1. Introduction
Mountain areas, due to their serious denivelations and difficult access, often consist of fragments of old-growth forests, so they are valuable, especially due to their biodiversity and genetic resources [
1]. The elevations of these areas have led to the development of vegetation belts with high diversity and spatial heterogeneity. Mountain forests also provide an important refuge for endemic and stenotopic species. Hoffmann et al. [
2] emphasized that climate changes will be detrimental for forests in central Europe, mainly due to the substantial increase in air temperatures as well as the insufficient amount of precipitation to counteract the resulting water stress in trees, reducing the abilities of trees to protect themselves against pathogens, increasing the risk of pest damage and diseases [
3]. These changes also affect the albedo of vegetation cover, which can lead to further climate alterations through positive feedback [
4]. These alterations may affect mountain forests, causing critical disturbances and irreversible changes [
5], as nearly 60% of this mountainous area is under intense human pressure [
6]. The dominant species in the forests of central Europe is Norway spruce (
Picea abies), which is preferred in forest management due to its fast growth and high productivity [
5,
6]. Bark beetle outbreaks, which result in the dynamic dieback of mature spruce trees, have led to the intensive development of new species below the plant canopy and, as a result, the appearance of deciduous tree species [
7]. In our previous study [
8], we confirmed that in 2015–2018, as a result of unfavorable phenomena that occurred in the Tatra Mountains, e.g., windthrow, bark beetle outbreaks, followed by sanitary cuts, approximately 29% (62 km
2) of Polish and Slovak coniferous tree stands died. The dynamics of these phenomena depends on many factors, such as habitats and vegetation belt (topography, topoclimate, and soils), as well as on the species composition of trees and their genotypes [
9]. The same issues have been observed in many other places [
10] and are expected to globally intensify [
11]. However, the impacts of individual disturbances on forest ecosystems are diverse, depending on the scale and frequency [
12]. Sometimes, these disturbances provide the opportunity to restore the ecosystem because, in many cases, the prior plantings were inconsistent with the habitat; after ecological disasters, the soil was protected from erosion by planting often genetically inappropriate available seedlings [
13]. For example, spruce should not be planted on rich soil in lower vegetation belts, where European beech (
Fagus sylvatica) and silver fir (
Abies alba) should dominate [
8]. Additionally, intensive disturbances covering large areas may lead to biotic homogenization [
14], limiting the ability of ecosystems to counteract or recover from negative phenomena, e.g., humus erosion, leaching of valuable minerals, and dieback of the rhizosphere [
15]. An additional threat is air pollution, which causes leaf chlorosis and weakened vegetation at the beginning of the growing season [
16].
The key element in environmental monitoring is the assessment of the changes that are occurring [
17]. In many countries, environmental monitoring is also required by law to regularly prepare national park management plans considering the dynamics and directions of these changes. The traditional monitoring methods are based on field verification of designated circular areas, and the obtained results were extrapolated to the remaining area being analyzed. However, current high-resolution multispectral images allow detailed identifications of individual tree species, the assessment of their condition, and observations for entire research areas with the same accuracy, producing average median F1-scores of 0.67–0.92 depending on the species [
18]. The current research problem is the assessment of classification algorithms, which, due to the high heterogeneity of the analyzed objects, require a diverse approach to identify selected species. The spatial and spectral resolutions of modern satellite sensors allow the registration of tree crowns on at least one pixel, which enables the identification of spectrally pure pixels and considerably facilitates the process of classification and assessment of individual species, even in mixed forests, achieving F1-scores of 0.76–0.90 depending on the data set [
19]. However, a dozen spectral bands may be insufficient to capture the spectral properties of individual species; therefore, an appropriate solution is using multitemporal data that reflect the characteristic features of individual species during vegetative development, e.g., leaf growth and discoloration [
19,
20,
21].
National parks protect old-growth forests (OGFs), which, due to their age, constitute valuable genetic resources [
22]. Spracklen and Spracklen [
22], based on the 10 and 20 m bands of Sentinel-2 and a random forest classifier, identified the dominant tree species of the eastern part of the Carpathians mountain range, which is where we have conducted our studies, in the western part of the range. For Norway spruce and beech, they achieved 95–98% producer accuracy (PA) and 85–90% user accuracy (UA), but for the rest of the other 17 species, the PA ranged between 25% and 60% [
22]. To differentiate the age of these species, the authors used a set of six indices and the grey level co-occurrence matrix. The outcomes allowed the identification of OGFs with an accuracy of approximately 85% for multitemporal Sentinel-2 images (summer and autumn). The random forest classifier and Sentinel-2 images were successfully used to identify tree species at the individual-tree scale of mixed forest in the Veluwe region (The Netherlands) [
23]. Based on the vegetation structure and tree composition, the authors identified 479 plots representing 1743 trees in the field. Sentinel-2 image acquisition focused only on the 10 m pixel size bands representing different phenological periods. In the first step, the tree tops were identified from the canopy height model (CHM); the same was applied to delineate tree crowns as an object-based approach according to the Popescu and Wynne method [
24] using the R-package
ForestTools [
25]. This allowed the extraction of structural features of the crown [
23]. The random-forest-based classification process consisted of a few scenarios, which focused on structural crown variables, one season, and multitemporal spectral properties. As the results of species classification, an accuracy of 78% was achieved based on all variables, and 73% was attained using only the spectral features of the multitemporal Sentinel-2 analyses. With a single image, the most accurate results were obtained in summer (71%) and the worst in winter (63%) [
23]. Bolyn et al. [
26] used Sentinel-2 images acquired from 13 dates (between spring and summer) for classification using a U-shaped neural network (UNet++) [
27]. The authors used 10 bands (10 and 20 m) from the time-weighted Sentinel-2 images [
28] to identify nine tree species in Wallonia (Belgium). The convolutional neural networks achieved a 73% overall accuracy (OA); the most accurate results were obtained for the tree species spruce, oak, beech, and Douglas fir, for which the PA and UA scores were above 70%. For poplars, larches, and birches, the PA was below 50%. These results are comparable to those produced by machine learning classifiers. However, the classification accuracy depends on the naturally occurring geographic zones and the heterogeneity of habitats. For example, Zagajewski et al. [
18], based on Sentinel-2 images, obtained a 74% UA, 67% PA, and 0.70 F1-score for SVM in the case of larch; slightly lower results were achieved by the random forest and artificial neural network classifiers. The same observation was obtained by Punalekar et al. [
29], who implemented a nonparametric extra tree classifier for Sentinel-2 images and nine vegetation indices for classifying larch in the whole of Wales with high accuracy: the F1-score was greater than 0.97.
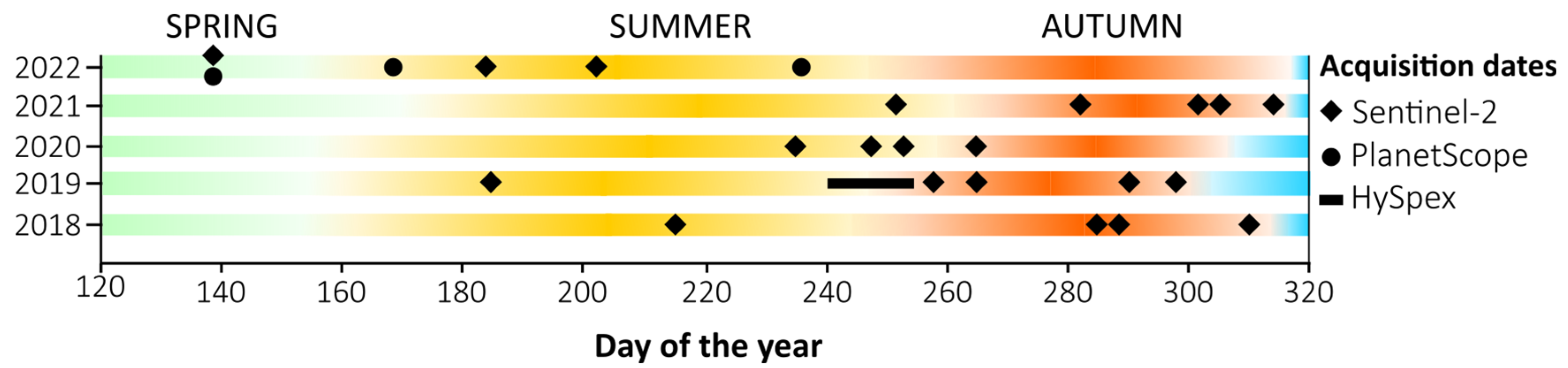
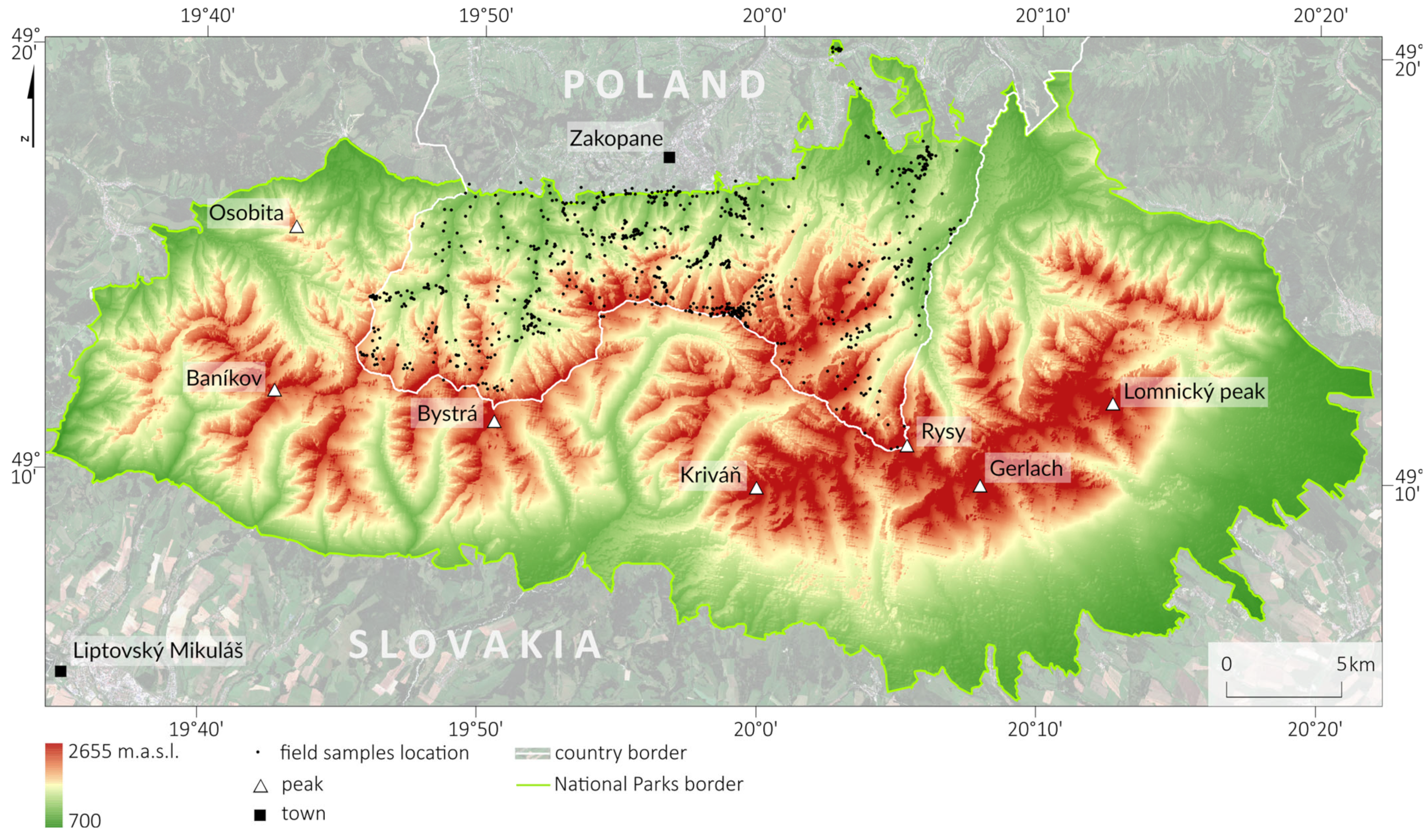
The high-mountain areas of central Europe are characterized by dynamic weather due to the influence of the Atlantic and continental climates, which result in the lifting of warm air masses, where condensation and cloudiness occur, which cause windbreaks, changing the age and species structure of forest stands. Therefore, obtaining scenes representing various stages of vegetation development is challenging. Hence, we need to analyze multiseasonal series to help capture phenology. Additionally, because the growing season in the mountains is short and the spectral properties of vegetation quickly change, we used multitemporal Sentinel-2 compositions. As the reference data, we collected field-verified polygons, representing pixels of all analyzed species located in different topographic patterns with different elevations, slopes, and aspects. In different topographic locations, the same species has different spectral signatures as a result of the influences of water vapor, insolation, wind, and temperature. These factors play key roles in leaf drying, which forces the plant to adopt protective mechanisms that affect spectral responses. Ecosystem heterogeneity (plant communities, rocks, dry trunks, branches, and other objects) generates mixels through the effect of the neighborhood, which influences the spectral responses registered by airborne and satellite sensors and thereby the classification results [
30]. The HySpex reference data have a spatial resolution of 2 m and a spectral resolution of 430 bands, which seems to be an advantage; however, the pixel size is comparable to the size of a tree crown. Additionally, the availability of long-term Sentinel-2 data substantially improves the ability to identify tree stands, especially when individual species implement different growth and development strategies during the vegetative period. Capturing spectral variation during the phenological season can remarkably increase the accuracy of the classification results and indicates which data selection scenarios allow us to obtain satisfactory results.
The use of data and open-source software allows for the constant monitoring of the environment without incurring burdensome costs, which is especially valuable for NGOs or nature conservation areas that have teams of several specialists dealing with nature monitoring, as well as well-qualified foresters who can verify the observed changes on an ongoing basis. Additionally, Sentinel-2 imaging covers the entire area at once, which reduces radiometric differences and ensures spectral coherence; the relatively short revisit time allows for the elimination of gaps created by single clouds. In comparison, airborne campaign planning is much more demanding due to size of the study area, elevation differences, and weather dynamics. As such, airborne campaigns often last for several days and include the acquisition of various fragments in different weather conditions, which may change the properties of the spectral analysis of the analyzed objects and thus increase the imaging cost. Satellite data allow the acquisition of objective and repeatable data for long-term environmental monitoring [
31], and the obtained results can be reliably applied to other mountain areas, which allows the identification of changes in alpine ecosystems located in different parts and zones in Europe and around the world.
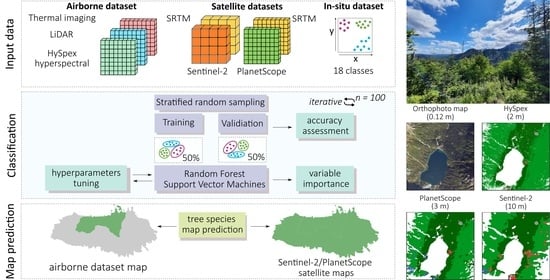
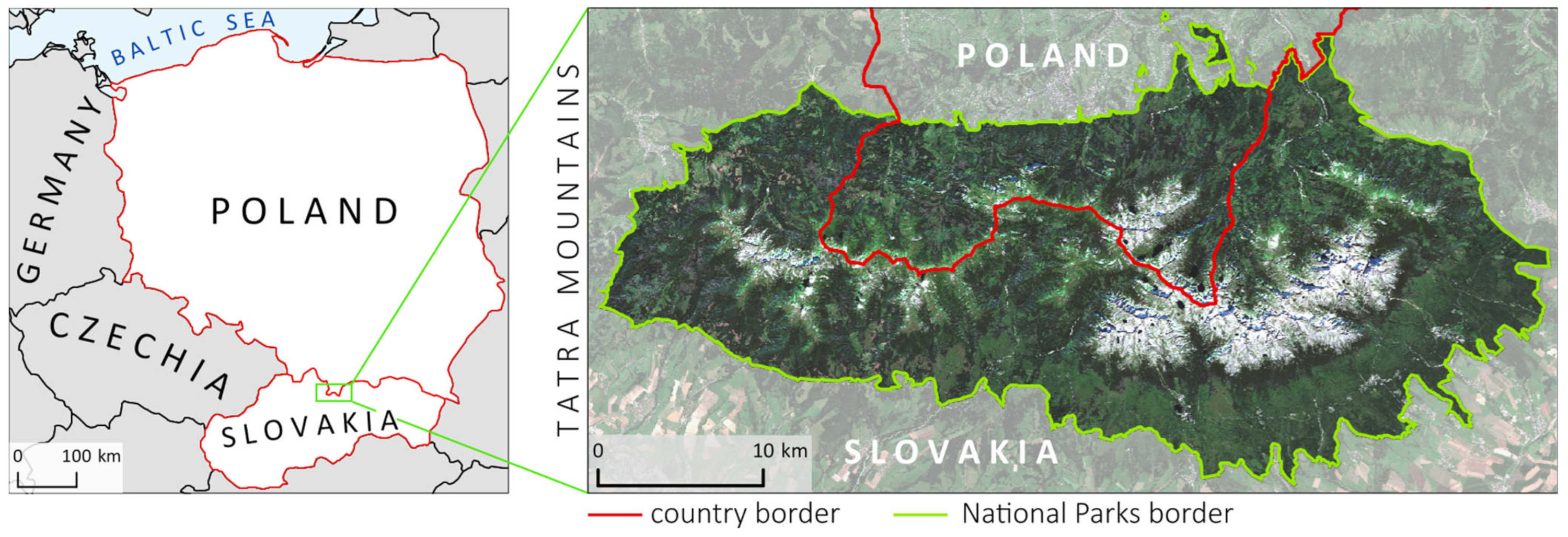
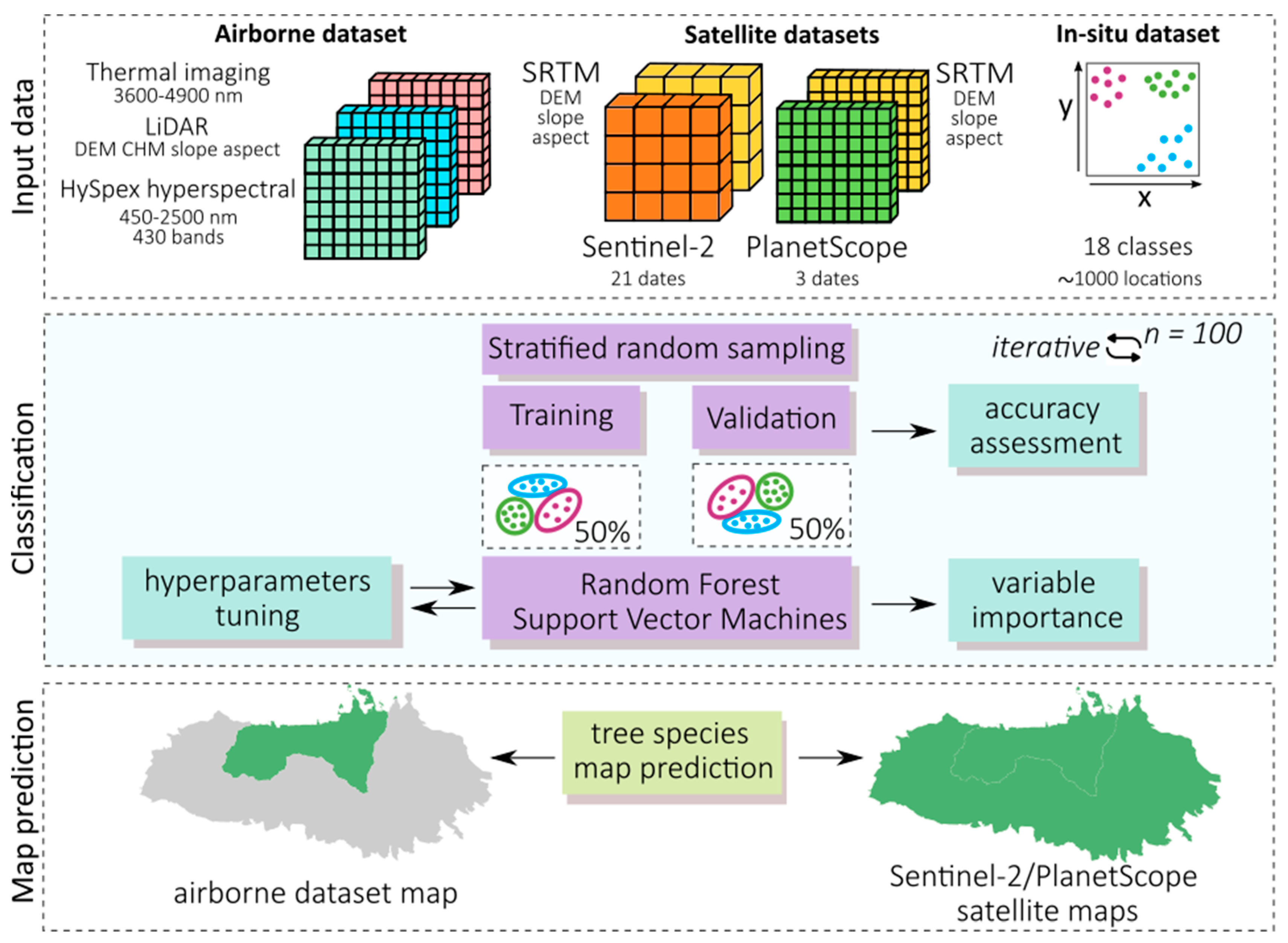
Our aim in this study was to assess the free Sentinel-2 data with commercial PlanetScope and airborne HySpex hyperspectral data for mapping the dominant woody species of the Tatra Mountains, which is a challenging research objects due to the substantial elevation differences in the area, as well as its various aspects and slopes, which create a mosaic of spectral features. Therefore, one of our objectives in this study was to assess the impact of topographic feature derivatives on the accuracy of the identification of coniferous and deciduous tree species. Because the greening-up period, maturity, senescence, and dormancy of individual species depend on elevation, slope, and aspect, the analyses for the same species depend on the topography. The Tatra forest ecosystem is characterized by high heterogeneity of species, which form different-sized groups of individual trees. Additionally, the variability in age, dry tree trunks, and branches causes a large mix of the signals recorded in individual pixels, which complicates the identification of the dominant objects; this directly affects the ability to identify individual species. Due to the occurrence of individual species in hard-to-reach places, e.g., Swiss stone pine (Pinus cembra, where single individuals are identified on mountain tops), species appear in all topographically analyzed patterns (elevation, slope, and aspect); therefore, an important issue in this study was testing different sizes of balanced samples to assess their impact on the classification result. For this reason, we applied an iterative classification method: we performed each classification iteration based on randomized selected patterns, which we independently selected for each iteration, and we repeated the whole process 100 times. This allowed us to test the spectral variability in the analyzed patterns and the impact of the number of training pixels on the obtained results. Therefore, the classification process was based on a large number of field-verified polygons to capture the diversity of forms of individual species occurring both on homogeneous plots and in the areas where several species coexisted. As classifiers, we used the R-based open-source programming libraries for the SVM and RF algorithms.
The main study question was to assess the classification accuracy of the dominant woody species in protected mountain areas based on airborne hyperspectral HySpex data as well as commercial high-resolution PlanetScope and open-access Sentinel-2 satellite data. We conducted the analyses to assess if investing in commercial imaging is profitable and if multitemporal Sentinel-2 images can be used to obtain sufficiently accurate spectral characteristics during the vegetative period to enable the successful identification of woody species of natural forests, which are much more heterogeneous in terms of tree composition and age compared with managed forests.
3. Results
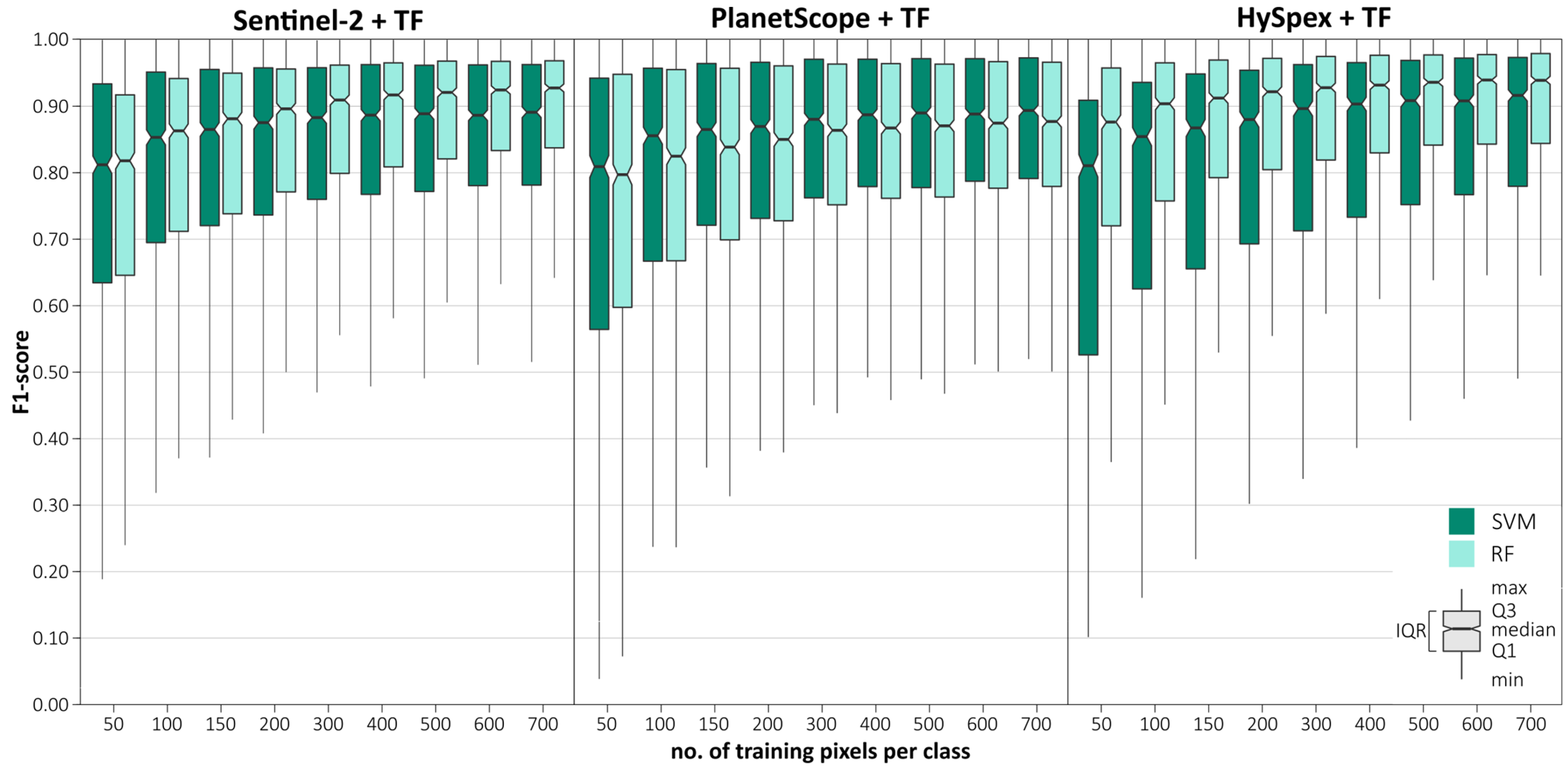
In the first stage, we aimed to determine the effect of the number of pixels on classification accuracy (from 50 to 700 pixels;
Figure 6). We obtained the highest median F1-score for the maximum number of pixels used (700): Sentinel-2 (0.93 RF; 0.89 SVM), PlanetScope (0.89 RF; 0.87 SVM), and HySpex (0.95 RF; 0.92 SVM). The results for the HySpex data were characterized by a high interquartile range (0.15–0.38), with a particularly large difference between the SVM and RF algorithms (by 0.06–0.15), followed by the PlanetScope data (0.17–0.31). We observed the narrowest range for the Sentinel-2 data (0.11–0.23); however, in this case, we found no difference between RF and SVM (0.01–0.05). As the number of pixels increased, the interquartile ranges of the results tended to decrease, by 0.03 per 100 pixels on average. The range increased by an average of 0.02 for every 100 pixels, with the largest increase observed between 50 and 200 pixels (by 0.04–0.09), whereas the difference between 50 and 700 pixels was 0.09–0.13. The results stabilized between 500 and 700 pixels and then changed little (0.01–0.02). Therefore, we considered the value of 700 pixels as optimal, which we used for further classification and analysis.
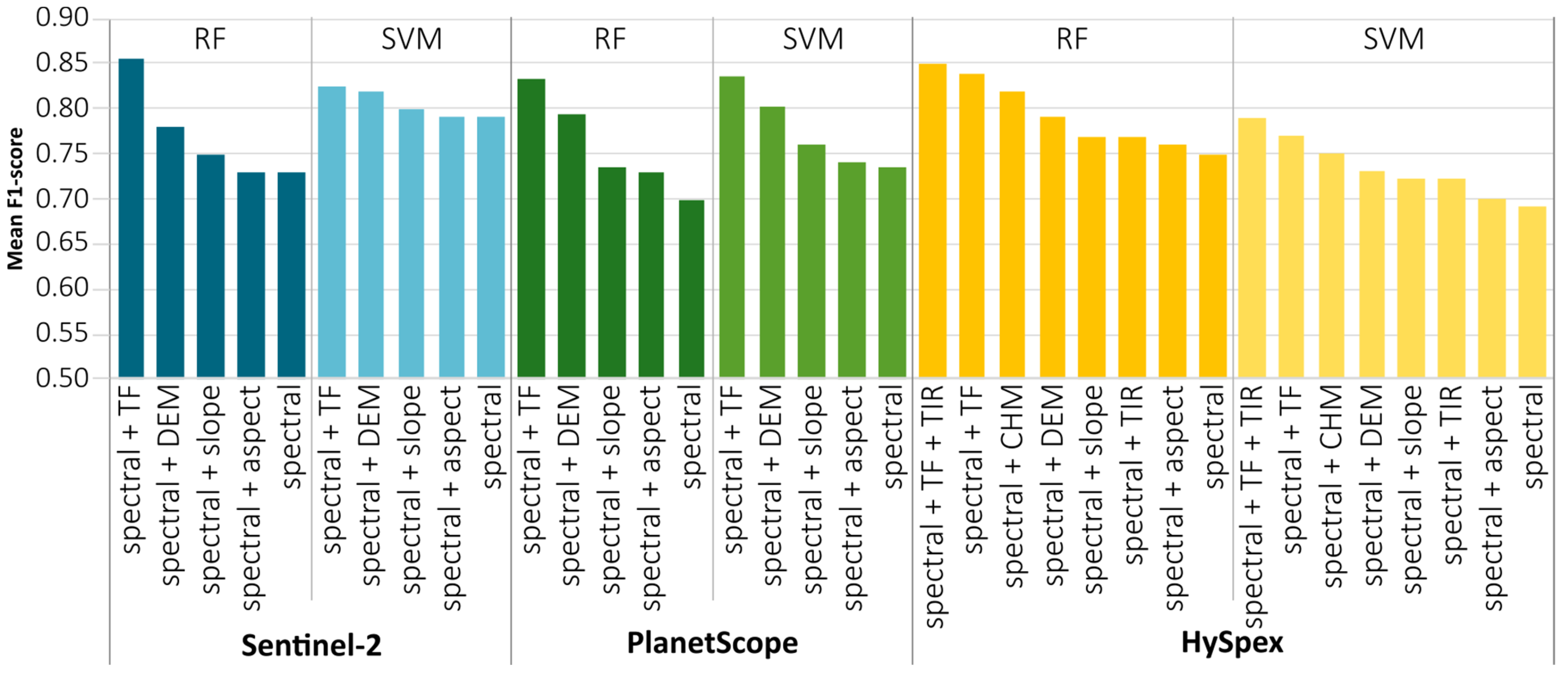
The average F1-score for the topographic features data (
Figure 7) oscillated around 0.77–0.86: Sentinel-2 (0.86 RF; 0.82 SVM), PlanetScope (0.83 RF; 0.84 SVM), and HySpex (0.84 RF; 0.77 SVM). The random forest algorithm performed better on the Sentinel-2 (by 0.04) and HySpex (by 0.07) data, whereas the SVM was more accurate on PlanetScope imagery (by 0.01). As shown by the results, using only spectral data, without any additional data, produced less accurate performance of 0.10 (range: 0.69–0.77) on average. Analyzing the influence of individual topographic variables (
Figure 7), the most accurate results were achieved by digital elevation model, which improved the results (by 0.04–0.10) in relation to the spectral data, followed by slope (by 0.01–0.03). Aspect least affected the result, only slightly increasing the classification accuracy (0.01–0.02), because the occurrence of forest stands in this area is only slightly influenced by direction. As such, aspect is more important in nonforest communities (shaded areas with accumulated snow affecting the type of vegetation). For hyperspectral data, we used airborne laser scanning derivatives, for which additional input was provided by the canopy height model, which increased the accuracy by 0.04, and thermal data, by 0.03. The combination of all topographic variables provided the best results, which increased the accuracy on average by 0.10 compared with those using spectral data. We found the smallest differences for Sentinel-2 imaging with the SVM algorithm, where the impact was small (by 0.05); we found the biggest differences for PlanetScope data with the RF classifier (by 0.14).
The next step was checking the relevance of the individual dates and channels of the PlanetScope (
Table 3) and Sentinel-2 (
Table 4) multitemporal composites. Comparing the influence of individual bands on the classification result using the mean decrease in accuracy (MDA) value, we observed the following: the most important period for the identification of forest stands is spring (month of May); this is when fully formed leaves of conifers can be observed. However, depending on the species, young leaves, needles, and shoots develop at difference paces; their color, shape, and size depend on the species as well. Another important observation was the usefulness of the red-edge bands, in the case of PlanetScope: the red-edge (B8), green (B3), red (B7;
Table 3), and for the Sentinel-2 the most informative were the 20 m channels B6, B5, B7, 10 m B2, and 60 m B9 (SWIR;
Table 4). This was expected because, due to plant pigments, the visible and red-edge range as well the cell structures in NIR play key roles in vegetation analyses. Notably, the red-edge range, which provides information about the condition of vegetation, played a key role in both cases.
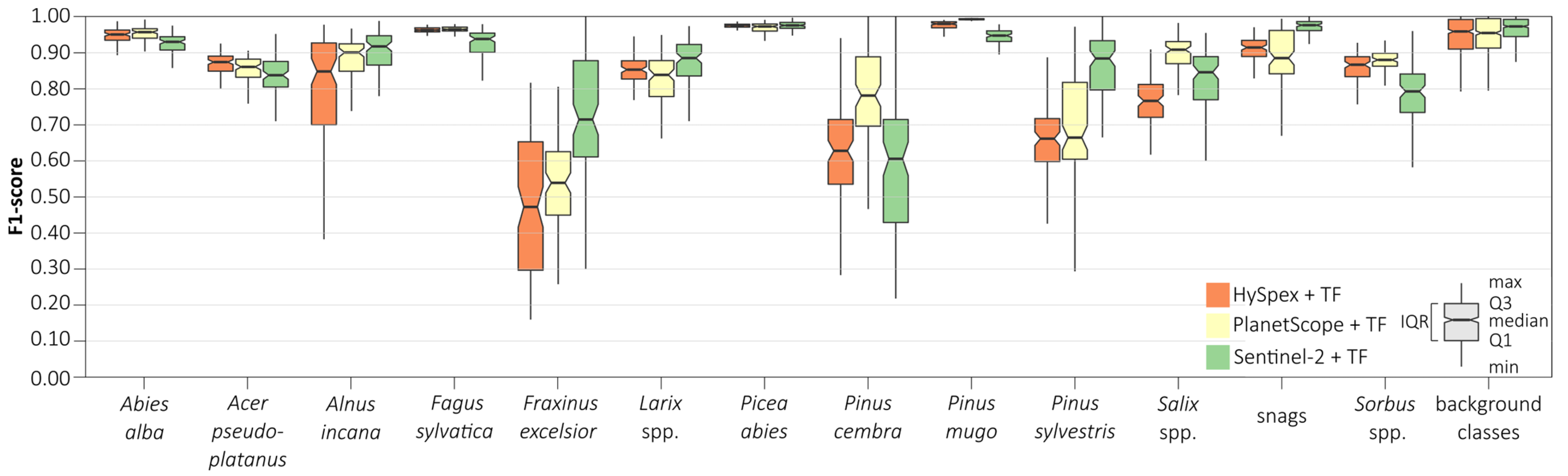
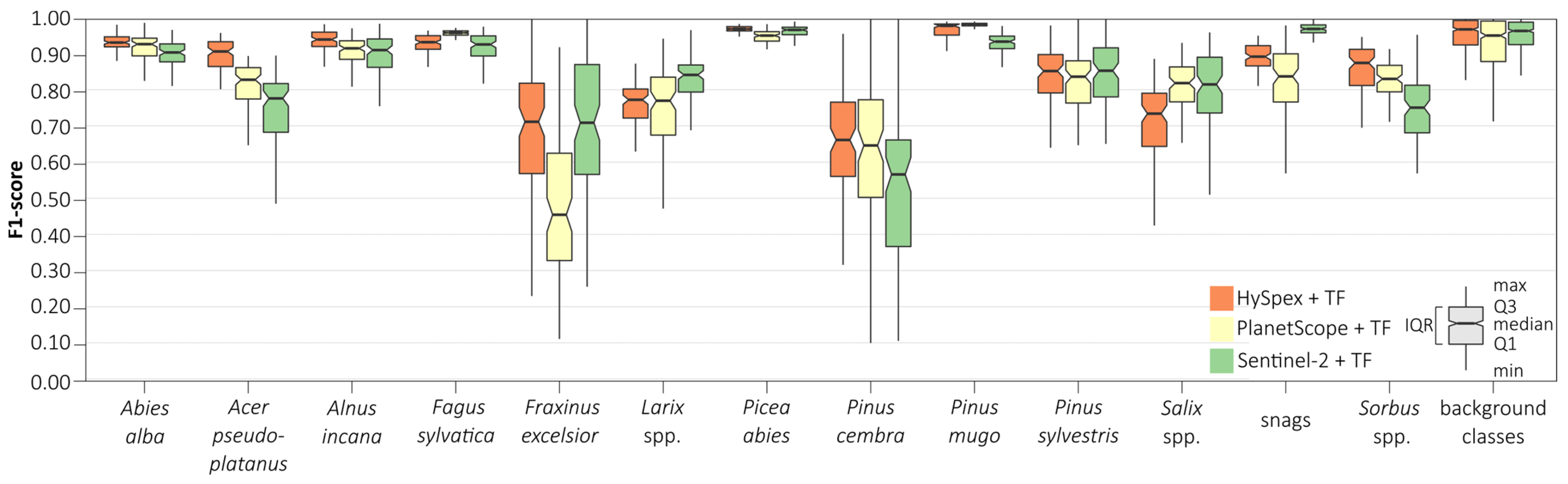
We conducted an iterative assessment of classification accuracy for each class depending on the dataset used based on the random forest (
Figure 8) and support vector machines (
Figure 9) classifiers. We obtained the highest median scores (above 0.90) for all remote sensing sets for the following classes: silver fir (
Abies alba; 0.91–0.95), European beech (
Fagus sylvatica; 0.92–0.96), Norway spruce (
Picea abies; 0.97–0.98), dwarf mountain pine (
Pinus mugo; 0.95–0.99), and grey alder (
Alnus incana; 0.87–0.92). We obtained the lowest ones for European ash (
Fraxinus excelsior; 0.44–0.72), Swiss stone pine (
Pinus cembra; 0.58–0.78), and Scots pine (
Pinus sylvestris; 0.67–0.85), which were also characterized by wide median differences (0.02–0.25) among the results obtained between the different remote sensing datasets. The widest interquartile range (IQR) was obtained for rare classes: European ash (
Fraxinus excelsior; 0.10–0.32), Swiss stone pine (
Pinus cembra; 0.20–0.27), and Scots pine (
Pinus sylvestris; 0.12–0.38); we obtained the narrowest IQR for silver fir (
Abies alba; 0.03–0.05), sycamore (
Acer pseudoplatanus; 0.04–0.08), European beech (
Fagus sylvatica; 0.01–0.05), Norway spruce (
Picea abies; 0.01–0.02), and dwarf mountain pine (
Pinus mugo; 0.01–0.02). We observed similar patterns for the SVM classifier, but these interquartile ranges for most classes were notably wider (by 0.01–0.07). Analyzing various classification scenarios, we found that random forest produced the most accurate classification results on Sentinel-2 and PlanetScope images, whereas support vector machine was more appropriate for HySpex data.
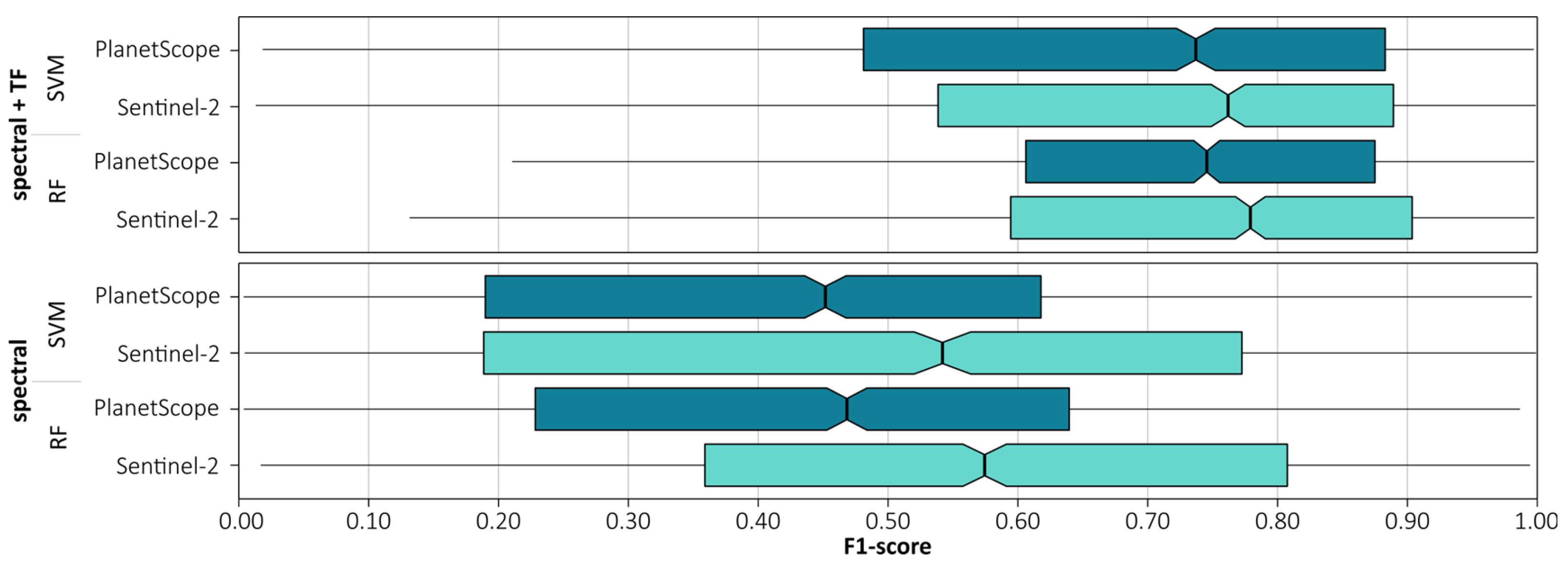
We also compared the classifications between PlanetScope and Sentinel-2 for the same acquisition date (19 May 2022) and time (approximately 9:00 UTC), allowing for the same conditions of solar irradiance and incidence angle (
Figure 10). Sentinel-2 spectral data outperformed PlanetScope data for both the random forest (0.58 vs. 0.47 F1-score) and support vector machine (0.53 vs. 0.45 F1-score) classifier. The differences between the classifiers substantially decreased when topographic derivatives were included, by an average of 0.23 points. However, the results obtained using Sentinel-2 data still scored higher (RF: 0.78, SVM: 0.76) than those obtained using PlanetScope data (RF: 0.74, SVM: 0.73), indicating the importance of including topographic product data in mountain vegetation analyses. Even with a single scene, high accuracy can be achieved.
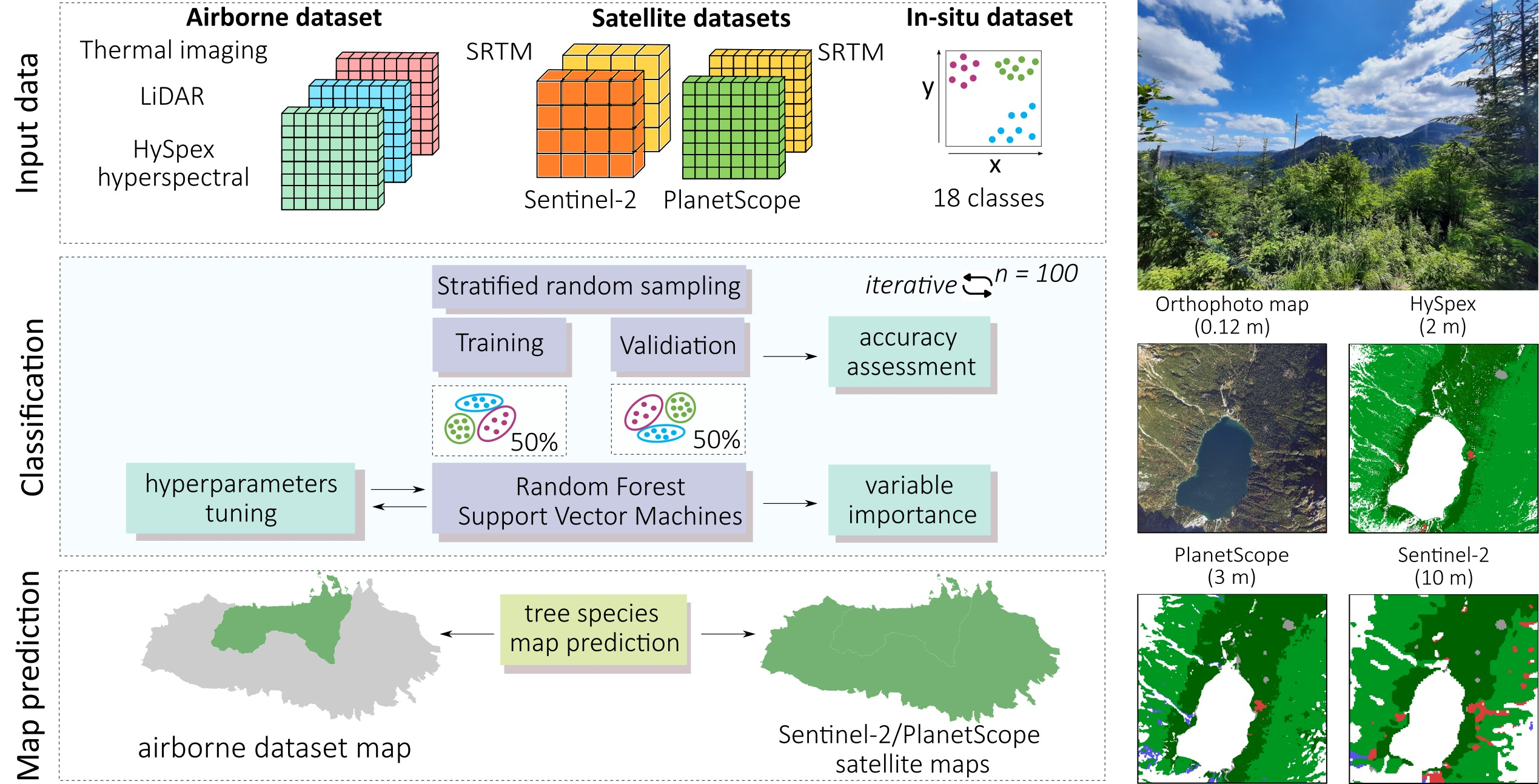
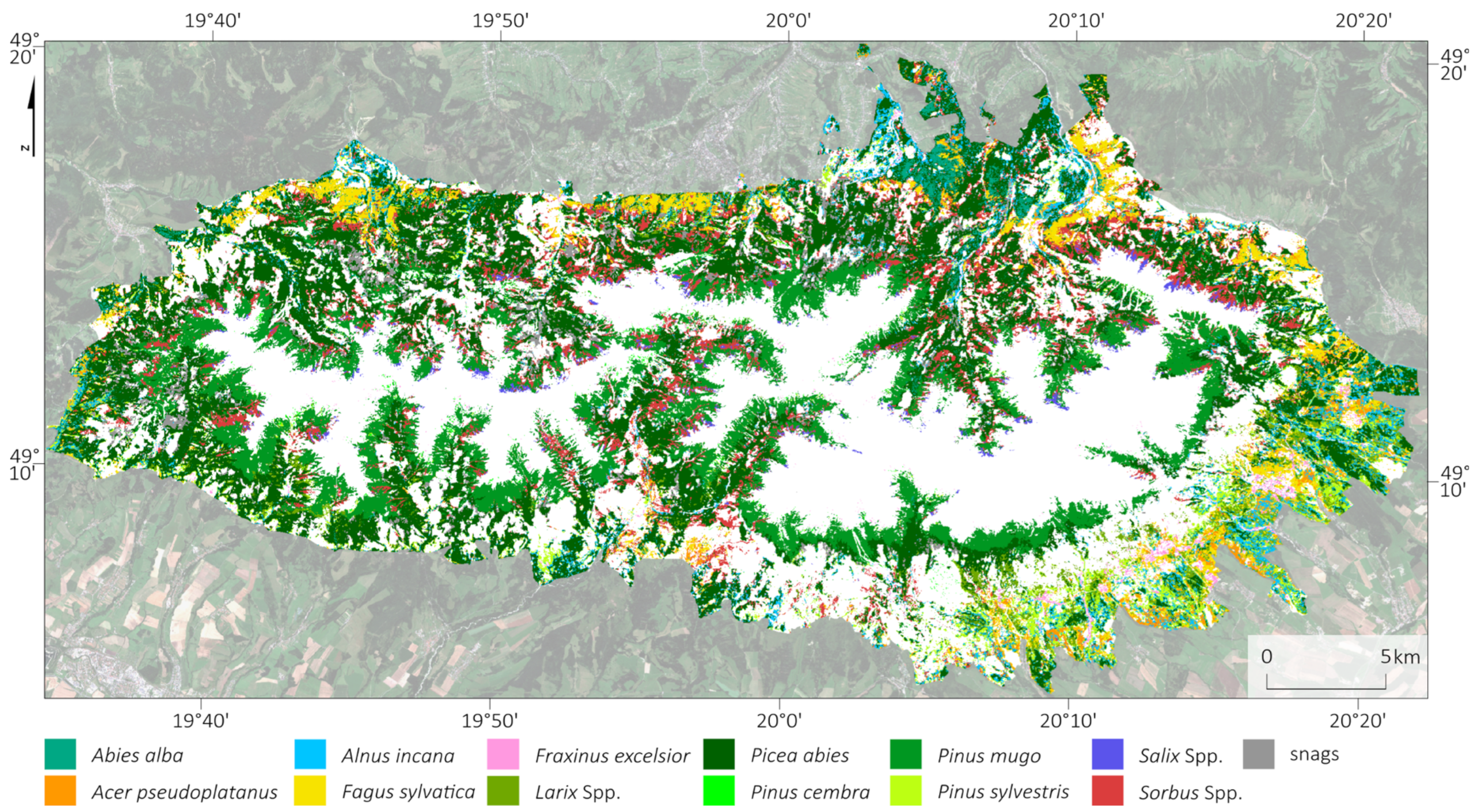
Based on the best classification results according to the mean F1-score for tree species classes, we classified the tree species in the Tatra Sentinel-2 area (
Figure 11;
Table 5), as well as for PlanetScope and HySpex imagery, the locations of which were compared with each other (
Figure 12). For this purpose, we selected three areas (dimensions: 1.5 km × 1.5 km = 225 ha) at different elevations and with different stand characteristics to reflect the diversity of the Tatra stands (deciduous, coniferous, and forests undergoing large-scale transformations) and to check the repeatability of the results. The obtained maps and error matrices (
Table 5 and
Appendix A) showed high convergence and a pattern of tree stand occurrence, with differences mainly due to the spatial resolution of the data (2, 3, and 10 m), the angle of sunlight incidence, and the effect of shadows. The maps were compared with a very-high-resolution orthophoto map (pixel size: 0.12 m).
4. Discussion
The results of our analyses confirmed the high potential of remote-sensing-based methods for the identification of forest species in protected areas. Despite the heterogeneity of the environment and topographic features, we successfully identified all dominant woody species of the Tatras, and the results should be considered satisfactory: for 10 out of 13 species, we obtained an average F1-score above 0.80, 6 of which were above 0.90. The accuracy of coniferous species identification was slightly higher on average (F1: 0.87) than for broadleaf species (F1: 0.84). Even these rarer species were well-identified on the Sentinel-2, PlanetScope, and HySpex images, allowing their successful identification on larger polygons in some areas of the TPN. However, in general, as they did not create compact and homogeneous surfaces, most of the identified patterns had error (IQR values were high) resulting from the share of other species, which created mixed tree crowns. Comparing our outcomes with those of other authors (
Table 6), our results obtained from HySpex images are a few percentage points higher than those of Shi et al. [
61], who used the same algorithm (RF). Our results are similar to those of Raczko et al. [
62], who used ANN and 280 spectral channels from APEX aerial imaging for the Karkonosze Mountains (mountain forests on the border of Poland and the Czech Republic). The proposed method of using multitemporal Sentinel-2 scenes (252-band data cube from 21 scenes representing different vegetation periods), allowed us to identify the studied species with higher accuracy than if we had used deep neural classifiers (CNN [
26] and XGB [
63]). Xi et al. [
64], comparing different algorithms, confirmed the dominance of RF over three other types of deep neural network (Conv1D, AlexNet, and LSTM) for a few species, which we identified (
Table 6), as well as for Manchurian ash classification. Additionally, RF produced results similar to those of Conv1D, AlexNet, LSTM, and SVM for Dahurian larch and white birch. RF scored worse than the best Conv1D classifier by approximately 6–8 percentage points in identifying Amur linden, Korean pine, and aspen [
64].
Waser et al. [
65], in mapping the dominant leaf type based on Sentinel-1/-2 data, found that UNet convolutional neural networks and RF obtained similar results; in some cases, UNet obtained worse results, depending on stand characteristics, type, and heterogeneity. Additionally, mapping a relatively low number of species (aspen, birch, pine, and spruce) on Sentinel-2 imagery, despite testing different types of CNN architectures, resulted in moderate F1-score of 0.68–0.76 using UNet with a ResNet encoder [
66] or 0.42–0.85 for feature pyramid network (FPN) with EfficientNet infrastructure [
67]. This demonstrates the difficulty of using deep learning methods to classify stands from medium-resolution satellite imagery (such as Sentinel-2), where spectral features are mainly relevant, especially for complex forest areas with diverse tree stands. Here, capturing the spatial patterns that occur in high-resolution imagery is difficult.
The identification of individual species mainly depends on the homogeneity and size of the analyzed patches. Notable analyses were published by Illarionova et al. [
67], who, using various methods of aggregation of individual patches, obtained classification accuracies (F1-score) in the range of 0.42–62 for aspen, 0.72–0.83 for birch, 0.81–0.84 for pine, and 0.74–0.76 for spruces. For the same species, but for compact and homogeneous patches, the accuracies were 0.77 for aspen, 0.90 for birch, 0.94 for pine, and 0.88 for spruce. We obtained similar observations from our analysis of our study area, where the species were well-known and well-identified, as shown in the box plots (
Figure 8 and
Figure 9). For example, ash, larch, pine, maple, and willow were clearly visible during field verification because, in areas covered by homogeneous and compact trees, the range of classified patterns coincided with the actual range of occurrence of these species. In situations where single trees formed a mix with other species of equal size, the classification result was incorrect. Based on Sentinel-2 images, we achieved the following average F1-score for the dominant tree species, which formed heterogeneous patches (
Table 6): ash, 0.81; larch, 0.84; pine, 0.83; and willow, 0.78. For comparison, other researchers obtained the following results: pine, 0.79–0.92; larch, 0.75 [
63]; larch, 0.93 [
29]; ash, 0.82; willow, 0.96 [
68]; pine, 0.78 [
69]; and larch, 0.70 [
18]. The resulting differences in the classification accuracies for woody species may be due to classes such as larch, Scots pine, or silver fir growing in small groups and often as admixtures in Norway spruce tree stands, which occupy the largest area and form compact homogeneous forests. Similar patterns were observed with rowan and willow, which coexist with dwarf pine, which form continuous, uniform areas. We encountered no difficulties in identifying these. We achieved high accuracies in identifying European beech trees and slightly lower accuracy in maple trees, which co-occur in stands due to their similar habitats, which could mutually influence the accuracies. Raczko and Zagajewski [
62] also obtained lower accuracies for a large mixture of woody species. The lower scores for European ash were likely due to this species growing linearly along streams and rivers, hindering identification due to mixing with water pixels. High accuracies were also obtained for snags, with only the PlanetScope data showing lower accuracies, which may have occurred because its sensor does not have a short-wave infrared (SWIR) channel that interacts with water content and distinguishes dry trunks. The results obtained for
Pinus cembra and
Fraxinus excelsior showed that despite the presence of single specimens, they were identifiable, but the scores were not high, but demonstrated the possibility of classifying rare species.
Table 6.
Comparison of obtained classification results (mean F1-score) for dominant tree species with those in the literature. S.f.—silver fir (Abies alba); Syc.—sycamore (Acer pseudoplatanus); G.a.—grey alder (Alnus incana); E.b.—European beech (Fagus sylvatica); E.a.—European ash (Fraxinus excelsior); Lar.—larch (Larix spp.); N.s.—Norway spruce (Picea abies); S.s.p.—Swiss stone pine (Pinus cembra); D.m.p.—dwarf mountain pine (Pinus mugo); S.p.—Scots pine (Pinus sylvestris); Wil.—willow (Salix spp.); Row.—rowan (Sorbus spp.). Results for same genera are marked in bold.
Table 6.
Comparison of obtained classification results (mean F1-score) for dominant tree species with those in the literature. S.f.—silver fir (Abies alba); Syc.—sycamore (Acer pseudoplatanus); G.a.—grey alder (Alnus incana); E.b.—European beech (Fagus sylvatica); E.a.—European ash (Fraxinus excelsior); Lar.—larch (Larix spp.); N.s.—Norway spruce (Picea abies); S.s.p.—Swiss stone pine (Pinus cembra); D.m.p.—dwarf mountain pine (Pinus mugo); S.p.—Scots pine (Pinus sylvestris); Wil.—willow (Salix spp.); Row.—rowan (Sorbus spp.). Results for same genera are marked in bold.
| Author | Sensor | Classifier | No. of
Classes | S.f. | Syc. | G.a. | E.b. | E.a. | Lar. | N.s. | S.s.p. | D.m.p. | S.p. | Wil. | Snags | Row. |
|---|
| Present paper | S-2 | RF | 13 | 0.90 | 0.80 | 0.88 | 0.91 | 0.68 | 0.84 | 0.97 | 0.52 | 0.93 | 0.83 | 0.78 | 0.97 | 0.75 |
| SVM | 0.90 | 0.78 | 0.88 | 0.92 | 0.81 | 0.80 | 0.98 | 0.32 | 0.95 | 0.75 | 0.76 | 0.97 | 0.79 |
| Planet Scope | RF | 0.91 | 0.81 | 0.91 | 0.96 | 0.48 | 0.76 | 0.95 | 0.62 | 0.99 | 0.81 | 0.82 | 0.83 | 0.84 |
| SVM | 0.94 | 0.83 | 0.86 | 0.96 | 0.47 | 0.79 | 0.93 | 0.73 | 0.99 | 0.64 | 0.88 | 0.86 | 0.86 |
| HySpex | RF | 0.93 | 0.90 | 0.94 | 0.93 | 0.69 | 0.76 | 0.97 | 0.65 | 0.97 | 0.83 | 0.71 | 0.89 | 0.86 |
| SVM | 0.94 | 0.84 | 0.78 | 0.95 | 0.39 | 0.82 | 0.97 | 0.56 | 0.97 | 0.60 | 0.72 | 0.89 | 0.83 |
| [61] | HySpex | RF | 5 | 0.83 | 0.81 | - | 0.80 | - | - | 84 | - | - | - | - | - | - |
| [62] | APEX | ANN | 6 | - | - | 0.86 | 0.90 | - | 0.77 | 0.92 | - | - | 0.78 | - | - | - |
| [63] | S-2 | XGB | 7 | - | - | - | 0.85 | - | 0.75 | 0.92 | - | - | 0.86 | - | - | - |
| [26] | S-2 | CNN | 8 | - | - | - | 0.73 | - | 0.51 | 0.84 | 0.58 | 0.58 | 0.58 | - | - | - |
| [65] | S-2 | AlexNet | 8 | - | - | - | - | 0.79 | 0.81 | 0.77 | 0.58 | 0.58 | 0.58 | - | - | - |
| RF | - | - | - | - | 0.79 | 0.83 | 0.73 | 0.68 | 0.68 | 0.68 | - | - | - |
| [70] | S-2 | RF | 8 | 0.80 | - | 0.83 | 0.89 | - | 0.76 | 0.77 | - | - | 0.80 | - | - | - |
| [71] | S-2 | SVM | 11 | 0.90 | 0.69 | 0.90 | 0.93 | - | 0.73 | 0.92 | - | - | 0.78 | - | - | - |
| [72] | S-2 | RF | 12 | - | 0.65 | 0.83 | 0.82 | 0.72 | 0.76 | 0.94 | - | - | 0.87 | - | - | - |
| [73] | S-2 | RF | 12 | - | 0.59 | 0.91 | 0.90 | 0.77 | 0.94 | 0.97 | 0.95 | 0.95 | 0.94 | - | - | - |
| [74] | S-2 | RF | 17 | 0.67 | 0.25 | 0.87 | 0.93 | - | 0.83 | 0.80 | - | - | 0.99 | - | - | - |
Considering the impact of individual topographic variables, the DEM was the most influential for the Sentinel-2 and PlanetScope satellite data, increasing the accuracy of the datasets by an average of 0.06 on the F1-score. Waśniewski et al. [
75], for Sentinel-2 data, and Ye et al. [
50], with PlanetScope data, showed the feature importance value of the DEM. The other variables, slope and aspect, marginally contributed to the classification accuracy (by 0.01–0.03 F1-score), which was confirmed by Bhattarai et al. [
76]. When using a dense series of multitemporal satellite data, the effect of individual variables must be determined to identify the most important spectral bands and acquisition dates for image classification. Shirazinejad et al. [
77] used, multitemporal compositions to increase the overall classification accuracy by an average of 0.28 compared with that of single Sentinel-2 scenes. Waśniewski et al. for Sentinel-2 obtained the highest variable importance values for Sentinel-2 channels B2, B5, B6 which also reached high values in our analyses. For mountain forest communities, Kovačević et al. [
78], using Sentinel-2 imagery, determined that the B11, B2, and B12, as well as B1 and B9, were most relevant for classification, which especially matched the B2 and B9 results in our analyses. In our case, all spectral bands obtained on May 19 scored high MDA values; the second highest MDA scores were obtained from the July 4 imagery (in mountain ecosystems, this is still the beginning of the vegetation period). The same observation was reported by Plakman et al. [
23], but the authors focused only on the original 10 m pixels. So, for them, the most important spectral bands were B4 (spring and winter), B2 (spring), B4 (autumn and summer), and then B8 (spring and summer). In our case, the most important bands were also near-infrared bands: B6, B5, B7, B2, and then B9 (
Table 3). This means that the pixels resampled down to 10 m still contain important spectral characteristics, allowing the identification of plant species. Based on our results of the influence of individual periods, we observed that deciduous and coniferous trees show large differences in color in many months, which applies in particular to months with decreasing MDA values: May, July, November, August, September, and October (in our analyses, we did not obtain any high-quality June Sentinel-2 image). The following topographic factors with the highest scores were relevant: digital elevation model (134), slope maps (22), and aspect maps (10). Gan et al. [
79], based on Gaussian process regression (GPR, which determines the similarity between samples) and random forest regression (RFR; which determines the degree of reduction of mean square error (MSE) of feature variables) and using nine vegetation indices, confirmed that early April, late June, mid-July, and late October are the key phenological periods for vegetation analysis. Excluding April, the rest of these periods were confirmed in our study. Similar observations were noted for the Sentinel-2 bands, which allowed to identify woody species of the Giant Mts.; the three best were from spring acquisition: B6, (NDVI created on the basis of autumn imagery), B4 and B5, followed by 9 autumn spectral bands (in order of informativeness: B8A, B12, B7, B11, B3, NDWI, B6, B5, B8), and summer scenes offered about 50% lower informativeness than the spring acquisitions [
18]. Further investigating the impact of individual dates, the highest MDA values were obtained for acquired imagery from May to July for both Sentinel-2 and PlanetScope datasets, with values gradually decreasing subsequently. Moreover, Shirazehad et al. [
77] for complex broeadleaf tree stands obtained the highest values for May and June which was also confirmed in both the Xi et al. [
64] and Karasiak et al. studies [
68]. This indicates the high relevance of acquiring imagery during the period of full vegetation development, which allows better differentiation of woody species classes than discolored forest stands during the autumn period. Analyzing the effect of the number of pixels, we found that the optimal value was 700 pixels; comparable values were obtained by Hamrouni et al. [
80] on a dense series of Sentinel-2 imagery (26–36 scenes) for 6 classes of tree species and types, where 750 samples were optimal; when increasing to 1000 pixels, the OA results increased by one percentage point for the random forest classifier. Additionally, in our previous study [
81] of land cover mapping for both Sentinel-2 and Landsat 8 data, we found that the 700 pixels produced acceptable results for both the RF and SVM classifiers. The obtained classification accuracies were satisfactory for any spatial resolution (2, 3, or 10 m), as also reported by Xu et al. [
82], where the effect of spatial resolution on classification accuracy was measured. Four data sets were used: Gaofen-2 (4 m), Sentinel-2 (10 m), Gaofen-1 (16 m), and Landsat 8 (30 m), to classify four tree species using NDVI and forest phenological metrics with random forest as the classifier. The overall accuracy for 10 m Sentinel-2 was higher (86%) than for 4 m resolution (84%); at 16 and 30 m resolution, the OA gradually decreased to 79%. Similar results were obtained for airborne hyperspectral AISA-Eagle II (with 2 m resolution with 256 spectral bands) with a 0.78–0.80 mean F1-score and 0.84–0.85 for Sentinel-2 satellite data [
30].
Heterogeneity, both in terms of composition and age of the analyzed forest communities, is a major challenge. This applies in particular to mountain national parks, which, due to the height differences, different terrains are exposed to the influence of strong winds and a wide range of incoming sun rays [
83,
84]. Due to the area being protected, a large part of the forest stand dies naturally or falls victim to wind throws. An additional complication is that humans have introduced fast-growing species, such as spruce, into the lower parts of the mountains, where deciduous trees, such as beeches with an admixture of fir, should naturally occur. Also important is the impact of anthropogenic pressure in the form of the construction of water-related recreation centers, which absorb groundwater, but gases are emitted through the exhaust of watercraft, affecting the acidification of the air, which leads to chlorosis. Acidified rains wash out valuable mineral substances from the soil, which then has limited ability to neutralize the effects due to the geological structure, i.e., the presence of granite in the substrate. Hence, the identification of individual woody species in mountainous national parks faces different challenges than that in species-homogenous managed forests growing in lowlands. Over time, suitable tools can be used to guide the development of effective methods of monitoring vegetation.
The obtained results are valuable as they confirm the future ability to continuously monitor the environment, both due to the open access to Sentinel-2 data and the algorithms, which do not require commercial software. The proposed methods should be developed for monitoring the lower belts of mountain areas, which are undergoing ecosystem reconstruction, as artificial spruce plantings have been eliminated by bark beetle outbreaks and windthrows, and suitable deciduous species, e.g., beech and fir, naturally resume their place in the ecosystem, which are desirable changes. However, this process is hindered by the long-occurring spruce; the dropping of their needles, which contain tannins that acidify the soil, has limited the ability of the soil to neutralize these substances through the granite substrate. This acidification has substantially affected soil microorganisms, including mycorrhiza, which markedly hinders the development of suitable plant species, increasing the risk of the appearance of alien invasive or native expansive species. The second challenge is spruce monitoring, which should even be conducted at sites in the upper belt, which is a natural habitat for this species. However, due to climate change, reduced snow thickness, faster snow melting, and the subsequent soil dehydration that leads to an increase in air temperature inside the stand, water stress is created. These phenomena also improve the conditions for bark beetle outbreaks, which has two or even three breeding cycles per season, exposing spruce stands to new insect attacks.
Global changes in the environment are leading to another negative consequence: the occupation of new habitats by species only previously existing in the lower belts, e.g., mountain pine shrubs, which outcompete the species on many valuable grasslands.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}











