

Spatial Analysis of Intra-Annual Reed Ecosystem Dynamics at Lake Neusiedl Using RGB Drone Imagery and Deep Learning

Abstract
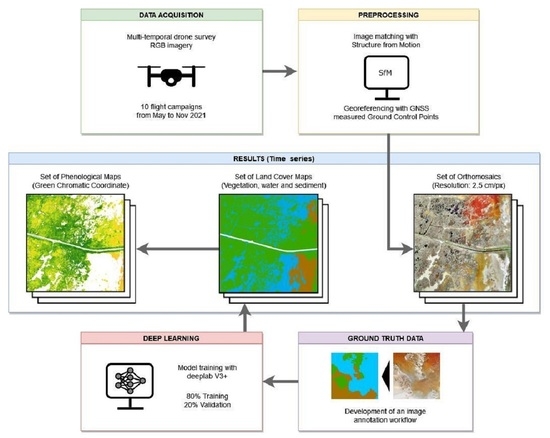
1. Introduction
- To provide a detailed workflow for land cover classification of high-resolution orthoimagery using deep learning image segmentation.
- To reveal the spatial and temporal variability of a reed ecosystem under intensive drought conditions.
- To disclose the phenological stages of Phragmites australis in the reed stands of Lake Neusiedl over the 2021 vegetation period.
2. Materials and Methods
2.1. Study Area
2.2. Data Preparation
2.2.1. UAV Data Acquisition
2.2.2. Photogrammetric Preprocessing
2.3. Semantic Segmentation
2.3.1. Image Annotation
- (a)
- The eight carefully selected ground truth areas were clipped from the orthomosaic and resampled to a resolution of 10 cm using the bilinear resampling technique. Resizing of the imagery was done to reduce processing time for the next step.
- (b)
- The resampled imagery was segmented into groups of adjacent pixels with similar spectral characteristics. The segmentation tool within the software can be used to define the spectral and spatial detail. It was found that the best segmentation results for distinguishing between the classes in our data were obtained when the spectral detail was set between 15 and 17 and the spatial detail set to 1. Furthermore, the minimum segment size was set to 500 pixels. With these parameter settings, while the segmentation algorithm produced fragmented patches that were smaller then required, it provided proper delineation between land cover classes along certain segment boundaries.
- (c)
- The raster output from the segmentation tool was vectorized for the next processing step.
- (d)
- Segments from the same semantic classes were manually merged. Merging was performed by visual interpretation of the polygons while overlaying the ground truth imagery. To speed up this process, segments smaller than 1 m² were removed and the resulting gaps were filled by adding the geometry of the gap to the adjacent polygon with which it shared the longest edge. This step provided fully overlapping semantic class polygons, with each polygon covering the entire area of a class along with its various intra-class variations contained in the ground truth imagery.
- (e)
- As a last step, the polygons were assigned to the corresponding land cover class in order to obtain a fully labeled polygon layer.
2.3.2. Deep Learning
2.4. Phenological Analysis
Phenocam Data
3. Results
3.1. Performance Metrics
3.2. Land Cover Classification
3.3. Vegetation Index
4. Discussion
4.1. Model Prediction
4.2. Spatio-Temporal Variability
4.3. Vegetation Phenology
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APOS | Austrian Positioning Service |
| CNN | Convolutional Neural Network |
| DOY | Day of the Year |
| EC | Eddy Covariance |
| FN | False Negative |
| FP | False Positive |
| GCC | Green Chromatic Coordinates |
| GCP | Ground Control Points |
| GNSS | Global Navigation Satellite System |
| GPU | Graphics Processing Unit |
| GSD | Ground Sampling Distance |
| IoU | Intersection over Union |
| LCC | Land Cover Classification |
| LTER | Long-Term Ecological Research |
| OA | Overall Accuracy |
| OBIA | Object-Based Image Analysis |
| RGB | Red–Green–Blue |
| RMSE | Root Mean Square Error |
| ROI | Region of Interest |
| RTK | Real-Time Kinematics |
| SD | Standard Deviation |
| SfM | Structure from Motion |
| TP | True Positive |
| UAV | Unmanned Aerial Vehicle |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Precision | Recall | F1-Score | IoU | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Orthomosaic Date | Image Chips Used for Training | Learning Rate | Accuracy of Last Epoch | Vegetation | Water | Sediment | Vegetation | Water | Sediment | Vegetation | Water | Sediment | Vegetation | Water | Sediment |
| 2021-05-05 | 1621 | 0.0004 | 0.945 | 0.958 | 0.967 | 0.892 | 0.941 | 0.946 | 0.952 | 0.950 | 0.956 | 0.921 | 0.851 | 0.837 | 0.629 |
| 2021-05-26 | 1642 | 0.0010 | 0.948 | 0.972 | 0.918 | 0.927 | 0.964 | 0.945 | 0.908 | 0.968 | 0.931 | 0.918 | 0.920 | 0.847 | 0.575 |
| 2021-06-18 | 1537 | 0.0010 | 0.958 | 0.977 | 0.959 | 0.931 | 0.965 | 0.917 | 0.975 | 0.971 | 0.938 | 0.953 | 0.924 | 0.731 | 0.784 |
| 2021-07-07 | 1574 | 0.0005 | 0.935 | 0.960 | 0.936 | 0.868 | 0.955 | 0.939 | 0.899 | 0.958 | 0.938 | 0.883 | 0.910 | 0.766 | 0.662 |
| 2021-07-27 | 1696 | 0.0008 | 0.939 | 0.948 | 0.878 | 0.944 | 0.957 | 0.912 | 0.924 | 0.952 | 0.895 | 0.934 | 0.887 | 0.476 | 0.801 |
| 2021-08-18 | 1642 | 0.0008 | 0.936 | 0.964 | 0.908 | 0.909 | 0.951 | 0.867 | 0.955 | 0.958 | 0.887 | 0.932 | 0.905 | 0.639 | 0.797 |
| 2021-09-10 | 1533 | 0.0005 | 0.950 | 0.951 | 0.952 | 0.947 | 0.964 | 0.906 | 0.951 | 0.958 | 0.928 | 0.949 | 0.890 | 0.603 | 0.835 |
| 2021-09-29 | 1706 | 0.0005 | 0.937 | 0.952 | 0.904 | 0.931 | 0.962 | 0.915 | 0.906 | 0.957 | 0.909 | 0.918 | 0.906 | 0.543 | 0.747 |
| 2021-10-20 | 1720 | 0.0005 | 0.939 | 0.958 | 0.923 | 0.906 | 0.953 | 0.953 | 0.899 | 0.955 | 0.938 | 0.903 | 0.904 | 0.689 | 0.680 |
| 2021-11-06 | 1741 | 0.0002 | 0.939 | 0.945 | 0.940 | 0.922 | 0.961 | 0.927 | 0.899 | 0.953 | 0.933 | 0.910 | 0.892 | 0.723 | 0.693 |
| Mean | 1641 | 0.0006 | 0.943 | 0.958 | 0.929 | 0.918 | 0.957 | 0.923 | 0.927 | 0.958 | 0.925 | 0.922 | 0.899 | 0.685 | 0.720 |
| SD | 71 | 0.0003 | 0.007 | 0.010 | 0.026 | 0.023 | 0.007 | 0.024 | 0.027 | 0.006 | 0.020 | 0.020 | 0.020 | 0.115 | 0.081 |
Appendix B


References
- Mitsch, W.J.; Bernal, B.; Hernandez, M.E. Ecosystem services of wetlands. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2015, 11, 1–4. [Google Scholar] [CrossRef]
- Čížková, H.; Kučera, T.; Poulin, B.; Květ, J. Ecological Basis of Ecosystem Services and Management of Wetlands Dominated by Common Reed (Phragmites australis): European Perspective. Diversity 2023, 15, 629. [Google Scholar] [CrossRef]
- Kallasvuo, M.; Lappalainen, A.; Urho, L. Coastal reed belts as fish reproduction habitats. Boreal Environ. Res. 2011, 16, 1–14. [Google Scholar]
- Yu, G.A.; Li, Z.; Yang, H.; Lu, J.; Huang, H.Q.; Yi, Y. Effects of riparian plant roots on the unconsolidated bank stability of meandering channels in the Tarim River, China. Geomorphology 2020, 351, 106958. [Google Scholar] [CrossRef]
- Nemeth, E.; Dvorak, M. Reed die-back and conservation of small reed birds at Lake Neusiedl, Austria. J. Ornithol. 2022, 163, 683–693. [Google Scholar] [CrossRef]
- Mirtl, M.; Bahn, M.; Battin, T.; Borsdorf, A.; Dirnböck, T.; Englisch, M.; Erschbamer, B.; Fuchsberger, J.; Gaube, V.; Grabherr, G.; et al. Research for the Future LTER-Austria White Paper—On the Status and Orientation of Process Oriented Ecosystem Research, Biodiversity and Conservation Research and Socio-Ecological Research in Austria; Technical Report; LTER-Austria Series; LTER: Vienna, Austria, 2015; Volume 2, ISBN 978-3-9503986-1-8. Available online: https://www.lter-austria.at (accessed on 5 August 2023).
- Díaz-Delgado, R.; Cazacu, C.; Adamescu, M. Rapid Assessment of Ecological Integrity for LTER Wetland Sites by Using UAV Multispectral Mapping. Drones 2019, 3, 3. [Google Scholar] [CrossRef]
- Csaplovics, E. Interpretation von Farbinfrarotbildern: Kartierung von Vegetationsschäden in Brixlegg; Schilfkartierung Neusiedler See. Geowiss. Mitteilungen 1982, 23, 178. [Google Scholar]
- Csaplovics, E.; Schmidt, J. Schilfkartierung Neusiedler See: Ausdehnung und Struktur der Schilfbestände des Neusiedler Sees—Projektmanagement, Erfassung und Kartierung des österreichischen Anteiles durch Luftbildklassifikation. 2011. Available online: https://naturschutzbund.at/files/bgl_homepage/projekte/lebensraeume/anlagen/AbschlussberichtSchilfkartierung.pdf (accessed on 5 August 2023).
- Nemeth, E.; Dvorak, M.; Knoll, T.; Kohler, B.; Mühlbacher, S.; Werba, F. Managementplan für den Neusiedler See als Teil des Europaschutzgebiets Neusiedler See—Nordöstliches Leithagebirge. 2014. Available online: https://www.burgenland.at/fileadmin/user_upload/Natura2000_MP__NeusiedlerSee_web.pdf (accessed on 5 August 2023).
- Csaplovics, E.; Nemeth, E. Airborne Optical Imaging in Support of Habitat Ecological Monitoring of the Austrian Reed Belt of Lake Neusiedl. In Proceedings of the GISScience RSGIS4HQ, Vienna, Austria, 23–25 September 2014; pp. 163–167. Available online: https://rsgis4hq.geo.tuwien.ac.at/fileadmin/editors/RSGIS4HQ/proceedings/RSGIS4HQ_Csaplovics.pdf (accessed on 5 August 2023).
- Márkus, I.; Király, G. A Fertő magyarországi nádasainak minősítése, osztályozása és térképezése (Qualification, classification and mapping of the reed stands of the Hungarian part of Lake Fertő). In Monografikus Tanulmányok a Fertő és a Hanság vidékéről. (Monographic Studies of Lake Fertő and Hanság Area); Fertô–Hanság Nemzeti Park Igazgatóság, Szaktudás Kiadó Ház: Budapest, Hungary, 2012; pp. 73–81. [Google Scholar]
- Dinka, M.; Ágoston-Szabó, E.; Szeglet, P. Comparison between biomass and C, N, P, S contents of vigorous and die-back reed stands of Lake Fertő/Neusiedler See. Biologia 2010, 65, 237–247. [Google Scholar] [CrossRef]
- Hackl, P.; Ledolter, J. A Statistical Analysis of the Water Levels at Lake Neusiedl. Austrian J. Stat. 2023, 52, 87–100. [Google Scholar] [CrossRef]
- Wolfram, G.; Herzig, A. Nährstoffbilanz Neusiedler See. Wien. Mitteilungen 2013, 228, 317–338. [Google Scholar]
- Hammer, U.T. Saline Lake Ecosystems of the World; Junk: Dordrecht, The Netherlands, 1986; p. 616. [Google Scholar]
- Soja, G.; Züger, J.; Knoflacher, M.; Kinner, P.; Soja, A.M. Climate impacts on water balance of a shallow steppe lake in Eastern Austria (Lake Neusiedl). J. Hydrol. 2013, 480, 115–124. [Google Scholar] [CrossRef]
- Steiner, G.M.; Englmaier, P.; Fink, M.; Grünweis, F.; Höfner, I.; Korner, I.; Ströhle, A.; Wolf, W. Mittelpunkte der Moorflächen aus dem Moorschutzkatalog (Steiner et al. 1992), 1992. Bundesministerium für Gesundheit und Umweltschutz, Wien. Available online: https://www.data.gv.at/katalog/de/dataset/moorschutzkatalog_1992_points (accessed on 5 August 2023).
- DJI. Mavic 2 Technische Daten. Available online: https://www.dji.com/at/mavic-2/info (accessed on 25 May 2023).
- Trimble R1—Datasheet, Model 2 GNSS SYSTEMS. Available online: https://geospatial.trimble.com/sites/geospatial.trimble.com/files/2021-07/022516-332B_TrimbleR10-2_DS_USL_0721_LR.pdf (accessed on 25 May 2023).
- Villanueva, J.K.S.; Blanco, A.C. Optimization of ground control point (GCP) configuration for unmanned aerial vehicle (UAV) survey using structure from motion (SfM). Int. Arch. Photogramm. Remote. Sens. Spat. Inf. Sci. 2019, XLII-4/W12, 167–174. [Google Scholar] [CrossRef]
- BEV. Bundesamt für Eich- und Vermessungswesen. 2019. Available online: https://www.bev.gv.at/ (accessed on 25 May 2023).
- Westoby, M.J.; Brasington, J.; Glasser, N.F.; Hambrey, M.J.; Reynolds, J.M. ‘Structure-from-Motion’ photogrammetry: A low-cost, effective tool for geoscience applications. Geomorphology 2012, 179, 300–314. [Google Scholar] [CrossRef]
- Deliry, S.I.; Avdan, U. Accuracy of Unmanned Aerial Systems Photogrammetry and Structure from Motion in Surveying and Mapping: A Review. J. Indian Soc. Remote Sens. 2021, 49, 1997–2017. [Google Scholar] [CrossRef]
- Pix4D Documentation. 2023. Available online: https://support.pix4d.com/hc/en-us/sections/360003718571-How-to-step-by-step-instructions (accessed on 25 May 2023).
- Shen, F.; Zeng, G. Semantic image segmentation via guidance of image classification. Neurocomputing 2019, 330, 259–266. [Google Scholar] [CrossRef]
- ESRI Inc. ArcGIS Pro, Version 3.0.3; ESRI Inc.: Redlands, CA, USA, 2022.
- ESRI. Deep Learning Libraries for ArcGIS Pro 3.0.3. 2022. Available online: https://github.com/Esri/deep-learning-frameworks.git (accessed on 25 May 2023).
- Tong, X.Y.; Xia, G.S.; Lu, Q.; Shen, H.; Li, S.; You, S.; Zhang, L. Land-cover classification with high-resolution remote sensing images using transferable deep models. Remote Sens. Environ. 2020, 237, 111322. [Google Scholar] [CrossRef]
- Zhang, X.; Han, L.; Han, L.; Zhu, L. How Well Do Deep Learning-Based Methods for Land Cover Classification and Object Detection Perform on High Resolution Remote Sensing Imagery? Remote Sens. 2020, 12, 417. [Google Scholar] [CrossRef]
- Volpi, M.; Tuia, D. Dense Semantic Labeling of Subdecimeter Resolution Images With Convolutional Neural Networks. IEEE Trans. Geosci. Remote Sens. 2017, 55, 881–893. [Google Scholar] [CrossRef]
- Chen, L.C.; Zhu, Y.; Papandreou, G.; Schroff, F.; Adam, H. Encoder-Decoder with Atrous Separable Convolution for Semantic Image Segmentation. arXiv 2018, arXiv:1802.02611. [Google Scholar] [CrossRef]
- Zhao, W.; Du, S. Learning multiscale and deep representations for classifying remotely sensed imagery. ISPRS J. Photogramm. Remote Sens. 2016, 113, 155–165. [Google Scholar] [CrossRef]
- Pashaei, M.; Kamangir, H.; Starek, M.J.; Tissot, P. Review and Evaluation of Deep Learning Architectures for Efficient Land Cover Mapping with UAS Hyper-Spatial Imagery: A Case Study Over a Wetland. Remote Sens. 2020, 12, 959. [Google Scholar] [CrossRef]
- Rahman, M.A.; Wang, Y. Optimizing Intersection-Over-Union in Deep Neural Networks for Image Segmentation. In Advances in Visual Computing; Bebis, G., Boyle, R., Parvin, B., Koracin, D., Porikli, F., Skaff, S., Entezari, A., Min, J., Iwai, D., Sadagic, A., et al., Eds.; Series Title: Lecture Notes in Computer Science; Springer International Publishing: Cham, Switzerland, 2016; Volume 10072, pp. 234–244. [Google Scholar] [CrossRef]
- Zheng, J.Y.; Hao, Y.Y.; Wang, Y.C.; Zhou, S.Q.; Wu, W.B.; Yuan, Q.; Gao, Y.; Guo, H.Q.; Cai, X.X.; Zhao, B. Coastal Wetland Vegetation Classification Using Pixel-Based, Object-Based and Deep Learning Methods Based on RGB-UAV. Land 2022, 11, 2039. [Google Scholar] [CrossRef]
- Anagnostis, A.; Tagarakis, A.C.; Kateris, D.; Moysiadis, V.; Sørensen, C.G.; Pearson, S.; Bochtis, D. Orchard Mapping with Deep Learning Semantic Segmentation. Sensors 2021, 21, 3813. [Google Scholar] [CrossRef] [PubMed]
- ESRI. ArcGIS API for Python—Arcgis.learn Module. 2022. Available online: https://developers.arcgis.com/python/api-reference/arcgis.learn.toc.html#deeplab (accessed on 25 May 2023).
- Sonnentag, O.; Hufkens, K.; Teshera-Sterne, C.; Young, A.M.; Friedl, M.; Braswell, B.H.; Milliman, T.; O’Keefe, J.; Richardson, A.D. Digital repeat photography for phenological research in forest ecosystems. Agric. For. Meteorol. 2012, 152, 159–177. [Google Scholar] [CrossRef]
- Filippa, G.; Cremonese, E.; Migliavacca, M.; Galvagno, M.; Folker, M.; Richardson, A.D.; Tomelleri, E. Package ‘Phenopix’: Process Digital Images of a Vegetation Cover; R Package Version 2.4.2; R Core Team: Vienna, Austria, 2020; Available online: https://cran.rstudio.com/web/packages/phenopix/phenopix.pdf (accessed on 25 May 2023).
- Filippa, G.; Cremonese, E.; Migliavacca, M.; Galvagno, M.; Forkel, M.; Wingate, L.; Tomelleri, E.; Di Morra Cella, U.; Richardson, A.D. Phenopix: A R package for image-based vegetation phenology. Agric. For. Meteorol. 2016, 220, 141–150. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing: R Foundation for Statistical Computing; Version 4.2.2; R Core Team: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 25 May 2023).
- Dowle, M.; Srinivasan, A. Data.table: Extension of ‘Data.Frame’; R Package Version 1.14.8; R Core Team: Vienna, Austria, 2023; Available online: https://CRAN.R-project.org/package=data.table (accessed on 25 May 2023).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 25 May 2023).
- Al-Najjar, H.A.H.; Kalantar, B.; Pradhan, B.; Saeidi, V.; Halin, A.A.; Ueda, N.; Mansor, S. Land Cover Classification from fused DSM and UAV Images Using Convolutional Neural Networks. Remote Sens. 2019, 11, 1461. [Google Scholar] [CrossRef]
- Bhatnagar, S.; Gill, L.; Ghosh, B. Drone Image Segmentation Using Machine and Deep Learning for Mapping Raised Bog Vegetation Communities. Remote Sens. 2020, 12, 2602. [Google Scholar] [CrossRef]
- Meneses, N.C.; Brunner, F.; Baier, S.; Geist, J.; Schneider, T. Quantification of Extent, Density, and Status of Aquatic Reed Beds Using Point Clouds Derived from UAV–RGB Imagery. Remote Sens. 2018, 10, 1869. [Google Scholar] [CrossRef]
- Meneses, N.C.; Baier, S.; Reidelstürz, P.; Geist, J.; Schneider, T. Modelling heights of sparse aquatic reed (Phragmites australis) using Structure from Motion point clouds derived from Rotary- and Fixed-Wing Unmanned Aerial Vehicle (UAV) data. Limnologica 2018, 72, 10–21. [Google Scholar] [CrossRef]
- Villa, P.; Laini, A.; Bresciani, M.; Bolpagni, R. A remote sensing approach to monitor the conservation status of lacustrine Phragmites australis beds. Wetl. Ecol. Manag. 2013, 21, 399–416. [Google Scholar] [CrossRef]
- Davranche, A.; Poulin, B.; Lefebvre, G. Reedbed monitoring using classification trees and SPOT-5 seasonal time series. In Proceedings of the International Symposium on Advanced Methods of Monitoring Reed Habitats in Europe, Illmitz, Austria, 25–26 November 2010; pp. 15–29. Available online: https://hal.science/hal-00692542 (accessed on 5 August 2023).
- Zlinszky, A. Mapping and conservation of the reed wetlands on Lake Balaton. Ph.D. Thesis, Eötvös Loránd University, Budapest, Hungary, 2013. [Google Scholar]
- Dienst, M.; Schmieder, K.; Ostendorp, W. Dynamik der Schilfröhrichte am Bodensee unter dem Einfluss von Wasserstandsvariationen (Effects of water level variations on the dynamics of the reed belts of Lake Constance). Limnologica 2004, 34, 29–36. [Google Scholar] [CrossRef]
- Sun, C.; Li, J.; Liu, Y.; Zhao, S.; Zheng, J.; Zhang, S. Tracking annual changes in the distribution and composition of saltmarsh vegetation on the Jiangsu coast of China using Landsat time series–based phenological parameters. Remote Sens. Environ. 2023, 284, 113370. [Google Scholar] [CrossRef]
- Tiškus, E.; Vaičiūtė, D.; Bučas, M.; Gintauskas, J. Evaluation of common reed ( Phragmites australis ) bed changes in the context of management using earth observation and automatic threshold. Eur. J. Remote Sens. 2023, 56, 2161070. [Google Scholar] [CrossRef]
- van Iersel, W.; Straatsma, M.; Addink, E.; Middelkoop, H. Monitoring height and greenness of non-woody floodplain vegetation with UAV time series. ISPRS J. Photogramm. Remote Sens. 2018, 141, 112–123. [Google Scholar] [CrossRef]
- Higgisson, W.; Cobb, A.; Tschierschke, A.; Dyer, F. Estimating the cover of Phragmites australis using unmanned aerial vehicles and neural networks in a semi–arid wetland. River Res. Appl. 2021, 37, 1312–1322. [Google Scholar] [CrossRef]
- Nechwatal, J.; Wielgoss, A.; Mendgen, K. Flooding events and rising water temperatures increase the significance of the reed pathogen Pythium phragmitis as a contributing factor in the decline of Phragmites australis. Hydrobiologia 2008, 613, 109–115. [Google Scholar] [CrossRef]
- Ostendorp, W. ‘Die-back’ of reeds in Europe—A critical review of literature. Aquat. Bot. 1989, 35, 5–26. [Google Scholar] [CrossRef]
- Lissner, J.; Schierup, H.H. Effects of salinity on the growth of Phragmites australis. Aquat. Bot. 1997, 55, 247–260. [Google Scholar] [CrossRef]
- Wasserportal Burgenland. Niederschlag: Illmitz-Biologische Station. 2023. Available online: https://wasser.bgld.gv.at/hydrographie/der-niederschlag/illmitz-biologische-station (accessed on 25 May 2023).
- Koch, M.; Koebsch, F.; Hahn, J.; Jurasinski, G. From meadow to shallow lake: Monitoring secondary succession in a coastal fen after rewetting by flooding based on aerial imagery and plot data. Mires Peat 2017, 19, 1. [Google Scholar] [CrossRef]
- Beyer, F.; Jansen, F.; Jurasinski, G.; Koch, M.; Schröder, B.; Koebsch, F. Drought years in peatland rewetting: Rapid vegetation succession can maintain the net CO2 sink function. Biogeosciences 2021, 18, 917–935. [Google Scholar] [CrossRef]
- Fukarek, F.; Henker, H.; Berg, C. Flora von Mecklenburg-Vorpommern: Farn-und Blütenpflanzen; Weissdorn-Verlag: Jena, Germany, 2006. [Google Scholar]
- Van Diggelen, R.; Middleton, B.; Bakker, J.; Grootjans, A.; Wassen, M. Fens and floodplains of the temperate zone: Present status, threats, conservation and restoration. Appl. Veg. Sci. 2006, 9, 157–162. [Google Scholar] [CrossRef]
- Klimkowska, A.; Van Diggelen, R.; Grootjans, A.P.; Kotowski, W. Prospects for fen meadow restoration on severely degraded fens. Perspect. Plant Ecol. Evol. Syst. 2010, 12, 245–255. [Google Scholar] [CrossRef]







| Flight Campaign | Average GSD 1 [cm] | Drone Survey Area [km²] | RMSE Georeferencing (N = 21) | RMSE Check Points (N = 6) | ||
|---|---|---|---|---|---|---|
| X Error [cm] | Y Error [cm] | X Error [cm] | Y Error [cm] | |||
| 2021-05-05 | 2.43 | 0.84 | 4.6 | 2.9 | 2.0 | 5.9 |
| 2021-05-26 | 2.39 | 0.92 | 23.9 | 11.7 | 3.2 | 3.6 |
| 2021-06-18 | 2.50 | 1.02 | 11.6 | 4.9 | 1.9 | 4.4 |
| 2021-07-07 | 2.49 | 0.99 | 24.2 | 8.9 | 10.7 | 5.0 |
| 2021-07-27 | 2.45 | 0.99 | 14.3 | 3.0 | 1.7 | 4.9 |
| 2021-08-18 | 2.41 | 0.99 | 15.5 | 10.2 | 3.9 | 4.2 |
| 2021-09-10 | 2.45 | 0.95 | 6.8 | 3.1 | 4.5 | 5.8 |
| 2021-09-29 | 2.36 | 0.97 | 5.2 | 3.8 | 3.8 | 5.2 |
| 2021-10-20 | 2.36 | 0.94 | 8.3 | 28.7 | 2.7 | 6.0 |
| 2021-11-06 | 2.32 | 1.00 | 3.0 | 1.4 | 3.2 | 5.6 |
| Average | 2.42 | 0.96 | 11.7 | 7.9 | 3.8 | 5.1 |
| Class | Precision | Recall | F1-Score | IoU 3 |
|---|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | |
| Vegetation | 0.958 ± 0.010 | 0.957 ± 0.007 | 0.958 ± 0.006 | 0.899 ± 0.020 |
| Water | 0.929 ± 0.026 | 0.923 ± 0.024 | 0.925 ± 0.020 | 0.685 ± 0.115 |
| Sediment | 0.918 ± 0.023 | 0.927 ± 0.027 | 0.922 ± 0.020 | 0.720 ± 0.081 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buchsteiner, C.; Baur, P.A.; Glatzel, S. Spatial Analysis of Intra-Annual Reed Ecosystem Dynamics at Lake Neusiedl Using RGB Drone Imagery and Deep Learning. Remote Sens. 2023, 15, 3961. https://doi.org/10.3390/rs15163961
Buchsteiner C, Baur PA, Glatzel S. Spatial Analysis of Intra-Annual Reed Ecosystem Dynamics at Lake Neusiedl Using RGB Drone Imagery and Deep Learning. Remote Sensing. 2023; 15(16):3961. https://doi.org/10.3390/rs15163961
Chicago/Turabian StyleBuchsteiner, Claudia, Pamela Alessandra Baur, and Stephan Glatzel. 2023. "Spatial Analysis of Intra-Annual Reed Ecosystem Dynamics at Lake Neusiedl Using RGB Drone Imagery and Deep Learning" Remote Sensing 15, no. 16: 3961. https://doi.org/10.3390/rs15163961
APA StyleBuchsteiner, C., Baur, P. A., & Glatzel, S. (2023). Spatial Analysis of Intra-Annual Reed Ecosystem Dynamics at Lake Neusiedl Using RGB Drone Imagery and Deep Learning. Remote Sensing, 15(16), 3961. https://doi.org/10.3390/rs15163961