



MODIS Time Series Reveal New Maximum Records of Defoliated Area by Ormiscodes amphimone in Deciduous Nothofagus Forests, Southern Chile
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
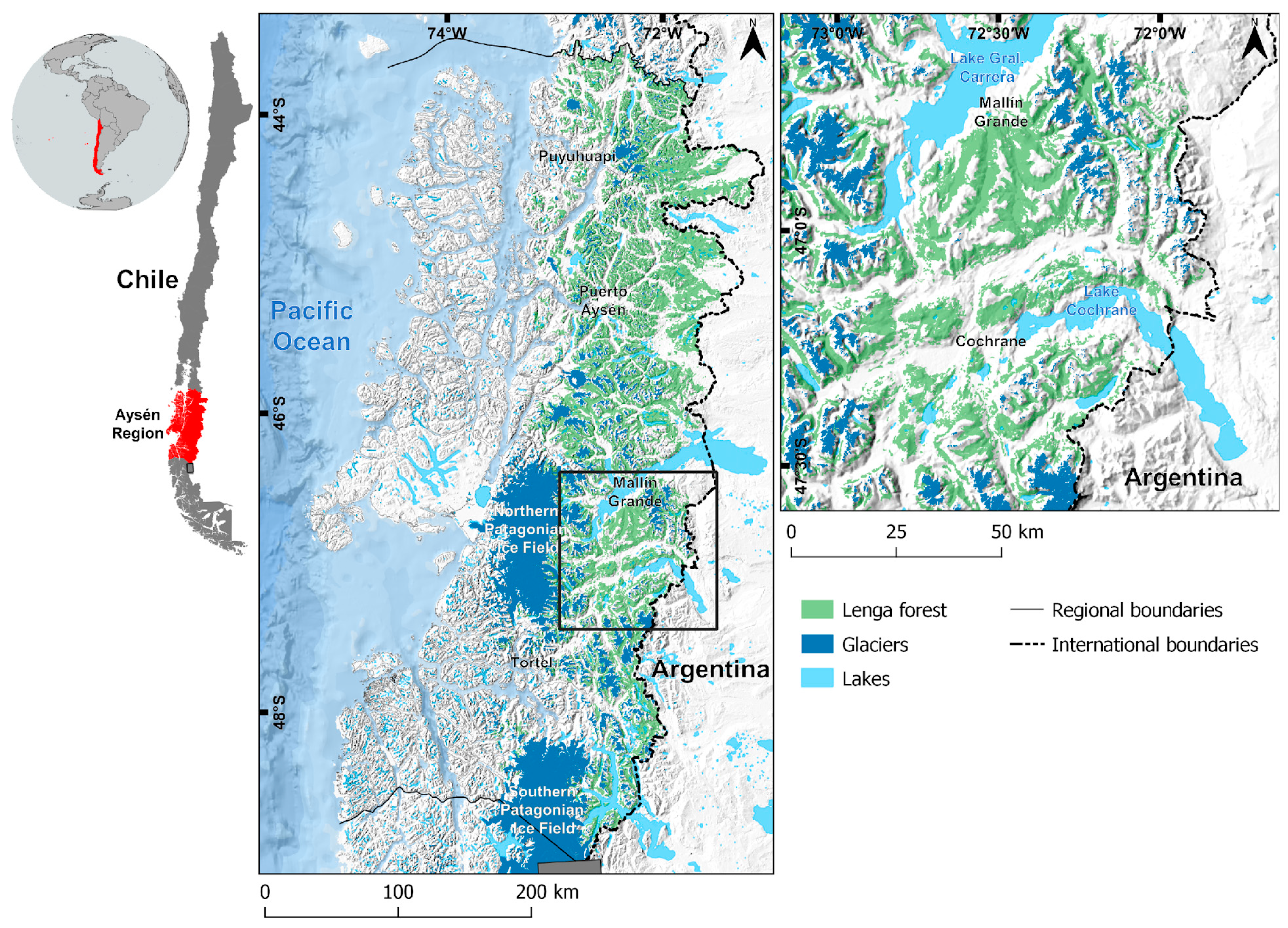
2.1. Study Area
2.2. Satellite Data
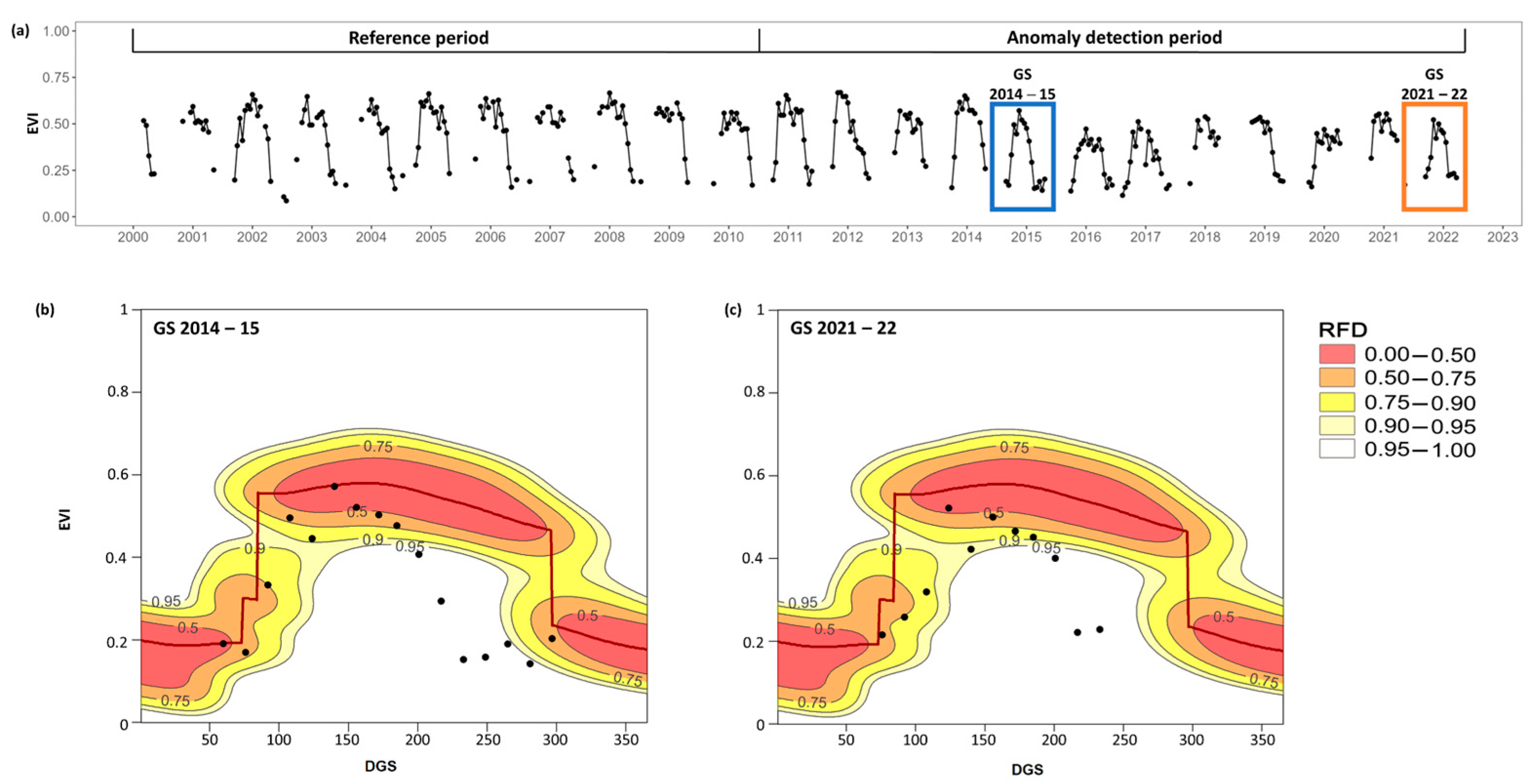
2.3. Outbreak Detection Using the “npphen” R-Package
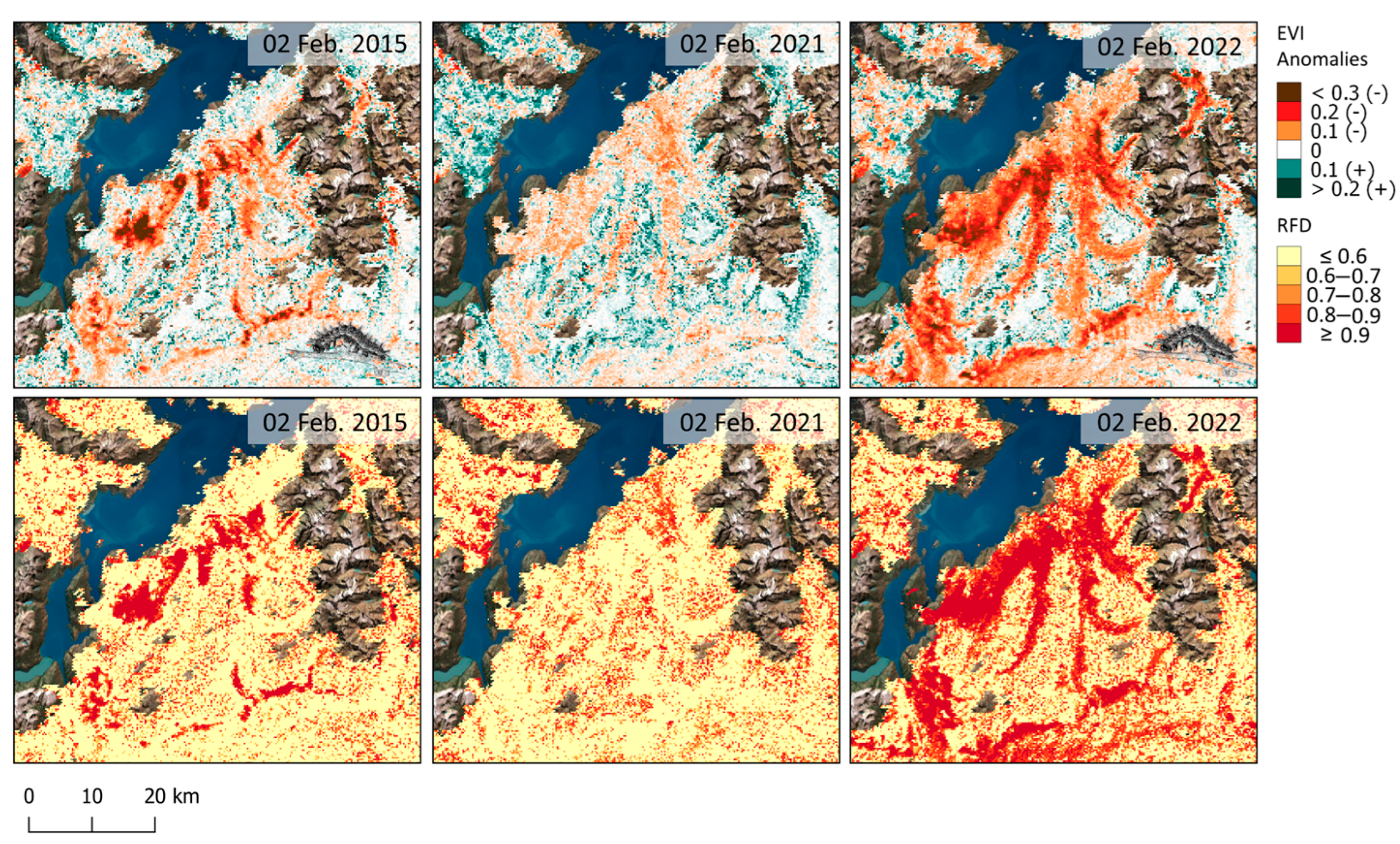
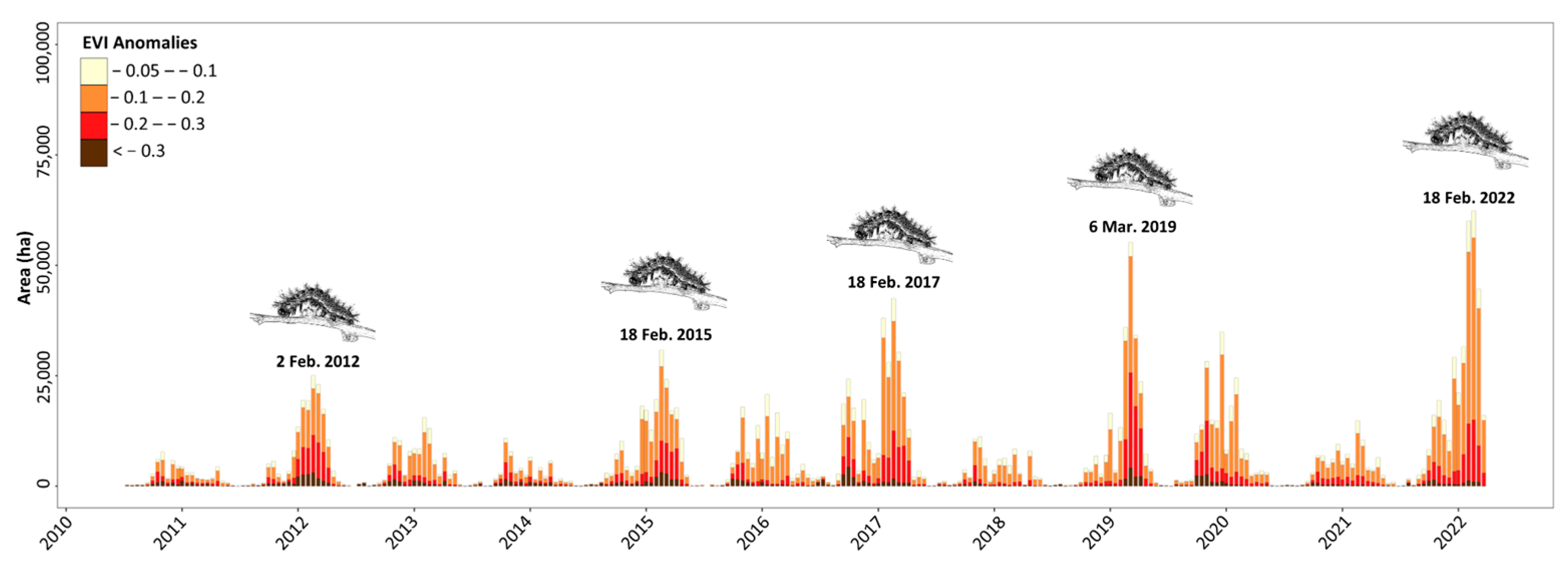
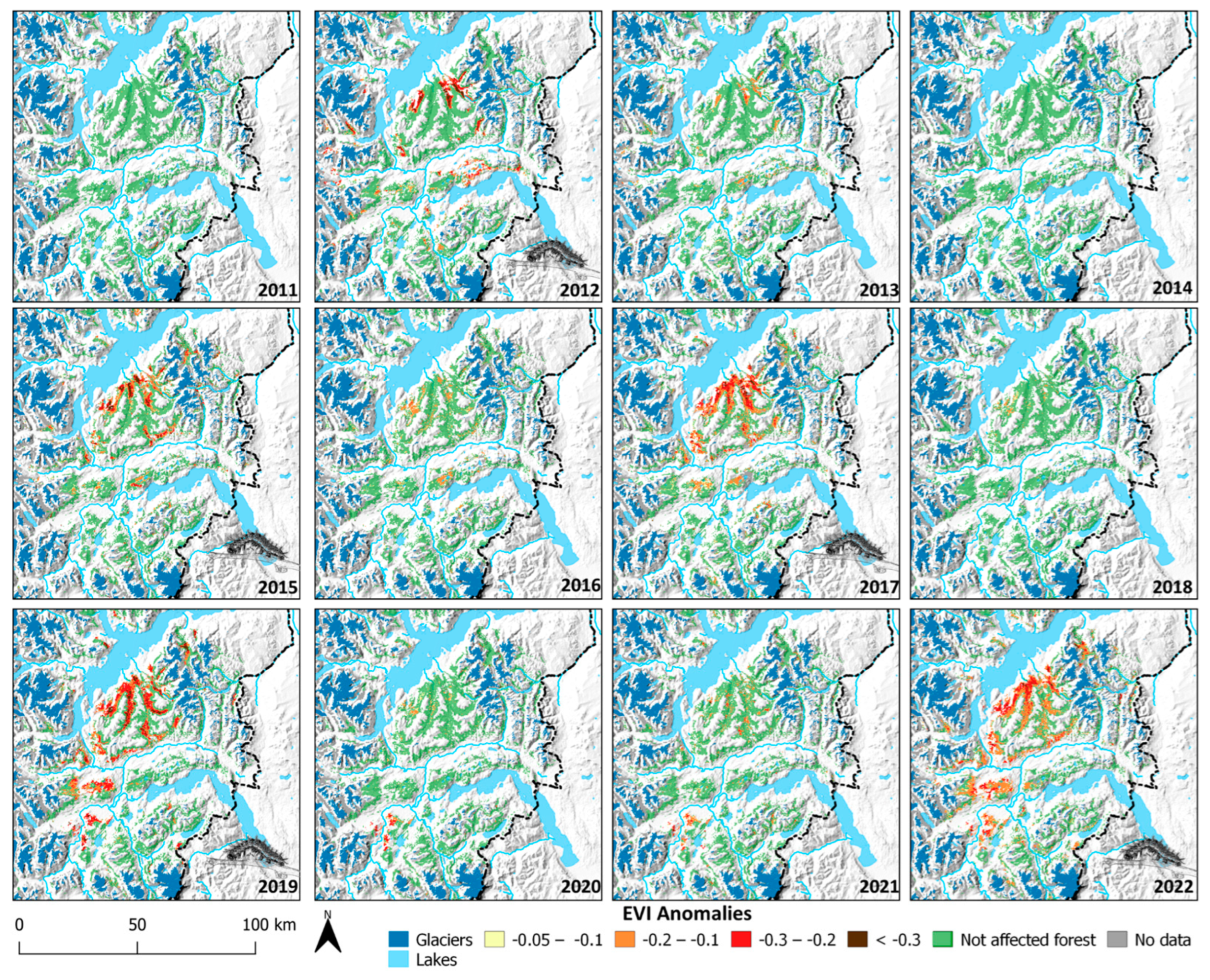
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gutiérrez, Á.G.; Chávez, R.O.; Domínguez-Concha, J.A.; Gibson-Carpintero, S.; Guerrero, I.P.; Rocco, R.; Urra, V.D.; Estay, S.A. Ormiscodes amphimone Outbreak Frequency Increased Since 2000 in Subantarctic Nothofagus pumilio Forests of Chilean Patagonia. In Forest Pest and Disease Management in Latin America; Springer: Berlin/Heidelberg, Germany, 2020; pp. 61–75. [Google Scholar]
- Paritsis, J. Ormiscodes Outbreak Dynamics: Impacts and Perspectives in a Warming World. In Forest Pest and Disease Management in Latin America: Modern Perspectives in Natural Forests and Exotic Plantations; Springer: Berlin/Heidelberg, Germany, 2020; pp. 77–88. [Google Scholar]
- Estay, S.A.; Chávez, R.O.; Rocco, R.; Gutiérrez, A.G. Quantifying Massive Outbreaks of the Defoliator Moth Ormiscodes amphimone in Deciduous Nothofagus-Dominated Southern Forests Using Remote Sensing Time Series Analysis. J. Appl. Entomol. 2019, 143, 787–796. [Google Scholar] [CrossRef]
- Chávez, R.O.; Rocco, R.; Gutiérrez, Á.G.; Dörner, M.; Estay, S.A. A Self-Calibrated Non-Parametric Time Series Analysis Approach for Assessing Insect Defoliation of Broad-Leaved Deciduous Nothofagus pumilio Forests. Remote Sens. 2019, 11, 204. [Google Scholar] [CrossRef]
- Angulo, A.O.; Lemaire, C.; Olivares, T.S. Catálogo Crítico e Ilustrado de Las Especies de La Familia Saturniidae En Chile (Lepidoptera: Saturniidae). Gayana 2004, 68, 20–42. [Google Scholar] [CrossRef]
- Paritsis, J.; Veblen, T.T. Dendroecological Analysis of Defoliator Outbreaks on Nothofagus pumilio and Their Relation to Climate Variability in the Patagonian Andes. Glob. Chang. Biol. 2011, 17, 239–253. [Google Scholar] [CrossRef]
- Garreaud, R.; Lopez, P.; Minvielle, M.; Rojas, M. Large-Scale Control on the Patagonian Climate. J. Clim. 2013, 26, 215–230. [Google Scholar] [CrossRef]
- Villalba, R.; Lara, A.; Boninsegna, J.A.; Masiokas, M.; Delgado, S.; Aravena, J.C.; Roig, F.A.; Schmelter, A.; Wolodarsky, A.; Ripalta, A. Large-Scale Temperature Changes across the Southern Andes: 20th-Century Variations in the Context of the Past 400 Years. Clim. Chang. 2003, 59, 177–232. [Google Scholar] [CrossRef]
- Gibson-Carpintero, S.; Venegas-González, A.; Urra, V.D.; Estay, S.A.; Gutiérrez, Á.G. Recent Increase in Autumn Temperature Has Stabilized Tree Growth in Forests near the Tree Lines in Chilean Patagonia. Ecosphere 2022, 13, e4266. [Google Scholar] [CrossRef]
- Chávez, R.O.; Estay, S.A.; Lastra, J.A.; Riquelme, C.G.; Olea, M.; Aguayo, J.; Decuyper, M. Npphen: An R-Package for Detecting and Mapping Extreme Vegetation Anomalies Based on Remotely Sensed Phenological Variability. Remote Sens. 2023, 15, 73. [Google Scholar] [CrossRef]
- Chávez, R.O.; Estay, S.A.; Lastra, J.A.; Riquelme, C. An R-Package for Vegetation Phenological Cycle and Anomaly Detection Using Remote Sensing Data; PUCV, AUCH: Valparaíso, Chile, 2022. [Google Scholar]
- Didan, K.; Barreto Munoz, A. MODIS Vegetation Index User’s Guide; The University of Arizona: Tucson, AZ, USA, 2019. [Google Scholar]
- Decuyper, M.; Chávez, R.O.; Lohbeck, M.; Lastra, J.A.; Tsendbazar, N.; Hackländer, J.; Herold, M.; Vågen, T.-G. Continuous Monitoring of Forest Change Dynamics with Satellite Time Series. Remote Sens. Environ. 2022, 269, 112829. [Google Scholar] [CrossRef]
- Paritsis, J.; Veblen, T.T.; Smith, J.M.; Holz, A. Spatial Prediction of Caterpillar (Ormiscodes) Defoliation in Patagonian Nothofagus Forests. Landsc. Ecol. 2011, 26, 791–803. [Google Scholar] [CrossRef]
- Rodríguez-Catón, M.; Villalba, R.; Srur, A.; Williams, A.P. Radial Growth Patterns Associated with Tree Mortality in Nothofagus pumilio Forest. Forests 2019, 10, 489. [Google Scholar] [CrossRef]
- Ward, S.F.; Aukema, B.H. Anomalous Outbreaks of an Invasive Defoliator and Native Bark Beetle Facilitated by Warm Temperatures, Changes in Precipitation and Interspecific Interactions. Ecography 2019, 42, 1068–1078. [Google Scholar] [CrossRef]
- Romashkin, I.; Neuvonen, S.; Tikkanen, O.-P. Northward Shift in Temperature Sum Isoclines May Favour Ips Typographus Outbreaks in European Russia. Agric. For. Entomol. 2020, 22, 238–249. [Google Scholar] [CrossRef]
- Nelson, W.A.; Bjørnstad, O.N.; Yamanaka, T. Recurrent Insect Outbreaks Caused by Temperature-Driven Changes in System Stability. Science 2013, 341, 796–799. [Google Scholar] [CrossRef] [PubMed]
- Estay, S.A.; Lima, M.; Bozinovic, F. The Role of Temperature Variability on Insect Performance and Population Dynamics in a Warming World. Oikos 2014, 123, 131–140. [Google Scholar] [CrossRef]
- IPCC Climate Change 2013: The Physical Science Basis; IPCC: Geneva, Switzerland, 2013; p. 1535.
- Paritsis, J.; Veblen, T.T. Temperature and Foliage Quality Affect Performance of the Outbreak Defoliator Ormiscodes amphimone (F.) (Lepidoptera: Saturniidae) in Northwestern Patagonia, Argentina. Rev. Chil. Hist. Nat. 2010, 83, 593–603. [Google Scholar] [CrossRef]
- Serra, M.N.; Quintero, C.; Rodríguez-Cabal, M.A.; Martínez, A.S.; Paritsis, J. Temperature and Host Plant Species Affect the Performance and Immunocompetence of an Outbreak Defoliator in Northwestern Patagonia. Ecol. Entomol. 2022, 47, 580–589. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estay, S.A.; Chávez, R.O.; Lastra, J.A.; Rocco, R.; Gutiérrez, Á.G.; Decuyper, M. MODIS Time Series Reveal New Maximum Records of Defoliated Area by Ormiscodes amphimone in Deciduous Nothofagus Forests, Southern Chile. Remote Sens. 2023, 15, 3538. https://doi.org/10.3390/rs15143538
Estay SA, Chávez RO, Lastra JA, Rocco R, Gutiérrez ÁG, Decuyper M. MODIS Time Series Reveal New Maximum Records of Defoliated Area by Ormiscodes amphimone in Deciduous Nothofagus Forests, Southern Chile. Remote Sensing. 2023; 15(14):3538. https://doi.org/10.3390/rs15143538
Chicago/Turabian StyleEstay, Sergio A., Roberto O. Chávez, José A. Lastra, Ronald Rocco, Álvaro G. Gutiérrez, and Mathieu Decuyper. 2023. "MODIS Time Series Reveal New Maximum Records of Defoliated Area by Ormiscodes amphimone in Deciduous Nothofagus Forests, Southern Chile" Remote Sensing 15, no. 14: 3538. https://doi.org/10.3390/rs15143538
APA StyleEstay, S. A., Chávez, R. O., Lastra, J. A., Rocco, R., Gutiérrez, Á. G., & Decuyper, M. (2023). MODIS Time Series Reveal New Maximum Records of Defoliated Area by Ormiscodes amphimone in Deciduous Nothofagus Forests, Southern Chile. Remote Sensing, 15(14), 3538. https://doi.org/10.3390/rs15143538