Capsaicin in Metabolic Syndrome



Abstract
1. Introduction
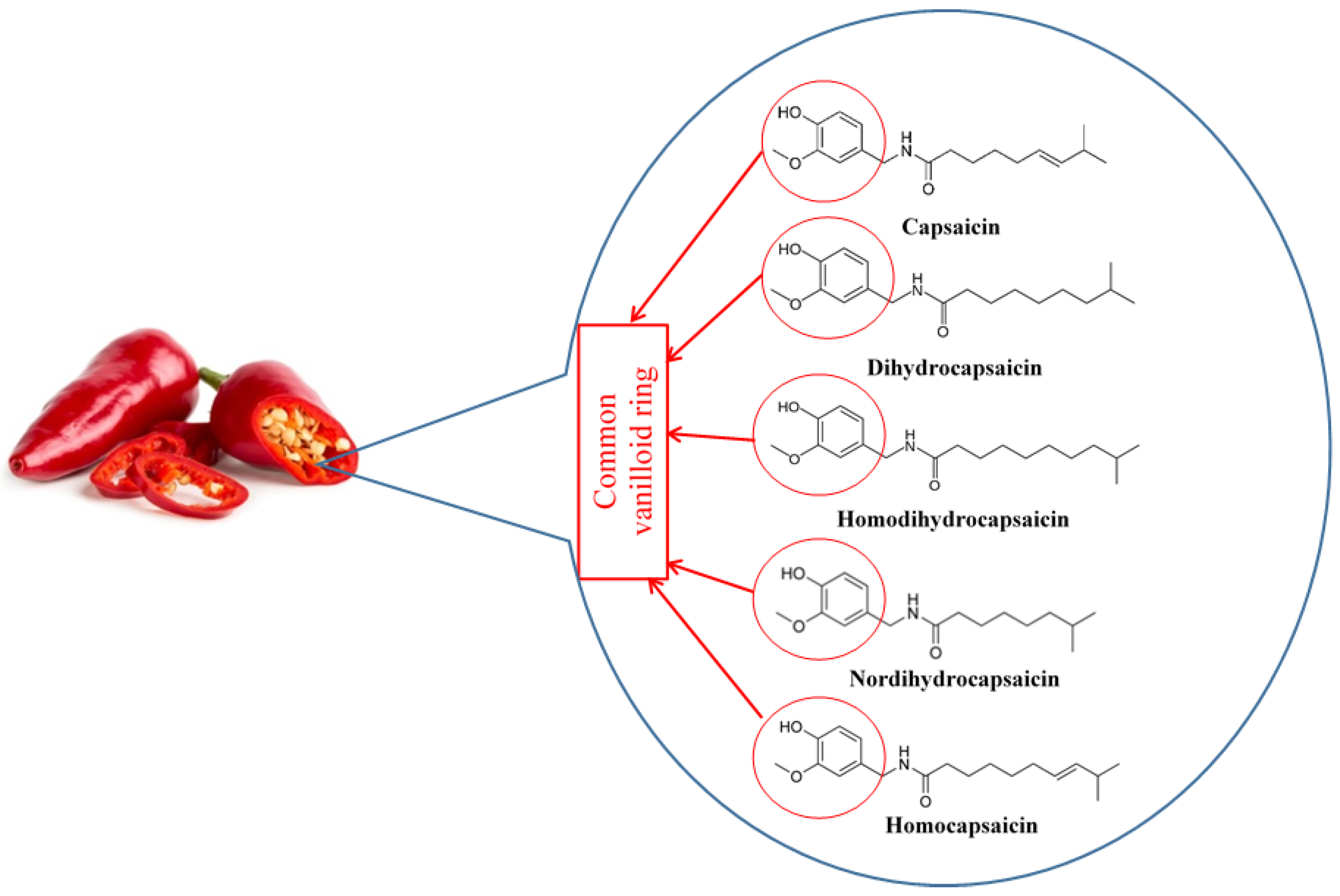
Capsaicin
2. Metabolic Disorders
2.1. Metabolic Syndrome
2.2. Insulin Resistance and Obesity
2.3. Mechanisms for the Development of Insulin Resistance and Obesity
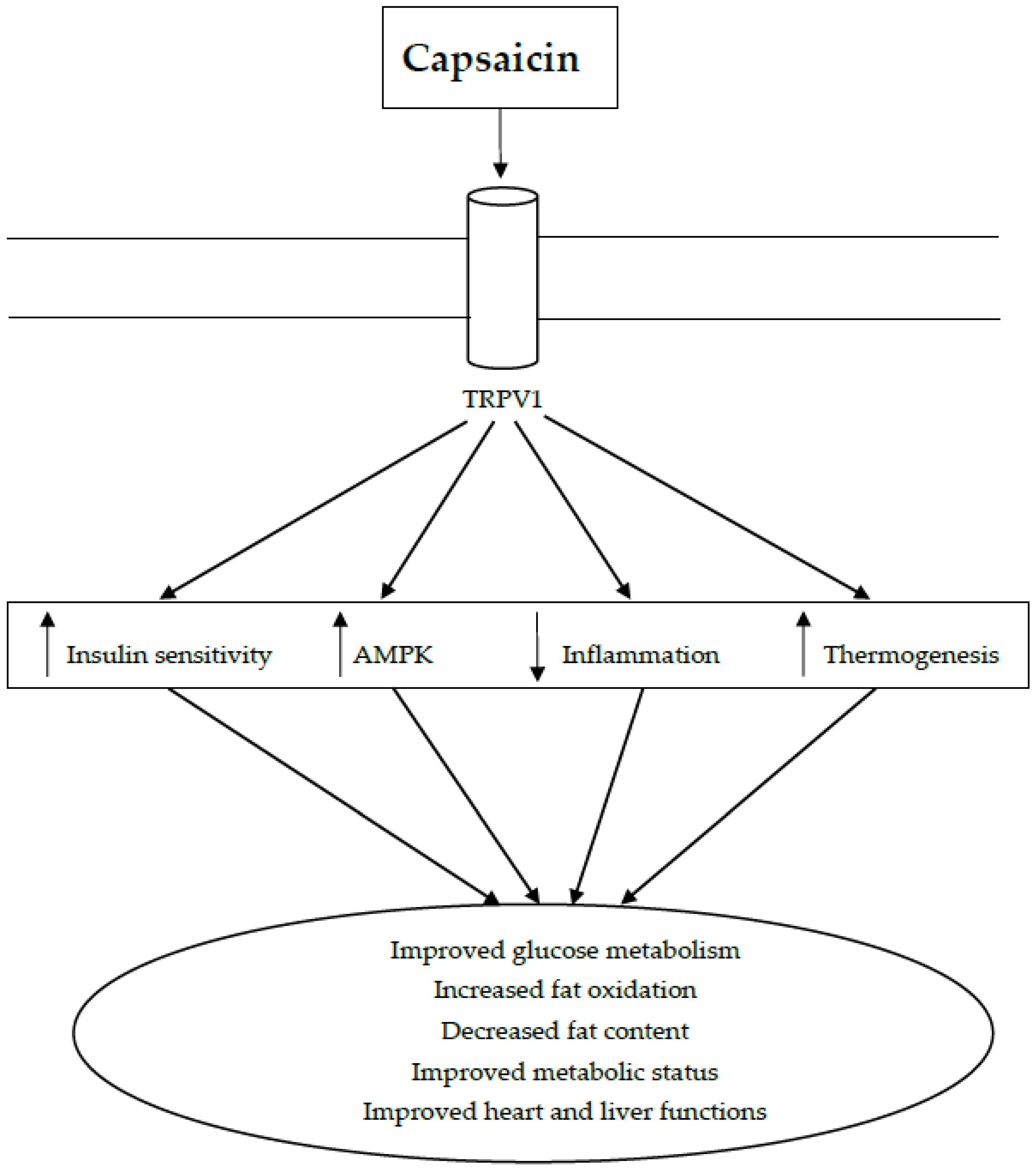
3. Capsaicin as a Treatment for Metabolic Syndrome
3.1. Capsaicin in Insulin Resistance and Glucose Metabolism
3.1.1. In Vitro Studies
3.1.2. Animal Studies
3.1.3. Human Studies
3.2. Capsaicin in Obesity and Dyslipidemia
3.2.1. In Vitro Studies
3.2.2. Animal Studies
3.2.3. Human Studies
3.3. Capsaicin in Vascular and Renal Function
3.4. Capsaicin in Non-Alcoholic Fatty Liver Disease
3.5. Anti-Inflammatory Actions of Capsaicin
3.6. Capsaicin in Oxidative Stress
3.7. Limitations in Clinical Use of Capsaicin
4. Conclusions and Future Directions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Thresh, J.C. Capsaicin, the active principle of capsicum fruits. Pharm. J. Trans. 1876, 7, 259–260. [Google Scholar]
- Nelson, E.K. The constitution of capsaicin, the pungent principle of capsicum. J. Am. Chem. Soc. 1919, 41, 1115–1121. [Google Scholar] [CrossRef]
- Späth, E.; Darling, S.F. Synthese des capsaicins. Eur. J. Inorg. Chem. 1930, 63, 737–743. [Google Scholar] [CrossRef]
- Rollyson, W.D.; Stover, C.A.; Brown, K.C.; Perry, H.E.; Stevenson, C.D.; McNees, C.A.; Ball, J.G.; Valentovic, M.A.; Dasgupta, P. Bioavailability of capsaicin and its implications for drug delivery. J. Control. Release 2014, 196, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Zahra, N.; Alim-un-Nisa, I.K.; Hina, S.; Javed, A.; Inam, S.M.; Malik, S.M.; Arshad, F. Estimation of capsaicin in different chilli varieties using different extraction techniques and HPLC method: A review. Pak. J. Food Sci. 2016, 26, 54–60. [Google Scholar]
- Pena-Alvarez, A.; Ramirez-Maya, E.; Alvarado-Suarez, L.A. Analysis of capsaicin and dihydrocapsaicin in peppers and pepper sauces by solid phase microextraction-gas chromatography-mass spectrometry. J. Chromatogr. A 2009, 1216, 2843–2847. [Google Scholar] [CrossRef] [PubMed]
- Patowary, P.; Pathak, M.P.; Zaman, K.; Raju, P.S.; Chattopadhyay, P. Research progress of capsaicin responses to various pharmacological challenges. Biomed. Pharmacother. 2017, 96, 1501–1512. [Google Scholar] [CrossRef] [PubMed]
- Groninger, H.; Schisler, R.E. Topical capsaicin for neuropathic pain #255. J. Palliat. Med. 2012, 15, 946–947. [Google Scholar] [CrossRef] [PubMed]
- Jones, V.M.; Moore, K.A.; Peterson, D.M. Capsaicin 8% topical patch (Qutenza)—A review of the evidence. J. Pain Palliat. Care Pharmacother. 2011, 25, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Bishnoi, M.; Khare, P.; Brown, L.; Panchal, S.K. Transient receptor potential (TRP) channels: A metabolic TR(i)P to obesity prevention and therapy. Obes. Rev. 2018. [Google Scholar] [CrossRef]
- Bevan, S.; Quallo, T.; Andersson, D.A. TRPV1. In Mammalian Transient Receptor Potential (TRP) Cation Channels: Volume I; Nilius, B., Flockerzi, V., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 222, pp. 207–245. [Google Scholar]
- Liao, M.; Cao, E.; Julius, D.; Cheng, Y. Structure of the TRPV1 ion channel determined by electron cryo-microscopy. Nature 2013, 504, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.K.; Guler, A.D.; Caterina, M.J. TRPV1 shows dynamic ionic selectivity during agonist stimulation. Nat. Neurosci. 2008, 11, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Hanson, S.M.; Newstead, S.; Swartz, K.J.; Sansom, M.S. Capsaicin interaction with TRPV1 channels in a lipid bilayer: Molecular dynamics simulation. Biophys. J. 2015, 108, 1425–1434. [Google Scholar] [CrossRef] [PubMed]
- Bae, C.; Anselmi, C.; Kalia, J.; Jara-Oseguera, A.; Schwieters, C.D.; Krepkiy, D.; Lee, C.W.; Kim, E.-H.; Kim, J.I.; Faraldo-Gómez, J.D. Structural insights into the mechanism of activation of the TRPV1 channel by a membrane-bound tarantula toxin. eLife 2016, 5, e11273. [Google Scholar] [CrossRef] [PubMed]
- Bang, S.; Yoo, S.; Oh, U.; Hwang, S.W. Endogenous lipid-derived ligands for sensory TRP ion channels and their pain modulation. Arch. Pharm. Res. 2010, 33, 1509–1520. [Google Scholar] [CrossRef] [PubMed]
- Dhaka, A.; Uzzell, V.; Dubin, A.E.; Mathur, J.; Petrus, M.; Bandell, M.; Patapoutian, A. TRPV1 is activated by both acidic and basic pH. J. Neurosci. 2009, 29, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Movahed, P.; Jönsson, B.A.; Birnir, B.; Wingstrand, J.A.; Jørgensen, T.D.; Ermund, A.; Sterner, O.; Zygmunt, P.M.; Högestätt, E.D. Endogenous unsaturated C18 N-acylethanolamines are vanilloid receptor (TRPV1) agonists. J. Biol. Chem. 2005, 280, 38496–38504. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, M.; Caterina, M.J.; Malmberg, A.B.; Rosen, T.A.; Gilbert, H.; Skinner, K.; Raumann, B.E.; Basbaum, A.I.; Julius, D. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 1998, 21, 531–543. [Google Scholar] [CrossRef]
- Zhang, L.L.; Yan Liu, D.; Ma, L.Q.; Luo, Z.D.; Cao, T.B.; Zhong, J.; Yan, Z.C.; Wang, L.J.; Zhao, Z.G.; Zhu, S.J.; et al. Activation of transient receptor potential vanilloid type-1 channel prevents adipogenesis and obesity. Circ. Res. 2007, 100, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Szolcsanyi, J.; Jancso-Gabor, A. Sensory effects of capsaicin congeners I. Relationship between chemical structure and pain-producing potency of pungent agents. Arzneimittelforschung 1975, 25, 1877–1881. [Google Scholar] [PubMed]
- Venkatachalam, K.; Montell, C. TRP channels. Annu. Rev. Biochem. 2007, 76, 387–417. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, E.S.; Fernandes, M.A.; Keeble, J.E. The functions of TRPA1 and TRPV1: Moving away from sensory nerves. Br. J. Pharmacol. 2012, 166, 510–521. [Google Scholar] [CrossRef] [PubMed]
- Rychkov, G.Y.; Barritt, G.J. Expression and function of TRP channels in liver cells. Adv. Exp. Med. Biol. 2011, 704, 667–686. [Google Scholar] [CrossRef] [PubMed]
- Zhong, B.; Wang, D.H. N-oleoyldopamine, a novel endogenous capsaicin-like lipid, protects the heart against ischemia-reperfusion injury via activation of TRPV1. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H728–H735. [Google Scholar] [CrossRef] [PubMed]
- Zhong, B.; Wang, D.H. TRPV1 gene knockout impairs preconditioning protection against myocardial injury in isolated perfused hearts in mice. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H1791–H1798. [Google Scholar] [CrossRef] [PubMed]
- Kassmann, M.; Harteneck, C.; Zhu, Z.; Nurnberg, B.; Tepel, M.; Gollasch, M. Transient receptor potential vanilloid 1 (TRPV1), TRPV4, and the kidney. Acta Physiol. 2013, 207, 546–564. [Google Scholar] [CrossRef] [PubMed]
- Frias, B.; Merighi, A. Capsaicin, nociception and pain. Molecules 2016, 21, 797. [Google Scholar] [CrossRef] [PubMed]
- Lotteau, S.; Ducreux, S.; Romestaing, C.; Legrand, C.; Van Coppenolle, F. Characterization of functional TRPV1 channels in the sarcoplasmic reticulum of mouse skeletal muscle. PLoS ONE 2013, 8, e58673. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Hazan, A.; Basu, A.; Zalcman, N.; Matzner, H.; Priel, A. Tyrosine residue in TRPV1 vanilloid binding pocket regulates deactivation kinetics. J. Biol. Chem. 2016, 291, 13855–13863. [Google Scholar] [CrossRef] [PubMed]
- Devesa, I.; Ferrándiz-Huertas, C.; Mathivanan, S.; Wolf, C.; Luján, R.; Changeux, J.-P.; Ferrer-Montiel, A. αCGRP is essential for algesic exocytotic mobilization of TRPV1 channels in peptidergic nociceptors. Proc. Natl. Acad. Sci. USA 2014, 111, 18345–18350. [Google Scholar] [CrossRef] [PubMed]
- Smutzer, G.; Devassy, R.K. Integrating TRPV1 receptor function with capsaicin psychophysics. Adv. Pharmacol. Sci. 2016, 2016, 1512457. [Google Scholar] [CrossRef] [PubMed]
- Smeets, A.J.; Westerterp-Plantenga, M.S. The acute effects of a lunch containing capsaicin on energy and substrate utilisation, hormones, and satiety. Eur. J. Nutr. 2009, 48, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Yan, Z.; Zhong, J.; Chen, J.; Ni, Y.; Li, L.; Ma, L.; Zhao, Z.; Liu, D.; Zhu, Z. Transient receptor potential vanilloid 1 activation enhances gut glucagon-like peptide-1 secretion and improves glucose homeostasis. Diabetes 2012, 61, 2155–2165. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, P.E.; El-kholy, W.; Riedel, M.J.; Salapatek, A.M.F.; Light, P.E.; Wheeler, M.B. The multiple actions of GLP-1 on the process of glucose-stimulated insulin secretion. Diabetes 2002, 51, S434–S442. [Google Scholar] [CrossRef] [PubMed]
- Parlevliet, E.T.; de Leeuw van Weenen, J.E.; Romijn, J.A.; Pijl, H. GLP-1 treatment reduces endogenous insulin resistance via activation of central GLP-1 receptors in mice fed a high-fat diet. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E318–E324. [Google Scholar] [CrossRef] [PubMed]
- Zsombok, A. Vanilloid receptors—Do they have a role in whole body metabolism? Evidence from TRPV1. J. Diabetes Complicat. 2013, 27, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Basith, S.; Cui, M.; Hong, S.; Choi, S. Harnessing the therapeutic potential of capsaicin and its analogues in pain and other diseases. Molecules 2016, 21, 966. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.K.; Vij, A.S.; Sharma, M. Mechanisms and clinical uses of capsaicin. Eur. J. Pharmacol. 2013, 720, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Fattori, V.; Hohmann, M.S.; Rossaneis, A.C.; Pinho-Ribeiro, F.A.; Verri, W.A. Capsaicin: Current understanding of its mechanisms and therapy of pain and other pre-clinical and clinical uses. Molecules 2016, 21, 844. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.L. A comprehensive definition for metabolic syndrome. Dis. Model. Mech. 2009, 2, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Ballestri, S.; Zona, S.; Targher, G.; Romagnoli, D.; Baldelli, E.; Nascimbeni, F.; Roverato, A.; Guaraldi, G.; Lonardo, A. Nonalcoholic fatty liver disease is associated with an almost twofold increased risk of incident type 2 diabetes and metabolic syndrome. Evidence from a systematic review and meta-analysis. J. Gastroenterol. Hepatol. 2016, 31, 936–944. [Google Scholar] [CrossRef] [PubMed]
- Nishida, K.; Otsu, K. Inflammation and metabolic cardiomyopathy. Cardiovasc. Res. 2017, 113, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Hernández, H.; Simental-Mendía, L.E.; Rodríguez-Ramírez, G.; Reyes-Romero, M.A. Obesity and inflammation: Epidemiology, risk factors, and markers of inflammation. Int. J. Endocrinol. 2013, 2013, 678159. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2017, 114, 1752–1761. [Google Scholar] [CrossRef] [PubMed]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative stress in obesity: A critical component in human diseases. Int. J. Mol. Sci. 2015, 16, 378–400. [Google Scholar] [CrossRef] [PubMed]
- Merone, L.; McDermott, R. Nutritional anti-inflammatories in the treatment and prevention of type 2 diabetes mellitus and the metabolic syndrome. Diabetes Res. Clin. Pract. 2017, 127, 238–253. [Google Scholar] [CrossRef] [PubMed]
- Gregorio, B.M.; De Souza, D.B.; de Morais Nascimento, F.A.; Pereira, L.M.; Fernandes-Santos, C. The potential role of antioxidants in metabolic syndrome. Curr. Pharm. Des. 2016, 22, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Samuel, V.T.; Shulman, G.I. Mechanisms for insulin resistance: Common threads and missing links. Cell 2012, 148, 852–871. [Google Scholar] [CrossRef] [PubMed]
- Ye, J. Mechanisms of insulin resistance in obesity. Front. Med. 2013, 7, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Roberts, C.K.; Hevener, A.L.; Barnard, R.J. Metabolic syndrome and insulin resistance: Underlying causes and modification by exercise training. Compr. Physiol. 2013, 3, 1–58. [Google Scholar] [CrossRef] [PubMed]
- Barnard, R.J.; Roberts, C.K.; Varon, S.M.; Berger, J.J. Diet-induced insulin resistance precedes other aspects of the metabolic syndrome. J. Appl. Physiol. 1998, 84, 1311–1315. [Google Scholar] [CrossRef] [PubMed]
- Parikh, R.M.; Mohan, V. Changing definitions of metabolic syndrome. Indian J. Endocrinol. Metab. 2012, 16, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Alberti, K.G.M.M.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.-C.; James, W.P.T.; Loria, C.M.; Smith, S.C. Harmonizing the metabolic syndrome: A joint interim statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International Atherosclerosis Society; and International Association for the Study of Obesity. Circulation 2009, 120, 1640–1645. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef]
- O’Neill, S.; O’Driscoll, L. Metabolic syndrome: A closer look at the growing epidemic and its associated pathologies. Obes. Rev. 2015, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pietilainen, K.H.; Kaprio, J.; Borg, P.; Plasqui, G.; Yki-Jarvinen, H.; Kujala, U.M.; Rose, R.J.; Westerterp, K.R.; Rissanen, A. Physical inactivity and obesity: A vicious circle. Obesity 2008, 16, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.A.; Kartashov, A.I.; Ebbeling, C.B.; Van Horn, L.; Slattery, M.L.; Jacobs, D.R., Jr.; Ludwig, D.S. Fast-food habits, weight gain, and insulin resistance (the CARDIA study): 15-year prospective analysis. Lancet 2005, 365, 36–42. [Google Scholar] [CrossRef]
- Boden, G. Obesity, insulin resistance and free fatty acids. Curr. Opin. Endocrinol. Diabetes Obes. 2011, 18, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Gungor, N.; Bacha, F.; Arslanian, S. Insulin resistance—Link to the components of the metabolic syndrome and biomarkers of endothelial dysfunction in youth. Diabetes Care 2007, 30, 2091–2097. [Google Scholar] [CrossRef] [PubMed]
- Juárez-López, C.; Klünder-Klünder, M.; Medina-Bravo, P.; Madrigal-Azcárate, A.; Mass-Díaz, E.; Flores-Huerta, S. Insulin resistance and its association with the components of the metabolic syndrome among obese children and adolescents. BMC Public Health 2010, 10, 318. [Google Scholar] [CrossRef] [PubMed]
- Berzigotti, A.; Abraldes, J.G. Impact of obesity and insulin-resistance on cirrhosis and portal hypertension. Gastroenterol. Hepatol. 2013, 36, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.; Pandya, V.; Whaley-Connell, A. Obesity and insulin resistance in resistant hypertension: Implications for the kidney. Adv. Chronic Kidney Dis. 2015, 22, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Czech, M.P. Insulin action and resistance in obesity and type 2 diabetes. Nat. Med. 2017, 23, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Carvajal, K.; Balderas-Villalobos, J.; Bello-Sanchez, M.D.; Phillips-Farfan, B.; Molina-Munoz, T.; Aldana-Quintero, H.; Gomez-Viquez, N.L. Ca2+ mishandling and cardiac dysfunction in obesity and insulin resistance: Role of oxidative stress. Cell Calcium 2014, 56, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Inoue, R.; Jensen, L.J.; Shi, J.; Morita, H.; Nishida, M.; Honda, A.; Ito, Y. Transient receptor potential channels in cardiovascular function and disease. Circ. Res. 2006, 99, 119–131. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F.; DiNicolantonio, J.J.; O’Keefe, J.H. Capsaicin may have important potential for promoting vascular and metabolic health. Open Heart 2015, 2, e000262. [Google Scholar] [CrossRef] [PubMed]
- Aronoff, S.L.; Berkowitz, K.; Shreiner, B.; Want, L. Glucose metabolism and regulation: Beyond insulin and glucagon. Diabetes Spectrum 2004, 17, 183–190. [Google Scholar] [CrossRef]
- Kim, S.H.; Hwang, J.T.; Park, H.S.; Kwon, D.Y.; Kim, M.S. Capsaicin stimulates glucose uptake in C2C12 muscle cells via the reactive oxygen species (ROS)/AMPK/p38 MAPK pathway. Biochem. Biophys. Res. Commun. 2013, 439, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Monsereenusorn, Y.; Glinsukon, T. Inhibitory effect of capsaicin on intestinal glucose absorption in vitro. Food Cosmet. Toxicol. 1978, 16, 469–473. [Google Scholar] [CrossRef]
- Han, J.; Isoda, H. Capsaicin induced the upregulation of transcriptional and translational expression of glycolytic enzymes related to energy metabolism in human intestinal epithelial cells. J. Agric. Food Chem. 2009, 57, 11148–11153. [Google Scholar] [CrossRef] [PubMed]
- Tolan, I.; Ragoobirsingh, D.; Morrison, E.Y. Isolation and purification of the hypoglycaemic principle present in Capsicum frutescens. Phytother. Res. 2004, 18, 95–96. [Google Scholar] [CrossRef] [PubMed]
- Tolan, I.; Ragoobirsingh, D.; Morrison, E.Y. The effect of capsaicin on blood glucose, plasma insulin levels and insulin binding in dog models. Phytother. Res. 2001, 15, 391–394. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Tsuyoshi, G.; Han, I.S.; Kawada, T.; Kim, Y.M.; Yu, R. Dietary capsaicin reduces obesity-induced insulin resistance and hepatic steatosis in obese mice fed a high-fat diet. Obesity 2010, 18, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.Y.; Kim, Y.S.; Ryu, S.Y.; Cha, M.R.; Yon, G.H.; Yang, H.J.; Kim, M.J.; Kang, S.; Park, S. Capsiate improves glucose metabolism by improving insulin sensitivity better than capsaicin in diabetic rats. J. Nutr. Biochem. 2013, 24, 1078–1085. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Tsuyoshi, G.; Le Ngoc, H.; Kim, H.M.; Tu, T.H.; Noh, H.J.; Kim, C.S.; Choe, S.Y.; Kawada, T.; Yoo, H.; et al. Dietary capsaicin attenuates metabolic dysregulation in genetically obese diabetic mice. J. Med. Food 2011, 14, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Song, J.X.; Ren, H.; Gao, Y.F.; Lee, C.Y.; Li, S.F.; Zhang, F.; Li, L.; Chen, H. Dietary capsaicin improves glucose homeostasis and alters the gut microbiota in obese diabetic ob/ob mice. Front. Physiol. 2017, 8, 602. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.J.; Kwon, D.Y.; Kim, M.J.; Kang, S.; Moon, N.R.; Daily, J.W.; Park, S. Red peppers with moderate and severe pungency prevent the memory deficit and hepatic insulin resistance in diabetic rats with Alzheimer’s disease. Nutr. Metab. 2015, 12, 9. [Google Scholar] [CrossRef] [PubMed]
- Şekeroğlu, V.; Aydın, B.; Atlı Şekeroğlu, Z.; Özdener Kömpe, Y. Hepatoprotective effects of capsaicin and alpha-tocopherol on mitochondrial function in mice fed a high-fat diet. Biomed. Pharmacother. 2018, 98, 821–825. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chen, J.; Ni, Y.; Feng, X.; Zhao, Z.; Wang, P.; Sun, J.; Yu, H.; Yan, Z.; Liu, D.; et al. TRPV1 activation prevents nonalcoholic fatty liver through UCP2 upregulation in mice. Pflugers Arch. 2012, 463, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.R.; Shin, M.K.; Yoon, D.J.; Kim, A.R.; Yu, R.; Park, N.H.; Han, I.S. Topical application of capsaicin reduces visceral adipose fat by affecting adipokine levels in high-fat diet-induced obese mice. Obesity 2013, 21, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.J.; Qin, Y.; Wang, L.; Zeng, Y.; Chang, H.; Wang, J.; Wang, B.; Wan, J.; Chen, S.H.; Zhang, Q.Y.; et al. Capsaicin-containing chili improved postprandial hyperglycemia, hyperinsulinemia, and fasting lipid disorders in women with gestational diabetes mellitus and lowered the incidence of large-for-gestational-age newborns. Clin. Nutr. 2016, 35, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, K.D.; Robertson, I.K.; Geraghty, D.P.; Ball, M.J. Effects of chili consumption on postprandial glucose, insulin, and energy metabolism. Am. J. Clin. Nutr. 2006, 84, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ma, X.; Zhang, L.; Sun, H.; Liu, X. Capsaicin reduces blood glucose by increasing insulin levels and glycogen content better than capsiate in streptozotocin-induced diabetic rats. J. Agric. Food Chem. 2017, 65, 2323–2330. [Google Scholar] [CrossRef] [PubMed]
- Medina-Contreras, J.M.L.; Colado-Velázquez Iii, J.; Gómez-Viquez, N.L.; Mailloux-Salinas, P.; Pérez-Torres, I.; Aranda-Fraustro, A.; Carvajal, K.; Bravo, G. Effects of topical capsaicin combined with moderate exercise on insulin resistance, body weight and oxidative stress in hypoestrogenic obese rats. Int. J. Obes. 2017, 41, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Gram, D.X.; Hansen, A.J.; Wilken, M.; Elm, T.; Svendsen, O.; Carr, R.D.; Ahrén, B.; Brand, C.L. Plasma calcitonin gene-related peptide is increased prior to obesity, and sensory nerve desensitization by capsaicin improves oral glucose tolerance in obese Zucker rats. Eur. J. Endocrinol. 2005, 153, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Gram, D.X.; Hansen, A.J.; Deacon, C.F.; Brand, C.L.; Ribel, U.; Wilken, M.; Carr, R.D.; Svendsen, O.; Ahren, B. Sensory nerve desensitization by resiniferatoxin improves glucose tolerance and increases insulin secretion in Zucker Diabetic Fatty rats and is associated with reduced plasma activity of dipeptidyl peptidase IV. Eur. J. Pharmacol. 2005, 509, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Gram, D.X.; Ahren, B.; Nagy, I.; Olsen, U.B.; Brand, C.L.; Sundler, F.; Tabanera, R.; Svendsen, O.; Carr, R.D.; Santha, P.; et al. Capsaicin-sensitive sensory fibers in the islets of Langerhans contribute to defective insulin secretion in Zucker diabetic rat, an animal model for some aspects of human type 2 diabetes. Eur. J. Neurosci. 2007, 25, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Oh, T.W.; Ohta, F. Capsaicin increases endurance capacity and spares tissue glycogen through lipolytic function in swimming rats. J. Nutr. Sci. Vitaminol. 2003, 49, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Yoshioka, M.; Suzuki, M. Capsaicin in diet does not affect glycogen contents in the liver and skeletal muscle of rats before and after exercise. J. Nutr. Sci. Vitaminol. 1996, 42, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, F.; Inoue, N.; Masamoto, Y.; Matsumura, S.; Kimura, W.; Kadowaki, M.; Higashi, T.; Tominaga, M.; Inoue, K.; Fushiki, T. Non-pungent capsaicin analogs (capsinoids) increase metabolic rate and enhance thermogenesis via gastrointestinal TRPV1 in mice. Biosci. Biotechnol. Biochem. 2009, 73, 2690–2697. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.-J.; Huang, W.-C.; Chiu, C.-C.; Liu, Y.-L.; Chiu, W.-C.; Chiu, C.-H.; Chiu, Y.-S.; Huang, C.-C. Capsaicin supplementation reduces physical fatigue and improves exercise performance in mice. Nutrients 2016, 8, 648. [Google Scholar] [CrossRef] [PubMed]
- Imaizumi, K.; Sato, S.; Kumazawa, M.; Arai, N.; Aritoshi, S.; Akimoto, S.; Sakakibara, Y.; Kawashima, Y.; Tachiyashiki, K. Capsaicinoids-induced changes of plasma glucose, free fatty acid and glycerol concentrations in rats. J. Toxicol. Sci. 2011, 36, 109–116. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Van de Wall, E.H.E.M.; Gram, D.X.; Strubbe, J.H.; Scheurink, A.J.W.; Koolhaas, J.M. Ablation of capsaicin-sensitive afferent nerves affects insulin response during an intravenous glucose tolerance test. Life Sci. 2005, 77, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Trudeau, F. Plasma glucose and capsaicin-sensitive nerves in the fed rat during exercise. Horm. Metab. Res. 1993, 25, 548–549. [Google Scholar] [CrossRef] [PubMed]
- Guillot, E.; Coste, A.; Angel, I. Involvement of capsaicin-sensitive nerves in the regulation of glucose tolerance in diabetic rats. Life Sci. 1996, 59, 969–977. [Google Scholar] [CrossRef]
- Morgan, A.E.; Lang, C.H. Involvement of capsaicin-sensitive nerves in regulating the hormone and glucose metabolic response to endotoxin. Am. J. Physiol. 1997, 273, E328–E335. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.F.; Jhamandas, K.H.; Livett, B.G. Capsaicin-sensitive nerves are required for glucostasis but not for catecholamine output during hypoglycemia in rats. Am. J. Physiol. 1990, 258, E212–E219. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, S.J.; Leighton, B.; DeFronzo, R.A. Neonatal de-afferentation of capsaicin-sensitive sensory nerves increases in vivo insulin sensitivity in conscious adult rats. Diabetologia 1998, 41, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, S.; Ahren, B. Capsaicin-induced sensory denervation increases glucose elimination in rodents. Diabetologia 1999, 42, 260–261. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lee, E.; Jung, D.Y.; Kim, J.H.; Patel, P.R.; Hu, X.; Lee, Y.; Azuma, Y.; Wang, H.-F.; Tsitsilianos, N.; Shafiq, U.; et al. Transient receptor potential vanilloid type-1 channel regulates diet-induced obesity, insulin resistance, and leptin resistance. FASEB J. 2015, 29, 3182–3192. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.; Yoshioka, M.; Kikuzato, S.; Kiyonaga, A.; Tanaka, H.; Shindo, M.; Suzuki, M. Dietary red pepper ingestion increases carbohydrate oxidation at rest and during exercise in runners. Med. Sci. Sports Exerc. 1997, 29, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Domotor, A.; Szolcsanyi, J.; Mozsik, G. Capsaicin and glucose absorption and utilization in healthy human subjects. Eur. J. Pharmacol. 2006, 534, 280–283. [Google Scholar] [CrossRef] [PubMed]
- Chaiyasit, K.; Khovidhunkit, W.; Wittayalertpanya, S. Pharmacokinetic and the effect of capsaicin in Capsicum frutescens on decreasing plasma glucose level. J. Med. Assoc. Thail. 2009, 92, 108–113. [Google Scholar]
- Rondanelli, M.; Opizzi, A.; Perna, S.; Faliva, M.; Solerte, S.B.; Fioravanti, M.; Klersy, C.; Edda, C.; Maddalena, P.; Luciano, S.; et al. Improvement in insulin resistance and favourable changes in plasma inflammatory adipokines after weight loss associated with two months’ consumption of a combination of bioactive food ingredients in overweight subjects. Endocrine 2013, 44, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Kroff, J.; Hume, D.J.; Pienaar, P.; Tucker, R.; Lambert, E.V.; Rae, D.E. The metabolic effects of a commercially available chicken peri-peri (African bird’s eye chilli) meal in overweight individuals. Br. J. Nutr. 2017, 117, 635–644. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Urbina, S.L.; Roberts, M.D.; Kephart, W.C.; Villa, K.B.; Santos, E.N.; Olivencia, A.M.; Bennett, H.M.; Lara, M.D.; Foster, C.A.; Purpura, M.; et al. Effects of twelve weeks of capsaicinoid supplementation on body composition, appetite and self-reported caloric intake in overweight individuals. Appetite 2017, 113, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Ludy, M.-J.; Moore, G.E.; Mattes, R.D. The effects of capsaicin and capsiate on energy balance: Critical review and meta-analyses of studies in humans. Chem. Senses 2012, 37, 103–121. [Google Scholar] [CrossRef] [PubMed]
- Saito, M. Capsaicin and related food ingredients reducing body fat through the activation of TRP and brown fat thermogenesis. Adv. Food Nutr. Res. 2015, 76, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Baskaran, P.; Krishnan, V.; Ren, J.; Thyagarajan, B. Capsaicin induces browning of white adipose tissue and counters obesity by activating TRPV1 channel-dependent mechanisms. Br. J. Pharmacol. 2016, 173, 2369–2389. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.L.; Yen, G.C. Effects of capsaicin on induction of apoptosis and inhibition of adipogenesis in 3T3-L1 cells. J. Agric. Food Chem. 2007, 55, 1730–1736. [Google Scholar] [CrossRef] [PubMed]
- Baboota, R.K.; Singh, D.P.; Sarma, S.M.; Kaur, J.; Sandhir, R.; Boparai, R.K.; Kondepudi, K.K.; Bishnoi, M. Capsaicin induces “brite” phenotype in differentiating 3T3-L1 preadipocytes. PLoS ONE 2014, 9, e103093. [Google Scholar] [CrossRef] [PubMed]
- Rohm, B.; Riedel, A.; Ley, J.P.; Widder, S.; Krammer, G.E.; Somoza, V. Capsaicin, nonivamide and trans-pellitorine decrease free fatty acid uptake without TRPV1 activation and increase acetyl-coenzyme A synthetase activity in Caco-2 cells. Food Funct. 2015, 6, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Kawada, T.; Hagihara, K.; Iwai, K. Effects of capsaicin on lipid metabolism in rats fed a high fat diet. J. Nutr. 1986, 116, 1272–1278. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Gao, B.; Tao, Y.; Guo, J.; Su, Z.Q. Antiobese effects of capsaicin-chitosan microsphere (CCMS) in obese rats induced by high fat diet. J. Agric. Food Chem. 2014, 62, 1866–1874. [Google Scholar] [CrossRef] [PubMed]
- Melnyk, A.; Himms-Hagen, J. Resistance to aging-associated obesity in capsaicin-desensitized rats one year after treatment. Obes. Res. 1995, 3, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Hirotani, Y.; Fukamachi, J.; Ueyama, R.; Urashima, Y.; Ikeda, K. Effects of capsaicin coadministered with eicosapentaenoic acid on obesity-related dysregulation in high-fat-fed mice. Biol. Pharm. Bull. 2017, 40, 1581–1585. [Google Scholar] [CrossRef] [PubMed]
- Mun, J.M.; Ok, H.M.; Kwon, O. Corn gluten hydrolysate and capsaicin have complimentary actions on body weight reduction and lipid-related genes in diet-induced obese rats. Nutr. Res. 2014, 34, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Diepvens, K.; Westerterp, K.R.; Westerterp-Plantenga, M.S. Obesity and thermogenesis related to the consumption of caffeine, ephedrine, capsaicin, and green tea. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R77–R85. [Google Scholar] [CrossRef] [PubMed]
- Kim Dong, H.; Joo Jeong, I.; Choi, J.W.; Yun Jong, W. Differential expression of skeletal muscle proteins in high-fat diet-fed rats in response to capsaicin feeding. Proteomics 2010, 10, 2870–2881. [Google Scholar] [CrossRef]
- Hur, K.Y.; Lee, M.-S. Gut microbiota and metabolic disorders. Diabetes Metab. J. 2015, 39, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Kweon, M.N. The gut microbiota: A key regulator of metabolic diseases. BMB Rep. 2016, 49, 536–541. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Shi, B. Gut microbiota as a potential target of metabolic syndrome: The role of probiotics and prebiotics. Cell Biosci. 2017, 7, 54. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Wang, B.; Kaliannan, K.; Wang, X.; Lang, H.; Hui, S.; Huang, L.; Zhang, Y.; Zhou, M.; Chen, M.; et al. Gut microbiota mediates the protective effects of dietary capsaicin against chronic low-grade inflammation and associated obesity induced by high-fat diet. mBio 2017, 8, e00470-17. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Xiong, S.; Zhu, Z. Dietary capsaicin protects cardiometabolic organs from dysfunction. Nutrients 2016, 8, 174. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Shen, M.; Zhao, X.; Zhu, H.; Yang, Y.; Lu, S.; Tan, Y.; Li, G.; Li, M.; Wang, J.; et al. Anti-obesity effect of capsaicin in mice fed with high-fat diet is associated with an increase in population of the gut bacterium Akkermansia muciniphila. Front. Microbiol. 2017, 8, 272. [Google Scholar] [CrossRef] [PubMed]
- Taghizadeh, M.; Farzin, N.; Taheri, S.; Mahlouji, M.; Akbari, H.; Karamali, F.; Asemi, Z. The effect of dietary supplements containing green tea, capsaicin and ginger extracts on weight loss and metabolic profiles in overweight women: A randomized double-blind placebo-controlled clinical trial. Ann. Nutr. Metab. 2017, 70, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, N.A.; Spillane, M.; La Bounty, P.; Grandjean, P.W.; Leutholtz, B.; Willoughby, D.S. Capsaicin and evodiamine ingestion does not augment energy expenditure and fat oxidation at rest or after moderately-intense exercise. Nutr. Res. 2013, 33, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Zhang, Y.; Zhu, X.; Liu, K.; Wang, X.; Chen, M.; Wang, J.; Chen, H.; Hui, S.; Huang, L.; et al. Healthy subjects differentially respond to dietary capsaicin correlating with specific gut enterotypes. J. Clin. Endocrinol. Metab. 2016, 101, 4681–4689. [Google Scholar] [CrossRef] [PubMed]
- Janssens, P.L.; Hursel, R.; Westerterp-Plantenga, M.S. Capsaicin increases sensation of fullness in energy balance, and decreases desire to eat after dinner in negative energy balance. Appetite 2014, 77, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Westerterp-Plantenga, M.S.; Smeets, A.; Lejeune, M.P. Sensory and gastrointestinal satiety effects of capsaicin on food intake. Int. J. Obes. 2005, 29, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Van Avesaat, M.; Troost, F.J.; Westerterp-Plantenga, M.S.; Helyes, Z.; Le Roux, C.W.; Dekker, J.; Masclee, A.A.; Keszthelyi, D. Capsaicin-induced satiety is associated with gastrointestinal distress but not with the release of satiety hormones. Am. J. Clin. Nutr. 2016, 103, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Achike, F.I.; To, N.H.; Wang, H.; Kwan, C.Y. Obesity, metabolic syndrome, adipocytes and vascular function: A holistic viewpoint. Clin. Exp. Pharmacol. Physiol. 2011, 38, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tesauro, M.; Cardillo, C. Obesity, blood vessels and metabolic syndrome. Acta Physiol. 2011, 203, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Wassink, A.M.; Olijhoek, J.K.; Visseren, F.L. The metabolic syndrome: Metabolic changes with vascular consequences. Eur. J. Clin. Investig. 2007, 37, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Hwang, M.-H.; Lee, S. Insulin resistance: Vascular function and exercise. Integr. Med. Res. 2016, 5, 198–203. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gluba, A.; Mikhailidis, D.P.; Lip, G.Y.; Hannam, S.; Rysz, J.; Banach, M. Metabolic syndrome and renal disease. Int. J. Cardiol. 2013, 164, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, D.H. Function and regulation of the vanilloid receptor in rats fed a high salt diet. J. Hypertens. 2003, 21, 1525–1530. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Luo, Z.; Ma, S.; Wong, W.T.; Ma, L.; Zhong, J.; He, H.; Zhao, Z.; Cao, T.; Yan, Z. Activation of TRPV1 by dietary capsaicin improves endothelium-dependent vasorelaxation and prevents hypertension. Cell Metab. 2010, 12, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wang, P.; Zhao, Z.; Cao, T.; He, H.; Luo, Z.; Zhong, J.; Gao, F.; Zhu, Z.; Li, L.; et al. Activation of transient receptor potential vanilloid 1 by dietary capsaicin delays the onset of stroke in stroke-prone Spontaneously Hypertensive Rats. Stroke 2011, 42, 3245–3251. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, D.H. Increased GFR and renal excretory function by activation of TRPV1 in the isolated perfused kidney. Pharmacol. Res. 2008, 57, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhong, J.; Zhao, Z.; Luo, Z.; Ma, S.; Sun, J.; He, H.; Zhu, T.; Liu, D.; Zhu, Z. Activation of TRPV1 reduces vascular lipid accumulation and attenuates atherosclerosis. Cardiovasc. Res. 2011, 92, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Yin, Y.; Liu, Y.; Pi, Y.; Guo, L.; Cao, X.; Gao, C.; Zhang, L.; Li, J. TRPV1 activation impedes foam cell formation by inducing autophagy in oxLDL-treated vascular smooth muscle cells. Cell Death Dis. 2014, 5, e1182. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Pu, Y.; Wang, P.; Chen, S.; Zhao, Y.; Liu, C.; Shang, Q.; Zhu, Z.; Liu, D. TRPV1-mediated UCP2 upregulation ameliorates hyperglycemia-induced endothelial dysfunction. Cardiovasc. Diabetol. 2013, 12, 69. [Google Scholar] [CrossRef] [PubMed]
- Fabbrini, E.; Sullivan, S.; Klein, S. Obesity and nonalcoholic fatty liver disease: Biochemical, metabolic and clinical implications. Hepatology 2010, 51, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Katsiki, N.; Mikhailidis, D.P.; Mantzoros, C.S. Non-alcoholic fatty liver disease and dyslipidemia: An update. Metabolism 2016, 65, 1109–1123. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, L.; Wang, F.; Chen, J.; Zhao, Y.; Wang, P.; Nilius, B.; Liu, D.; Zhu, Z. Dietary capsaicin prevents nonalcoholic fatty liver disease through transient receptor potential vanilloid 1-mediated peroxisome proliferator-activated receptor delta activation. Pflugers Arch. 2013, 465, 1303–1316. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Luo, H.; Jiang, Y.; Chen, P. Dietary capsaicin and antibiotics act synergistically to reduce non-alcoholic fatty liver disease induced by high fat diet in mice. Oncotarget 2017, 8, 38161–38175. [Google Scholar] [CrossRef] [PubMed]
- Bitencourt, S.; Stradiot, L.; Verhulst, S.; Thoen, L.; Mannaerts, I.; van Grunsven, L.A. Inhibitory effect of dietary capsaicin on liver fibrosis in mice. Mol. Nutr. Food Res. 2015, 59, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation, metaflammation and immunometabolic disorders. Nature 2017, 542, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; Saltiel, A.R. Inflammatory links between obesity and metabolic disease. J. Clin. Investig. 2011, 121, 2111–2117. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.; Azevedo, I. Chronic inflammation in obesity and the metabolic syndrome. Mediat. Inflamm. 2010, 2010, 289645. [Google Scholar] [CrossRef] [PubMed]
- Mraz, M.; Haluzik, M. The role of adipose tissue immune cells in obesity and low-grade inflammation. J. Endocrinol. 2014, 222, R113–R127. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Vicario, C.; Alcaraz-Quiles, J.; Garcia-Alonso, V.; Rius, B.; Hwang, S.H.; Titos, E.; Lopategi, A.; Hammock, B.D.; Arroyo, V.; Claria, J. Inhibition of soluble epoxide hydrolase modulates inflammation and autophagy in obese adipose tissue and liver: Role for omega-3 epoxides. Proc. Natl. Acad. Sci. USA 2015, 112, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Iyer, A.; Lim, J.; Poudyal, H.; Reid, R.C.; Suen, J.Y.; Webster, J.; Prins, J.B.; Whitehead, J.P.; Fairlie, D.P.; Brown, L. An inhibitor of phospholipase A2 group IIA modulates adipocyte signaling and protects against diet-induced metabolic syndrome in rats. Diabetes 2012, 61, 2320–2329. [Google Scholar] [CrossRef] [PubMed]
- Hui, D.Y.; Cope, M.J.; Labonte, E.D.; Chang, H.T.; Shao, J.; Goka, E.; Abousalham, A.; Charmot, D.; Buysse, J. The phospholipase A2 inhibitor methyl indoxam suppresses diet-induced obesity and glucose intolerance in mice. Br. J. Pharmacol. 2009, 157, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
- Lundh, M.; Galbo, T.; Poulsen, S.S.; Mandrup-Poulsen, T. Histone deacetylase 3 inhibition improves glycaemia and insulin secretion in obese diabetic rats. Diabetes Obes. Metab. 2015, 17, 703–707. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Taliyan, R. Histone deacetylase inhibitors: Future therapeutics for insulin resistance and type 2 diabetes. Pharmacol. Res. 2016, 113, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, A.; Fiorino, E.; Longo, R.; Barilla, S.; Mitro, N.; Cermenati, G.; Giudici, M.; Caruso, D.; Mai, A.; Guerrini, U.; et al. Attenuation of diet-induced obesity and induction of white fat browning with a chemical inhibitor of histone deacetylases. Int. J. Obes. 2017, 41, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Hong, F.; Wang, J.; Wang, J.; Zhao, X.; Wang, S.; Xue, T.; Xu, J.; Zheng, X.; Zhai, Y. DBZ is a putative PPARg agonist that prevents high fat diet-induced obesity, insulin resistance and gut dysbiosis. Biochim. Biophys. Acta 2017, 1861, 2690–2701. [Google Scholar] [CrossRef] [PubMed]
- Roberts, C.K.; Sindhu, K.K. Oxidative stress and metabolic syndrome. Life Sci. 2009, 84, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Mahjoub, S.; Masrour-Roudsari, J. Role of oxidative stress in pathogenesis of metabolic syndrome. Casp. J. Intern. Med. 2012, 3, 386–396. [Google Scholar]
- Lee, C.Y.; Kim, M.; Yoon, S.W.; Lee, C.H. Short-term control of capsaicin on blood and oxidative stress of rats in vivo. Phytother. Res. 2003, 17, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Luqman, S.; Rizvi Syed, I. Protection of lipid peroxidation and carbonyl formation in proteins by capsaicin in human erythrocytes subjected to oxidative stress. Phytother. Res. 2006, 20, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, M.; Zhang, J.; Peng, W.; Firempong, C.K.; Deng, W.; Wang, Q.; Wang, S.; Shi, F.; Yu, J.; et al. Improved oral bioavailability of capsaicin via liposomal nanoformulation: Preparation, in vitro drug release and pharmacokinetics in rats. Arch. Pharm. Res. 2015, 38, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Jiang, X.Y.; Zhu, Y.; Omari-Siaw, E.; Deng, W.W.; Yu, J.N.; Xu, X.M.; Zhang, W.M. Oral delivery of capsaicin using MPEG-PCL nanoparticles. Acta Pharmacol. Sin. 2015, 36, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Sasahara, I.; Furuhata, Y.; Iwasaki, Y.; Inoue, N.; Sato, H.; Watanabe, T.; Takahashi, M. Assessment of the biological similarity of three capsaicin analogs (Capsinoids) found in non-pungent chili pepper (CH-19 Sweet) fruits. Biosci. Biotechnol. Biochem. 2010, 74, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Bode, A.M.; Dong, Z. The two faces of capsaicin. Cancer Res. 2011, 71, 2809–2814. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Laviada, I.; Rodríguez-Henche, N. The potential antitumor effects of capsaicin. In Capsaicin as a Therapeutic Molecule; Abdel-Salam, O.M.E., Ed.; Springer: Basel, Switzerland, 2014; pp. 181–208. [Google Scholar]
- Boulinguiez, A.; Staels, B.; Duez, H.; Lancel, S. Mitochondria and endoplasmic reticulum: Targets for a better insulin sensitivity in skeletal muscle? Biochim. Biophys. Acta 2017, 1862, 901–916. [Google Scholar] [CrossRef] [PubMed]
- Yashiro, K.; Tonson, A.; Pecchi, E.; Vilmen, C.; Le Fur, Y.; Bernard, M.; Bendahan, D.; Giannesini, B. Capsiate supplementation reduces oxidative cost of contraction in exercising mouse skeletal muscle in vivo. PLoS ONE 2015, 10, e0128016. [Google Scholar] [CrossRef] [PubMed]
- Faraut, B.; Giannesini, B.; Matarazzo, V.; Le Fur, Y.; Rougon, G.; Cozzone, P.J.; Bendahan, D. Capsiate administration results in an uncoupling protein-3 downregulation, an enhanced muscle oxidative capacity and a decreased abdominal fat content in vivo. Int. J. Obes. 2009, 33, 1348–1355. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.; Poudyal, H.; Panchal, S.K. Functional foods as potential therapeutic options for metabolic syndrome. Obes. Rev. 2015, 16, 914–941. [Google Scholar] [CrossRef] [PubMed]
- Baboota, R.K.; Khare, P.; Mangal, P.; Singh, D.P.; Bhutani, K.K.; Kondepudi, K.K.; Kaur, J.; Bishnoi, M. Dihydrocapsiate supplementation prevented high-fat diet–induced adiposity, hepatic steatosis, glucose intolerance, and gut morphological alterations in mice. Nutr. Res. 2018, 51, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Irii, H.; Tahara, Y.; Ishii, H.; Hirao, A.; Udagawa, H.; Hiramoto, M.; Yasuda, K.; Takanishi, A.; Shibata, S.; et al. Synthesis of a new (6)-gingerol analogue and its protective effect with respect to the development of metabolic syndrome in mice fed a high-fat diet. J. Med. Chem. 2011, 54, 6295–6304. [Google Scholar] [CrossRef] [PubMed]
- Diwan, V.; Poudyal, H.; Brown, L. Piperine attenuates cardiovascular, liver and metabolic changes in high carbohydrate, high fat-fed rats. Cell Biochem. Biophys. 2013, 67, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Hochkogler, C.M.; Lieder, B.; Rust, P.; Berry, D.; Meier, S.M.; Pignitter, M.; Riva, A.; Leitinger, A.; Bruk, A.; Wagner, S.; et al. A 12-week intervention with nonivamide, a TRPV1 agonist, prevents a dietary-induced body fat gain and increases peripheral serotonin in moderately overweight subjects. Mol. Nutr. Food Res. 2017, 61, 1600731. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Animal Model/Human | Capsaicin Dose (Duration) | Effects on Glucose Metabolism | Mechanism(s) to Improve Glucose & Insulin Responses |
|---|---|---|---|
| db/db mice and TRPV1−/− mice [34] | 0.01% of diet (24 weeks) | ↑ insulin sensitivity ↓ basal blood glucose | ↑ TRPV1 expression ↑ intestinal GLP1 secretion |
| C57BL/6 mice [74] | 0.015% of diet (10 weeks) | ↓ basal blood glucose ↓ glucose intolerance ↓ basal blood insulin | ↑ TRPV1 activity ↓ PPARγ activity ↓ NF-κB activity ↓ inflammatory cytokines ↓ IRS-1 ↑ GLUT4 expression ↑ adiponectin ↓ leptin |
| Sprague Dawley rats [75] | 0.025% of diet (8 weeks) | ↓ basal blood glucose ↓ basal blood insulin ↓ glucose intolerance ↑ insulin sensitivity ↓ insulin intolerance | ↓ leptin ↑ pancreatic β-cell mass ↓ pancreatic islet cell apoptosis ↑ ratio of β:α pancreatic cells ↑ pAkt/PEPCK & pAMPK signaling post-TRPV1 activation |
| KKAy mice [76] | 0.015% of diet (3 weeks) | ↓ basal blood insulin ↓ basal blood glucose | ↓ inflammatory cytokines ↑ adiponectin ↑ AdipoR2 expression |
| ob/ob mice [77] | 0.01% or 0.02% of diet (6 weeks) | ↓ basal blood insulin ↓ basal blood glucose ↓ glucose intolerance ↑ insulin sensitivity ↓ insulin intolerance | ↓ ghrelin ↓ inflammatory cytokines ↑ GLP-1 ↑ butyrate |
| Sprague Dawley rats [78] | Chillies equal to 1% of diet (7 weeks) | ↓ basal blood insulin ↓ HOMA-IR ↑ insulin sensitivity ↓ insulin intolerance | ↑ pancreatic β-cell mass ↓ pancreatic islet cell apoptosis ↓ β-amyloid accumulation |
| Swiss albino mice [79] | 5 mg/kg/day (8 weeks) | ↓ basal blood glucose ↓ HOMA-IR | ↑ glucose 6-phosphate dehydrogenase ↑ glutathione-S-transferases |
| C57BL/6 mice and TRPV1−/− mice [80] | 0.01% of diet (24 weeks) | ↓ basal blood glucose | ↑ hepatic β-oxidation ↑ TRPV1 expression and activity ↑ UCP2 expression and activity |
| C57BL/6 mice [81] | 100 mg of 0.075% capsaicin cream/day (7 weeks) | ↓ basal blood glucose ↑ insulin sensitivity | ↑ adiponectin ↑ PPARα, PPARγ, visfatin, adipsin ↓ inflammatory cytokines |
| Women with gestational diabetes mellitus [82] | 5 mg/day (4 weeks) | ↓ 2 h postprandial blood glucose ↓ 2 h postprandial blood insulin ↓ 2 h postprandial HOMA-IR | ↓ calcitonin gene-related peptide |
| Humans [83] | 30 mg/day (4 weeks) | ↓ postprandial insulin | ↓ postprandial C-peptide ↑ C-peptide/insulin quotient |
| TRPV1 Agonist | Animal Model | Dose (Duration) | Responses |
|---|---|---|---|
| Capsiate [75] | Pancreatectomized rats | 0.025% of diet (8 weeks) | ↓ body weight gain ↓ visceral fat ↓ leptin ↓ basal blood glucose ↑ glucose tolerance ↑ insulin sensitivity ↑ pancreatic β-cell mass ↓ pancreatic islet cell apoptosis ↑ ratio of β:α pancreatic cells ↓ hepatic triglyceride content ↑ hepatic glycogen content ↑ pAkt/PEPCK & pAMPK signaling post-TRPV1 activation |
| Dihydrocapsiate [174] | High-fat diet-fed mice | 2 mg/kg/day and 10 mg/kg/day (12 weeks) | ↓ body weight gain ↓ WAT lipid accumulation ↓ BAT lipid accumulation ↓ hepatic triglyceride content ↓ blood triglycerides ↓ blood insulin ↓ glucose intolerance ↑ energy expenditure & mitochondrial biogenesis gene expression ↑ intestinal crypt depth, muscularis thickness & goblet cells ↓ gut Firmicutes ↓ host energy availability ↑ TRPV1 expression & activity |
| Resiniferatoxin [112] | High-fat diet-fed mice | 200 µg/kg (4 weeks) | ↓ body weight gain ↑ locomotor activity |
| 6-gingerol or aza-6-gingerol [175] | High-fat diet-fed mice | 0.06% of diet (12 weeks) | ↓ body weight gain ↓ visceral fat accumulation ↓ leptin ↓ blood insulin ↓ basal blood glucose ↓ glucose intolerance ↓ hepatic lipogenic enzymes |
| Piperine [176] | High-carbohydrate, high-fat diet-fed rats | ~30 mg/kg/day (8 weeks) | ↓ body weight ↓ systolic blood pressure ↓ glucose intolerance ↓ visceral fat accumulation ↓ hepatic fibrosis and fat deposition ↓ cardiac collagen deposition ↑ cardiac function |
| Nonivamide [177] | Moderately overweight men | 0.15 mg/day (12 weeks) | ↓ body fat ↑ postprandial serotonin ↑ satiety |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panchal, S.K.; Bliss, E.; Brown, L. Capsaicin in Metabolic Syndrome. Nutrients 2018, 10, 630. https://doi.org/10.3390/nu10050630
Panchal SK, Bliss E, Brown L. Capsaicin in Metabolic Syndrome. Nutrients. 2018; 10(5):630. https://doi.org/10.3390/nu10050630
Chicago/Turabian StylePanchal, Sunil K., Edward Bliss, and Lindsay Brown. 2018. "Capsaicin in Metabolic Syndrome" Nutrients 10, no. 5: 630. https://doi.org/10.3390/nu10050630
APA StylePanchal, S. K., Bliss, E., & Brown, L. (2018). Capsaicin in Metabolic Syndrome. Nutrients, 10(5), 630. https://doi.org/10.3390/nu10050630