



Outbreak Mechanism of Locust Plagues under Dynamic Drought and Flood Environments Based on Time Series Remote Sensing Data: Implication for Identifying Potential High-Risk Locust Areas

 ,
, 
 ,
,  , and
, and 
Abstract
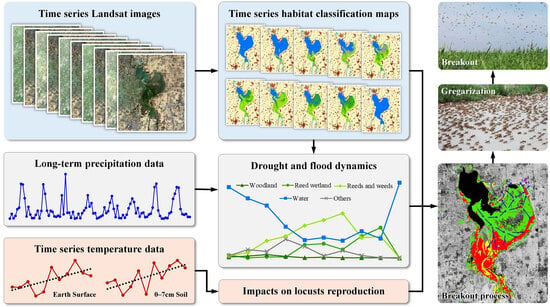
:
1. Introduction
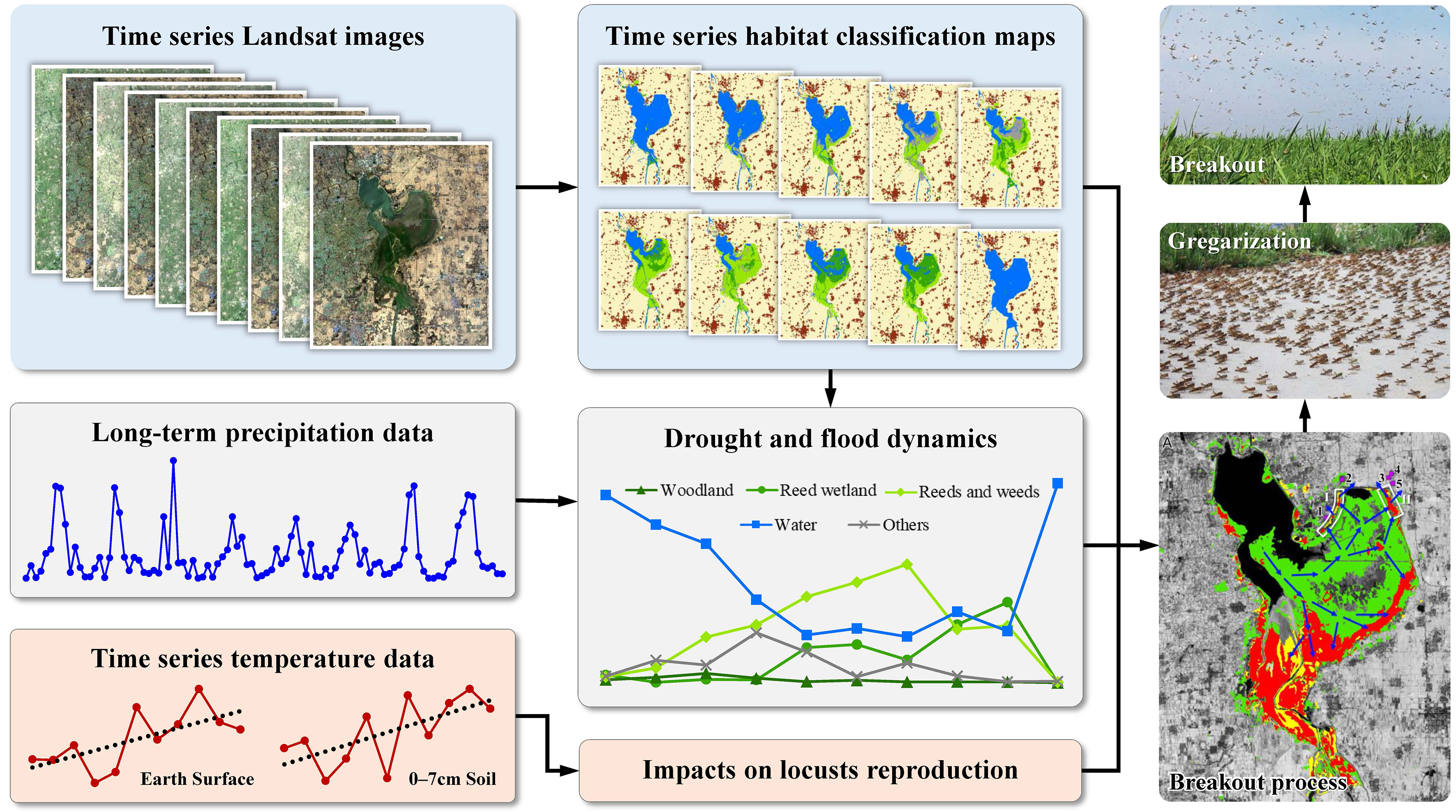
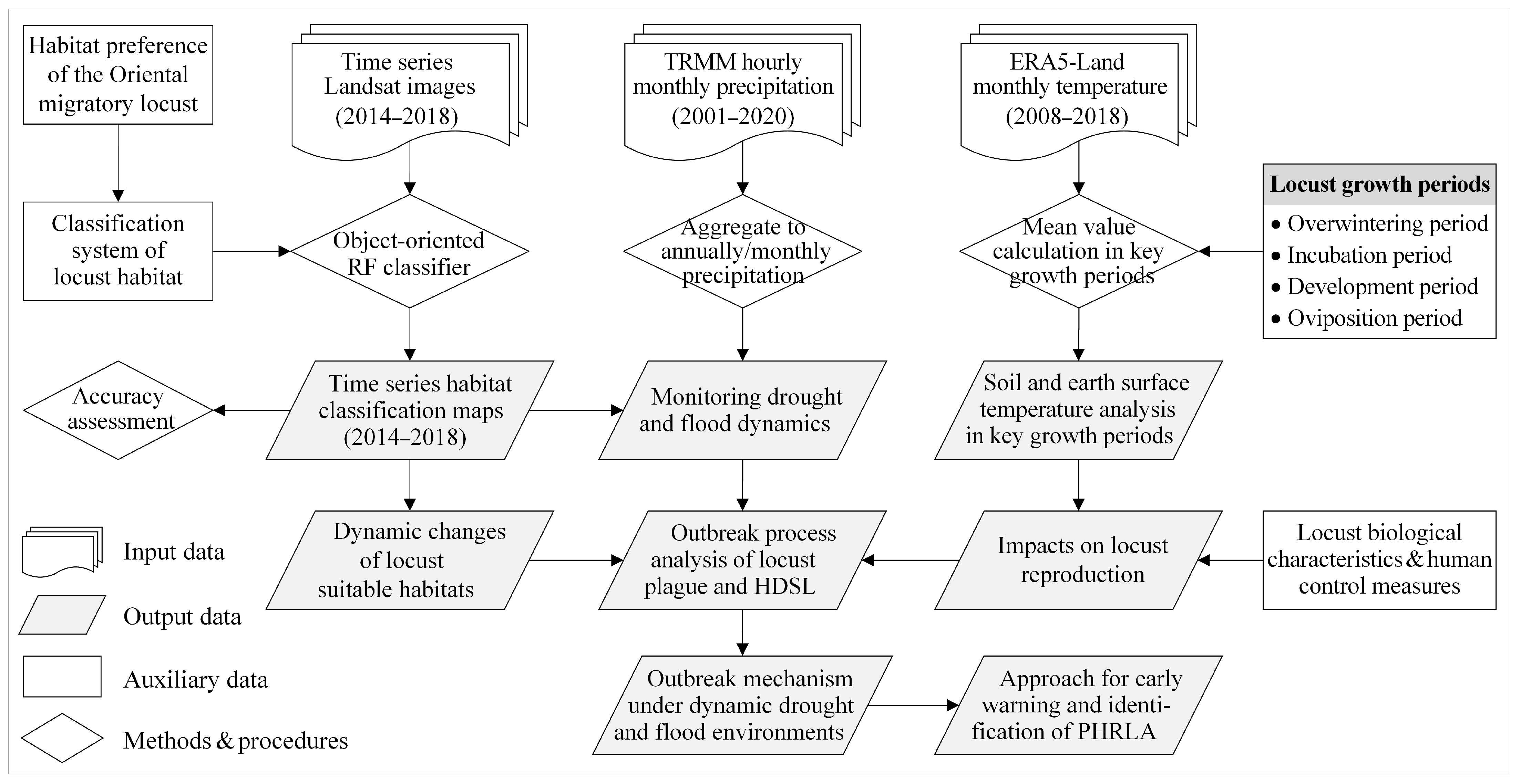
2. Materials and Methods
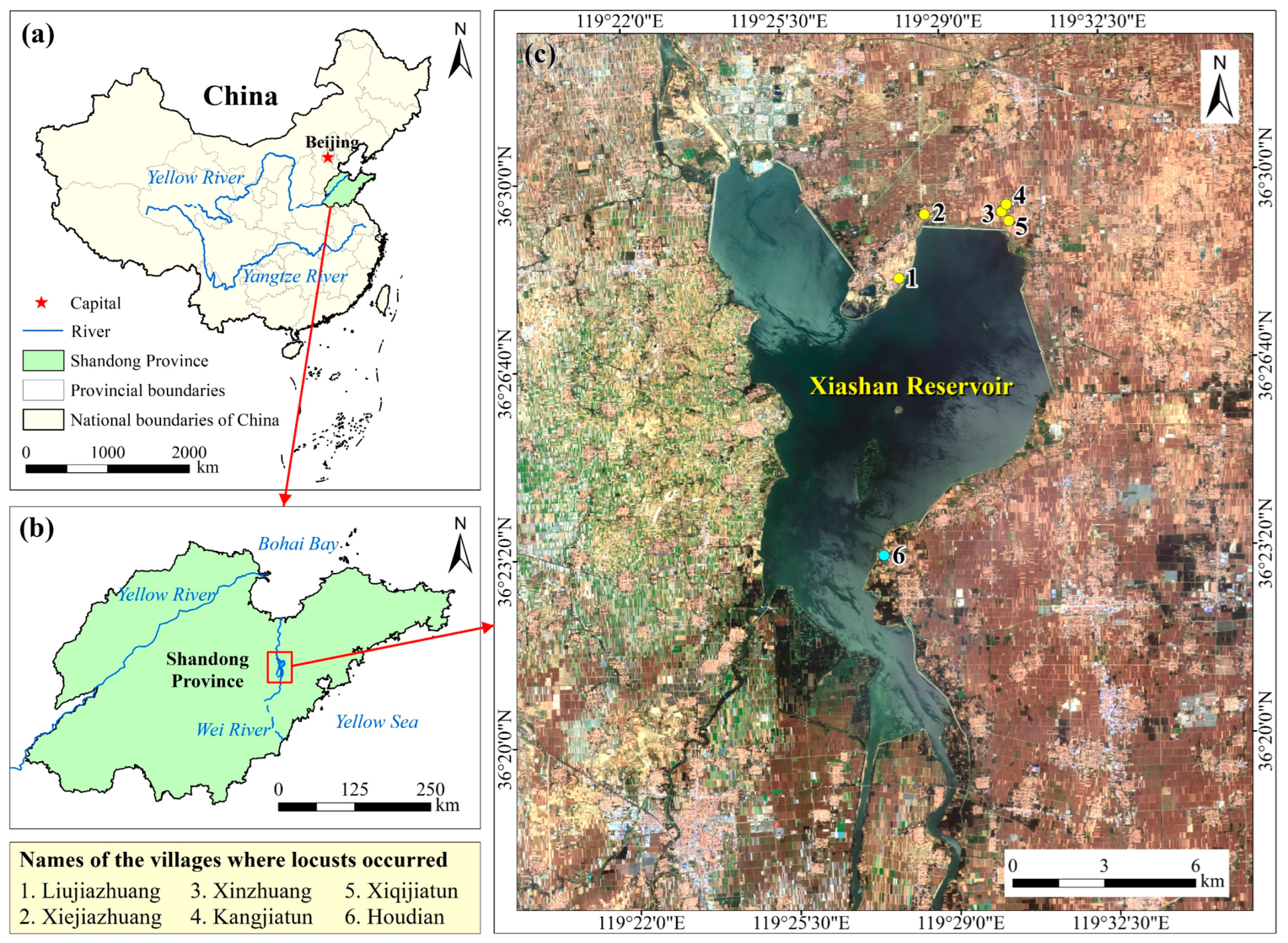
2.1. Study Area
2.2. Biological Characteristics of the OML
2.2.1. Annual Life History
2.2.2. Behavior Habits
2.3. Data Acquisition and Processing
2.3.1. Satellite Remote Sensing Images
2.3.2. Remote Sensing-Based Meteorological Products
2.3.3. Ground Survey Data of Locusts
2.3.4. Habitat Classes and Reference Data Selection
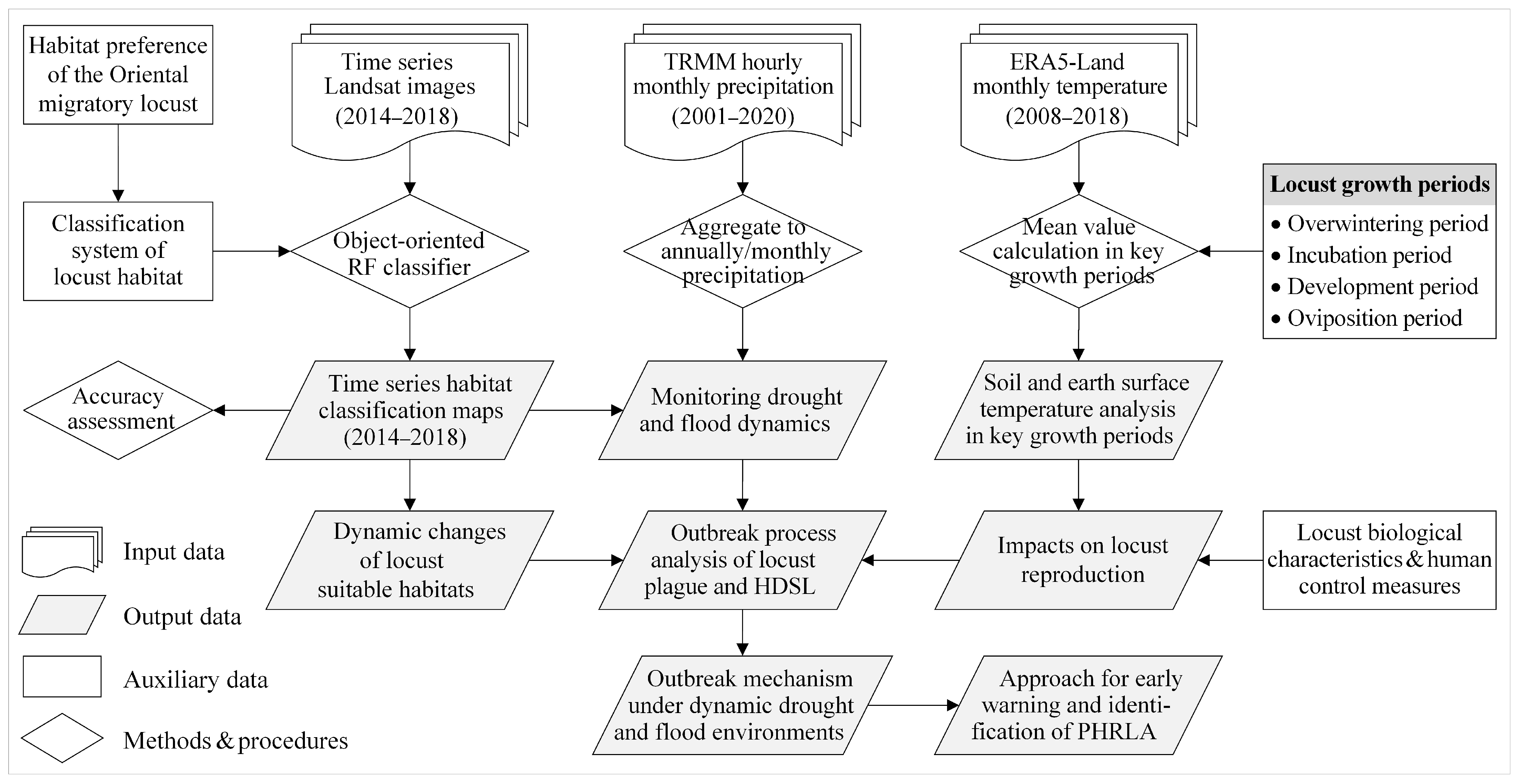
2.4. Methods
2.4.1. Object-Oriented Random Forest Classifier for Habitat Classification
2.4.2. Monitoring Drought and Flood Dynamics Based on Precipitation
2.4.3. Analysis of Temperature Conditions in Key Locust Growth Periods
3. Results
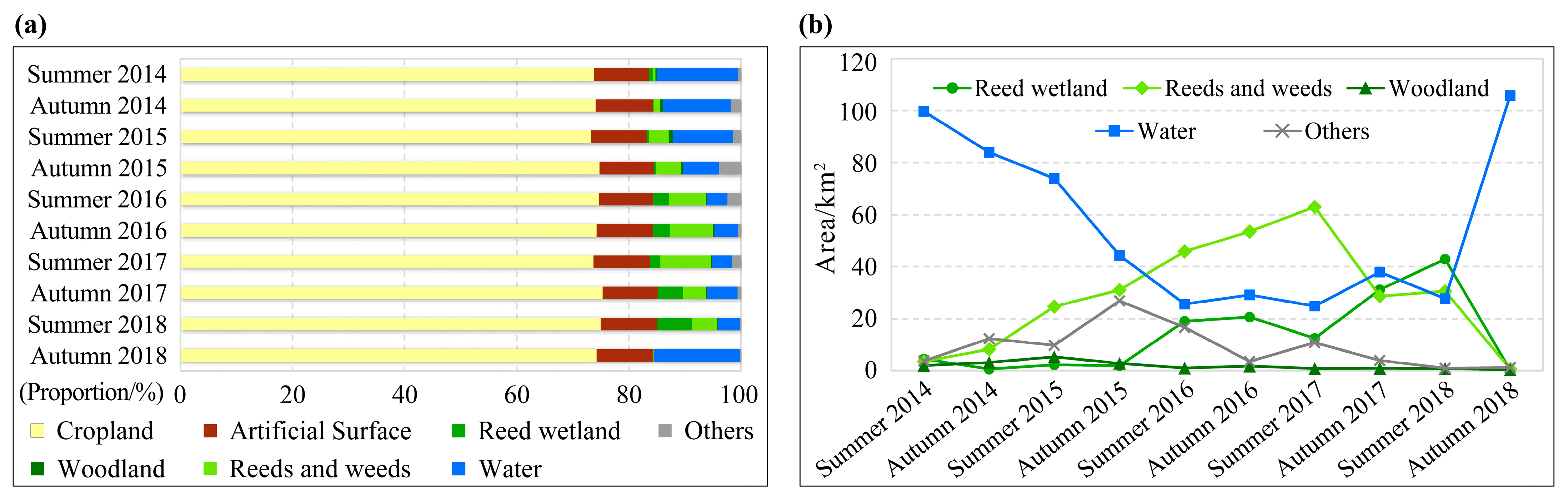
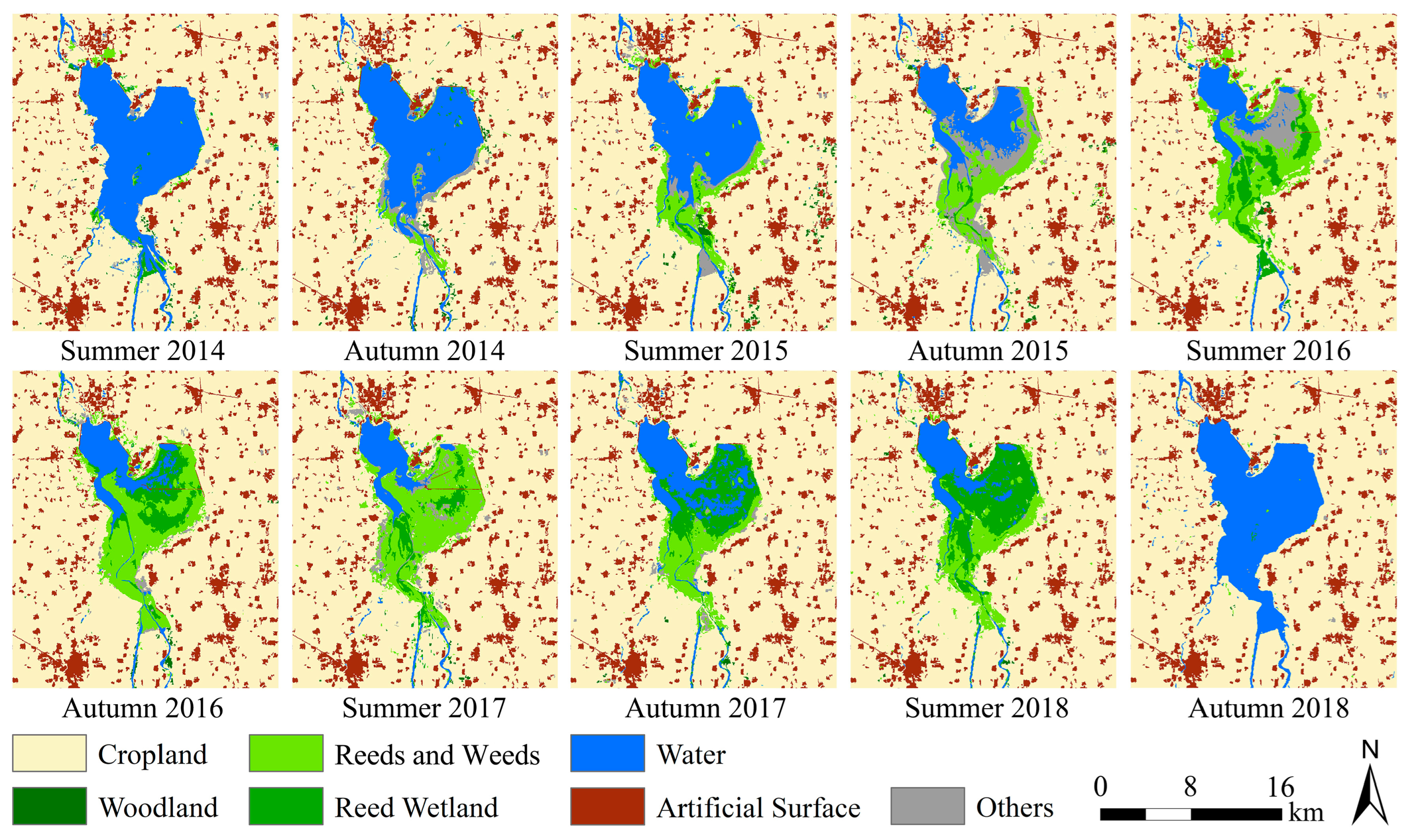
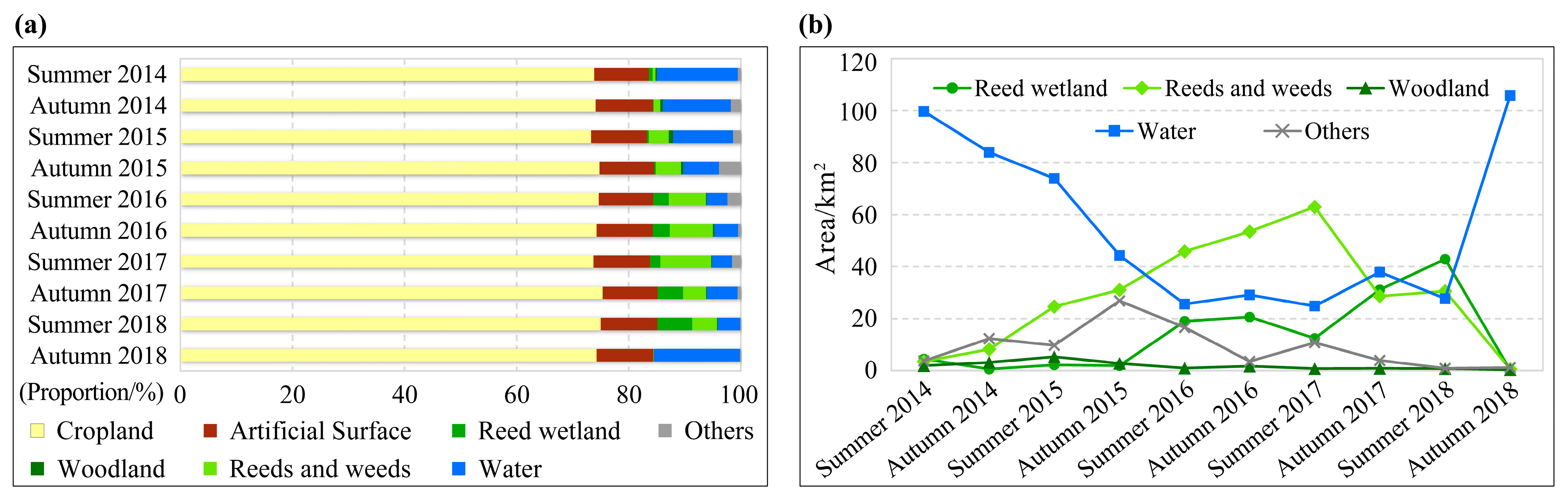
3.1. Habitat Classification Accuracy and Time Series Habitat Maps
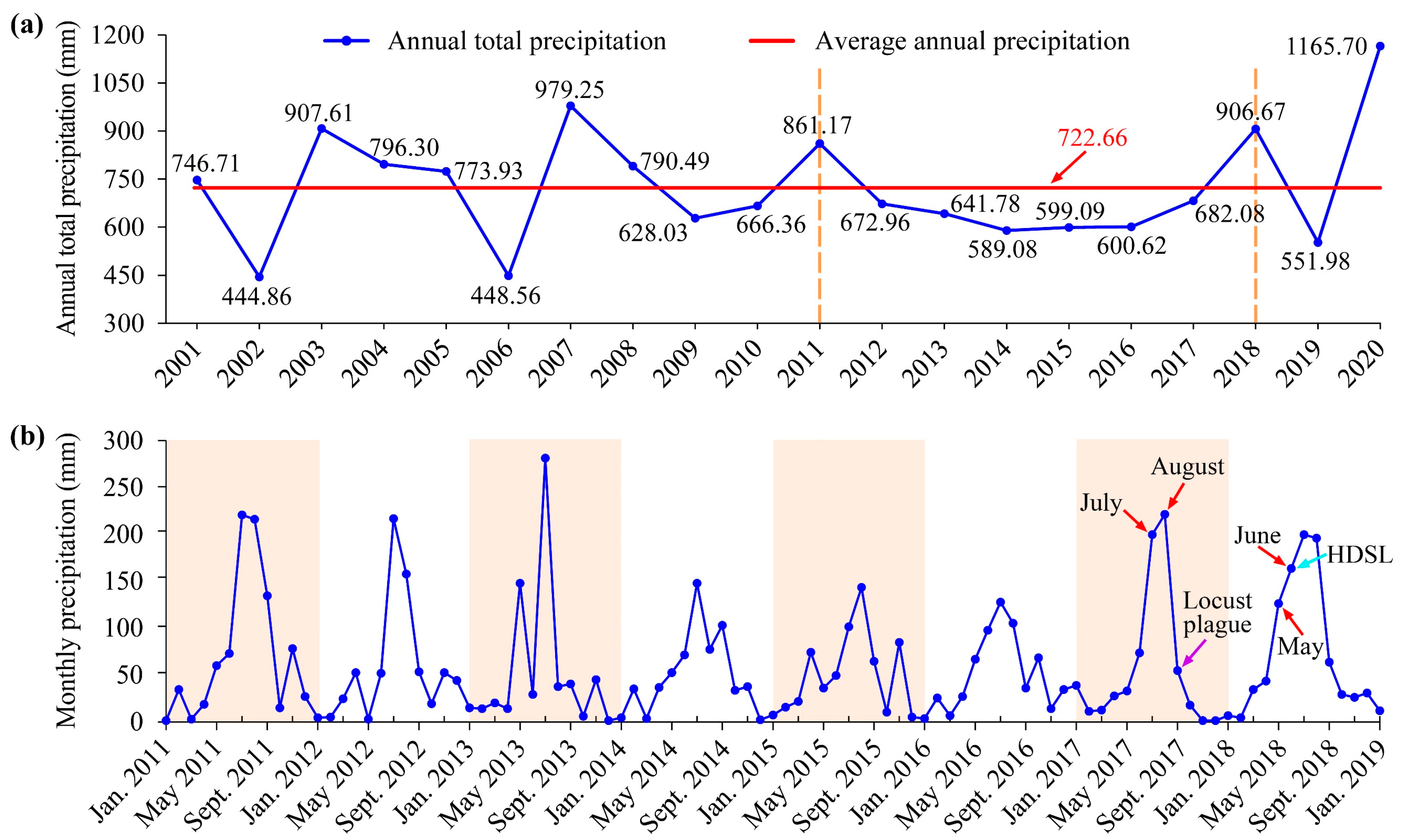
3.2. Dynamic Drought and Flood Environments of the Study Area
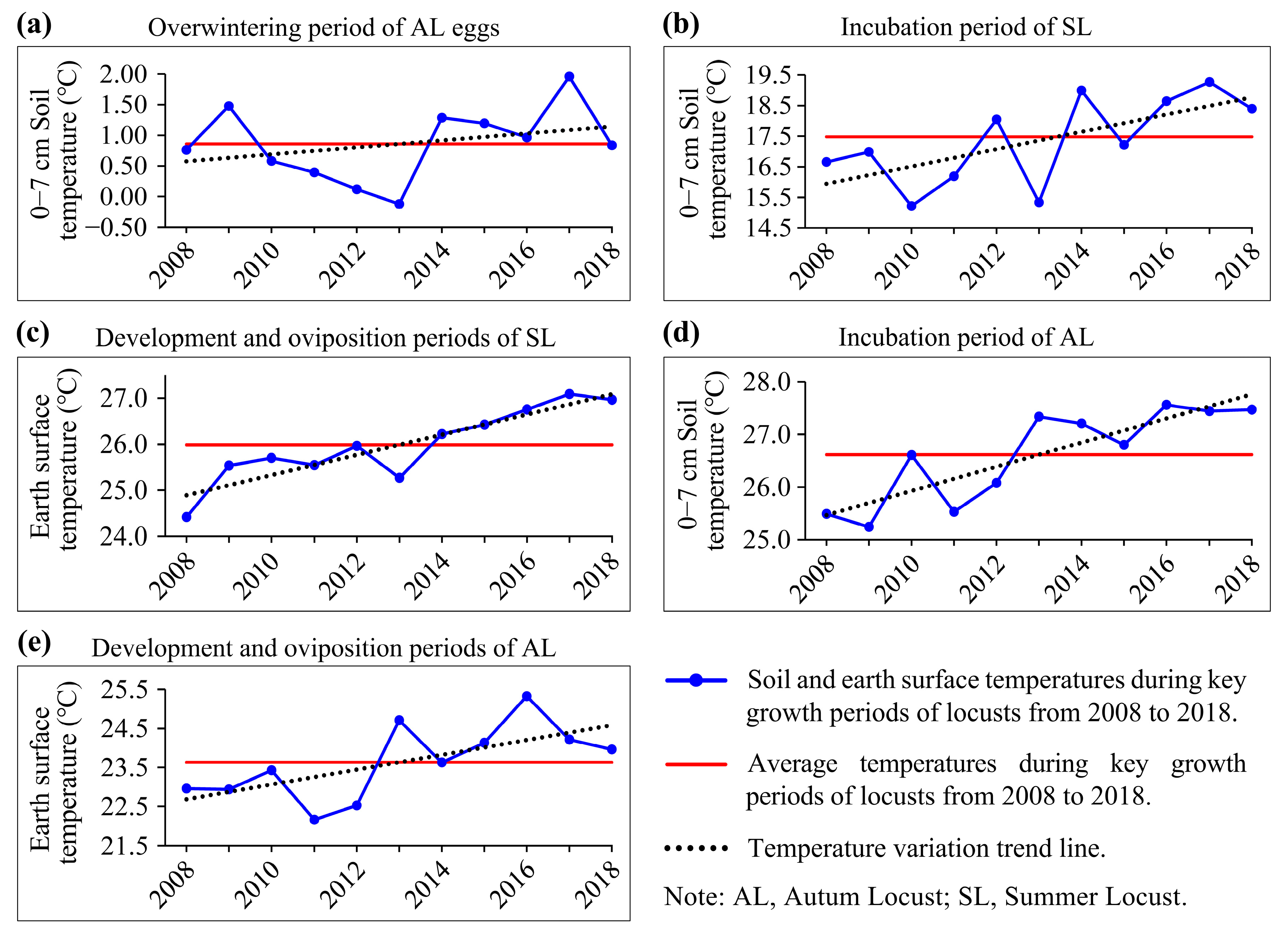
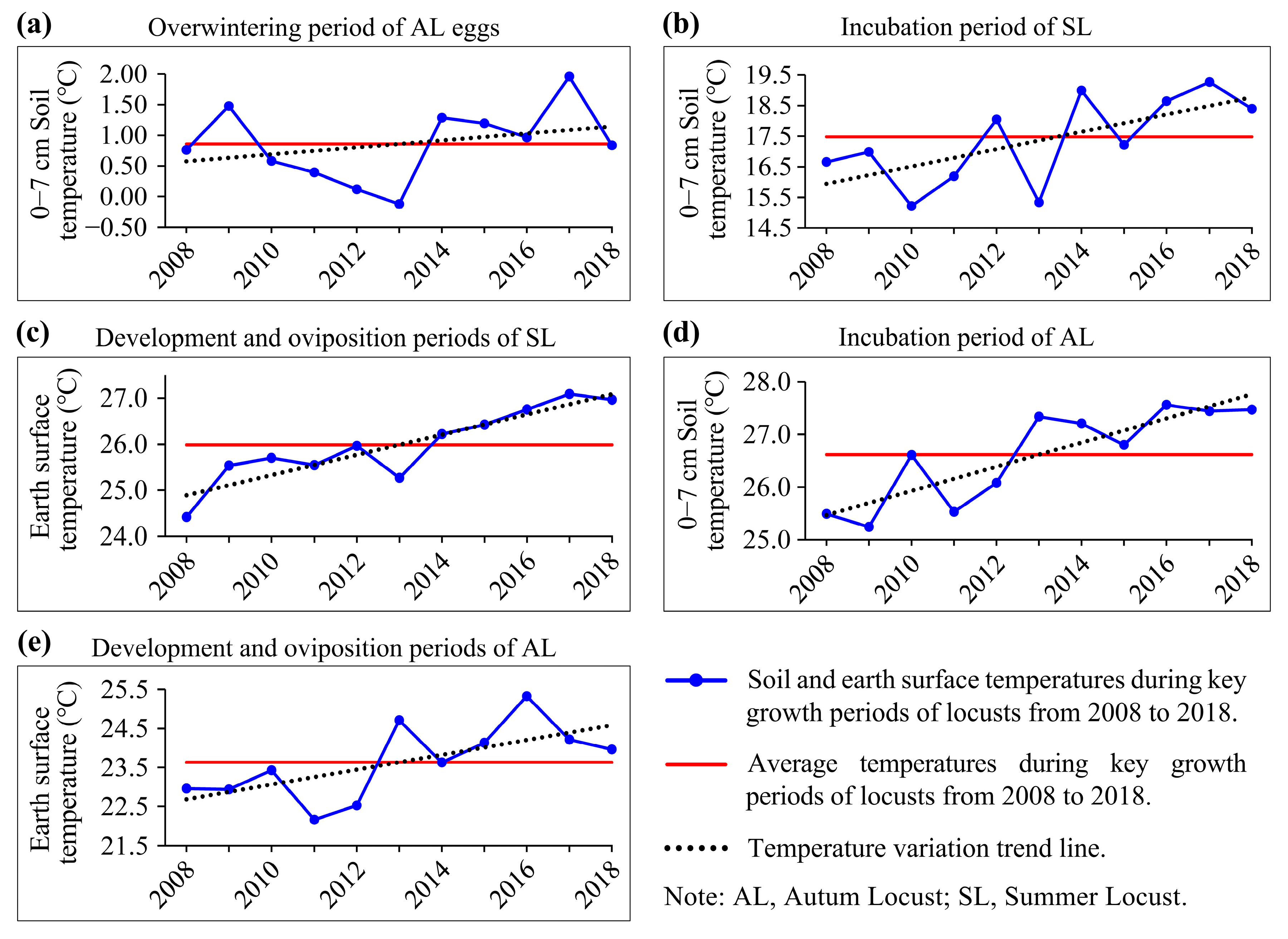
3.3. Impacts of Temperature Conditions on Locust Reproduction
4. Discussion
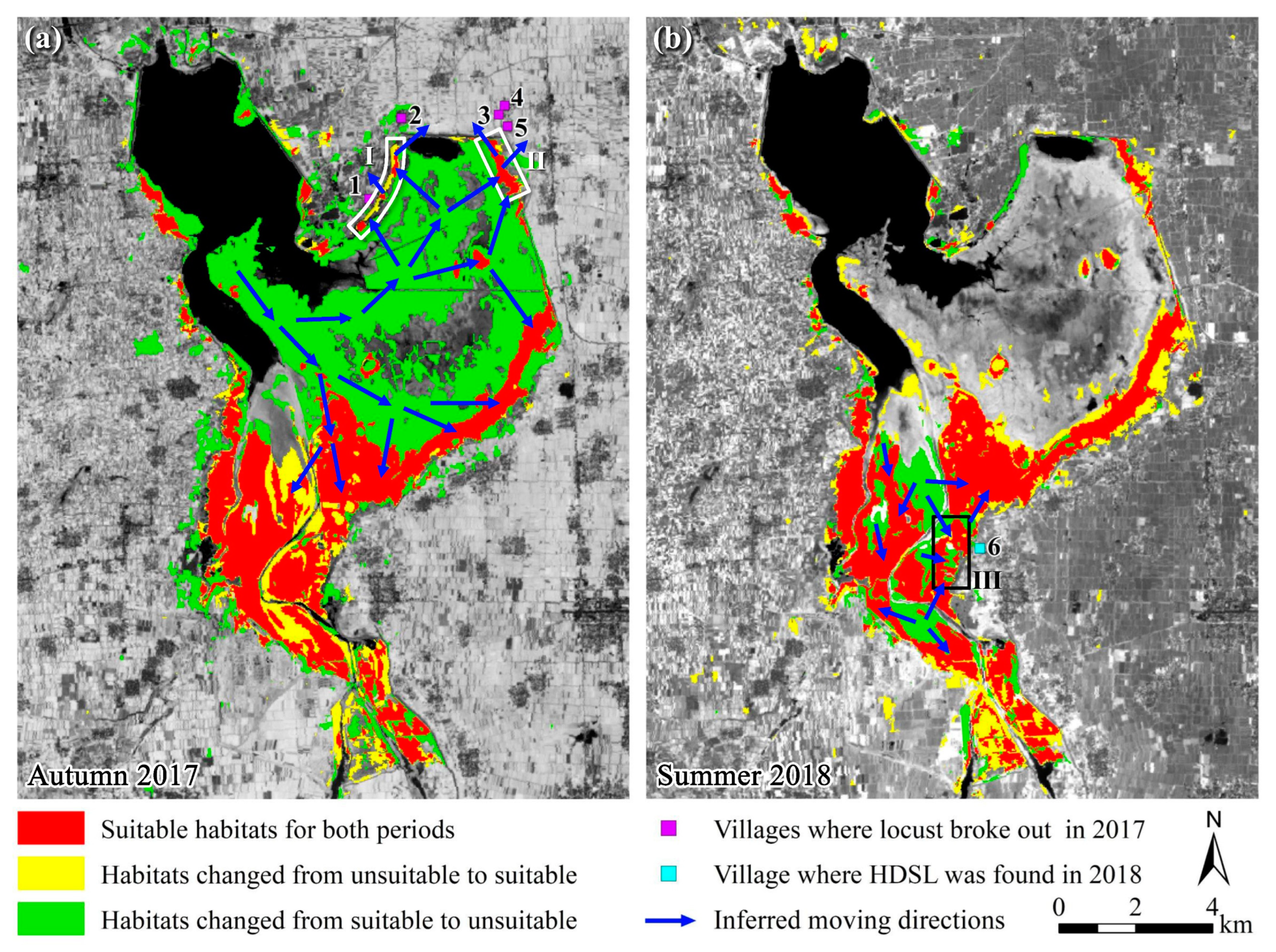
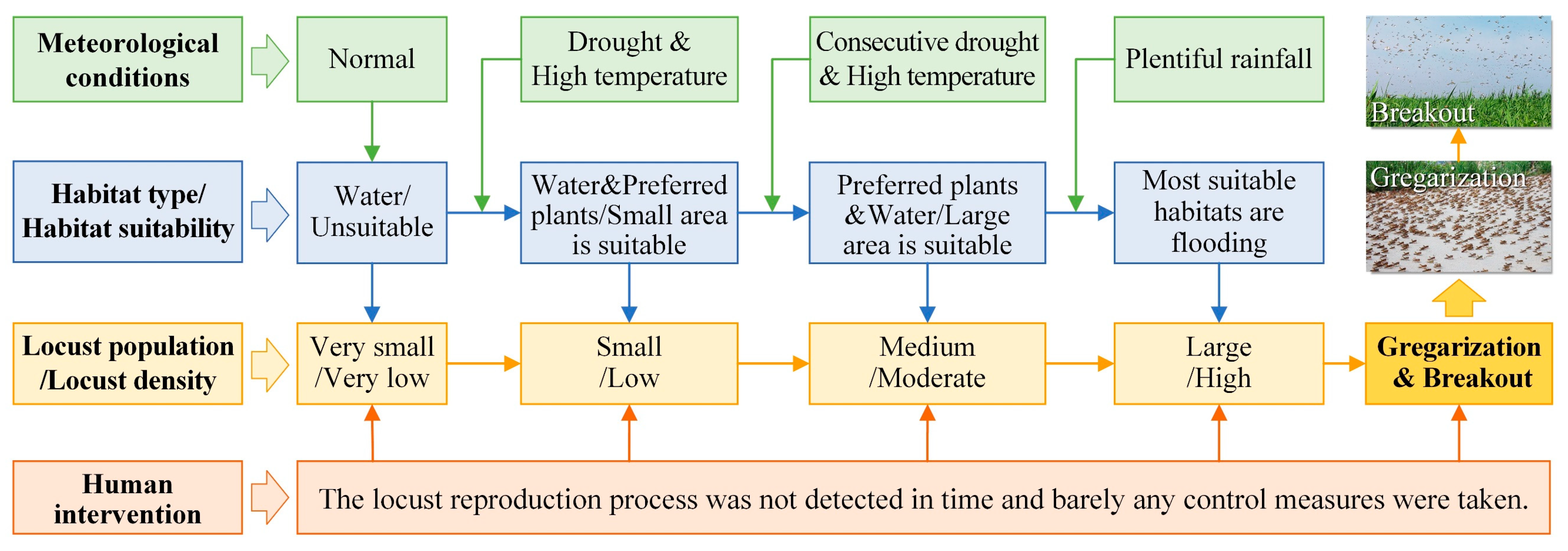
4.1. Outbreak Process of the Locust Plague and the HDSL
- Consecutive drought and high temperature promoted locust reproduction
- 2.
- Flooding-induced habitat compression promoted locust gregarization
4.2. Outbreak Mechanism of Locust Plagues under Drought and Flood Dynamics
4.3. Approach for Early Warning and Identifying Potential High-Risk Locust Areas
- Monitoring drought and flood patterns. Firstly, select water bodies (e.g., rivers, lakes, reservoirs, etc.) and their surrounding areas as the primary focus areas considering the “water attachment” feature of OMLs. Next, assess whether the annual precipitation in the target area over the past 3–5 years has been below the ten-year average (assuming no locusts have been reported in the focus area over the past decade) to detect droughts. If no drought occurs, the locust outbreak risk is low. If a drought is just beginning, continuous meteorological monitoring of the target area is necessary. If a consecutive drought occurs, proceed to the second step. If the target area experiences increased rainfall following a prolonged drought, proceed to the third step.
- Monitoring variations in suitable habitats. Carry out multi-year habitat classification mapping from the year prior to the commencement of a consecutive drought to the current year and analyze the area changes in the suitable habitat. If there is a consistent increase in the suitable habitat area, it suggests that locusts may have completed multi-generation reproduction, potentially leading to the formation of HDSL. In such cases, it is necessary to strengthen field investigations and take measures promptly to reduce the locust population and density, thereby achieving an early warning of the PHRLA and mitigating the risk of locust outbreaks.
- Identifying potential high-risk locust areas. If the target area experiences a sudden increase in rainfall after a prolonged drought and the suitable habitat area shows a trend of an initial increase followed by a decrease, there is a higher risk of a locust plague. First, generate the distribution maps of the suitable habitats before and after rainfall. Then, based on variations in suitable habitat size and distribution and the surrounding topography of water bodies before and after the rainfall, analyze the potential locust movement directions and speculate the possible locations of HDSL. Furthermore, based on the land cover types in the surrounding areas, especially the croplands, identify the PHRLA.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, E.; Chen, Z. Comprehensive Control Technical Manual of Oriental Migratory Locust; China Agriculture Press: Beijing, China, 2010. [Google Scholar]
- Tu, X.; Hu, G.; Fu, X.; Zhang, Y.; Ma, J.; Wang, Y.; Gould, P.J.L.; Du, G.; Su, H.; Zhang, Z.; et al. Mass windborne migrations extend the range of the migratory locust in East China. Agric. For. Entomol. 2020, 22, 41–49. [Google Scholar] [CrossRef]
- Wang, X.; Li, G.; Xu, W.; Kong, D.; Gao, X.; Wang, S.; Feng, C. The locust plagues in the Yangtze River Delta of China during the Ming and Qing Dynasties. Nat. Hazards 2022, 115, 2333–2350. [Google Scholar] [CrossRef]
- Meynard, C.N.; Lecoq, M.; Chapuis, M.P.; Piou, C. On the relative role of climate change and management in the current desert locust outbreak in East Africa. Glob. Chang. Biol. 2020, 26, 3753–3755. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Ma, N.L.; Zhang, D.; Zhou, Q.; Yue, X.; Khoo, S.C.; Yang, H.; Guan, R.; Chen, H.; Zhang, X.; et al. A review of historical and recent locust outbreaks: Links to global warming, food security and mitigation strategies. Environ. Res. 2020, 191, 110046. [Google Scholar] [CrossRef] [PubMed]
- Salih, A.A.M.; Baraibar, M.; Mwangi, K.K.; Artan, G. Climate change and locust outbreak in East Africa. Nat. Clim. Chang. 2020, 10, 584–585. [Google Scholar] [CrossRef]
- Youngblood, J.P.; Cease, A.J.; Talal, S.; Copa, F.; Medina, H.E.; Rojas, J.E.; Trumper, E.V.; Angilletta, M.J.; Harrison, J.F. Climate change expected to improve digestive rate and trigger range expansion in outbreaking locusts. Ecol. Monogr. 2023, 93, e1550. [Google Scholar] [CrossRef]
- Cressman, K. Role of remote sensing in desert locust early warning. J. Appl. Remote Sens. 2013, 7, 75098. [Google Scholar] [CrossRef]
- Zhang, L.; Lecoq, M.; Latchininsky, A.; Hunter, D. Locust and Grasshopper Management. Annu. Rev. Entomol. 2019, 64, 15–34. [Google Scholar] [CrossRef]
- Sun, R.; Huang, W.; Dong, Y.; Zhao, L.; Zhang, B.; Ma, H.; Geng, Y.; Ruan, C.; Xing, N.; Chen, X.; et al. Dynamic forecast of Desert Locust presence using machine learning with a multivariate time lag sliding window technique. Remote Sens. 2022, 14, 747. [Google Scholar] [CrossRef]
- Lomer, C.J.; Bateman, R.P.; Dent, D.; Groote, H.; Douro-Kpindou, O.K.; Kooyman, C.; Langewald, J.; Ouambama, Z.; Peveling, R.; Thomas, M. Development of strategies for the incorporation of biological pesticides into the integrated management of locusts and grasshoppers. Agric. For. Entomol. 2011, 1, 71–88. [Google Scholar] [CrossRef]
- Adriaansen, C.; Woodman, J.D.; Deveson, E.; Drake, V.A. Chapter 4.1—The Australian Plague Locust—Risk and Response. In Biological and Environmental Hazards, Risks, and Disasters; Shroder, J.F., Sivanpillai, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 67–86. [Google Scholar]
- Maute, K.; French, K.; Story, P.; Bull, C.M.; Hose, G.C. Short and long-term impacts of ultra-low-volume pesticide and biopesticide applications for locust control on non-target arid zone arthropods. Agric. Ecosyst. Environ. 2017, 240, 233–243. [Google Scholar] [CrossRef]
- Yang, P.; Ren, B. Promoting the application of green crop pest management technologies—Review on the key issues in the national technical schemes of major crop pest management for 2011 to 2017. Plant Prot. 2018, 44, 6–8. [Google Scholar]
- Yao, X.; Zhu, D.; Yun, W.; Peng, F.; Li, L. A WebGIS-based decision support system for locust prevention and control in China. Comput. Electron. Agric. 2017, 140, 148–158. [Google Scholar] [CrossRef]
- Ji, R.; Xie, B.Y.; Li, D.M.; Li, Z.; Zhang, X. Use of MODIS data to monitor the oriental migratory locust plague. Agric. Ecosyst. Environ. 2004, 104, 615–620. [Google Scholar] [CrossRef]
- Shi, Y.; Huang, W.; Dong, Y.; Peng, D.; Zheng, Q.; Yang, P. The influence of landscape’s dynamics on the Oriental Migratory Locust habitat change based on the time-series satellite data. J. Environ. Manag. 2018, 218, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Klein, I.; Uereyen, S.; Eisfelder, C.; Pankov, V.; Oppelt, N.; Kuenzer, C. Application of geospatial and remote sensing data to support locust management. Int. J. Appl. Earth Obs. 2023, 117, 103212. [Google Scholar] [CrossRef]
- Ellenburg, W.L.; Mishra, V.; Roberts, J.B.; Limaye, A.S.; Case, J.L.; Blankenship, C.B.; Cressman, K. Detecting Desert Locust Breeding Grounds: A Satellite-Assisted Modeling Approach. Remote Sens. 2021, 13, 1276. [Google Scholar] [CrossRef]
- Gómez, D.; Salvador, P.; Sanz, J.; Casanova, C.; Taratiel, D.; Casanova, J.L. Desert locust detection using Earth observation satellite data in Mauritania. J. Arid Environ. 2019, 164, 29–37. [Google Scholar] [CrossRef]
- Moustafa, O.R.M.; Cressman, K. Using the enhanced vegetation index for deriving risk maps of desert locust (Schistocerca gregaria, Forskal) breeding areas in Egypt. J. Appl. Remote Sens. 2014, 8, 84897. [Google Scholar] [CrossRef]
- Waldner, F.O.; Ebbe, M.A.B.; Cressman, K.; Defourny, P. Operational monitoring of the Desert Locust habitat with Earth Observation: An assessment. ISPRS Int. J. Geo-Inf. 2015, 4, 2379–2400. [Google Scholar] [CrossRef]
- Cissé, S.; Ghaout, S.; Ebbe, M.A.B.; Kamara, S.; Piou, C. Field verification of the prediction model on desert locust adult phase status from density and vegetation. J. Insect Sci. 2016, 16, 74. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Zhao, L.; Huang, W.; Dong, Y.; Ma, H.; Guo, A.; Ren, Y.; Xing, N.; Huang, Y.; Sun, R.; et al. A landscape-based habitat suitability model (LHS model) for Oriental Migratory Locust area extraction at large scales: A case study along the Middle and Lower Reaches of the Yellow River. Remote Sens. 2022, 14, 1058. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, W.; Ye, H.; Lu, L. Study on the Identification of Habitat Suitability Areas for the Dominant Locust Species Dasyhippus barbipes in Inner Mongolia. Remote Sens. 2023, 15, 1718. [Google Scholar] [CrossRef]
- Löw, F.; Waldner, F.; Latchininsky, A.; Biradar, C.; Bolkart, M.; Colditz, R.R. Timely monitoring of Asian Migratory locust habitats in the Amudarya delta, Uzbekistan using time series of satellite remote sensing vegetation index. J. Environ. Manag. 2016, 183, 562–575. [Google Scholar] [CrossRef]
- Sun, Z.; Ye, H.; Huang, W.; Qimuge, E.; Bai, H.; Nie, C.; Lu, L.; Qian, B.; Wu, B. Assessment on Potential Suitable Habitats of the Grasshopper Oedaleus decorus asiaticus in North China based on MaxEnt Modeling and Remote Sensing Data. Insects 2023, 14, 138. [Google Scholar] [CrossRef]
- Kimathi, E.; Tonnang, H.E.Z.; Subramanian, S.; Cressman, K.; Abdel-Rahman, E.M.; Tesfayohannes, M.; Niassy, S.; Torto, B.; Dubois, T.; Tanga, C.M.; et al. Prediction of breeding regions for the desert locust Schistocerca gregaria in East Africa. Sci. Rep. 2020, 10, 11937. [Google Scholar] [CrossRef]
- Shroder, J.F.; Sivanpillai, R. Biological and Environmental Hazards, Risks, and Disasters; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Zhang, Z.; Cazelles, B.; Tian, H.; Christian Stige, L.; Bräuning, A.; Stenseth, N.C. Periodic temperature-associated drought/flood drives locust plagues in China. Proc. R. Soc. B Biol. Sci. 2008, 276, 823–831. [Google Scholar] [CrossRef]
- Lin, K.E.; Wang, P.K.; Pai, P.; Lin, Y.; Wang, C. Historical droughts in the Qing dynasty (1644–1911) of China. Clim. Past 2020, 16, 911–931. [Google Scholar] [CrossRef]
- Fei, J.; Zhou, J. The drought and locust plague of 942–944 AD in the Yellow River Basin, China. Quat. Int. 2016, 394, 115–122. [Google Scholar] [CrossRef]
- Li, G.; Kong, D.; Li, F.; Liu, Q.; Wang, H. Reviews and prospects on studies of locust breeding area evolution and drainage network change in China during the historical period. Trop. Geogr. 2017, 37, 226–237. [Google Scholar]
- Xiao, L. Spatial-temporal distribution of locust plague and its relationship with flood/drought in North China during the Qing Dynasty. J. Palaeogeogr. 2018, 20, 1113–1122. (In Chinese) [Google Scholar]
- Yu, G.; Shen, H.; Liu, J. Impacts of climate change on historical locust outbreaks in China. J. Geophys. Res. 2009, 114, D18104. [Google Scholar] [CrossRef]
- Tratalos, J.A.; Cheke, R.A.; Healey, R.G.; Stenseth, N.C. Desert locust populations, rainfall and climate change: Insights from phenomenological models using gridded monthly data. Clim. Res. 2010, 43, 229–239. [Google Scholar] [CrossRef]
- Li, G.; Liu, Q.; Wang, H.; Kong, D.; Yang, X. Spatiotemporal characteristics and environmental response of locust plague in Jiangsu Province during the past one thousand years. Int. J. Nat. Disasters 2015, 24, 66–76. [Google Scholar]
- Huang, B.; Li, G.; Li, F.; Kong, D.; Wang, Y. The 1855 to 1859 locust plague in China. Nat. Hazards 2019, 95, 529–545. [Google Scholar] [CrossRef]
- Wei, Y. Analysis and Research on the Scheme of Reservoir Capacity Increase of Xiashan Reservoir. Master’s Thesis, Shandong University, Jinan, China, 2015. [Google Scholar]
- Tian, L.; Xing, F.; Wang, E.; Wang, X.; Zheng, Z. Changes and distributions of phytoplankton community of Xiashan Reservoir. Water Technol. 2021, 15, 17–21. [Google Scholar]
- Ma, J.; Han, X. Mechanism and Method of Remote Sensing Monitoring of Oriental Migratory Locust Plague; Science Press: Beijing, China, 2004. [Google Scholar]
- Nishide, Y.; Suzuki, T.; Tanaka, S. The hatching time of Locusta migratoria under out door conditions: Role of temperature and adaptive significance. Physiol. Entomol. 2017, 42, 146–155. [Google Scholar] [CrossRef]
- Zhang, L.; Li, H. The impacts of nymph population densities and stadium of Locusta migratoria manilensis (Meyen) on the changes of its social type to the scattered type. Plant Prot. Technol. Ext. 2002, 22, 3–5. [Google Scholar]
- Stige, L.C.; Chan, K.S.; Zhang, Z.; Frank, D.; Stenseth, N.C. Thousand-year-long Chinese time series reveals climatic forcing of decadal locust dynamics. Proc. Natl. Acad. Sci. USA 2007, 104, 16188–16193. [Google Scholar] [CrossRef]
- Crooks, W.T.; Cheke, R.A. Soil moisture assessments for brown locust Locustana pardalina breeding potential using synthetic aperture radar. J. Appl. Remote Sens. 2014, 8, 2378–2391. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, K.; Wang, J.; Jia, H.; Ma, L.; Li, Y.; Duan, J. Assessment of genetic diversity and chemical composition among seven black locust populations from Northern China. Biochem. Syst. Ecol. 2020, 90, 104010. [Google Scholar] [CrossRef]
- Cressman, K. Chapter 4.2—Desert Locust. In Biological and Environmental Hazards, Risks, and Disasters; Shroder, J.F., Sivanpillai, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 87–105. [Google Scholar] [CrossRef]
- Guo, X.; Yu, Q.; Chen, D.; Wei, J.; Yang, P.; Yu, J.; Wang, X.; Kang, L. 4-Vinylanisole is an aggregation pheromone in locusts. Nature 2020, 584, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Huffman, G.J.; Bolvin, D.T.; Nelkin, E.J.; Wolff, D.B.; Adler, R.F.; Gu, G.; Hong, Y.; Bowman, K.P.; Stocker, E.F. The TRMM Multisatellite Precipitation Analysis (TMPA): Quasi-Global, Multiyear, Combined-Sensor Precipitation Estimates at Fine Scales. J. Hydrometeorol. 2007, 8, 38–55. [Google Scholar] [CrossRef]
- Muñoz-Sabater, J.; Dutra, E.; Agustí-Panareda, A.; Albergel, C.; Arduini, G.; Balsamo, G.; Boussetta, S.; Choulga, M.; Harrigan, S.; Hersbach, H.; et al. ERA5-Land: A state-of-the-art global reanalysis dataset for land applications. Earth Syst. Sci. Data 2021, 13, 4349–4383. [Google Scholar] [CrossRef]
- Ma, S. The population dynamics of the Oriental migratory locust (Locust migratoria manilensis Meyen) in China. Acta Entomol. Sin. 1958, 8, 1–40. [Google Scholar]
- Qi, X.; Wang, X.; Xu, H.; Kang, L. Influence of soil moisture on egg cold hardiness in the migratory locust Locusta migratoria (Orthoptera: Acridiidae). Physiol. Entomol. 2007, 32, 219–224. [Google Scholar] [CrossRef]
- Zhu, E. Occurrence and Management of the Oriental Migratory Locust in China; China Agriculture Press: Beijing, China, 1999. [Google Scholar]
- Yang, Q.; Liu, W.; Huang, C.; Zhu, J.; Zhang, Z.; Zhu, J.; Xie, F. Occurring analysis on high-density spot of Locusta migartoria and suggestions on its monitoring and controlling in China in 2017. China Plant Prot. 2018, 38, 37–39+47. [Google Scholar]
- Zha, Y.; Ni, S.; Yang, S. An effective approach to automatically extract urban land-use from TM imagery. J. Remote Sens. 2003, 7, 37–40. [Google Scholar]
- Prasomsup, W.; Piyatadsananon, P.; Aunphoklang, W.; Boonrang, A. Extraction Technic for Built-up Area Classification in Landsat 8 Imagery. Int. J. Environ. Sci. Dev. 2020, 11, 15–20. [Google Scholar] [CrossRef]
- Achanta, R.; Süsstrunk, S. Superpixels and Polygons Using Simple Non-Iterative Clustering. In Proceedings of the 2017 IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Honolulu, HI, USA, 21–26 July 2017; pp. 4895–4904. [Google Scholar]
- Nik, N.; Martono, E.; Putra, N.S.; Suputa. Hatching of migratory locust (Locusta migratoria, L.) (Orthoptera: Acrididae) eggs at several of texture and moisture levels in semi-field laboratory. EurAsian J. Biosci. 2020, 14, 4457–4465. [Google Scholar]
- Woodman, J.D. Surviving a flood: Effects of inundation period, temperature and embryonic development stage in locust eggs. Bull. Entomol. Res. 2015, 105, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Han, X. Study on Remote Sensing Mechanism and Methods for East Asian Migratory Locust Hazard Monitoring. Ph.D. Thesis, Institute of Remote Sensing Application, China Academy of Science, Beijing, China, 2003. [Google Scholar]
- Ji, R.; Li, D.; Xie, B.; Li, Z.; Meng, D. Spatial distribution of oriental migratory locust (Orthoptera: Acrididae) egg pod populations: Implications for site-specific pest management. Environ. Entomol. 2006, 35, 1244–1248. [Google Scholar] [CrossRef]
- Wu, T.; Hao, S.; Kang, L. Effects of Soil Temperature and Moisture on the Development and Survival of Grasshopper Eggs in Inner Mongolian Grasslands. Front. Ecol. Evol. 2021, 9, 727911. [Google Scholar] [CrossRef]
- Latchininsky, A.; Sword, G.; Sergeev, M.; Cigliano, M.; Lecoq, M. Locusts and Grasshoppers: Behavior, Ecology, and Biogeography. Psyche 2011, 2011, 578327. [Google Scholar] [CrossRef]
- Wang, X.; Li, G.; Wang, S.; Feng, C.; Xu, W.; Nie, Q.; Liu, Q. The effect of environmental changes on locust outbreak dynamics in the downstream area of the Yellow River during the Ming and Qing Dynasties. Sci. Total Environ. 2023, 877, 162921. [Google Scholar] [CrossRef]
- Simpson, S.J.; McCaffery, A.R.; Hagele, B.F. A behavioural analysis of phase change in the desert locust. Biol. Rev. 1999, 74, 461–480. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years | Summer | Autumn | ||
|---|---|---|---|---|
| Date | Cloud Coverage | Date | Cloud Coverage | |
| 2014 | 26 May | Cloudless | 22 September | Cloudless |
| 2015 | 5 June | 0.19% | 25 September | Cloudless |
| 2016 | 16 June | Cloudless | 20 September | 0.87% |
| 2017 | 25 May | Cloudless | 23 September | Cloudless |
| 2018 | 6 June | Cloudless | 26 September | 0.08% |
| Habitat Class | Description |
|---|---|
| Cropland | Dry land used for cultivating food crops, such as wheat, corn, millet, sorghum, rice, etc., and vegetable greenhouses used for planting potatoes, cabbages, radish, leeks, and so on. Due to routine tillage activities such as plowing and irrigation, locust eggs laid in cropland usually cannot survive, but the crops could still provide food supply when locusts break out and migrate in. |
| Reed wetland | Annual pure reeds; sometimes the bottom part is submerged by water, with vegetation coverage usually higher than 75%. Because of the high soil moisture content, locusts cannot lay eggs and hatch. And it is also tricky for nymphs to develop due to the high canopy closure. When locusts become adults and flyable, reed wetlands can provide abundant food supplies since the reeds are their favorite plant food. |
| Reeds and weeds | Mixed area of reeds and other gramineous weeds on wet or semi-dry soil, with 15–75% vegetation coverage. The vegetation and soil conditions provide ideal habitats for locust breeding. The low vegetation coverage regions (15–50%) offer perfect places for locust oviposition and nymph development; the high vegetation coverage regions (50–75%) can provide adequate host plants for locust feeding. |
| Woodland | Timber plantations along rivers, reservoirs, and around villages, as well as natural trees along rivers and ornamental trees in urban area parks. This habitat class is unsuitable for locust breeding due to their dietary incompatibility with the prevailing vegetation. |
| Water | Reservoirs, rivers, artificial lakes, and irrigation ditches. Water resources are invariably interconnected with soil moisture and temperature, thereby exerting a subsequent impact on the growth of the surrounding vegetation. Hence, during specific periods, the adjacent surroundings of water resources can furnish conducive habitats for locusts. |
| Artificial Surface | Towns, villages, roads, and other water facilities, such as artificial dams and ditches, which are unsuitable for locust breeding. |
| Others | Bare lands with low vegetation coverage, such as reservoir mudflats and mining areas, which are highly unsuitable for the propagation of locusts. |
| Period | Number of Reference Pixels | |||||||
|---|---|---|---|---|---|---|---|---|
| Cropland | Reed Wetland | Reeds and Weeds | Woodland | Water | Artificial Surface | Others | Total | |
| Summer 2014 | 7452 | 410 | 97 | 23 | 2324 | 5150 | 311 | 15,767 |
| Autumn 2014 | 6772 | 115 | 969 | 120 | 2153 | 5289 | 1464 | 16,882 |
| Summer 2015 | 6637 | 483 | 4453 | 43 | 2633 | 5661 | 1972 | 21,882 |
| Autumn 2015 | 7026 | 266 | 4265 | 57 | 2844 | 5661 | 2172 | 22,291 |
| Summer 2016 | 6319 | 652 | 3948 | 26 | 1412 | 5180 | 2599 | 20,136 |
| Autumn 2016 | 6420 | 1763 | 6469 | 160 | 1166 | 5367 | 489 | 21,834 |
| Summer 2017 | 6991 | 1564 | 4202 | 21 | 733 | 4522 | 1279 | 19,312 |
| Autumn 2017 | 6420 | 1187 | 3192 | 160 | 2035 | 5228 | 704 | 18,926 |
| Summer 2018 | 5822 | 2564 | 4097 | 181 | 1384 | 5238 | 294 | 19,580 |
| Autumn 2018 | 6392 | 18 | 146 | 60 | 3038 | 5198 | 255 | 15,107 |
| Time | PA and UA (%) | Habitat Type | OA (%) | Kappa | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cropland | Reed Wetland | Reeds and Weeds | Woodland | Water | Artificial Surface | Others | ||||
| Summer 2014 | PA | 94.69 | 94.39 | 60.00 | 95.61 | 99.35 | 90.52 | 91.02 | 93.68 | 0.89 |
| UA | 95.24 | 80.80 | 68.57 | 66.46 | 99.89 | 92.61 | 74.51 | |||
| Autumn 2014 | PA | 95.13 | 100.00 | 94.62 | 86.67 | 99.63 | 93.24 | 94.50 | 94.87 | 0.91 |
| UA | 97.82 | 100.00 | 75.69 | 60.47 | 99.63 | 92.80 | 81.13 | |||
| Summer 2015 | PA | 92.34 | 88.55 | 90.08 | 87.10 | 99.85 | 90.73 | 97.64 | 92.59 | 0.89 |
| UA | 95.67 | 86.47 | 81.18 | 81.00 | 99.25 | 90.45 | 88.76 | |||
| Autumn 2015 | PA | 90.65 | 88.24 | 92.93 | 82.31 | 99.35 | 90.07 | 94.57 | 91.58 | 0.88 |
| UA | 96.33 | 60.61 | 88.17 | 67.60 | 99.35 | 87.65 | 79.20 | |||
| Summer 2016 | PA | 92.82 | 99.46 | 91.03 | 92.13 | 99.20 | 95.23 | 93.65 | 93.82 | 0.90 |
| UA | 96.29 | 98.40 | 82.63 | 60.29 | 99.60 | 94.81 | 85.82 | |||
| Autumn 2016 | PA | 92.86 | 98.30 | 93.66 | 97.62 | 100.00 | 93.10 | 90.51 | 93.57 | 0.90 |
| UA | 96.52 | 96.65 | 87.86 | 72.35 | 99.65 | 92.32 | 64.41 | |||
| Summer 2017 | PA | 91.52 | 98.59 | 95.63 | 94.81 | 99.17 | 92.02 | 93.14 | 93.09 | 0.90 |
| UA | 95.33 | 97.42 | 96.32 | 68.22 | 98.36 | 90.23 | 79.50 | |||
| Autumn 2017 | PA | 95.23 | 97.61 | 94.55 | 90.98 | 100.00 | 92.75 | 91.06 | 94.85 | 0.91 |
| UA | 97.07 | 99.19 | 93.57 | 76.10 | 99.51 | 90.86 | 75.93 | |||
| Summer 2018 | PA | 94.73 | 95.69 | 95.73 | 94.94 | 99.52 | 93.40 | 95.29 | 94.80 | 0.91 |
| UA | 97.57 | 99.05 | 90.78 | 86.67 | 97.47 | 89.28 | 68.94 | |||
| Autumn 2018 | PA | 94.42 | 100.00 | 88.89 | 87.93 | 99.66 | 94.32 | 92.09 | 94.81 | 0.90 |
| UA | 98.15 | 50.00 | 74.16 | 75.00 | 99.15 | 90.41 | 71.51 | |||
 | ||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, L.; Li, H.; Huang, W.; Dong, Y.; Geng, Y.; Ma, H.; Chen, J. Outbreak Mechanism of Locust Plagues under Dynamic Drought and Flood Environments Based on Time Series Remote Sensing Data: Implication for Identifying Potential High-Risk Locust Areas. Remote Sens. 2023, 15, 5206. https://doi.org/10.3390/rs15215206
Zhao L, Li H, Huang W, Dong Y, Geng Y, Ma H, Chen J. Outbreak Mechanism of Locust Plagues under Dynamic Drought and Flood Environments Based on Time Series Remote Sensing Data: Implication for Identifying Potential High-Risk Locust Areas. Remote Sensing. 2023; 15(21):5206. https://doi.org/10.3390/rs15215206
Chicago/Turabian StyleZhao, Longlong, Hongzhong Li, Wenjiang Huang, Yingying Dong, Yun Geng, Huiqin Ma, and Jinsong Chen. 2023. "Outbreak Mechanism of Locust Plagues under Dynamic Drought and Flood Environments Based on Time Series Remote Sensing Data: Implication for Identifying Potential High-Risk Locust Areas" Remote Sensing 15, no. 21: 5206. https://doi.org/10.3390/rs15215206
APA StyleZhao, L., Li, H., Huang, W., Dong, Y., Geng, Y., Ma, H., & Chen, J. (2023). Outbreak Mechanism of Locust Plagues under Dynamic Drought and Flood Environments Based on Time Series Remote Sensing Data: Implication for Identifying Potential High-Risk Locust Areas. Remote Sensing, 15(21), 5206. https://doi.org/10.3390/rs15215206