Direct and Long-Term Metabolic Consequences of Lowly vs. Highly-Digestible Starch in the Early Post-Weaning Diet of Mice

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
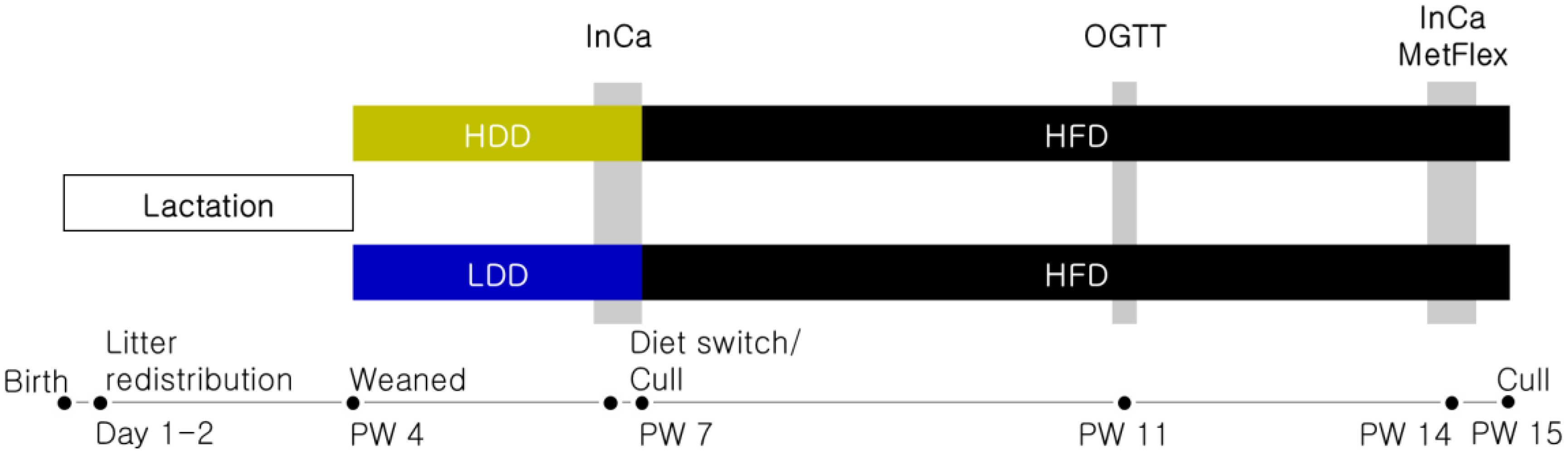
2.1. Animal Model
2.2. Experimental Diets
2.3. Oral Glucose Tolerance Test
2.4. Indirect Calorimetry (InCa) and Metabolic Flexibility
2.5. Sacrifice
2.6. Serum Measurements
2.7. Hepatic Triglycerides and Glycogen Content
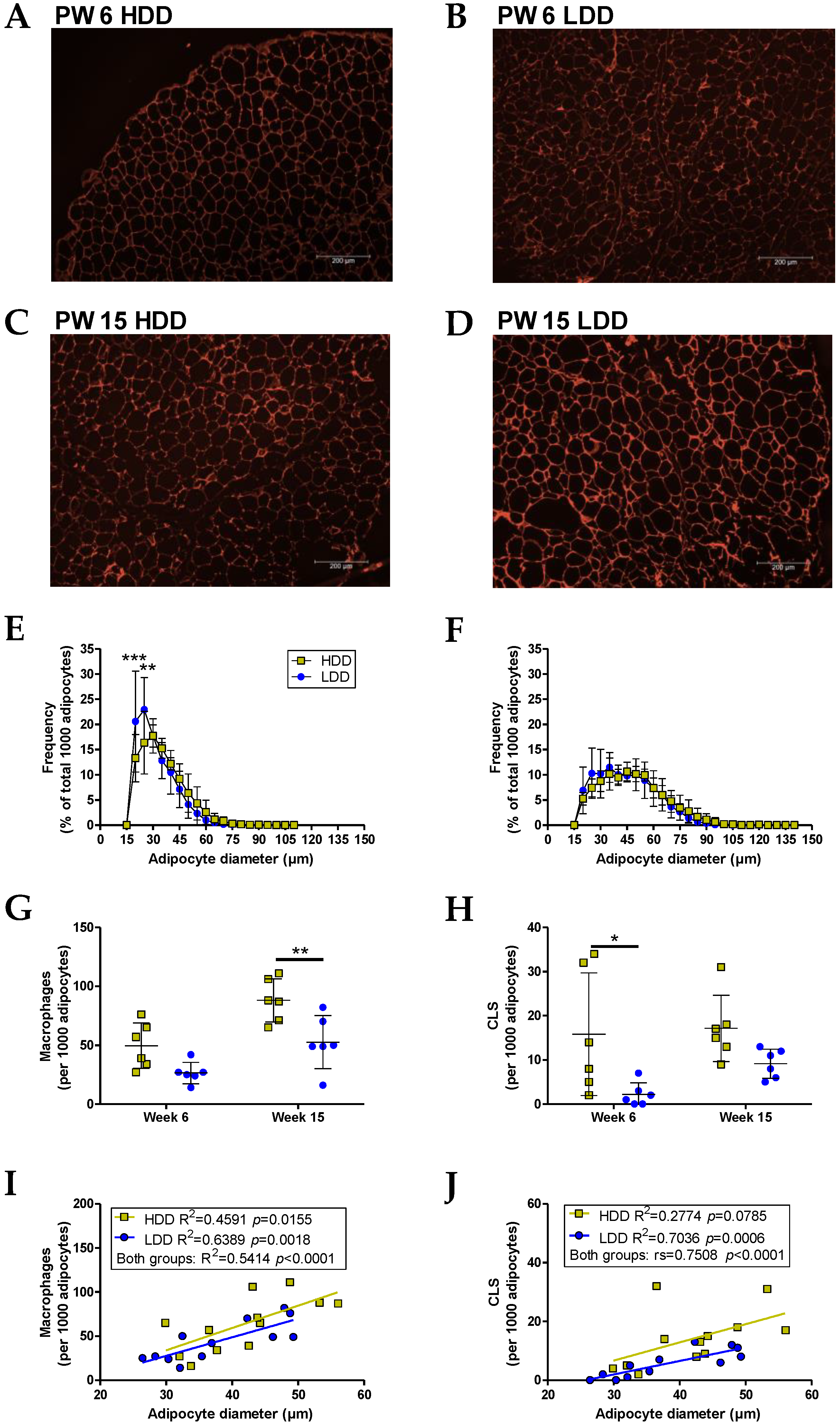
2.8. Gonadal White Adipose Tissue (gWAT) Histology
2.9. Quantitative Real-Time Reverse-Transcription Polymerase Chain Reaction (RT-qPCR)
2.10. Data Analysis
3. Results
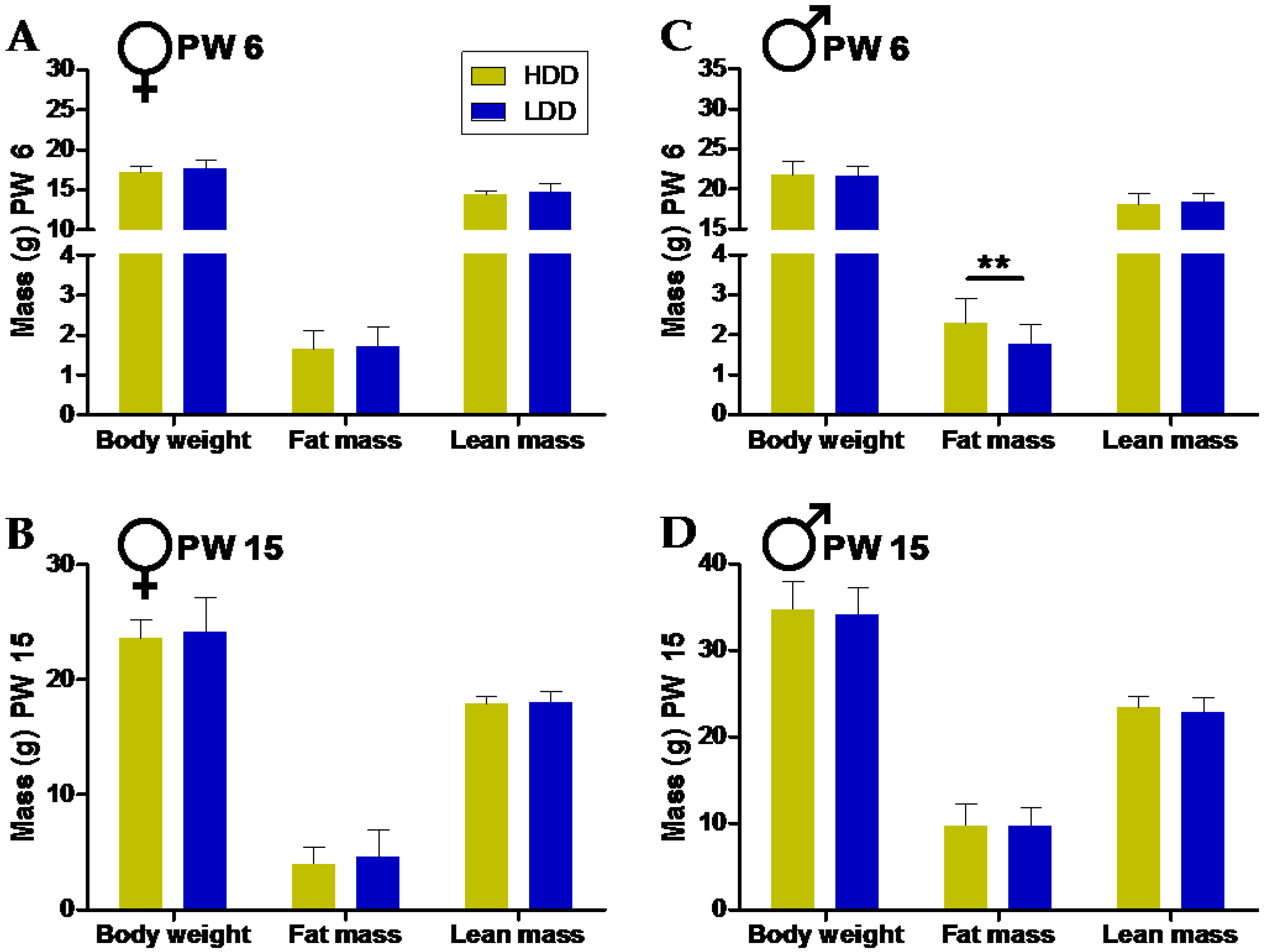
3.1. Direct and Long-Term Effects on Body Weight and Body Composition by Post-Weaning Starches
3.2. Direct and Long-Term Effects on Basal Metabolism
3.3. Direct and Long-Term Effects on Other Physiological Parameters at Sacrifice
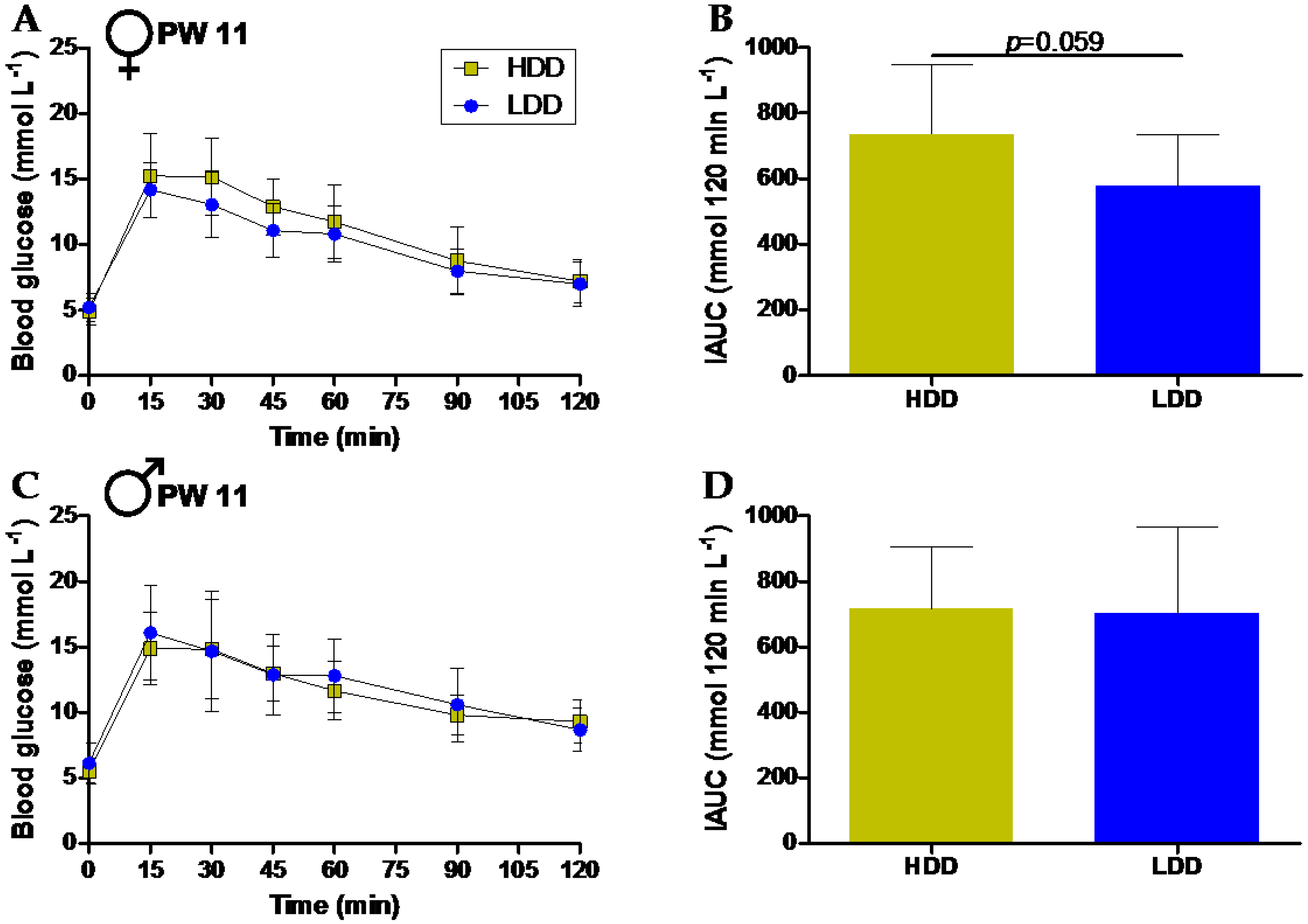
3.4. Long-Term Effects on Glucose Tolerance
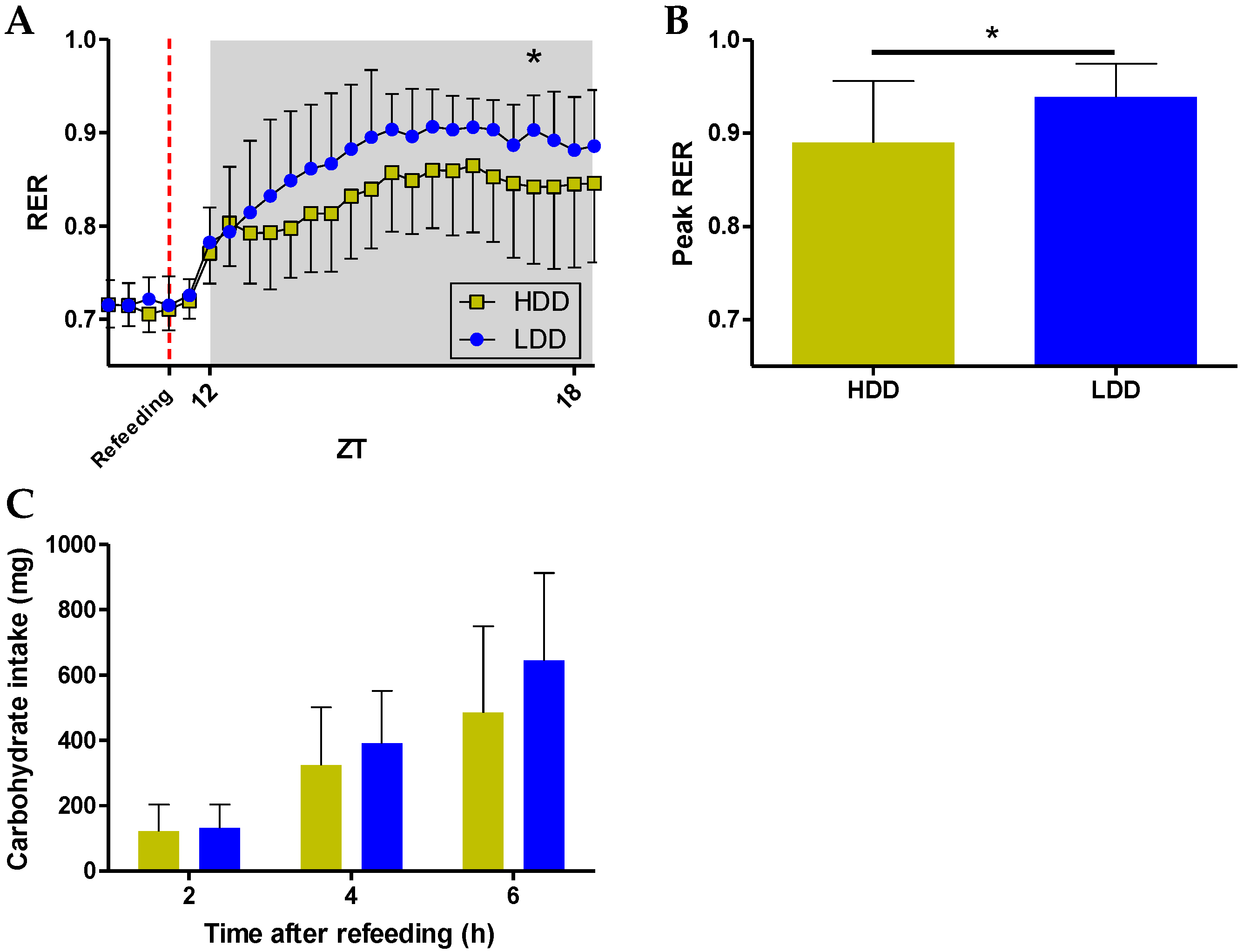
3.5. Long-Term Effects on Metabolic Flexibility
3.6. Direct and Long-Term Effets on Adipose Tissue in Females
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Langley-Evans, S.C. Nutrition in early life and the programming of adult disease: A review. J. Hum. Nutr. Diet. 2015, 28 (Suppl. 1), 1–14. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B.; Demmelmair, H.; Grote, V.; Prell, C.; Weber, M. High protein intake in young children and increased weight gain and obesity risk. Am. J. Clin. Nutr. 2016, 103, 303–304. [Google Scholar] [CrossRef] [PubMed]
- Tang, M. Protein intake during the first two years of life and its association with growth and risk of overweight. Int. J. Environ. Res. Public Health 2018, 15. [Google Scholar] [CrossRef] [PubMed]
- Muhlhausler, B.S.; Ailhaud, G.P. Omega-6 polyunsaturated fatty acids and the early origins of obesity. Curr. Opin. Endocrinol. Diabetes Obes. 2013, 20, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Muhlhausler, B.S.; Gibson, R.A.; Makrides, M. Effect of long-chain polyunsaturated fatty acid supplementation during pregnancy or lactation on infant and child body composition: A systematic review. Am. J. Clin. Nutr. 2010, 92, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, D.S.; Hu, F.B.; Tappy, L.; Brand-Miller, J. Dietary carbohydrates: Role of quality and quantity in chronic disease. BMJ 2018, 361, k2340. [Google Scholar] [CrossRef] [PubMed]
- Schwingshackl, L.; Hobl, L.P.; Hoffmann, G. Effects of low glycaemic index/low glycaemic load vs. high glycaemic index/high glycaemic load diets on overweight/obesity and associated risk factors in children and adolescents: A systematic review and meta-analysis. Nutr. J. 2015, 14, 87. [Google Scholar] [CrossRef] [PubMed]
- Van Schothorst, E.M.; Bunschoten, A.; Schrauwen, P.; Mensink, R.P.; Keijer, J. Effects of a high-fat, low- versus high-glycemic index diet: Retardation of insulin resistance involves adipose tissue modulation. FASEB J. 2009, 23, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Calleja, J.M.S.; Konstanti, P.; Swarts, H.J.M.; Bouwman, L.M.S.; Garcia-Campayo, V.; Billecke, N.; Oosting, A.; Smidt, H.; Keijer, J.; van Schothorst, E.M. Non-invasive continuous real-time in vivo analysis of microbial hydrogen production shows adaptation to fermentable carbohydrates in mice. Sci. Rep. 2018, 8, 15351. [Google Scholar] [CrossRef] [PubMed]
- Elia, M.; Cummings, J.H. Physiological aspects of energy metabolism and gastrointestinal effects of carbohydrates. Eur. J. Clin. Nutr. 2007, 61 (Suppl. 1), S40–S47. [Google Scholar] [CrossRef] [PubMed]
- Shaoul, R.; Tiosano, D.; Hochberg, Z. Evo-devo of child growth: The role of weaning in the transition from infancy to childhood. Crit. Rev. Food Sci. Nutr. 2016, 56, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Wopereis, H.; Oozeer, R.; Knipping, K.; Belzer, C.; Knol, J. The first thousand days–intestinal microbiology of early life: Establishing a symbiosis. Pediatr. Allergy Immunol. 2014, 25, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Fewtrell, M.; Bronsky, J.; Campoy, C.; Domellof, M.; Embleton, N.; Fidler Mis, N.; Hojsak, I.; Hulst, J.M.; Indrio, F.; Lapillonne, A.; et al. Complementary feeding: A position paper by the european society for paediatric gastroenterology, hepatology, and nutrition (ESPGHAN) committee on nutrition. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Woo Baidal, J.A.; Locks, L.M.; Cheng, E.R.; Blake-Lamb, T.L.; Perkins, M.E.; Taveras, E.M. Risk factors for childhood obesity in the first 1000 days: A systematic review. Am. J. Prev. Med. 2016, 50, 761–779. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations. Carbohydrates in Human Nutrition: Report of a Joint FAO/WHO Expert Consultation, Rome, 14–18 April 1997; FAO: Rome, Italy, 1998. [Google Scholar]
- Penfold, N.C.; Ozanne, S.E. Developmental programming by maternal obesity in 2015: Outcomes, mechanisms, and potential interventions. Horm. Behav. 2015, 76, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.S.; Srinivasan, M. Metabolic programming in the immediate postnatal life. Ann. Nutr. Metab. 2011, 58 (Suppl. 2), 18–28. [Google Scholar] [CrossRef] [PubMed]
- Gugusheff, J.; Sim, P.; Kheng, A.; Gentili, S.; Al-Nussairawi, M.; Brand-Miller, J.; Muhlhausler, B. The effect of maternal and post-weaning low and high glycaemic index diets on glucose tolerance, fat deposition and hepatic function in rat offspring. J. Dev. Orig. Health Dis. 2015, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Oosting, A.; van Vlies, N.; Kegler, D.; Schipper, L.; Abrahamse-Berkeveld, M.; Ringler, S.; Verkade, H.J.; van der Beek, E.M. Effect of dietary lipid structure in early postnatal life on mouse adipose tissue development and function in adulthood. Br. J. Nutr. 2014, 111, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Wielinga, P.Y.; Harthoorn, L.F.; Verschuren, L.; Schoemaker, M.H.; Jouni, Z.E.; van Tol, E.A.; Kleemann, R.; Kooistra, T. Arachidonic acid/docosahexaenoic acid-supplemented diet in early life reduces body weight gain, plasma lipids, and adiposity in later life in ApoE*3Leiden mice. Mol. Nutr. Food Res. 2012, 56, 1081–1089. [Google Scholar] [CrossRef] [PubMed]
- Minana-Solis Mdel, C.; Escobar, C. Post-weaning protein malnutrition in the rat produces short and long term metabolic impairment, in contrast to earlier and later periods. Int. J. Biol. Sci. 2008, 4, 422–432. [Google Scholar] [CrossRef] [PubMed]
- He, Y.H.; Li, S.T.; Wang, Y.Y.; Wang, G.; He, Y.; Liao, X.L.; Sun, C.H.; Li, Y. Postweaning low-calcium diet promotes later-life obesity induced by a high-fat diet. J. Nutr. Biochem. 2012, 23, 1238–1244. [Google Scholar] [CrossRef] [PubMed]
- Shahkhalili, Y.; Mace, K.; Moulin, J.; Zbinden, I.; Acheson, K.J. The fat:carbohydrate energy ratio of the weaning diet programs later susceptibility to obesity in male Sprague Dawley rats. J. Nutr. 2011, 141, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Bouwman, L.M.S.; Fernandez-Calleja, J.M.S.; Swarts, H.J.M.; van der Stelt, I.; Oosting, A.; Keijer, J.; van Schothorst, E.M. No adverse programming by post-weaning dietary fructose of body weight, adiposity, glucose tolerance, or metabolic flexibility. Mol. Nutr. Food Res. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Hoevenaars, F.P.; van Schothorst, E.M.; Horakova, O.; Voigt, A.; Rossmeisl, M.; Pico, C.; Caimari, A.; Kopecky, J.; Klaus, S.; Keijer, J. BIOCLAIMS standard diet (BIOsd): A reference diet for nutritional physiology. Genes Nutr. 2012, 7, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Duivenvoorde, L.P.; van Schothorst, E.M.; Swarts, H.M.; Kuda, O.; Steenbergh, E.; Termeulen, S.; Kopecky, J.; Keijer, J. A difference in fatty acid composition of isocaloric high-fat diets alters metabolic flexibility in male C57BL/6JOlaHsd mice. PLoS ONE 2015, 10, e0128515. [Google Scholar] [CrossRef] [PubMed]
- Muoio, D.M. Metabolic inflexibility: When mitochondrial indecision leads to metabolic gridlock. Cell 2014, 159, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Krisman, C.R. A method for the colorimetric estimation of glycogen with iodine. Anal. Biochem. 1962, 4, 17–23. [Google Scholar] [CrossRef]
- Cinti, S.; Mitchell, G.; Barbatelli, G.; Murano, I.; Ceresi, E.; Faloia, E.; Wang, S.; Fortier, M.; Greenberg, A.S.; Obin, M.S. Adipocyte death defines macrophage localization and function in adipose tissue of obese mice and humans. J. Lipid Res. 2005, 46, 2347–2355. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, A.E.; Jones, T.R.; Lamprecht, M.R.; Clarke, C.; Kang, I.H.; Friman, O.; Guertin, D.A.; Chang, J.H.; Lindquist, R.A.; Moffat, J.; et al. Cellprofiler: Image analysis software for identifying and quantifying cell phenotypes. Genome Biol. 2006, 7, R100. [Google Scholar] [CrossRef] [PubMed]
- Cellprofiler. Available online: http://forum.cellprofiler.org/t/adipocyte-h-e-cell-profiler-pipeline/1460 (accessed on 31 July 2018).
- Van Schothorst, E.M.; Franssen-van Hal, N.; Schaap, M.M.; Pennings, J.; Hoebee, B.; Keijer, J. Adipose gene expression patterns of weight gain suggest counteracting steroid hormone synthesis. Obes. Res. 2005, 13, 1031–1041. [Google Scholar] [CrossRef] [PubMed]
- Fischbach, M.A.; Sonnenburg, J.L. Eating for two: How metabolism establishes interspecies interactions in the gut. Cell Host Microbe 2011, 10, 336–347. [Google Scholar] [CrossRef] [PubMed]
- Van Ommen, B.; Keijer, J.; Heil, S.G.; Kaput, J. Challenging homeostasis to define biomarkers for nutrition related health. Mol. Nutr. Food Res. 2009, 53, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Stroeve, J.H.; van Wietmarschen, H.; Kremer, B.H.; van Ommen, B.; Wopereis, S. Phenotypic flexibility as a measure of health: The optimal nutritional stress response test. Genes Nutr. 2015, 10, 459. [Google Scholar] [CrossRef] [PubMed]
- Horakova, O.; Medrikova, D.; van Schothorst, E.M.; Bunschoten, A.; Flachs, P.; Kus, V.; Kuda, O.; Bardova, K.; Janovska, P.; Hensler, M.; et al. Preservation of metabolic flexibility in skeletal muscle by a combined use of n-3 PUFA and rosiglitazone in dietary obese mice. PLoS ONE 2012, 7, e43764. [Google Scholar] [CrossRef] [PubMed]
- Sparks, L.M.; Ukropcova, B.; Smith, J.; Pasarica, M.; Hymel, D.; Xie, H.; Bray, G.A.; Miles, J.M.; Smith, S.R. Relation of adipose tissue to metabolic flexibility. Diabetes Res. Clin. Pract. 2009, 83, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Boutens, L.; Stienstra, R. Adipose tissue macrophages: Going off track during obesity. Diabetologia 2016, 59, 879–894. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Moschen, A.R. Food, immunity, and the microbiome. Gastroenterology 2015, 148, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid. Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Zeevi, D.; Korem, T.; Zmora, N.; Israeli, D.; Rothschild, D.; Weinberger, A.; Ben-Yacov, O.; Lador, D.; Avnit-Sagi, T.; Lotan-Pompan, M.; et al. Personalized nutrition by prediction of glycemic responses. Cell 2015, 163, 1079–1094. [Google Scholar] [CrossRef] [PubMed]
- Muñiz Pedrogo, D.A.; Jensen, M.D.; Van Dyke, C.T.; Murray, J.A.; Woods, J.A.; Chen, J.; Kashyap, P.C.; Nehra, V. Gut microbial carbohydrate metabolism hinders weight loss in overweight adults undergoing lifestyle intervention with a volumetric diet. Mayo Clin. Proc. 2018, 93, 1104–1110. [Google Scholar] [CrossRef] [PubMed]
- Bindels, L.B.; Segura Munoz, R.R.; Gomes-Neto, J.C.; Mutemberezi, V.; Martinez, I.; Salazar, N.; Cody, E.A.; Quintero-Villegas, M.I.; Kittana, H.; de Los Reyes-Gavilan, C.G.; et al. Resistant starch can improve insulin sensitivity independently of the gut microbiota. Microbiome 2017, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Darko, K.O.; Huang, Y.; He, C.; Yang, H.; He, S.; Li, J.; Li, J.; Hocher, B.; Yin, Y. Resistant starch regulates gut microbiota: Structure, biochemistry and cell signalling. Cell Physiol. Biochem. 2017, 42, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.J.; Senior, A.M.; Bell-Anderson, K.S. Metabolic effects of high glycaemic index diets: A systematic review and meta-analysis of feeding studies in mice and rats. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.A. Resistant starch and energy balance: Impact on weight loss and maintenance. Crit. Rev. Food Sci. Nutr. 2014, 54, 1158–1166. [Google Scholar] [CrossRef] [PubMed]
- Grove, K.L.; Fried, S.K.; Greenberg, A.S.; Xiao, X.Q.; Clegg, D.J. A microarray analysis of sexual dimorphism of adipose tissues in high-fat-diet-induced obese mice. Int. J. Obes. 2010, 34, 989–1000. [Google Scholar] [CrossRef] [PubMed]
- Karastergiou, K.; Fried, S.K. Cellular mechanisms driving sex differences in adipose tissue biology and body shape in humans and mouse models. Adv. Exp. Med. Biol. 2017, 1043, 29–51. [Google Scholar] [CrossRef] [PubMed]
- Charrier, J.A.; Martin, R.J.; McCutcheon, K.L.; Raggio, A.M.; Goldsmith, F.; Goita, M.; Senevirathne, R.N.; Brown, I.L.; Pelkman, C.; Zhou, J.; et al. High fat diet partially attenuates fermentation responses in rats fed resistant starch from high-amylose maize. Obesity 2013, 21, 2350–2355. [Google Scholar] [CrossRef] [PubMed]
- Topping, D.L.; Clifton, P.M. Short-chain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.E.; Tang, Z.; Kerbois, C.; Delville, C.; Konstantopedos, P.; Bruel, A.; Derous, D.; Green, C.; Aspden, R.M.; Goodyear, S.R.; et al. The effects of graded levels of calorie restriction: I. Impact of short term calorie and protein restriction on body composition in the C57BL/6 mouse. Oncotarget 2015, 6, 15902–15930. [Google Scholar] [CrossRef] [PubMed]
- Aiken, C.E.; Ozanne, S.E. Sex differences in developmental programming models. Reproduction 2013, 145, R1–R13. [Google Scholar] [CrossRef] [PubMed]
- Della Torre, S.; Mitro, N.; Meda, C.; Lolli, F.; Pedretti, S.; Barcella, M.; Ottobrini, L.; Metzger, D.; Caruso, D.; Maggi, A. Short-term fasting reveals amino acid metabolism as a major sex-discriminating factor in the liver. Cell Metab. 2018, 28, 256–267.e5. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Wang, L.; Yang, D.; Li, L.; Togo, J.; Wu, Y.; Liu, Q.; Li, B.; Li, M.; Wang, G.; et al. Dietary fat, but not protein or carbohydrate, regulates energy intake and causes adiposity in mice. Cell Metab. 2018, 28, 415–431.e4. [Google Scholar] [CrossRef] [PubMed]
- Oosting, A.; Kegler, D.; Wopereis, H.J.; Teller, I.C.; van de Heijning, B.J.; Verkade, H.J.; van der Beek, E.M. Size and phospholipid coating of lipid droplets in the diet of young mice modify body fat accumulation in adulthood. Pediatr. Res. 2012, 72, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Baars, A.; Oosting, A.; Engels, E.; Kegler, D.; Kodde, A.; Schipper, L.; Verkade, H.J.; van der Beek, E.M. Milk fat globule membrane coating of large lipid droplets in the diet of young mice prevents body fat accumulation in adulthood. Br. J. Nutr. 2016, 115, 1930–1937. [Google Scholar] [CrossRef] [PubMed]
- Vickers, M.H. Early life nutrition, epigenetics and programming of later life disease. Nutrients 2014, 6, 2165–2178. [Google Scholar] [CrossRef] [PubMed]
- Reichetzeder, C.; Dwi Putra, S.E.; Li, J.; Hocher, B. Developmental origins of disease—Crisis precipitates change. Cell. Physiol. Biochem. 2016, 39, 919–938. [Google Scholar] [CrossRef] [PubMed]
- Miles, J.L.; Huber, K.; Thompson, N.M.; Davison, M.; Breier, B.H. Moderate daily exercise activates metabolic flexibility to prevent prenatally induced obesity. Endocrinology 2009, 150, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Holness, M.J.; Zariwala, G.; Walker, C.G.; Sugden, M.C. Adipocyte pyruvate dehydrogenase kinase 4 expression is associated with augmented ppargamma upregulation in early-life programming of later obesity. FEBS Open Bio 2012, 2, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.L.; Soeters, M.R.; Wust, R.C.I.; Houtkooper, R.H. Metabolic flexibility as an adaptation to energy resources and requirements in health and disease. Endocr. Rev. 2018. [Google Scholar] [CrossRef] [PubMed]
- Marco, A.; Kisliouk, T.; Weller, A.; Meiri, N. High fat diet induces hypermethylation of the hypothalamic pomc promoter and obesity in post-weaning rats. Psychoneuroendocrinology 2013, 38, 2844–2853. [Google Scholar] [CrossRef] [PubMed]
- Kellermayer, R.; Balasa, A.; Zhang, W.; Lee, S.; Mirza, S.; Chakravarty, A.; Szigeti, R.; Laritsky, E.; Tatevian, N.; Smith, C.W.; et al. Epigenetic maturation in colonic mucosa continues beyond infancy in mice. Hum. Mol. Genet. 2010, 19, 2168–2176. [Google Scholar] [CrossRef] [PubMed]
- Gluckman, P.D.; Hanson, M.A.; Buklijas, T. A conceptual framework for the developmental origins of health and disease. J. Dev. Orig. Health Dis. 2010, 1, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.J.; Atkinson, F.S.; Ramalingam, N.; Buyken, A.E.; Brand-Miller, J.C. Effects of human milk and formula on postprandial glycaemia and insulinaemia. Eur. J. Clin. Nutr. 2015, 69, 939–943. [Google Scholar] [CrossRef] [PubMed]
- Le Huerou-Luron, I.; Blat, S.; Boudry, G. Breast- v. Formula-feeding: Impacts on the digestive tract and immediate and long-term health effects. Nutr. Res. Rev. 2010, 23, 23–36. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | HDD | LDD | HFD |
|---|---|---|---|
| Casein | 212.2 | 212.0 | 233.5 |
| l-Cysteine | 3.0 | 3.0 | 3.0 |
| Amylose mix (AmyloGel 03003) 1 | 0.0 | 568.6 | 0.0 |
| Amylopectin (C*Gel 04201) 2 | 568.6 | 0.0 | 0.0 |
| Wheat starch | 0.0 | 0.0 | 285.6 |
| Maltodextrin | 0.0 | 0.0 | 100.0 |
| Glucose | 0.0 | 0.0 | 70.0 |
| Coconut oil | 21.4 | 21.4 | 0.0 |
| Sunflower oil | 83.1 | 83.1 | 0.0 |
| Flaxseed oil | 14.2 | 14.2 | 4.0 |
| Palm oil | 0.0 | 0.0 | 206.3 |
| Cholesterol | 0.03 | 0.03 | 0.097 |
| Cellulose | 50.0 | 50.0 | 50.0 |
| Mineral mix (AIN-93G-MX) | 35.0 | 35.0 | 35.0 |
| Vitamin mix (AIN-93-VX) | 10.0 | 10.0 | 10.0 |
| Choline bitartrate | 2.5 | 2.5 | 2.5 |
| Total (g) | 1000.0 | 1000.0 | 1000.0 |
| Gross energy density (kJ g1) 3 | 18.9 | 19.5 | 20.8 |
| Calculated energy density (kJ g−1) 4 | 17.9 | 17.9 | 19.8 |
| Protein (energy%) | 20.1 | 20.1 | 20.0 |
| Carbohydrate (energy%) | 54.9 | 54.9 | 40.0 |
| Fat (energy%) | 25.0 | 25.0 | 40.0 |
| Females | Males | |||||||
|---|---|---|---|---|---|---|---|---|
| PW 6 | PW 14 | PW 6 | PW 14 | |||||
| Parameter | HDD | LDD | HDD | LDD | HDD | LDD | HDD | LDD |
| EE (24 h, kJ h−1) | 1.60 ± 0.08 | 1.65 ± 0.09 | 1.90 ± 0.12 | 1.90 ± 0.12 | 1.80 ± 0.12 | 1.77 ± 0.11 | 2.11 ± 0.25 | 2.15 ± 0.25 |
| EE (LP, kJ h−1) | 1.46 ± 0.08 | 1.52 ± 0.11 | 1.78 ± 0.13 | 1.79 ± 0.11 | 1.66 ± 0.13 | 1.61 ± 0.12 | 2.01 ± 0.23 | 2.04 ± 0.24 |
| EE (DP, kJ h−1) | 1.75 ± 0.09 | 1.78 ± 0.09 | 2.03 ± 0.13 | 2.02 ± 0.14 | 1.95 ± 0.12 | 1.93 ± 0.10 | 2.21 ± 0.27 | 2.27 ± 0.26 |
| RER (24 h) | 0.84 ± 0.04 | 0.84 ± 0.04 | 0.86 ± 0.04 | 0.86 ± 0.04 | 0.88 ± 0.03 | 0.85 ± 0.03 # | 0.85 ± 0.02 | 0.85 ± 0.02 |
| RER (LP) | 0.81 ± 0.05 | 0.82 ± 0.05 | 0.86 ± 0.04 | 0.86 ± 0.04 | 0.85 ± 0.03 | 0.82 ± 0.04 * | 0.85 ± 0.03 | 0.86 ± 0.02 |
| RER (DP) | 0.87 ± 0.05 | 0.87 ± 0.04 | 0.86 ± 0.05 | 0.86 ± 0.04 | 0.91 ± 0.03 | 0.88 ± 0.02 * | 0.84 ± 0.02 | 0.84 ± 0.02 |
| Activity (24 h, counts × 104) | 6.93 (6.13, 7.45) | 6.55 (5.88, 6.72) | 5.07 (3.89, 7.07) | 4.56 (3.85, 6.23) | 4.95 (4.13, 6.06) | 5.43 (4.43, 5.70) | 2.93 (2.31, 3.81) | 2.98 (2.37, 3.51) |
| Activity (LP, counts × 104) | 1.46 (1.35, 1.83) | 1.46 (1.27, 1.75) | 1.36 (1.02, 1.89) | 1.03 (0.84, 1.71) | 1.4 (1.06, 1.74) | 1.10 (0.97, 1.28) | 0.80 (0.66, 1.39) | 0.84 (0.71, 1.00) |
| Activity (DP, counts × 104) | 5.45 (4.52, 5.57) | 4.47 (4.32, 5.06) | 3.42 (2.81, 5.24) | 3.35 (2.80, 4.73) | 3.65 (2.99, 4.28) | 4.31 (3.37, 4.33) | 1.88 (1.56, 2.50) | 2.07 (1.64, 2.54) |
| H2 (24 h, mL) | 0.18 (0.14, 0.26) | 1.64 (1.14, 2.12) § | 0.24 (0.21, 0.32) | 0.34 (0.28, 0.39) | 0.24 (0.18, 0.35) | 1.47 (1.11, 1.90) § | 0.46 (0.36, 0.75) | 0.38 (0.32, 0.72) |
| H2 (LP, mL) | 0.08 (0.06, 0.13) | 0.60 (0.46, 0.74) § | 0.10 (0.09, 0.16) | 0.19 (0.14, 0.20) | 0.10 (0.08, 0.14) | 0.52 (0.40, 0.61) § | 0.21 (0.16, 0.35) | 0.18 (0.15, 0.36) |
| H2 (DP, mL) | 0.11 (0.07, 0.14) | 1.07 (0.66, 1.41) § | 0.14 (0.11, 0.17) | 0.15 (0.14, 0.20) | 0.14 (0.09, 0.21) | 0.99 (0.67, 1.32) § | 0.25 (0.20, 0.40) | 0.21 (0.17, 0.36) |
| Females | Males | |||||||
|---|---|---|---|---|---|---|---|---|
| PW 6 | PW 15 | PW 6 | PW 15 | |||||
| Parameter | HDD | LDD | HDD | LDD | HDD | LDD | HDD | LDD |
| Cumulative GE intake (MJ) | 0.90 (0.87, 0.91) | 1.01 (0.98, 1.03) § | 3.36 (3.26, 3.51) | 3.24 (3.12, 3.50) | 1.00 (0.97, 1.03) | 1.13 (1.10, 1.15) § | 3.81 (3.61, 4.09) | 3.73 (3.58, 4.06) |
| gWAT (mg) | 65 (58, 83) | 65 (48, 77) | 197 (178, 297) | 240 (181, 422) | 140 (122, 168) | 117 (91, 145) | 667 (568, 758) | 796 (592, 885) |
| mWAT (mg) | 104 (83, 139) | 108 (81, 114) | 202 (183, 303) | 244 (180, 383) | 176 (150, 200) | 140 (117, 173) | 562 (474, 797) | 627 (523, 806) |
| Liver (g) | 0.72 (0.66, 0.77) | 0.67 (0.62, 0.75) | 0.89 (0.85, 0.95) | 0.89 (0.83, 1.01) | 1.01 (0.93, 1.06) | 1.05 (0.96, 1.11) | 1.20 (1.08, 1.52) | 1.23 (1.11, 1.49) |
| Liver (g g−1 body weight) | 0.042 (0.039, 0.044) | 0.039 (0.036, 0.042) | 0.039 (0.037, 0.039) | 0.039 (0.036, 0.040) | 0.048 (0.044, 0.049) | 0.049 (0.045, 0.051) | 0.036 (0.033, 0.042) | 0.036 (0.034, 0.041) |
| Liver TG (mg g−1 wet tissue) | n.m. | n.m. | 36.1 ± 13.9 | 37.6 ± 14.2 | n.m. | n.m. | n.m. | n.m. |
| Liver glycogen (mg g−1 wet tissue) | n.m. | n.m. | 53.3 ± 24.0 | 68.7 ± 39.2 | n.m. | n.m. | n.m. | n.m. |
| Pancreas (mg) | 235 (200, 262) | 255 (228, 296) | 322 (299, 347) | 319 (281, 380) | 261 (254, 287) | 282 (248, 303) | 383 (344, 534) | 363 (326, 489) |
| Small intestine (cm) | 31.9 (30.8, 32.2) | 33.8 (32.8, 36.3) # | 33.3 (32.5, 33.7) | 33.6 (32.9, 34.8) | 33.3 (32.8, 34.0) | 35.4 (35.0, 37.2) § | 35.0 (34.4, 36.8) | 35.7 (34.3, 37.4) |
| Small intestine (g) | 0.60 (0.56, 0.61) | 0.72 (0.67, 0.84) ¥ | 0.73 (0.69, 0.74) | 0.72 (0.68, 0.79) | 0.68 (0.65, 0.70) | 0.78 (0.74, 0.89) § | 0.84 (0.80, 0.94) | 0.84 (0.79, 0.94) |
| Cecum contents (mg) | 108 (90, 116) | 225 (204, 295) § | 104 (91, 125) | 108 (87, 143) | 152 (121, 162) | 273 (250, 359) § | 196 (164, 203) | 140 (123, 171) * |
| Colon (mg) | 92 (87, 97) | 135 (126, 152) § | 118 (109, 122) | 112 (109, 123) | 104 (97, 109) | 153 (141, 166) § | 139 (126, 151) | 136 (129, 159) |
| Blood glucose (mmol L−1) | 5.0 ± 0.7 | 4.7 ± 1.0 | 4.9 ± 0.5 | 5.3 ± 0.9 | 5.8 ± 0.9 | 6.2 ± 0.7 | 5.6 ± 0.9 | 5.8 ± 0.9 |
| Serum insulin (ng mL−1) | 0.73 (0.56, 0.87) | 0.60 (0.48, 0.72) | 0.62 (0.58, 1.11) | 0.83 (0.65, 1.60) | 1.03 (0.90, 1.12) | 0.83 (0.72, 1.04) | 2.09 (1.71, 3.20) | 2.22 (1.93, 2.61) |
| Serum leptin (ng mL−1) | 1.6 (1.2, 2.7) | 1.3 (0.9, 2.5) | 3.9 (2.9, 8,0) | 5.0 (2.8, 13.6) | 2.9 (2.1, 3.3) | 1.8 (1.3, 2.1) # | 52.1 (22.2, 125.9) | 65.3 (41.6, 101.6) |
| Serum adiponectin (µg mL−1) | n.m. | n.m. | 12.3 ± 1.1 | 12.2 ± 1.5 | n.m. | n.m. | n.m. | n.m. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Calleja, J.M.S.; Bouwman, L.M.S.; Swarts, H.J.M.; Oosting, A.; Keijer, J.; Van Schothorst, E.M. Direct and Long-Term Metabolic Consequences of Lowly vs. Highly-Digestible Starch in the Early Post-Weaning Diet of Mice. Nutrients 2018, 10, 1788. https://doi.org/10.3390/nu10111788
Fernández-Calleja JMS, Bouwman LMS, Swarts HJM, Oosting A, Keijer J, Van Schothorst EM. Direct and Long-Term Metabolic Consequences of Lowly vs. Highly-Digestible Starch in the Early Post-Weaning Diet of Mice. Nutrients. 2018; 10(11):1788. https://doi.org/10.3390/nu10111788
Chicago/Turabian StyleFernández-Calleja, José M. S., Lianne M. S. Bouwman, Hans J. M. Swarts, Annemarie Oosting, Jaap Keijer, and Evert M. Van Schothorst. 2018. "Direct and Long-Term Metabolic Consequences of Lowly vs. Highly-Digestible Starch in the Early Post-Weaning Diet of Mice" Nutrients 10, no. 11: 1788. https://doi.org/10.3390/nu10111788
APA StyleFernández-Calleja, J. M. S., Bouwman, L. M. S., Swarts, H. J. M., Oosting, A., Keijer, J., & Van Schothorst, E. M. (2018). Direct and Long-Term Metabolic Consequences of Lowly vs. Highly-Digestible Starch in the Early Post-Weaning Diet of Mice. Nutrients, 10(11), 1788. https://doi.org/10.3390/nu10111788