Impact of Genetic Variability on Physiological Responses to Caffeine in Humans: A Systematic Review


 ,
,  ,
, 


Abstract
1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Selection Criteria
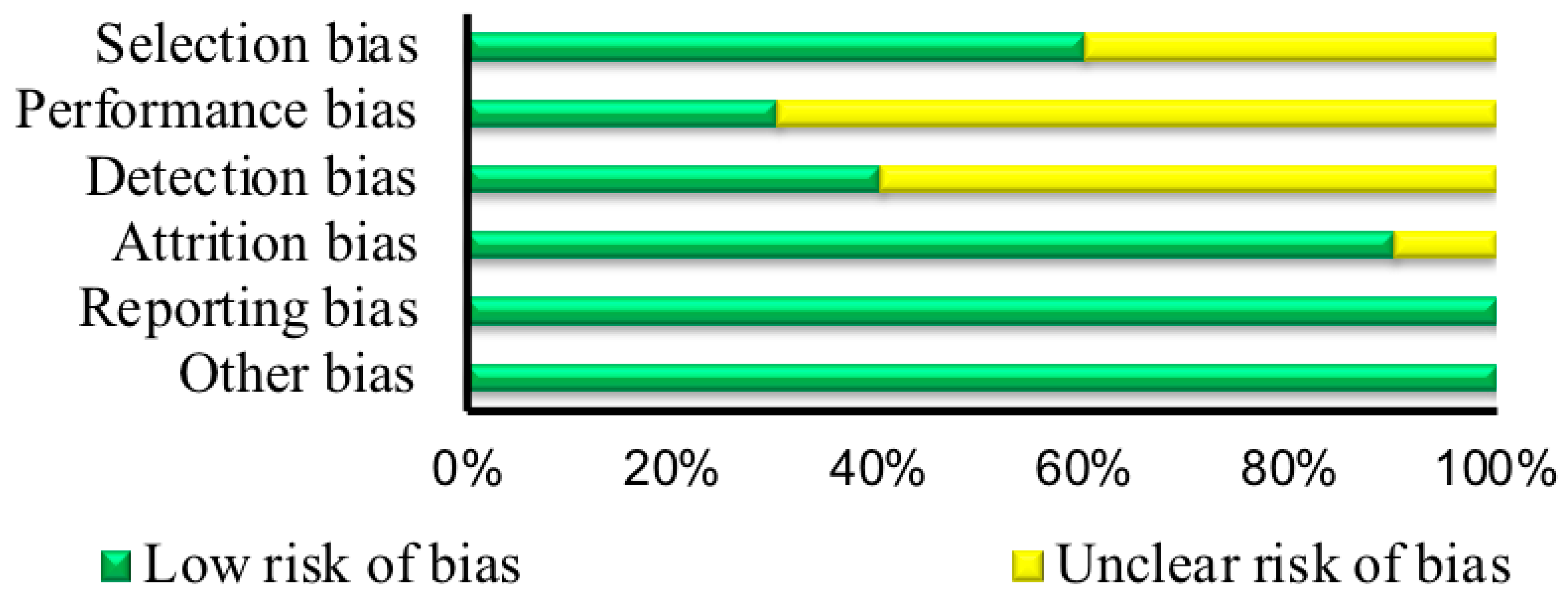
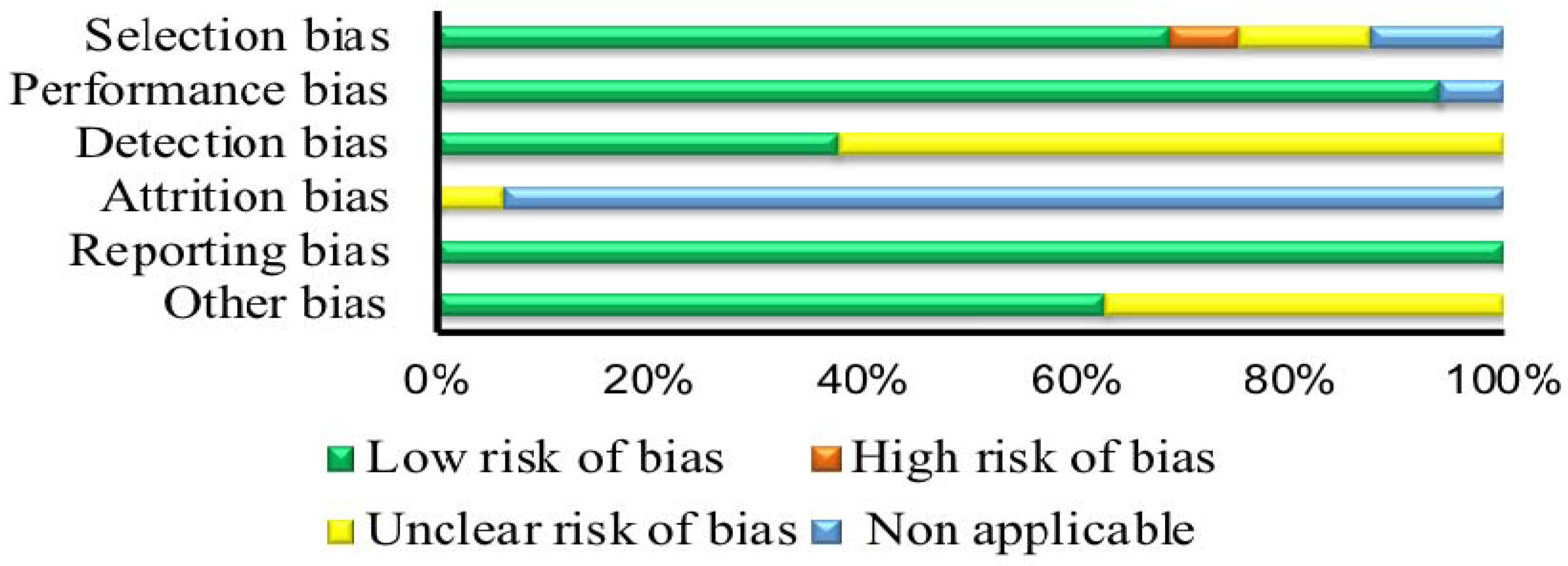
2.3. Risk of Bias Assessment
2.4. Data Extraction and Analysis
3. Results
3.1. Searching Procedure Results
3.2. Characteristics of the Included Studies
3.3. Risk of Bias Assessment
3.4. Reporting of the Outcomes
3.4.1. Habitual Use
3.4.2. Anxiogenic Consequences
3.4.3. Ergogenic Consequences
3.5. Other Outcomes
4. Discussion
4.1. Habitual Use
4.2. Anxiogenic Consequences
4.3. Ergogenic Consequences
4.4. Quality of Evidence, Limitations, and Potential Biases in the Review Process
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Magkos, F.; Kavouras, S.A. Caffeine use in sports, pharmacokinetics in man, and cellular mechanisms of action. Crit. Rev. Food Sci. Nutr. 2005, 45, 535–562. [Google Scholar] [CrossRef] [PubMed]
- Lelo, A.; Birkett, D.J.; Robson, R.A.; Miners, J.O. Comparative pharmacokinetics of caffeine and its primary demethylated metabolites paraxanthine, theobromine and theophylline in man. Br. J. Clin. Pharmacol. 1986, 22, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Sachse, C.; Brockmoller, J.; Bauer, S.; Roots, I. Functional significance of a C→A polymorphism in intron 1 of the cytochrome p450 cyp1a2 gene tested with caffeine. Br. J. Clin. Pharmacol. 1999, 47, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B. Astra award lecture. Adenosine, adenosine receptors and the actions of caffeine. Pharmacol. Toxicol. 1995, 76, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Salmi, P.; Chergui, K.; Fredholm, B.B. Adenosine-dopamine interactions revealed in knockout mice. J. Mol. Neurosci. 2005, 26, 239–244. [Google Scholar] [CrossRef]
- Childs, E.; Hohoff, C.; Deckert, J.; Xu, K.; Badner, J.; de Wit, H. Association between adora2a and drd2 polymorphisms and caffeine-induced anxiety. Neuropsychopharmacology 2008, 33, 2791–2800. [Google Scholar] [CrossRef] [PubMed]
- McMahon, G.; Taylor, A.E.; Davey Smith, G.; Munafò, M.R. Phenotype refinement strengthens the association of ahr and cyp1a1 genotype with caffeine consumption. PLoS ONE 2014, 9, e103448. [Google Scholar] [CrossRef] [PubMed]
- Eskelinen, M.H.; Kivipelto, M. Caffeine as a protective factor in dementia and alzheimer’s disease. J. Alzheimers Dis. 2010, 20, S167–S174. [Google Scholar] [CrossRef] [PubMed]
- Bohn, S.K.; Ward, N.C.; Hodgson, J.M.; Croft, K.D. Effects of tea and coffee on cardiovascular disease risk. Food Funct. 2012, 3, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, M.C.; Byrne, E.M.; Esko, T.; Nalls, M.A.; Ganna, A.; Paynter, N.; Monda, K.L.; Amin, N.; Fischer, K.; Renstrom, F.; et al. Genome-wide meta-analysis identifies six novel loci associated with habitual coffee consumption. Mol. Psychiatry 2015, 20, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Alsene, K.; Deckert, J.; Sand, P.; de Wit, H. Association between a2a receptor gene polymorphisms and caffeine-induced anxiety. Neuropsychopharmacology 2003, 28, 1694–1702. [Google Scholar] [CrossRef] [PubMed]
- Rogers, P.J.; Hohoff, C.; Heatherley, S.V.; Mullings, E.L.; Maxfield, P.J.; Evershed, R.P.; Deckert, J.; Nutt, D.J. Association of the anxiogenic and alerting effects of caffeine with adora2a and adora1 polymorphisms and habitual level of caffeine consumption. Neuropsychopharmacology 2010, 35, 1973–1983. [Google Scholar] [CrossRef] [PubMed]
- Graham, T.E. Caffeine and exercise: Metabolism, endurance and performance. Sports Med. 2001, 31, 785–807. [Google Scholar] [CrossRef] [PubMed]
- Ganio, M.S.; Klau, J.F.; Casa, D.J.; Armstrong, L.E.; Maresh, C.M. Effect of caffeine on sport-specific endurance performance: A systematic review. J. Strength Cond. Res. 2009, 23, 315–324. [Google Scholar] [CrossRef]
- Doherty, M.; Smith, P.M. Effects of caffeine ingestion on exercise testing: A meta-analysis. Int. J. Sport Nutr. Exerc. Metab. 2004, 14, 626–646. [Google Scholar] [CrossRef]
- Goldstein, E.R.; Ziegenfuss, T.; Kalman, D.; Kreider, R.; Campbell, B.; Wilborn, C.; Taylor, L.; Willoughby, D.; Stout, J.; Graves, B.S.; et al. International society of sports nutrition position stand: Caffeine and performance. J. Int. Soc. Sports Nutr. 2010, 7, 5. [Google Scholar] [CrossRef]
- Jenkins, N.T.; Trilk, J.L.; Singhal, A.; O’Connor, P.J.; Cureton, K.J. Ergogenic effects of low doses of caffeine on cycling performance. Int. J. Sport Nutr. Exerc. Metab. 2008, 18, 328–342. [Google Scholar] [CrossRef]
- Meyers, B.M.; Cafarelli, E. Caffeine increases time to fatigue by maintaining force and not by altering firing rates during submaximal isometric contractions. J. Appl. Physiol. 2005, 99, 1056–1063. [Google Scholar] [CrossRef]
- Doherty, M.; Smith, P.M.; Davison, R.C.; Hughes, M.G. Caffeine is ergogenic after supplementation of oral creatine monohydrate. Med. Sci. Sports Exerc. 2002, 34, 1785–1792. [Google Scholar] [CrossRef]
- Womack, C.J.; Saunders, M.J.; Bechtel, M.K.; Bolton, D.J.; Martin, M.; Luden, N.D.; Dunham, W.; Hancock, M. The influence of a cyp1a2 polymorphism on the ergogenic effects of caffeine. J. Int. Soc. Sports Nutr. 2012, 9, 7. [Google Scholar] [CrossRef]
- Loy, B.D.; O’Connor, P.J.; Lindheimer, J.B.; Covert, S.F. Caffeine is ergogenic for adenosine a2a receptor gene (adora2a) t allele homozygotes: A pilot study. J. Caffeine Res. 2015, 5, 2. [Google Scholar] [CrossRef]
- Harris, J.D.; Quatman, C.E.; Manring, M.M.; Siston, R.A.; Flanigan, D.C. How to write a systematic review. Am. J. Sports Med. 2014, 42, 2761–2768. [Google Scholar] [CrossRef]
- Davey, J.; Turner, R.M.; Clarke, M.J.; Higgins, J.P. Characteristics of meta-analyses and their component studies in the cochrane database of systematic reviews: A cross-sectional, descriptive analysis. BMC Med. Res. Methodol. 2011, 11, 160. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, M.; Berkman, N.D.; Dryden, D.M.; Hartling, L. Assessing Risk of Bias and Confounding in Observational Studies of Interventions or Exposure: Further Development of the RTI Item Bank. Available online: http://www.effectivehealthcare.ahrq.gov/reports/final.cfm (accessed on 22 August 2018).
- Algrain, H.A.; Thomas, R.M.; Carrillo, A.E.; Ryan, E.J.; Kim, C.-H.; Lettanll, R.B.; Ryan, E.J. The effects of a polymorphism in the cytochrome p450 cyp1a2 gene on performance enhancement with caffeine in recreational cyclists. J. Caffeine Res. 2015, 6, 1. [Google Scholar] [CrossRef]
- Cornelis, M.C.; El-Sohemy, A.; Campos, H. Genetic polymorphism of the adenosine a2a receptor is associated with habitual caffeine consumption. Am. J. Clin. Nutr. 2007, 86, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, M.C.; Monda, K.L.; Yu, K.; Paynter, N.; Azzato, E.M.; Bennett, S.N.; Berndt, S.I.; Boerwinkle, E.; Chanock, S.; Chatterjee, N.; et al. Genome-wide meta-analysis identifies regions on 7p21 (ahr) and 15q24 (cyp1a2) as determinants of habitual caffeine consumption. PLoS Genet. 2011, 7, e1002033. [Google Scholar] [CrossRef]
- Djordjevic, N.; Ghotbi, R.; Bertilsson, L.; Jankovic, S.; Aklillu, E. Induction of cyp1a2 by heavy coffee consumption in serbs and swedes. Eur. J. Clin. Pharmacol. 2008, 64, 381–385. [Google Scholar] [CrossRef]
- Domschke, K.; Gajewska, A.; Winter, B.; Herrmann, M.J.; Warrings, B.; Muhlberger, A.; Wosnitza, K.; Glotzbach, E.; Conzelmann, A.; Dlugos, A.; et al. Adora2a gene variation, caffeine, and emotional processing: A multi-level interaction on startle reflex. Neuropsychopharmacology 2012, 37, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Domschke, K.; Klauke, B.; Winter, B.; Gajewska, A.; Herrmann, M.J.; Warrings, B.; Muhlberger, A.; Wosnitza, K.; Dlugos, A.; Naunin, S.; et al. Modification of caffeine effects on the affect-modulated startle by neuropeptide s receptor gene variation. Psychopharmacology 2012, 222, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Gajewska, A.; Blumenthal, T.D.; Winter, B.; Herrmann, M.J.; Conzelmann, A.; Muhlberger, A.; Warrings, B.; Jacob, C.; Arolt, V.; Reif, A.; et al. Effects of adora2a gene variation and caffeine on prepulse inhibition: A multi-level risk model of anxiety. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 40, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Giersch, G.E.; Boyett, J.C.; Hargens, T.A.; Luden, N.D.; Saunders, L.J.; Daley, H.M.; Hughey, C.A.; El-Sohemy, A.; Womack, C.J. The effect of the cyp1a2 163 C>A polymorphism on caffeine metabolism and subsequent cycling performance. J. Caffeine Adenosine Res. 2018, 8, 2. [Google Scholar] [CrossRef]
- Guest, N.; Corey, P.; Vescovi, J.; El-Sohemy, A. Caffeine, cyp1a2 genotype, and endurance performance in athletes. Med. Sci. Sports Exerc. 2018, 50, 1570–1578. [Google Scholar] [CrossRef] [PubMed]
- Josse, A.R.; Da Costa, L.A.; Campos, H.; El-Sohemy, A. Associations between polymorphisms in the ahr and cyp1a1-cyp1a2 gene regions and habitual caffeine consumption. Am. J. Clin. Nutr. 2012, 96, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Luciano, M.; Zhu, G.; Kirk, K.M.; Gordon, S.D.; Heath, A.C.; Montgomery, G.W.; Martin, N.G. “No thanks, it keeps me awake”: The genetics of coffee-attributed sleep disturbance. Sleep 2007, 30, 1378–1386. [Google Scholar] [CrossRef]
- Pataky, M.W.; Womack, C.J.; Saunders, M.J.; Goffe, J.L.; D’Lugos, A.C.; El-Sohemy, A.; Luden, N.D. Caffeine and 3-km cycling performance: Effects of mouth rinsing, genotype, and time of day. Scand. J. Med. Sci. Sports 2016, 26, 613–619. [Google Scholar] [CrossRef]
- Pirastu, N.; Kooyman, M.; Robino, A.; van der Spek, A.; Navarini, L.; Amin, N.; Karssen, L.C.; Van Duijn, C.M.; Gasparini, P. Non-additive genome-wide association scan reveals a new gene associated with habitual coffee consumption. Sci. Rep. 2016, 6, 31590. [Google Scholar] [CrossRef]
- Puente, C.; Abián-Vicén, J.; Del Coso, J.; Lara, B.; Salinero, J.J. The cyp1a2 -163C>A polymorphism does not alter the effects of caffeine on basketball performance. PLoS ONE 2018, 13, e0195943. [Google Scholar] [CrossRef] [PubMed]
- Salinero, J.J.; Lara, B.; Ruiz-Vicente, D.; Areces, F.; Puente-Torres, C.; Gallo-Salazar, C.; Pascual, T.; Del Coso, J. Cyp1a2 genotype variations do not modify the benefits and drawbacks of caffeine during exercise: A pilot study. Nutrients 2017, 9, 269. [Google Scholar] [CrossRef]
- Soares, R.N.; Schneider, A.; Valle, S.C.; Schenkel, P.C. The influence of cyp1a2 genotype in the blood pressure response to caffeine ingestion is affected by physical activity status and caffeine consumption level. Vascul. Pharmacol. 2018, 106, 67–73. [Google Scholar] [CrossRef]
- Thomas, R.M.; Algrain, H.A.; Ryan, E.J.; Popojas, A.; Carrigan, P.; Abdulrahman, A.; Carrillo, A.E. Influence of a cyp1a2 polymorphism on post-exercise heart rate variability in response to caffeine intake: A double-blind, placebo-controlled trial. Ir. J. Med. Sci. 2017, 186, 285–291. [Google Scholar] [CrossRef]
- Urry, E.; Jetter, A.; Landolt, H.P. Assessment of cyp1a2 enzyme activity in relation to type-2 diabetes and habitual caffeine intake. Nutr. Metab. 2016, 13, 66. [Google Scholar] [CrossRef] [PubMed]
- Margulis, A.V.; Pladevall, M.; Riera-Guardia, N.; Varas-Lorenzo, C.; Hazell, L.; Berkman, N.D.; Viswanathan, M.; Perez-Gutthann, S. Quality assessment of observational studies in a drug-safety systematic review, comparison of two tools: The Newcastle-Ottawa scale and the RTI item bank. Clin. Epidemiol. 2014, 6, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Al-Saleh, M.A.; Armijo-Olivo, S.; Thie, N.; Seikaly, H.; Boulanger, P.; Wolfaardt, J.; Major, P. Morphologic and functional changes in the temporomandibular joint and stomatognathic system after transmandibular surgery in oral and oropharyngeal cancers: Systematic review. J. Otolaryngol. Head Neck Surg. 2012, 41, 345–360. [Google Scholar] [PubMed]
- Higgins, J.P.; Altman, D.G.; Gotzsche, P.C.; Juni, P.; Moher, D.; Oxman, A.D.; Savovic, J.; Schulz, K.F.; Weeks, L.; Sterne, J.A.; et al. The cochrane collaboration’s tool for assessing risk of bias in randomised trials. BMJ 2011, 343, d5928. [Google Scholar] [CrossRef] [PubMed]
- Puente, C.; Abian-Vicen, J.; Salinero, J.J.; Lara, B.; Areces, F.; Del Coso, J. Caffeine improves basketball performance in experienced basketball players. Nutrients 2017, 9, 1033. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.; Green, S. Cochrane Handbook for Systematic Reviews of Interventions; Version 5.1.0; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011. [Google Scholar]



{kind=link}
{kind=link}
| First Author | Design | Participants | Main Outcome | Secondary Outcome |
|---|---|---|---|---|
| Algrain [25] | Controlled Trial | Male (M) = 13 Female (F) = 7 | Polymorphism in CYP1A2 gene (AA and C-allele carriers) did not impact ergogenic benefit of caffeine in recreational cyclists (p > 0.05) | |
| Alsene [11] | Randomized Controlled Trial | 94 healthy, infrequent caffeine users | 1976T/T and 2592Tins/Tins genotypes report greater increase in anxiety after caffeine administration (p < 0.05) | |
| Childs [6] | Randomized Controlled Trial | 102 healthy individuals (M = 51 and F = 51) who consumed less than 300 mg caffeine per week | ADORA2A TT genotype reported highest anxiety (VAS) (4.6 ± 1.9) and ADORA2A CC (−7.5 ± 3.7) reported the least anxiety after 150 mg of caffeine but was not significant when data for European-American participants were considered (p = 0.1); caffeine-induced anxiety was associated with dopamine receptor 2 gene (DRD2) polymorphism | |
| Cornelis [26] | Cross-sectional | n = 2735 | ADORA2A, but not CYP1A2, genotype was associated with different amounts of caffeine intake; compared to persons consuming <100 mg caffeine/day, odds ratios for having the ADORA2A TT genotype were 0.74, 0.63, and 0.57 for persons consuming 100–200, 200–400, and >400 mg caffeine/day, respectively | Association more pronounced among current smokers compared to nonsmokers |
| Cornelis [27] | Cross-sectional | 47,341 individuals of European descent | Two loci-7p21 (p = 2.4 × 10−19), near AHR, and 15q24 (p = 5.2 × 10−14), near CYP1A1 and CYP1A2; both candidates as CYP1A2 caffeine metabolizers | |
| Cornelis [10] | Cross-sectional | Coffee consumers of European ancestry n = 91,462 African American ancestry n = 7964 | Eight loci, six being novel, met genome-wide significance (log10Bayes factor >5.64); loci near genes potentially involved in pharmacokinetics (ABCG2, AHR, POR, and CYP1A2) and pharmacodynamics (BDNF and SLC6A4). Loci related to metabolic traits (GCKR and MLXIPL) | |
| Djordjevic [28] | Single-group interventional design | 126 Healthy Serbians, 64 nonsmoking (from their previous study) | Inducing effect of CYP1A2 activity with heavy coffee consumption among Serbian (p = 0.022) and Swedish (p = 0.016) participants carrying the CYP1A2-163 C > A polymorphism | |
| Domschke [29] | Controlled Trial | M = 56 and F = 54 healthy individuals | Startle magnitude highest for unpleasant pictures and lowest for pleasant pictures across ADORA2A genotypes (p < 0.001). TT (risk) genotype carriers had highest startle magnitude in the caffeine condition in response to unpleasant pictures, occurring mostly among females | Females of this group had higher startle magnitudes than males |
| Domschke [30] | Controlled Trial | 58 M and 66 F healthy proband | ADORA2A TT risk genotype carriers had significantly increased startle magnitude in response to neutral stimuli (p = 0.02) and a significant decrease in startle magnitude in response to unpleasant stimuli (p = 0.02) in caffeine compared to placebo condition; no change in AA/AT nonrisk genotype | |
| Gajewska [31] | Randomized Controlled Trial | 57 M and 57 F healthy individuals controlled for anxiety sensitivity | Prepulse inhibition was influenced by genetics (ADORA2A 1976C/T); impaired prepulse facilitation in anxiety sensitive ADORA2A TT group in response to caffeine compared to placebo (t(56) =2.16, p = 0.04) | |
| Giersch [32] | Controlled Trial | 20 male subjects between age of 18–45 years | CYP1A2 C-allele carriers had higher serum caffeine one hour after caffeine ingestion (C-allele carriers = 14.2 ± 1.8 ppm, AA homozygotes = 11.7 ± 1.7 ppm, p = 0.001). No difference between genotypes in caffeine metabolites (p > 0.05). Main effect of caffeine on performance (p = 0.03); no caffeine by genotype interaction (p > 0.05) | |
| Guest [33] | Randomized Controlled Trial | In CYP1A2 AA genotype, cycling time decreased by 4.8% (p = 0.0005) and 6.8% (p < 0.0001) with 2 and 4 mg kg−1 caffeine consumption, respectively. (2 and 4 mg kg−1); in CC genotype, cycling time increased by 13.7% (p = 0.04) with caffeine consumption (4 mg kg−1), no effects were observed among AC genotype | Competitive male athletes n = 101 | 4 mg kg−1 caffeine decreased cycling time by 3% vs. placebo |
| Josse [34] | Cross-sectional | n = 1639 nonsmokers and n = 884 current smokers | Subjects who consumed >400 mg caffeine compared to who consumed <100 mg caffeine were more likely to be carriers of T, C, or T alleles for rs6968865, rs4410790, and rs2472297, respectively; corresponding Odds Ratios and 95% confidence intervals (CIs) were 1.41 (1.03, 1.93), 1.41 (1.04, 1.92), and 1.55 (1.01, 2.36) | |
| Loy [21] | Randomized Controlled Trial | Women with high self-reported caffeine sensitivity and low daily caffeine consumption, TT n = 6, CT/CC n = 6 | Caffeine proved to be ergogenic for ADORA2A TT allele homozygotes (6.85 ± 4.41 kJ) but not ergogenic for CT/CC alleles (−2.70 ± 5.64 kJ) (d = −1.89) | |
| Luciano [35] | Cross-sectional | 3808 Australian adult twin pairs (n = 1799 monozygous pairs and n = 2009 dizygous pairs) | Genes not typically associated with sleep disturbance were implicated in coffee-attributed insomnia | |
| McMahon [7] | Cross-sectional | 4460–7520 women | Caffeine consumption was associated with CYP1A1 (Betas = 8.7 to 21.4, p-values = 1.59 × 10−3 to 3.33 × 10−10) and AHR (Betas = 4.0 to 14.6, p-values = 1.1510−1 to 3.34 × 10−6) genotypes; association not strengthened with combined allelic score (1.28% of phenotypic variance) | |
| Pataky [36] | Randomized Controlled Trial | 25 M and 13 F recreational cyclists from James Madison University | CYP1A2 AC heterozygotes experienced greater power output (6%) with caffeine ingestion. Caffeine ingestion favored AC heterozygotes compared to AA homozygotes when performance gains were compared to placebo (5.1 ± 6.1%, p = 0.12) | |
| Pirastu [37] | Cross-sectional | 370 individuals from Puglia, Italy and 843 individuals from Friuli Venezia Region, Italy | PDSS2 gene shown in sample was linked to negative regulation of the expression of caffeine metabolism genes in several tissues (e.g., subcutaneous adipose tissue −0.27, skeletal muscle −0.52) | |
| Puente [38] | Randomized Controlled Trial | 10 men and 9 women elite basketball players | CYP1A2 genotype (rs762551) AA improved Abalakov jump height (p = 0.03) with caffeine consumption, while C-allele carriers remained unchanged (p = 0.33); Sprint was not improved in either genotype with caffeine, while number of body impacts increased in both AA (4.1 ± 5.3%; p = 0.02) and C-allele carriers (3.3 ± 3.2%; p = 0.01) | |
| Rogers [12] | Randomized Controlled Trial | 162 non/low and 217 medium/high caffeine consumers | ADORA2A (rs5751876) TT genotype showed largest increase in anxiety after caffeine (mean ± standard error ) for caffeine = 1.65 ± 0.15 and for placebo = 0.95 ± 0.17, p < 0.01) | |
| Sachse [3] | Single-group interventional design | 185 healthy Caucasian nonsmokers and 51 smokers | Among smokers (n = 51), subjects who possessed the CYP1A2 (rs762551) AA genotype metabolized caffeine (100 mg) faster 1.37 (AC –0.88; and CC – 0.82) relative to C-allele carriers while utilizing a 5 h paraxanthine/caffeine ratio as the outcome measure (p = 0.008) | |
| Salinero [39] | Randomized Controlled Trial | 21 healthy active participants | Caffeine ingestion increased peak and mean power in both AA and C-allele carriers of the CYP1A2 gene (p > 0.05); no difference in Wingate test performance between AA and C-allele carriers (p > 0.05) | 31.3% of C-allele carriers reported increased nervousness after caffeine ingestion |
| Soares [40] | Single-group interventional design | 37 individuals between ages of 19–50 | Systolic blood pressure (BP) increased with caffeine ingestion only among individuals with CYP1A2 AA genotype (p < 0.05); both CYP1A2 AA and AC had high diastolic BP after caffeine ingestion (p < 0.05); physical activity only modulated the BP responses to acute caffeine ingestion in AC individuals | |
| Thomas [41] | Controlled Trial | CYP1A2 AA (F = 4 and M = 7), C-allele carriers (F= 3 and M = 6) | No difference in heart rate variability between CYP1A2*1F polymorphisms (i.e., AA and C-allele carriers) measured at baseline and postexercise (p > 0.05) | |
| Urry [42] | Case–Control Study | 57 subjects with type 2 diabetes (T2D) and 146 non-T2D | CYP1A2 enzyme activity was significantly higher in T2D compared to control group (p = 0.004) | |
| Womack [20] | Randomized Controlled Trial | Trained male cyclists n = 35 | Caffeine supplementation reduced 40 km time greater in CYP1A2 AA homozygotes (4.9%) than in C-allele carriers (1.8%) (p < 0.05) |
| First Author | Random Sequence Generation | Allocation Concealment | Blinding of Participants and Researchers | Blinding of Outcome Assessment | Incomplete Outcome Data | Selective Reporting | Other Bias |
|---|---|---|---|---|---|---|---|
| Randomized Controlled Trials (RCTs) | |||||||
| Gajewska [31] | ? | ? | ? | ? | + | + | + |
| Alsene [11] | ? | ? | ? | + | + | + | + |
| Pataky [36] | ? | ? | ? | + | + | + | + |
| Rogers [12] | ? | ? | ? | ? | + | + | + |
| Puente [38] | ? | + | + | ? | + | + | + |
| Guest [33] | + | + | ? | + | + | + | + |
| Salinero [39] | ? | + | + | ? | + | + | + |
| Loy [21] | + | ? | + | ? | ? | + | + |
| Womack [20] | ? | + | ? | + | + | + | + |
| Childs [6] | ? | ? | ? | ? | + | + | + |
| First Author | Selection Bias | Performance Bias | Detection Bias | Attrition Bias | Selective Outcome | Confounding |
|---|---|---|---|---|---|---|
| Non-RCT | ||||||
| Djordjevic [28] | - | + | ? | o | + | ? |
| McMahon [7] | o | o | ? | ? | + | + |
| Soares [40] | + | + | ? | o | + | + |
| Giersch [32] | + | + | + | o | + | + |
| Domschke [30] | + | + | + | o | + | + |
| Sachse [3] | + | + | ? | o | + | + |
| Urry [42] | ? | + | ? | o | + | + |
| Pirastu [37] | ? | + | + | o | + | ? |
| Josse [34] | + | + | ? | o | + | ? |
| Cornelis [27] | + | + | ? | o | + | ? |
| Domschke [29] | + | + | + | o | + | + |
| Cornelis [10] | + | + | ? | o | + | ? |
| Thomas [41] | + | + | + | o | + | + |
| Algrain [25] | + | + | + | o | + | + |
| Cornelis [26] | + | + | ? | o | + | + |
| Luciano [35] | o | + | ? | o | + | ? |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fulton, J.L.; Dinas, P.C.; Carrillo, A.E.; Edsall, J.R.; Ryan, E.J.; Ryan, E.J. Impact of Genetic Variability on Physiological Responses to Caffeine in Humans: A Systematic Review. Nutrients 2018, 10, 1373. https://doi.org/10.3390/nu10101373
Fulton JL, Dinas PC, Carrillo AE, Edsall JR, Ryan EJ, Ryan EJ. Impact of Genetic Variability on Physiological Responses to Caffeine in Humans: A Systematic Review. Nutrients. 2018; 10(10):1373. https://doi.org/10.3390/nu10101373
Chicago/Turabian StyleFulton, Jacob L., Petros C. Dinas, Andres E. Carrillo, Jason R. Edsall, Emily J. Ryan, and Edward J. Ryan. 2018. "Impact of Genetic Variability on Physiological Responses to Caffeine in Humans: A Systematic Review" Nutrients 10, no. 10: 1373. https://doi.org/10.3390/nu10101373
APA StyleFulton, J. L., Dinas, P. C., Carrillo, A. E., Edsall, J. R., Ryan, E. J., & Ryan, E. J. (2018). Impact of Genetic Variability on Physiological Responses to Caffeine in Humans: A Systematic Review. Nutrients, 10(10), 1373. https://doi.org/10.3390/nu10101373