Potential Prebiotic Properties of Nuts and Edible Seeds and Their Relationship to Obesity


Abstract
1. Introduction
2. Materials and Methods
3. Relationship between Nut Consumption and Body Adiposity
4. Nutritional Attributes and Bioactive Compounds of Nuts and Edible Seeds
5. Potential of Nut and Edible Seed Consumption on the Remodeling of Gut Microbiota
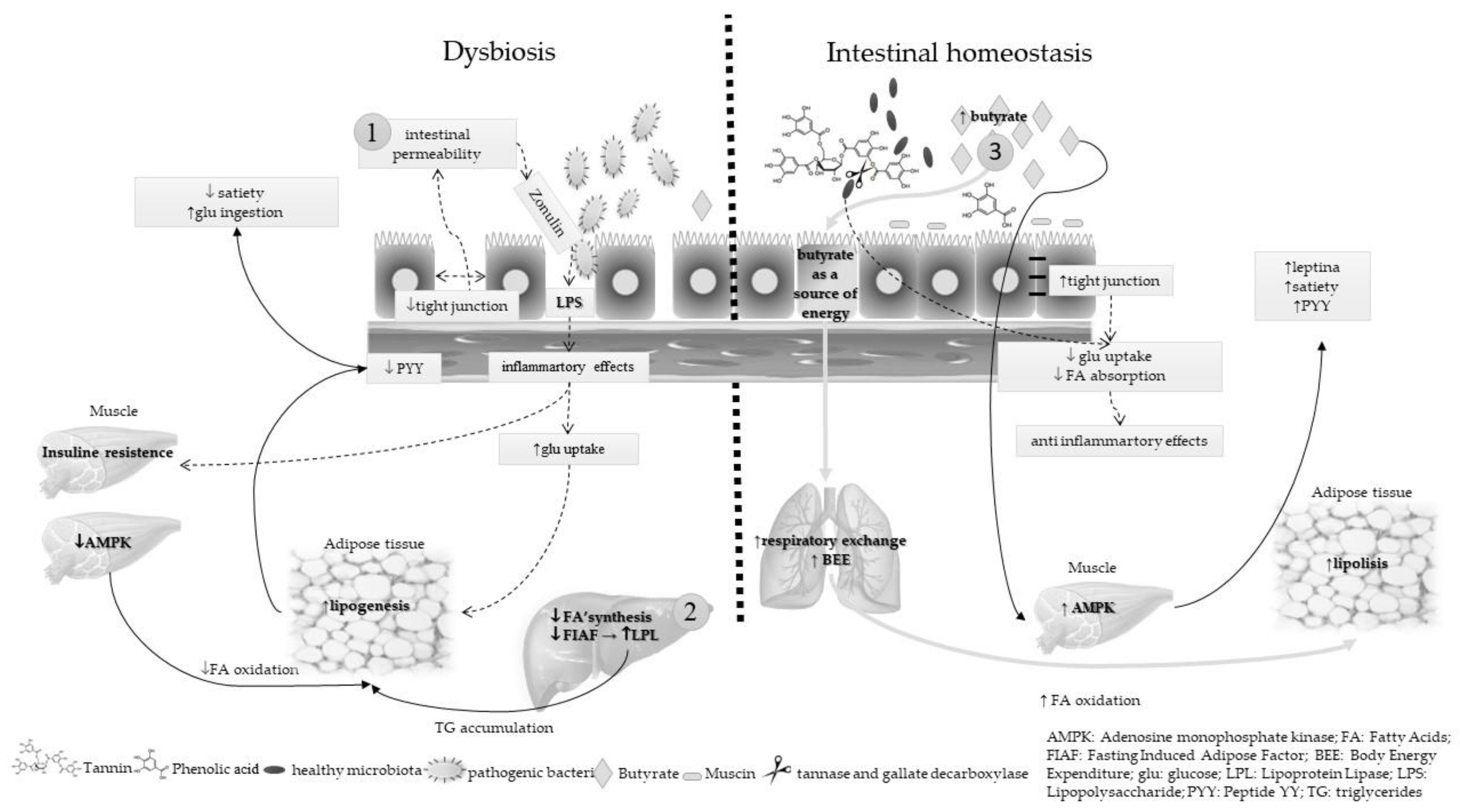
6. Microbiota and Energy Metabolism: Dysbiosis and Intestinal Homeostasis
7. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Samouda, H.; Ruiz-Castell, M.; Bocquet, V.; Kuemmerle, A.; Chioti, A.; Dadoun, F.; Kandala, N.; Stranges, S. Geographical variation of overweight, obesity and related risk factors: Findings from the European health examination survey in Luxembourg, 2013–2015. PLoS ONE 2018, 6, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Conklin, A.I.; Daoud, A.; Shimkhada, R.; Ponce, N.A. The impact of rising food prices on obesity in women: A longitudinal analysis of 31 low-income and middle-income countries from 2000 to 2014. Int. J. Obes. 2018, 17, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bes-Rastrollo, M.; Wedick, N.M.; Martinez-Gonzalez, M.A.; Li, T.Y.; Sampson, L.; Hu, F.B. Prospective study of nut consumption, long-term weight change, and obesity risk in women. Am. J. Clin. Nutr. 2009, 89, 1913–1919. [Google Scholar] [CrossRef] [PubMed]
- Hosseinpour-Niazi, S.; Hosseini, S.; Mirmiran, P.; Azizi, F. Prospective study of nut consumption and incidence of metabolic syndrome: Tehran lipid and glucose study. Nutrients 2017, 9, 1056. [Google Scholar] [CrossRef] [PubMed]
- Jaceldo-Siegl, K.; Haddad, E.; Oda, K.; Fraser, G.E.; Sabaté, J. Tree nuts are inversely associated with metabolic syndrome and obesity: The Adventist health study-2. PLoS ONE 2014, 9, 85133. [Google Scholar] [CrossRef] [PubMed]
- Mejia, S.B.; Kendall, C.W.C.; Viguiliouk, E.; Augustin, L.S.; Há, V.; Cozma, A.I.; Mirrahimi, A.; Maroleanu, A.; Chiavaroli, L.; Leiter, L.A.; et al. Effect of tree nuts on metabolic syndrome criteria: A systematic review and meta-analysis of randomized controlled trials. BMJ Open 2014, 4, e004660. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Pan, A.; Yu, Z.; Qi, Q.; Lu, L.; Zhang, G.; Yu, D.; Zong, G.; Zhou, Y.; Chen, X.; et al. Lifestyle counseling and supplementation with flaxseed or walnuts influence the management of metabolic syndrome. J. Nutr. 2010, 11, 1937–1942. [Google Scholar] [CrossRef] [PubMed]
- Foster, G.D.; Shantz, K.L.; Veur, S.S.V.; Oliver, T.L.; Lent, M.R.; Virus, A.; Szapary, P.O.; Rader, D.J.; Zemel, B.S.; Gilden-Tsai, A. A randomized trial of the effects of an almond-enriched, hypocaloric diet in the treatment of obesity. Am. J. Clin. Nutr. 2012, 2, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Wien, M.A.; Sabaté, J.M.; Iklé, D.N.; Cole, S.E.; Kandeel, F.R. Almonds vs. complex carbohydrates in a weight reduction program. Int. J. Obes. Relat. Metab. Disord. 2003, 27, 1365–1372. [Google Scholar] [CrossRef] [PubMed]
- Somerset, S.M.; Graham, A.; Markwell, L.B.K. Isoenergetic replacement of dietary saturated with monounsaturated fat via macadamia nuts enhances endothelial function in overweight subjects. E-SPEN J. 2013, 8, 113–119. [Google Scholar] [CrossRef]
- Gulati, S.; Misra, A.; Pandey, R.M.; Bhatt, S.P.; Saluja, S. Effects of pistachio nuts on body composition, metabolic, inflammatory and oxidative stress parameters in Asian Indians with metabolic syndrome: A 24-wk, randomized control trial. Nutrition 2014, 30, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, J.; Tan, S.Y.; Mattes, R.D. Almond consumption during energy restriction lowers truncal fat and blood pressure in compliant overweight or obese adults. J. Nutr. 2016, 12, 2513–2519. [Google Scholar] [CrossRef] [PubMed]
- Kendall, C.W.C.; West, S.G.; Augustin, L.S.; Esfahani, A.; Vidgen, E.; Bashyam, B.; Sauder, K.A.; Campbell, J.; Chiavaroli, L.; Jenkins, A.L.; et al. Acute effects of pistachio consumption on glucose and insulin, satiety hormones and endothelial function in the metabolic syndrome. Eur. J. Clin. Nutr. 2014, 68, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Keogh, J.B.; Clifton, P.M. Benefits of nut consumption on insulin resistance and cardiovascular risk factors: Multiple potential mechanisms of actions. Nutrients 2017, 9, 1271. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, M.; Sathe, S.K. Chemical composition of selected edible nut seeds. J. Agric. Food Chem. 2006, 54, 4705–4714. [Google Scholar] [CrossRef] [PubMed]
- Kornsteiner, M.; Wagner, K.; Elmadfa, I. Tocopherols and total phenolics in 10 different nut types. Food Chem. 2006, 98, 381–387. [Google Scholar] [CrossRef]
- Yang, J. Brazil nuts and associated health benefits: A review. LWT-Food Sci. Technol. 2009, 42, 1573–1580. [Google Scholar] [CrossRef]
- Carrera-Quintanar, L.; López Roa, R.I.; Quintero-Fabián, S.; Sánchez-Sánchez, M.A.; Vizmanos, B.; Ortuño-Sahagún, D. Phytochemicals that influence gut microbiota as prophylactics and for the treatment of obesity and inflammatory diseases. Mediators Inflamm. 2018, 2018, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Vidra, N.; Bijlsma, M.J.; Trias-Llimo´s, S.; Janssen, F. Past trends in obesity-attributable mortality in eight European countries: An application of age-period-cohort analysis. Int. J. Public. Health 2018, 63, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, D.K.; Renuka; Puniya, M.; Shandilya, U.K.; Dhewa, T.; Kumar, N.; Kumar, S.; Puniya, A.K.; Shukla, P. Gut microbiota modulation and its relationship with obesity using prebiotic fibers and probiotics: A review. Front. Microbiol. 2017, 8, 563–580. [Google Scholar] [CrossRef] [PubMed]
- Rock, C.L.; Flatt, S.W.; Barkai, H.S.; Pakiz, B.; Heath, D.D. Walnut consumption in a weight reduction intervention: Effects on body weight, biological measures, blood pressure and satiety. Nutr. J. 2017, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tapsell, L.C.; Batterham, M.J.; Teuss, G.; Tan, S.Y.; Dalton, S.; Quick, C.J.; Gillen, L.J.; Charlton, K.E. Long-term effects of increased dietary polyunsaturated fat from walnuts on metabolic parameters in type II diabetes. Eur. J. Clin. Nutr. 2009, 63, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Song, R.; Nguyen, C.; Zerlin, A.; Karp, H.; Naowamondhol, K.; Thames, G.; Gao, K.; Li, L.; Tseng, C.H.; et al. Pistachio nuts reduce triglycerides and body weight by comparison to refined carbohydrate snack in obese subjects on a 12-week weight loss program. J. Am. Coll. Nutr. 2010, 29, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Bitok, E.; Rajaram, S.; Jaceldo-Siegl, K.; Oda, K.; Sala-Vila, A.; Serra-Mir, M.; Ros, E.; Sabaté, J. Effects of long-term walnut supplementation on body weight in free-living elderly: Results of a randomized controlled trial. Nutrients 2018, 10, 1317. [Google Scholar] [CrossRef] [PubMed]
- U.S. Department of Agriculture. Dietary Guide for Americans 2010; USDA: Washington, DC, USA, 2010.
- Bento, A.P.N.; Cominetti, C.; Simões Filho, A.; Naves, M.M.V. Baru almond improves lipid profile in mildly hypercholesterolemic subjects: A randomized, controlled, crossover study. Nutr. Metabol. Cardiov. Dis. 2014, 24, 1330–1336. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Park, W.J. Unsaturated fatty acids, desaturases, and human health. J. Med. Food 2014, 17, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.G.O.; Fernandes, D.C.; Alves, A.M.; Freitas, J.B.; Naves, M.M.V. Nutritional quality and protein value of exotic almonds and nut from the Brazilian Savanna compared to peanut. Food Res. Inter. 2011, 44, 2319–2325. [Google Scholar] [CrossRef]
- Institute of Medicine. Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium, and Carotenoids; National Academy Press: Washington, DC, USA, 2000. [Google Scholar]
- Wong, S.K.; Chin, K.-Y.; Suhaimi, F.H.; Ahmad, F.; Ima-Nirwan, S. Vitamin E as a potential interventional treatment for metabolic syndrome: Evidence from animal and human studies. Front. Pharmacol. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, M.; Katsuragi, Y.; Kusuhara, M.; Higashi, K.; Miyajima, E.; Mizuno, K.; Obata, T.; Ohmori, R.; Ohsuzu, F.; et al. Phytosterols dissolved in diacylglycerol oil reinforce the cholesterol lowering effect of low-dose pravastatin treatment. Nutr. Metabol. Cardiov. Dis. 2008, 20, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Ren, F.; Wei, H.; Liu, L.; Shen, T.; Xu, S.; Wei, J.; Ren, J.; Ni, H. Combination of berberine and evodiamine inhibits intestinal cholesterol absorption in high fat diet induced hyperlipidemic rats. Lip. Health Dis. 2017, 16, 239. [Google Scholar] [CrossRef] [PubMed]
- Sharavana, G.; Joseph, G.S.; Baskaran, V. Lutein attenuates oxidative stress markers and ameliorates glucose homeostasis through polyol pathway in heart and kidney of STZ-induced hyperglycemic rat model. Eur. J. Nutr. 2017, 56, 2475–2485. [Google Scholar] [CrossRef] [PubMed]
- Santhakumar, A.B.; Battino, M.; Alvarez-Suarez, J.M. Dietary polyphenols: Structures, bioavailability and protective effects against atherosclerosis. Food Chem. Toxicol. 2018, 113, 49–65. [Google Scholar] [CrossRef] [PubMed]
- U.S. Department of Agriculture. National nutrient database for standard reference. In National Nutrient Database for Nuts, Dry Roasted, Without Salt Added; USDA: Washington, DC, USA, 2016/2017. [Google Scholar]
- Alves, A.M.; Fernandes, D.C.; Borges, J.F.; Sousa, A.G.O.; Naves, M.M.V. Oleaginosas nativas do Cerrado apresentam perfis de ácidos graxos favoráveis à saúde cardiovascular. Rev. Nutr. 2016, 29, 859–866. [Google Scholar] [CrossRef]
- Siqueira, A.P.S.; Pacheco, M.T.B.; Naves, M.M.V. Nutritional quality and bioactive compounds of partially defatted baru almond flour. Food Sci. Technol. 2015, 35, 127–131. [Google Scholar] [CrossRef]
- Santiago, G.L.; Oliveira, I.G.; Horst, M.A.; Naves, M.M.V.; Silva, M.R. Peel and pulp of baru (Dipteryx Alata Vog.) provide high fiber, phenolic content and antioxidant capacity. Food Sci. Technol. 2016, 2, 244–249. [Google Scholar] [CrossRef]
- Bamberger, C.; Rossmeier, A.; Lechner, K.; Wu, L.; Waldmann, E.; Fischer, S.; Stark, R.G.; Altenhofer, J.; Henze, K.; Parhofer, K.G. A walnut-enriched diet affects gut microbiome in healthy caucasian subjects: A randomized, controlled trial. Nutrients 2018, 10, 244. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Nueno-Palop, C.; Bisignano, G.; Wickham, M.S.J.; Narbad, A. Potential prebiotic properties of almond (Amygdalus communis L.) seeds. Appl. Environ. Microbiol. 2008, 7, 4264–4270. [Google Scholar] [CrossRef] [PubMed]
- Ukhanova, M.; Wang, X.; Baer, D.; Novotny, J.A.; Fredborg, M.; Mai, V. Effects of almond and pistachio consumption on gut microbiota composition in a randomised cross-over human feeding study. Br. J. Nutr. 2014, 111, 2146–2152. [Google Scholar] [CrossRef] [PubMed]
- Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C. Interactions of gut microbiota with dietary polyphenols and consequences to human health. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Selma, M.V.; González-Sarrías, A.; Salas-Salvadó, J.; Andrés-Lacueva, C.; Alasalvar, C.; Örem, A.; Tomás-Barnerán, F.A.; Espín, J.C. The gut microbiota metabolism of pomegranate or walnut ellagitannins yields two urolithin-metabotypes taht correlate with cardiometabolic risk biomarkers: Comparison between normoweight, overweight-obesity and metabolic syndrome. Clin. Nutr. 2018, 37, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Lamuel-Raventos, R.M.; Onge, M.-P.S. Prebiotic nut compounds and human microbiota. Cric. Rev. Food Sci. Nutr. 2017, 57, 3154–3163. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, W.; Huang, G.; Zhanga, W.; Ni, L. In vitro and in vivo evaluation of the prebiotic effect of raw and roasted almonds (Prunus amygdalus). J. Sci. Food Agric. 2016, 96, 1836–1843. [Google Scholar] [CrossRef] [PubMed]
- Blaiotta, G.; Gatta, B.L.; Capua, M.D.; Luccia, A.D.; Coppola, R.; Aponte, M. Effect of chestnut extract and chestnut fiber on viability of potential probiotic Lactobacillus strains under gastrointestinal tract conditions. Food Microbiol. 2013, 36, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Byerley, L.O.; Samuelson, D.; Blanchard IV, E.; Luo, M.; Lorenzen, B.N.; Banks, S.; Ponder, M.A.; Welsh, D.A.; Taylor, C.M. Changes in the gut microbial communities following addition of walnuts to the diet. J. Nutr. Biochem. 2017, 48, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Lin, X.; Huang, G.; Zhang, W.; Rao, P.; Ni, L. Prebiotic effects of almonds and almond skins on intestinal microbiotain healthy adult humans. Anaerobe 2014, 26, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Holscher, H.D.; Taylor, A.M.; Swanson, K.S.; Novotny, J.A.; Baer, D.J. Almond consumption and processing affects the composition of the gastrointestinal microbiota of healthy adult men and women: A randomized controlled trial. Nutrients 2018, 10, 126. [Google Scholar] [CrossRef] [PubMed]
- Castaner, O.; Goday, A.; Park, Y.-M.; Lee, S.-H.; Magkos, F.; Shiow, S.-A.T.E.; Schröder, H. The gut microbiome profile in obesity: A systematic review. Inter. J. Endocrinol. Rev. 2018, 2018, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mokkala, K.; Pellonper, O.; Roytio, H.; Pussinen, P.; Ronnemaa, T.; Laitinen, K. Increased intestinal permeability, measured by serum zonulin, is associated with metabolic risk markers in overweight pregnant women. Metabolism 2017, 69, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Cheng, J.; Duncan, A.E.; Kau, A.L. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 2013, 341, 1241–2149. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Kardum, N.; Glibetic, M. Polyphenols and their interactions with other dietary compounds: Implications for human health. Adv. Food Nutr. Res. 2018, 84, 103–144. [Google Scholar] [PubMed]
- Ueda, S.; Nomoto, R.; Yoshida, K.; Osawa, R. Comparison of three tannases cloned from closely related lactobacillus species: L. plantarum, L. paraplantarum, and L. pentosus. BMC Microbiol. 2014, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Reference | Sample | Study Design | Study Findings | |
|---|---|---|---|---|
| Nut/Edible Seed (Dose Daily, Time of Intervention) | Significant Result | p Value | ||
| Wu et al. 2010 [7] | 283 adults with metabolic syndrome | Walnut (30 g, 12 weeks) | ⇓ waist circumference | <0.05 |
| Foster et al. 2012 [8] | 123 overweight and obese adults | Almond (56 g, 6 months) | ⇓ body weight | 0.04 |
| Wien et al. 2013 [9] | 65 overweight and obese adults | Almond (84 g, 24 weeks) | ⇓ body weight | <0.05 |
| ⇓ waist circumference | ||||
| ⇓ body fat | ||||
| Somerset et al. 2013 [10] | 64 overweight and obese adults | Macadamia (50% E from fat as MUFA,~50 g, 10 weeks) | ⇓ waist circumference | <0.05 |
| Gulati et al. 2014 [11] | 60 adults with metabolic syndrome | Pistachio (20% E, ~50 g, 3 weeks) | ⇓ waist circumference | 0.02 |
| Dhillon et al. 2016 [12] | 86 overweight and obese adults | Almond (42 g, 12 weeks) | ⇓ body fat | 0.04 |
| Nutrients and Bioactive Compounds | Nuts | Edible Seeds | References | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Almond | Brazil Nut | Cashew | Hazelnut | Pecan | Pistachio | Walnut | Baru Almond | Peanut | |||
| Energy (kcal) | 246 | 284 | 244 | 275 | 302 | 242 | 278 | 232 | 250 | [28,35] | |
| Total protein (g) | 8.28 | 5.92 | 7.99 | 5.98 | 3.19 | 8.42 | 5.72 | 12.72 | 12.58 | [15,28] | |
| Lipid (g) | Total | 18.43 | 28.35 | 18.58 | 26.12 | 28.13 | 19.16 | 27.41 | 18.14 | 18.72 | [15,36] |
| Fat acids (g/100 g oil) | SFA | 9.09 | 25.35 | 21.12 | 9.11 | 8.35 | 14.24 | 11.76 | 15.47 | 19.37 | [15,36] |
| PUFA | 29.31 | 45.61 | 17.19 | 7.79 | 24.92 | 51.47 | 72.96 | 31.71 | 37.76 | [15,36] | |
| MUFA | 61.60 | 29.04 | 61.18 | 83.10 | 66.73 | 34.29 | 15.28 | 51.57 | 42.72 | [15,36] | |
| MUFA/SFA | 6.7 | 1.1 | 2.9 | 9.1 | 8.0 | 2.4 | 1.3 | 3.3 | 2.2 | [15,36] | |
| Dietary fiber (g) | Total | 5.31 | 2.85 | 1.28 | 4.00 | 4.00 | 4.38 | 3.02 | 3.91 | 2.21 | [28,35] |
| Soluble | - | - | - | - | - | - | - | 0.86 | 0.58 | [28] | |
| Insoluble | - | - | - | - | - | - | - | 3.05 | 1.63 | [28] | |
| α-TE (mg/100 g oil) | 25.0 | 4.3 | 1.3 | 33.1 | 3.7 | 7.3 | 5.5 | 11.6 | 11.6 | [16,37] | |
| Phytosterols (mg/100 g oil) | 218 | 193 | 199 | 110 | 196 | 559 | 124 | - | 173 | [17] | |
| Carotenoids (µg) | Total | ND | ND | ND | ND | ND | 2040 | ND | 5 | - | [16,37] |
| Lutein | ND | ND | ND | ND | ND | 1870 | ND | - | ND | [16] | |
| β-carotene | ND | ND | ND | ND | ND | 170 | ND | - | ND | [16] | |
| Phenolics (mg) | Total (range) | 102 (55–194) | 48 (43–57) | 58 (56–60) | 124 (43–184) | 546 (434–614) | 369 (209–613) | 691 (444–872) | 309 | 179 (139–235) | [16,38] |
| Flavonoids | 40 | 46 | 27 | 49 | 300 | 61 | 317 | - | 62 | [17] | |
| Tannin | 123 | 4 | 17 | 98 | 374 | 94 | 145 | 239 | - | [15,37] | |
| Reference | Aim | Intervention | Study Type | Study Design | Study Findings |
|---|---|---|---|---|---|
| Mandalari et al. 2008 [40] | To investigate the potential prebiotic effect of almonds in vitro by using mixed fecal bacterial cultures | Finely ground whole (FG) and defatted (DG) almonds | In vitro | In vitro gastric and duodenal digestion of the almond samples and fractions were subsequently used as substrates for the colonic model in which the composition and metabolic activity of gut bacteria populations were evaluated | Whole almond: Bifidobacteria ⇑ Eubacterium rectale and butyrate production ⇑ |
| Blaiotta et al. 2013 [46] | To evaluate chestnut components as probiotic carriers by examining the effect on the viability of selected lactic acid bacteria (LAB) | Chestnut extracts and chestnut fiber (LAB viability during 180 min) | In vitro | Simulated gastric (with pepsin) and bile (with pancreatin) juices were prepared and added to cultured LAB cells (12 strains) with chestnut fiber or chestnut extracts | Lactobacillus paracasei GG ⇑ Lactobacillus rhamnosus ⇑ Lactobacillus casei ⇑ Streptococcus macedonicus ⇓ Streptococcus thermophilus ⇓ |
| Liu et al. 2016 [45] | To compare the fermentation properties of raw and roasted almonds | Predigested raw and roasted almonds (0, 2.5, 5, 10, 15%) | In vitro | Hydrolyzed raw and roasted almonds under simulated gastric and duodenal digestion were added to cultured Lactobacillus acidophilus, Bifidobacterium breve, and Escherichia coli and incubated anaerobically at 37° for 48 h | Lactobacillus acidophilus ⇑ Bifidobacterium breve ⇑ Escherichia coli ⇓ |
| Liu et al. 2016 [45] | To test the prebiotic effect of raw and roasted almonds on faecal and caecal bacteria | Raw and roasted almonds (5 g/kg BW - 1 g/day for 4 weeks via intragastric | Animal model | Male specific-pathogen-free (SPF) Wistar rats (30) with 10-week-old. They were randomly divided into three groups (10 rats per group) according to feeding regime: control, raw almonds, and roasted almonds | Lactobacillus ssp. ⇑ Bifidobacterium ssp. ⇑ Enteroccocus ssp. ⇓ Escherichia coli ⇓ |
| Byerley et al. 2017 [47] | To investigate if walnuts modulate the gut microbiome and promote their health benefit | Walnuts (approximately 1.7 g/day for 6 or 10 weeks) | Animal model | Male Fischer 344 rats (20) in two groups: 1) control diet, 2) walnut diet, with 11% walnuts replacing protein (casein), fat (oil), and fiber (cellulose) of the control diet. Fecal samples were collected from descending colon at the sacrifice | Lactobacillus ⇑ Ruminococcaceae ⇑ Roseburia ⇑ Bacteroides ⇓ Anaerotruncus ⇓ Alphaproteobacteria ⇓ |
| Ukhanova et al. 2014 [41] | To evaluate if intake of nuts affects bacterial or fungal microbiota composition | Almond (0, 1.5 and 3 servings/day: 0, 42.5 or 85 g for 18 days each treatment) | Randomized, controlled, crossover trial | Healthy adults (n = 18) with three 18 d feeding periods separated by a washout period of 2 weeks. During the 3 treatment periods, the same-base typical low-fiber American diet was provided | Lactobacillus ⇒ Bifidobacteria ⇒ |
| Pistachio (0, 42.5 or 85g/day for 18 days each treatment) | Healthy adults (n = 16) with three 18 d feeding periods separated by a washout period of 2 weeks. The provided diet was the same for almond treatment | Butyrate-producing bacteria ⇑ Bifidobacteria ⇒ Lactobacillus ⇒ lactic acid bacteria ⇓ | |||
| Liu et al. 2014 [48] | To investigate the prebiotic effects of almond and almond skin intake in healthy humans | Roasted almonds (56 g/day) and almond skin (10 g/day) for 6 weeks | Randomized, controlled trial | Healthy adult volunteers (n = 48, 16 for each treatment) consumed almonds (56 g), almond skin (10 g), or commercial fructooligosaccharides (8 g) (as positive control) daily for 6 weeks. Diet was provided by the school canteen, which excluded peanuts or other nuts | Lactobacillus ssp. ⇑ Bifidobacterium ssp. ⇑ Escherichia coli ⇒ Clostridum perfringens ⇓ |
| Holscher et al. 2018 [49] | To assess the interrelationship of almond consumption and processing on the gastrointestinal microbiota(bacterial genera) | Whole almonds (WA); whole, roasted almonds; roasted, chopped almonds (CA), and almond butter (42 g)/day for 21 days each treatment) | Randomized, controlled, crossover trial | Healthy adults (n = 18), controlled-feeding, five periods of 3 weeks, crossover study with washouts between diet periods was conducted. Treatments included: (1) zero servings/day of almonds (control); (2) 1.5 servings (42 g)/day of whole almonds; (3) 1.5 servings/day of whole, roasted almonds; (4) 1.5 servings/day of roasted, chopped almonds; and (5) 1.5 servings/day of almond butter | CA: Lachnospira ⇑ Roseburia ⇑ Oscillospira ⇑ WA: Dialister ⇑ |
| Bamberger et al. 2018 [39] | To investigate the effect of walnut intake on the gut microbiome composition and microbial diversity | Walnut (43 g/day) for 8 weeks | Randomized, controlled, crossover trial | Healthy nonsmoking subjects (n = 135) older than 50 years (men and postmenopausal women) subjects were randomized to 2 different diet phases, walnut-enriched diet (43 g/day) or nut-free control diet, for 8 weeks (separated by a 4-week washout) | Butyrate-producing bacteria ⇑ Ruminococcaceae ⇑ Bifidobacteria ⇑ Clostridium sp. ⇓ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sugizaki, C.S.A.; Naves, M.M.V. Potential Prebiotic Properties of Nuts and Edible Seeds and Their Relationship to Obesity. Nutrients 2018, 10, 1645. https://doi.org/10.3390/nu10111645
Sugizaki CSA, Naves MMV. Potential Prebiotic Properties of Nuts and Edible Seeds and Their Relationship to Obesity. Nutrients. 2018; 10(11):1645. https://doi.org/10.3390/nu10111645
Chicago/Turabian StyleSugizaki, Clara S. A., and Maria Margareth V. Naves. 2018. "Potential Prebiotic Properties of Nuts and Edible Seeds and Their Relationship to Obesity" Nutrients 10, no. 11: 1645. https://doi.org/10.3390/nu10111645
APA StyleSugizaki, C. S. A., & Naves, M. M. V. (2018). Potential Prebiotic Properties of Nuts and Edible Seeds and Their Relationship to Obesity. Nutrients, 10(11), 1645. https://doi.org/10.3390/nu10111645