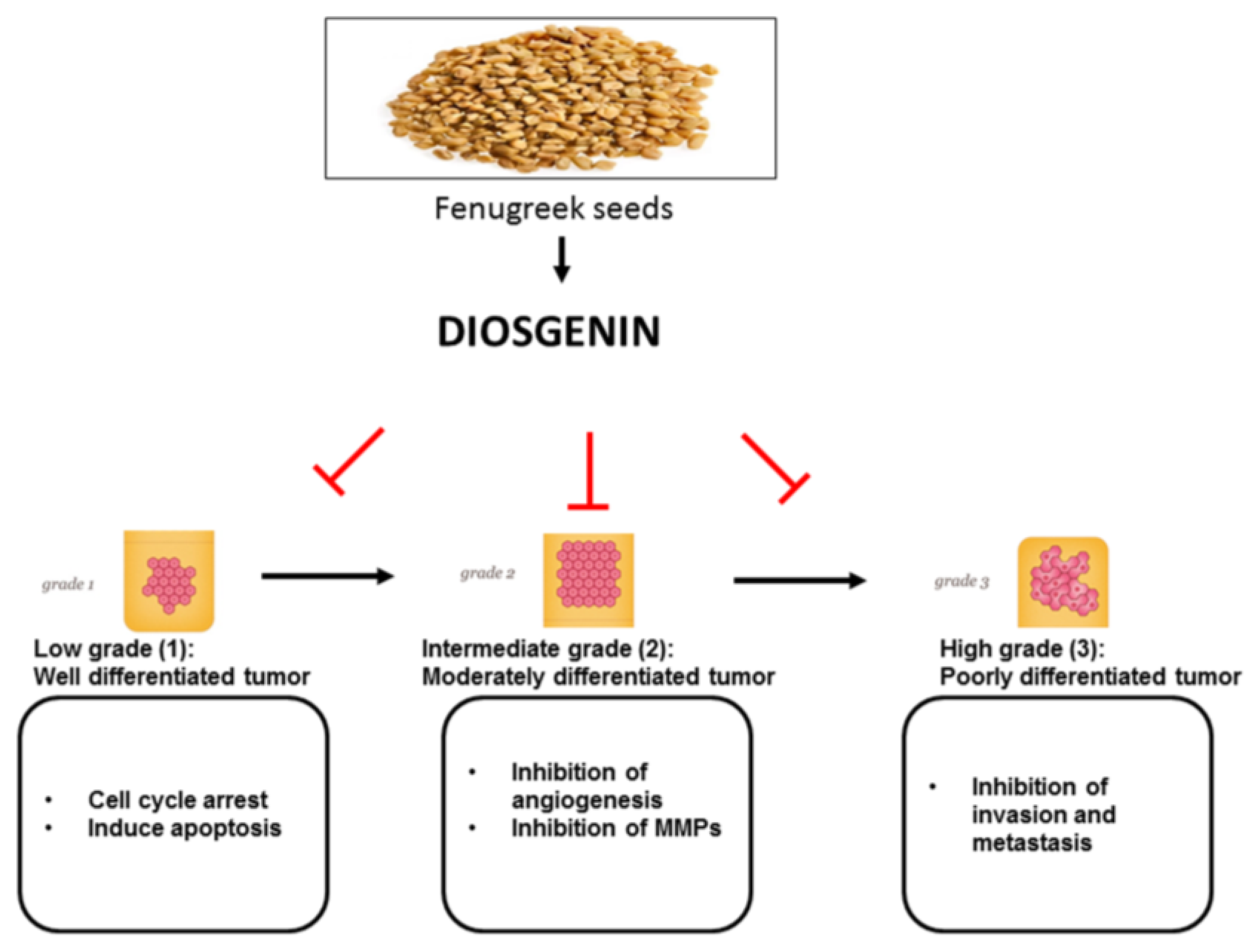
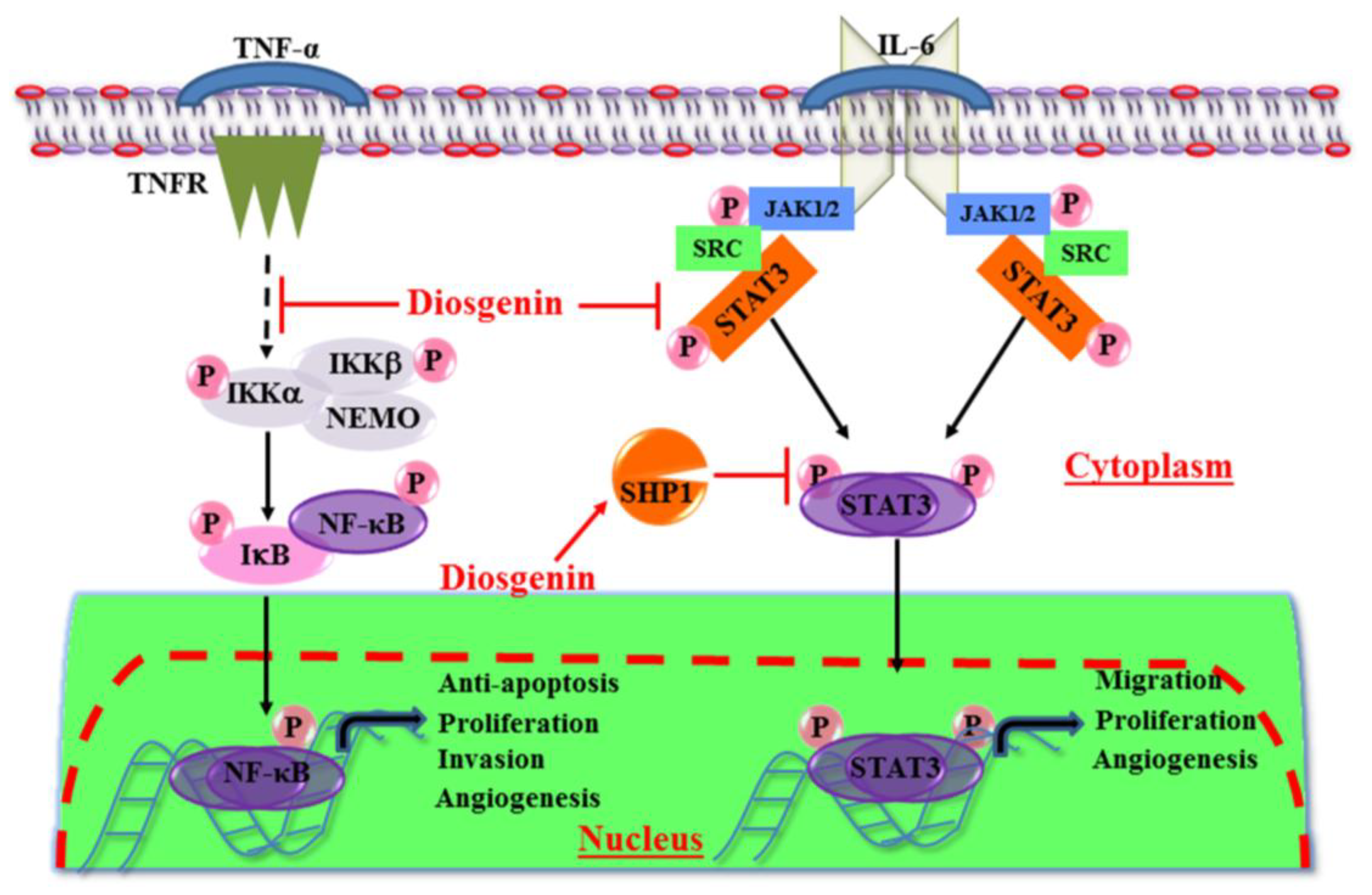
3. In Vitro Anti-Cancer Effects of Diosgenin
4. In Vivo Anti-Cancer Effects of Diosgenin
5. Semisynthetic Derivatives of Diosgenin That Exhibit Anti-Cancer Activity
6. Conclusions and Future Perspectives
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2017. CA Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Rethinking the war on cancer. Lancet 2014, 383, 558–563. [Google Scholar] [CrossRef]
- Floor, S.L.; Dumont, J.E.; Maenhaut, C.; Raspe, E. Hallmarks of cancer: Of all cancer cells, all the time? Trends Mol. Med. 2012, 18, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Pietras, K.; Ostman, A. Hallmarks of cancer: Interactions with the tumor stroma. Exp. Cell Res. 2010, 316, 1324–1331. [Google Scholar] [CrossRef] [PubMed]
- Massague, J.; Obenauf, A.C. Metastatic colonization by circulating tumour cells. Nature 2016, 529, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Chaffer, C.L.; Weinberg, R.A. How does multistep tumorigenesis really proceed? Cancer Discov. 2015, 5, 22–24. [Google Scholar] [CrossRef] [PubMed]
- Iwatsuki, M.; Mimori, K.; Yokobori, T.; Ishi, H.; Beppu, T.; Nakamori, S.; Baba, H.; Mori, M. Epithelial-mesenchymal transition in cancer development and its clinical significance. Cancer Sci. 2010, 101, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Almendro, V.; Marusyk, A.; Polyak, K. Cellular heterogeneity and molecular evolution in cancer. Annu. Rev. Pathol. 2013, 8, 277–302. [Google Scholar] [CrossRef] [PubMed]
- Steeg, P.S. Targeting metastasis. Nat. Rev. Cancer 2016, 16, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Dabrosin, C.; Yin, X.; Fuster, M.M.; Arreola, A.; Rathmell, W.K.; Generali, D.; Nagaraju, G.P.; El-Rayes, B.; Ribatti, D.; et al. Broad targeting of angiogenesis for cancer prevention and therapy. Semin. Cancer Biol. 2015, 35, S224–S243. [Google Scholar] [CrossRef] [PubMed]
- Sethi, G.; Ahn, K.S.; Aggarwal, B.B. Targeting nuclear factor-kappa B activation pathway by thymoquinone: Role in suppression of antiapoptotic gene products and enhancement of apoptosis. Mol. Cancer Res. 2008, 6, 1059–1070. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.H.; Sethi, G.; Kuo, P.L. Novel medicines and strategies in cancer treatment and prevention. BioMed Res. Int. 2014, 2014, 474078. [Google Scholar] [CrossRef] [PubMed]
- Ahn, K.S.; Sethi, G.; Aggarwal, B.B. Nuclear factor-kappa B: From clone to clinic. Curr. Mol. Med. 2007, 7, 619–637. [Google Scholar] [CrossRef] [PubMed]
- Chai, E.Z.; Shanmugam, M.K.; Arfuso, F.; Dharmarajan, A.; Wang, C.; Kumar, A.P.; Samy, R.P.; Lim, L.H.; Wang, L.; Goh, B.C.; et al. Targeting transcription factor stat3 for cancer prevention and therapy. Pharmacol. Ther. 2016, 162, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhang, J.; Arfuso, F.; Chinnathambi, A.; Zayed, M.E.; Alharbi, S.A.; Kumar, A.P.; Ahn, K.S.; Sethi, G. NF-kappaB in cancer therapy. Arch. Toxicol. 2015, 89, 711–731. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Kannaiyan, R.; Sethi, G. Targeting cell signaling and apoptotic pathways by dietary agents: Role in the prevention and treatment of cancer. Nutr. Cancer 2011, 63, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Deorukhkar, A.; Krishnan, S.; Sethi, G.; Aggarwal, B.B. Back to basics: How natural products can provide the basis for new therapeutics. Expert Opin. Investig. Drugs 2007, 16, 1753–1773. [Google Scholar] [CrossRef] [PubMed]
- Kneller, R. The importance of new companies for drug discovery: Origins of a decade of new drugs. Nat. Rev. Drug Discov. 2010, 9, 867–882. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A.; Sethi, G. Bioactive natural products in cancer prevention and therapy: Progress and promise. Semin. Cancer Biol. 2016, 40–41, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J. Developing natural product drugs: Supply problems and how they have been overcome. Pharmacol. Ther. 2016, 162, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J. Bioactive cyclic molecules and drug design. Expert Opin. Drug Discov. 2018, 13, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Lee, J.H.; Chai, E.Z.; Kanchi, M.M.; Kar, S.; Arfuso, F.; Dharmarajan, A.; Kumar, A.P.; Ramar, P.S.; Looi, C.Y.; et al. Cancer prevention and therapy through the modulation of transcription factors by bioactive natural compounds. Semin. Cancer Biol. 2016, 40–41, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Nguyen, A.H.; Kumar, A.P.; Tan, B.K.; Sethi, G. Targeted inhibition of tumor proliferation, survival, and metastasis by pentacyclic triterpenoids: Potential role in prevention and therapy of cancer. Cancer Lett. 2012, 320, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Warrier, S.; Kumar, A.P.; Sethi, G.; Arfuso, F. Potential role of natural compounds as anti-angiogenic agents in cancer. Curr. Vasc. Pharmacol. 2017, 15, 503–519. [Google Scholar] [CrossRef] [PubMed]
- Mishra, K.P.; Ganju, L.; Sairam, M.; Banerjee, P.K.; Sawhney, R.C. A review of high throughput technology for the screening of natural products. Biomed. Pharmacother. 2008, 62, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Grothaus, P.G.; Newman, D.J. Impact of natural products on developing new anti-cancer agents. Chem. Rev. 2009, 109, 3012–3043. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J. Natural products as leads to potential drugs: An old process or the new hope for drug discovery? J. Med. Chem. 2008, 51, 2589–2599. [Google Scholar] [CrossRef] [PubMed]
- Chai, E.Z.; Siveen, K.S.; Shanmugam, M.K.; Arfuso, F.; Sethi, G. Analysis of the intricate relationship between chronic inflammation and cancer. Biochem. J. 2015, 468, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sung, B.; Prasad, S.; Yadav, V.R.; Aggarwal, B.B. Cancer cell signaling pathways targeted by spice-derived nutraceuticals. Nutr. Cancer 2012, 64, 173–197. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Current status of marine-derived compounds as warheads in anti-tumor drug candidates. Mar. Drugs 2017, 15, 99. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Drugs and drug candidates from marine sources: An assessment of the current “state of play”. Planta Med. 2016, 82, 775–789. [Google Scholar] [CrossRef] [PubMed]
- El Bairi, K.; Ouzir, M.; Agnieszka, N.; Khalki, L. Anticancer potential of trigonella foenum graecum: Cellular and molecular targets. Biomed. Pharmacother. 2017, 90, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Shabbeer, S.; Sobolewski, M.; Anchoori, R.K.; Kachhap, S.; Hidalgo, M.; Jimeno, A.; Davidson, N.; Carducci, M.A.; Khan, S.R. Fenugreek: A naturally occurring edible spice as an anticancer agent. Cancer Biol. Ther. 2009, 8, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Kunnumakkara, A.B.; Harikumar, K.B.; Tharakan, S.T.; Sung, B.; Anand, P. Potential of spice-derived phytochemicals for cancer prevention. Planta Med. 2008, 74, 1560–1569. [Google Scholar] [CrossRef] [PubMed]
- Kenny, O.; Smyth, T.J.; Hewage, C.M.; Brunton, N.P. Antioxidant properties and quantitative UPLC-MS analysis of phenolic compounds from extracts of Fenugreek (Trigonella foenum-graecum) seeds and bitter melon (Momordica charantia) fruit. Food Chem. 2013, 141, 4295–4302. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.A.; Kumar, P. Fenugreek: A review on its nutraceutical properties and utilization in various food products. J. Saudi Soc. Agric. Sci. 2018, 17, 97–106. [Google Scholar] [CrossRef]
- Kim, J.K.; Park, S.U. An update on the biological and pharmacological activities of diosgenin. EXCLI J. 2018, 17, 24–28. [Google Scholar] [PubMed]
- Bahmani, M.; Shirzad, H.; Mirhosseini, M.; Mesripour, A.; Rafieian-Kopaei, M. A review on ethnobotanical and therapeutic uses of Fenugreek (Trigonella foenum-graceum L.). J. Evid. Based Complement. Altern. Med. 2016, 21, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, K. Fenugreek (Trigonella foenum-graecum): A review of health beneficial physiological effects. Food Rev. Int. 2006, 22, 203–224. [Google Scholar] [CrossRef]
- Jesus, M.; Martins, A.P.; Gallardo, E.; Silvestre, S. Diosgenin: Recent highlights on pharmacology and analytical methodology. J. Anal. Methods Chem. 2016, 2016, 4156293. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.F.; Weng, C.J.; Sethi, G.; Hu, D.N. Natural bioactives and phytochemicals serve in cancer treatment and prevention. Evid. Based Complement. Altern. Med. 2013, 2013, 698190. [Google Scholar] [CrossRef] [PubMed]
- Djerassi, C.; Rosenkranz, G.; Pataki, J.; Kaufmann, S. Steroids, xxvii. Synthesis of allopregnane-3beta, 11beta, 17alpha-, 20beta, 21-pentol from cortisone and diosgenin. J. Biol. Chem. 1952, 194, 115–118. [Google Scholar] [PubMed]
- Patel, K.; Gadewar, M.; Tahilyani, V.; Patel, D.K. A review on pharmacological and analytical aspects of diosmetin: A concise report. Chin. J. Integr. Med. 2013, 19, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tang, Y.M.; Yu, S.L.; Han, Y.W.; Kou, J.P.; Liu, B.L.; Yu, B.Y. Advances in the pharmacological activities and mechanisms of diosgenin. Chin. J. Nat. Med. 2015, 13, 578–587. [Google Scholar] [CrossRef]
- Tang, Y.; Yi, T.; Chen, H.; Zhao, Z.; Liang, Z.; Chen, H. Quantitative comparison of multiple components in dioscorea nipponica and d. Panthaica by ultra-high performance liquid chromatography coupled with quadrupole time-of-flight mass spectrometry. Phytochem. Anal. 2013, 24, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.N.; Pang, Y.X.; He, X.C.; Zhang, Y.Z.; Zhang, J.Y.; Zhao, Z.Z.; Yi, T.; Chen, H.B. UPLC-QTOF-MS identification of metabolites in rat biosamples after oral administration of dioscorea saponins: A comparative study. J. Ethnopharmacol. 2015, 165, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Ciura, J.; Szeliga, M.; Grzesik, M.; Tyrka, M. Next-generation sequencing of representational difference analysis products for identification of genes involved in diosgenin biosynthesis in Fenugreek (Trigonella foenum-graecum). Planta 2017, 245, 977–991. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, K.; Ghosh, A.; Kumar, V.; Chaudhary, S.; Srivastava, N.; Katudia, K.; Tiwari, T.; Chikara, S.K. De novo transcriptome sequencing in Trigonella foenum-graecum L. to identify genes involved in the biosynthesis of diosgenin. Plant. Genome 2013, 6, 1–11. [Google Scholar] [CrossRef]
- Chiang, C.T.; Way, T.D.; Tsai, S.J.; Lin, J.K. Diosgenin, a naturally occurring steroid, suppresses fatty acid synthase expression in HER2-overexpressing breast cancer cells through modulating Akt, mTOR and JNK phosphorylation. FEBS Lett. 2007, 581, 5735–5742. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Fernandez, P.P.; Rajendran, P.; Hui, K.M.; Sethi, G. Diosgenin, a steroidal saponin, inhibits STAT3 signaling pathway leading to suppression of proliferation and chemosensitization of human hepatocellular carcinoma cells. Cancer Lett. 2010, 292, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Moalic, S.; Liagre, B.; Corbiere, C.; Bianchi, A.; Dauca, M.; Bordji, K.; Beneytout, J.L. A plant steroid, diosgenin, induces apoptosis, cell cycle arrest and cox activity in osteosarcoma cells. FEBS Lett. 2001, 506, 225–230. [Google Scholar] [CrossRef]
- Chen, P.S.; Shih, Y.W.; Huang, H.C.; Cheng, H.W. Diosgenin, a steroidal saponin, inhibits migration and invasion of human prostate cancer pc-3 cells by reducing matrix metalloproteinases expression. PLoS ONE 2011, 6, e20164. [Google Scholar] [CrossRef] [PubMed]
- Raju, J.; Rao, C.V. Diosgenin, a steroid saponin constituent of yams and fenugreek: Emerging evidence for applications. In Bioactive Compounds Medicine in Phytomedicine; Rasooli, I., Ed.; InTech.: London, UK, 2012; pp. 125–142. [Google Scholar]
- Shishodia, S.; Aggarwal, B.B. Diosgenin inhibits osteoclastogenesis, invasion, and proliferation through the downregulation of Akt, I kappa B kinase activation and NF-kappa B-regulated gene expression. Oncogene 2006, 25, 1463–1473. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Dey, K.K.; Dey, G.; Pal, I.; Majumder, A.; MaitiChoudhury, S.; kundu, S.C.; Mandal, M. Antineoplastic and apoptotic potential of traditional medicines thymoquinone and diosgenin in squamous cell carcinoma. PLoS ONE 2012, 7, e46641. [Google Scholar] [CrossRef] [PubMed]
- Lepage, C.; Leger, D.Y.; Bertrand, J.; Martin, F.; Beneytout, J.L.; Liagre, B. Diosgenin induces death receptor-5 through activation of p38 pathway and promotes trail-induced apoptosis in colon cancer cells. Cancer Lett. 2011, 301, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Romero-Hernandez, L.L.; Merino-Montiel, P.; Montiel-Smith, S.; Meza-Reyes, S.; Vega-Baez, J.L.; Abasolo, I.; Schwartz, S., Jr.; Lopez, O.; Fernandez-Bolanos, J.G. Diosgenin-based thio(seleno)ureas and triazolyl glycoconjugates as hybrid drugs. Antioxidant and antiproliferative profile. Eur. J. Med. Chem. 2015, 99, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Liao, A.; Lee, K.K.; Ban, J.S.; Yang, H.S.; Im, Y.J.; Chun, C. Design, synthesis of methotrexate-diosgenin conjugates and biological evaluation of their effect on methotrexate transport-resistant cells. Steroids 2016, 116, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Fan, J.; Wang, Q.; Ju, D.; Feng, M.; Li, J.; Guan, Z.B.; An, D.; Wang, X.; Ye, L. Diosgenin induces ros-dependent autophagy and cytotoxicity via mtor signaling pathway in chronic myeloid leukemia cells. Phytomedicine 2016, 23, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Jagadeesan, J.; Langeswaran, K.; Gowthamkumar, S.; Balasubramanian, M.P. Diosgenin exhibits beneficial efficiency on human mammary carcinoma cell line MCF-7 and against N-nitroso-N-methylurea (NMU) induced experimental mammary carcinoma. Biomed. Prev. Nutr. 2013, 3, 381–388. [Google Scholar] [CrossRef]
- Albini, A.; Tosetti, F.; Li, V.W.; Noonan, D.M.; Li, W.W. Cancer prevention by targeting angiogenesis. Nat. Rev. Clin. Oncol. 2012, 9, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Tumor angiogenesis: Therapeutic implications. N. Engl. J. Med. 1971, 285, 1182–1186. [Google Scholar] [PubMed]
- Folkman, J. Role of angiogenesis in tumor growth and metastasis. Semin. Oncol. 2002, 29, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Angiogenesis: An organizing principle for drug discovery? Nat. Rev. Drug Discov. 2007, 6, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Selim, S.; Al Jaouni, S. Anti-inflammatory, antioxidant and antiangiogenic activities of diosgenin isolated from traditional medicinal plant, costus speciosus (Koen ex.Retz.) Sm. Nat. Prod. Res. 2016, 30, 1830–1833. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Chen, H.; Li, G.; Zhu, H.; Gao, Y.; Zhang, L.; Sun, J. Diosgenin inhibits the migration of human breast cancer MDA-MB-231 cells by suppressing VAV2 activity. Phytomedicine 2014, 21, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.J.; Tang, Q.J.; Zhang, C.A.; Qin, Z.F.; Pang, B.; Wei, P.K.; Liu, B.; Chou, Y.N. Anti-proliferation and anti-invasion effects of diosgenin on gastric cancer BGC-823 cells with HIF-1alpha shrnas. Int. J. Mol. Sci. 2012, 13, 6521–6533. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Koduru, S.; Kumar, R.; Venguswamy, G.; Kyprianou, N.; Damodaran, C. Diosgenin targets AKT-mediated prosurvival signaling in human breast cancer cells. Int. J. Cancer 2009, 125, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Sowmyalakshmi, S.; Ranga, R.; Gairola, C.G.; Chendil, D. Effect of diosgenin (Fenugreek) on breast cancer cells. Proc. Am. Assoc. Cancer Res. 2005, 46, 1382. [Google Scholar]
- Corbiere, C.; Liagre, B.; Bianchi, A.; Bordji, K.; Dauca, M.; Netter, P.; Beneytout, J.L. Different contribution of apoptosis to the antiproliferative effects of diosgenin and other plant steroids, hecogenin and tigogenin, on human 1547 osteosarcoma cells. Int. J. Oncol. 2003, 22, 899–905. [Google Scholar] [CrossRef] [PubMed]
- Liagre, B.; Bertrand, J.; Leger, D.Y.; Beneytout, J.L. Diosgenin, a plant steroid, induces apoptosis in COX-2 deficient K562 cells with activation of the p38 map kinase signalling and inhibition of NF-kappaB binding. Int. J. Mol. Med. 2005, 16, 1095–1101. [Google Scholar] [CrossRef] [PubMed]
- Leger, D.Y.; Liagre, B.; Corbiere, C.; Cook-Moreau, J.; Beneytout, J.L. Diosgenin induces cell cycle arrest and apoptosis in hel cells with increase in intracellular calcium level, activation of CPLA2 and COX-2 overexpression. Int. J. Oncol. 2004, 25, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Corbiere, C.; Liagre, B.; Terro, F.; Beneytout, J.L. Induction of antiproliferative effect by diosgenin through activation of p53, release of apoptosis-inducing factor (AIF) and modulation of caspase-3 activity in different human cancer cells. Cell. Res. 2004, 14, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Cai, B.; Cui, C.B.; Liu, H.W.; Wu, C.F.; Yao, X.S. Diosgenin-3-O-alpha-l-rhamnopyranosyl-(1→4)-beta-d-glucopyranoside obtained as a new anticancer agent from dioscorea futschauensis induces apoptosis on human colon carcinoma HCT-15 cells via mitochondria-controlled apoptotic pathway. J. Asian Nat. Prod. Res. 2004, 6, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Raju, J.; Patlolla, J.M.; Swamy, M.V.; Rao, C.V. Diosgenin, a steroid saponin of Trigonella foenum graecum (Fenugreek), inhibits azoxymethane-induced aberrant crypt foci formation in F344 rats and induces apoptosis in HT-29 human colon cancer cells. Cancer Epidemiol. Biomarkers Prev. 2004, 13, 1392–1398. [Google Scholar] [PubMed]
- Malisetty, V.S.; Patlolla, J.M.R.; Raju, J.; Marcus, L.A.; Choi, C.L.; Rao, C.V. Chemoprevention of colon cancer by diosgenin, a steroidal saponin constituent of fenugreek. Proc. Am. Assoc. Cancer Res. 2005, 46, 2473. [Google Scholar]
- Miyoshi, N.; Nagasawa, T.; Mabuchi, R.; Yasui, Y.; Wakabayashi, K.; Tanaka, T.; Ohshima, H. Chemoprevention of azoxymethane/dextran sodium sulfate-induced mouse colon carcinogenesis by freeze-dried yam sanyaku and its constituent diosgenin. Cancer Prev. Res. (Phila.) 2011, 4, 924–934. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.L.; Zhang, Y.J.; Gao, W.Y.; Man, S.L.; Wang, Y. In vitro and in vivo anticancer activity of steroid saponins of paris polyphylla var. Yunnanensis. Exp. Oncol. 2009, 31, 27–32. [Google Scholar] [PubMed]
- Marker, R.E.; Sterols, C.V. The preparation of testosterone and related compounds from sarsasapogenin and diosgenin. J. Am. Chem. Soc. 1940, 62, 2543–2547. [Google Scholar] [CrossRef]
- Tietze, L.F.; Bell, H.P.; Chandrasekhar, S. Natural product hybrids as new leads for drug discovery. Angew. Chem. Int. Ed. 2003, 42, 3996–4028. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Sanchez, L.; Hernandez-Linares, M.G.; Escobar, M.L.; Lopez-Munoz, H.; Zenteno, E.; Fernandez-Herrera, M.A.; Guerrero-Luna, G.; Carrasco-Carballo, A.; Sandoval-Ramirez, J. Antiproliferative, cytotoxic, and apoptotic activity of steroidal oximes in cervicouterine cell lines. Molecules 2016, 21, 1533. [Google Scholar] [CrossRef] [PubMed]
- Masood Ur, R.; Mohammad, Y.; Fazili, K.M.; Bhat, K.A.; Ara, T. Synthesis and biological evaluation of novel 3-O-tethered triazoles of diosgenin as potent antiproliferative agents. Steroids 2017, 118, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Xie, F.; Zhao, G.D.; Wang, D.F.; Lou, H.X.; Liu, Z.P. Synthetic studies towards 1alpha-hydroxysolasodine from diosgenin and the unexpected tetrahydrofuran ring opening in the birch reduction process. Steroids 2015, 104, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Hamid, A.A.; Kaushal, T.; Ashraf, R.; Singh, A.; Chand Gupta, A.; Prakash, O.; Sarkar, J.; Chanda, D.; Bawankule, D.U.; Khan, F.; et al. (22beta,25R)-3beta-hydroxy-spirost-5-en-7-iminoxy-heptanoic acid exhibits anti-prostate cancer activity through caspase pathway. Steroids 2017, 119, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; More, P.; Derle, A.; Kitture, R.; Kale, T.; Gorain, M.; Avasthi, A.; Markad, P.; Kundu, G.C.; Kale, S.; et al. Diosgenin functionalized iron oxide nanoparticles as novel nanomaterial against breast cancer. J. Nanosci. Nanotechnol. 2015, 15, 9464–9472. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Class of Compounds | Phytochemical Constituents | Reference |
|---|---|---|
| Steroidal sapogenins | Diosgenin, Yamogenin, Smilagenin, Sarsasapogenin, Tigogenin, Neotigogenin, Gitogenin, Yuccagenin, Saponaretin | [40] |
| Flavonoids | Quercetin, Rutin, Vitexin, Isovitexin | [40] |
| Saponins | Graecunins, Fenugrin B, Fenugreekine, Trigofoenosides A–G | [40] |
| Alkaloids | Trimethylamine, Neurin, Trigonelline, Choline, Gentianine, Carpaine, and Betain | [40] |
| Fibers | Gum, Neutral detergent fiber | [40] |
| Lipids | Lipids, Triacylglycerols, Diacylglycerols, Monoacylglycerols, Phosphatidylcholine, Phosphatidylethanolamine, Phosphatidylinositol, Free fatty acids | [40] |
| Others | Coumarin, Amino acids, Vitamins, Minerals. 28% Mucilage; 22% Proteins; 5% of a stronger swelling, Bitter fixed oil | [40] |
| Cancer model | Cell Lines | Diosgenin Dose | Molecular Target | References |
|---|---|---|---|---|
| Breast carcinoma | Estrogen receptor positive and estrogen receptor negative human breast cancer MCF-7 and MDA 231 cells | 20 μM and 30 μM | Inhibition of cell proliferation Induces apoptosis | [72] |
| MDA-MB-231 breast cancer cells | 20 μM, 40 μM, and 60 μM | Downregulation of Bcl2 | [57] | |
| Her2 over-expressing breast cancer cells | 5–20 μM | Modulation of Akt, mTOR, and JNK phosphorylation | [53] | |
| MCF-7 breast cancer cells | 20 μM and 40 μM | Upregulation of p53 tumor suppressor gene | [73] | |
| Hepatocellular carcinoma | C3A, HUH-7, and HepG2 cells | 50 μM and 100 μM | Downregulation of STAT3 signaling pathway Upregulates SH-PTP2 expression Induces apoptosis Potentiates the apoptotic effects of doxorubicin and paclitaxel | [54] |
| Prostate carcinoma | PC3 cells | 5 μM, 10 μM, and 20 μM | Downregulates NF-κB signaling pathway Inhibits matrix metalloproteinases Inhibits invasion and migration of cells | [56] |
| Osteosarcoma | 1547 cells | 40 μM, 80 μM, and 100 μM | Inhibits cell proliferation Induces apoptosis | [55] |
| 1547 cells | 40 μM | Inhibits cell proliferation Induces apoptosis Upregulation of p53 tumor suppressor gene | [74] | |
| Human erythroleukemia | HEL cells, K562 cells | 40 μM | Inhibits NF-κB signaling pathway | [75] |
| HEL cells | 40 μM | Inhibits proliferation Induces apoptosis Upregulation of p21 | [76] | |
| Human Laryngocarcinoma Human Melanoma | HEp-2 cells M4Beu cells | 40 μM | Inhibits cell proliferation Induces caspase-3 dependent apoptosis Upregulates p53 tumor suppressor gene | [77] |
| Human cancer cells | Human epithelial carcinoma cell line (A431), human NSCLC cell line (A549), human ovarian cancer cell line (A2780), Human erythroleukemia (K562) and Dukes’ type C, colorectal adenocarcinoma (HCT-15) | 10 mmol/L | Induces apoptosis via mitochondrial dependent pathway | [78] |
| Multiple myeloma (U266), leukemia (U937), and breast cancer (MCF-7) | 50 μM and 100 μM | Inhibits NF-κB signaling pathway | [58] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sethi, G.; Shanmugam, M.K.; Warrier, S.; Merarchi, M.; Arfuso, F.; Kumar, A.P.; Bishayee, A. Pro-Apoptotic and Anti-Cancer Properties of Diosgenin: A Comprehensive and Critical Review. Nutrients 2018, 10, 645. https://doi.org/10.3390/nu10050645
Sethi G, Shanmugam MK, Warrier S, Merarchi M, Arfuso F, Kumar AP, Bishayee A. Pro-Apoptotic and Anti-Cancer Properties of Diosgenin: A Comprehensive and Critical Review. Nutrients. 2018; 10(5):645. https://doi.org/10.3390/nu10050645
Chicago/Turabian StyleSethi, Gautam, Muthu K. Shanmugam, Sudha Warrier, Myriam Merarchi, Frank Arfuso, Alan Prem Kumar, and Anupam Bishayee. 2018. "Pro-Apoptotic and Anti-Cancer Properties of Diosgenin: A Comprehensive and Critical Review" Nutrients 10, no. 5: 645. https://doi.org/10.3390/nu10050645
APA StyleSethi, G., Shanmugam, M. K., Warrier, S., Merarchi, M., Arfuso, F., Kumar, A. P., & Bishayee, A. (2018). Pro-Apoptotic and Anti-Cancer Properties of Diosgenin: A Comprehensive and Critical Review. Nutrients, 10(5), 645. https://doi.org/10.3390/nu10050645