

Trends of Aboveground Net Primary Productivity of Patagonian Meadows, the Omitted Ecosystem in Desertification Studies

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
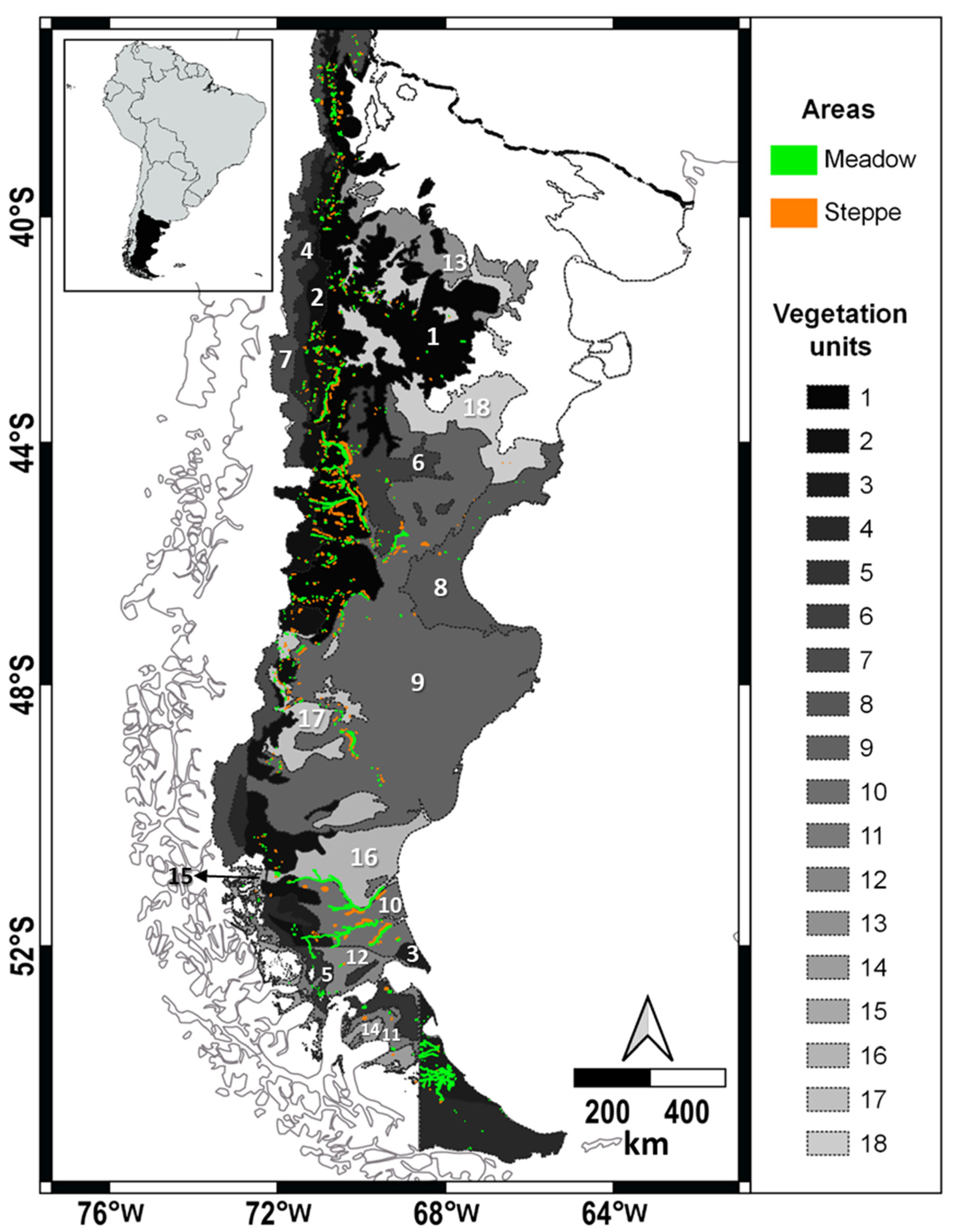
2.1. Study Area
2.2. Data
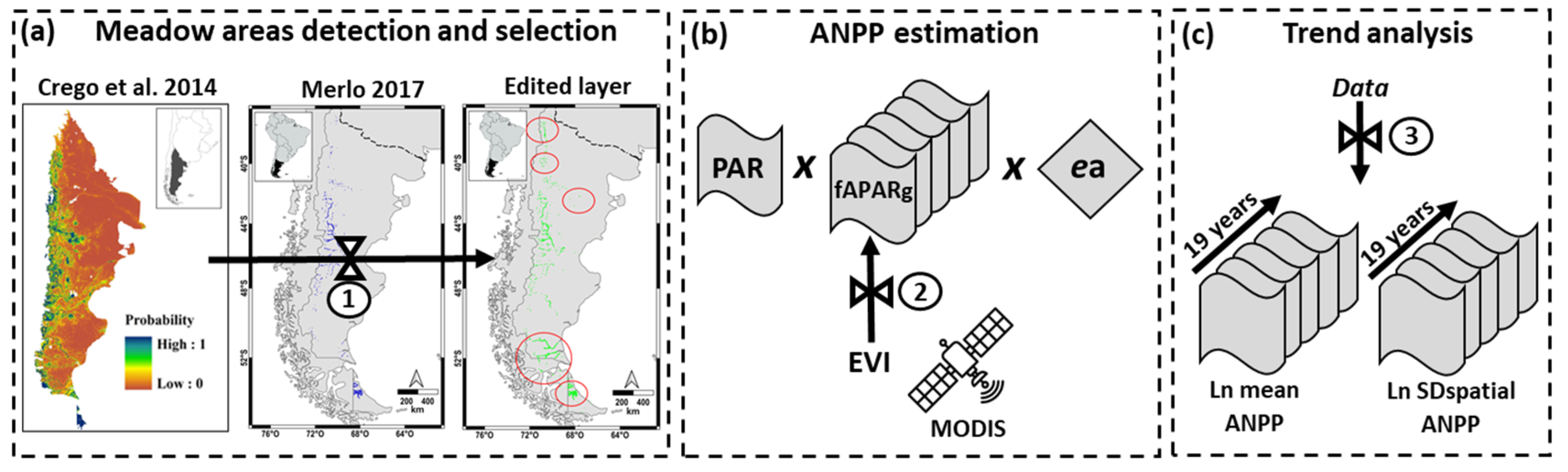
2.2.1. Meadow Area Detection and Selection
2.2.2. ANPP Estimation
2.2.3. Geographic and Climatic Data
2.2.4. Database Processing
2.3. Analysis
3. Results
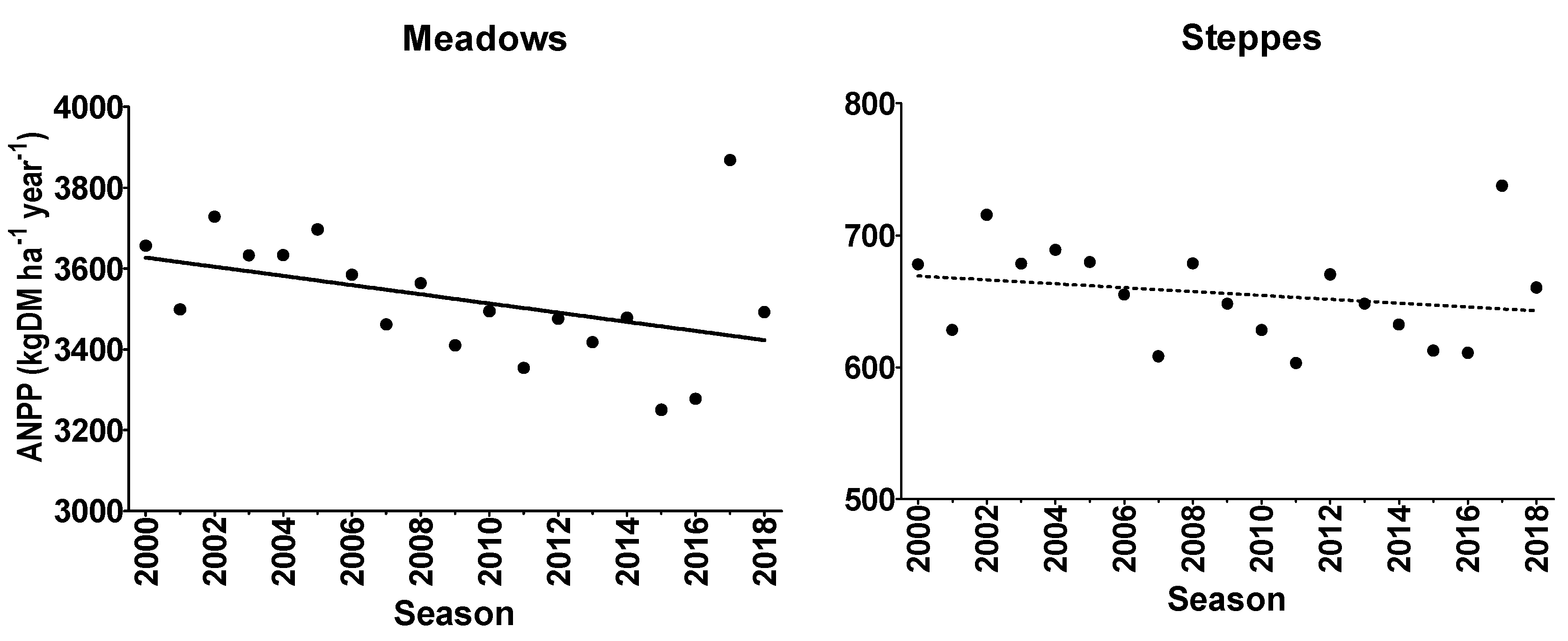
3.1. ANPP Trends
3.1.1. Average ANPP Trends
3.1.2. Relative Rate of Change of ANPP by Pair of Meadow and Steppe Areas
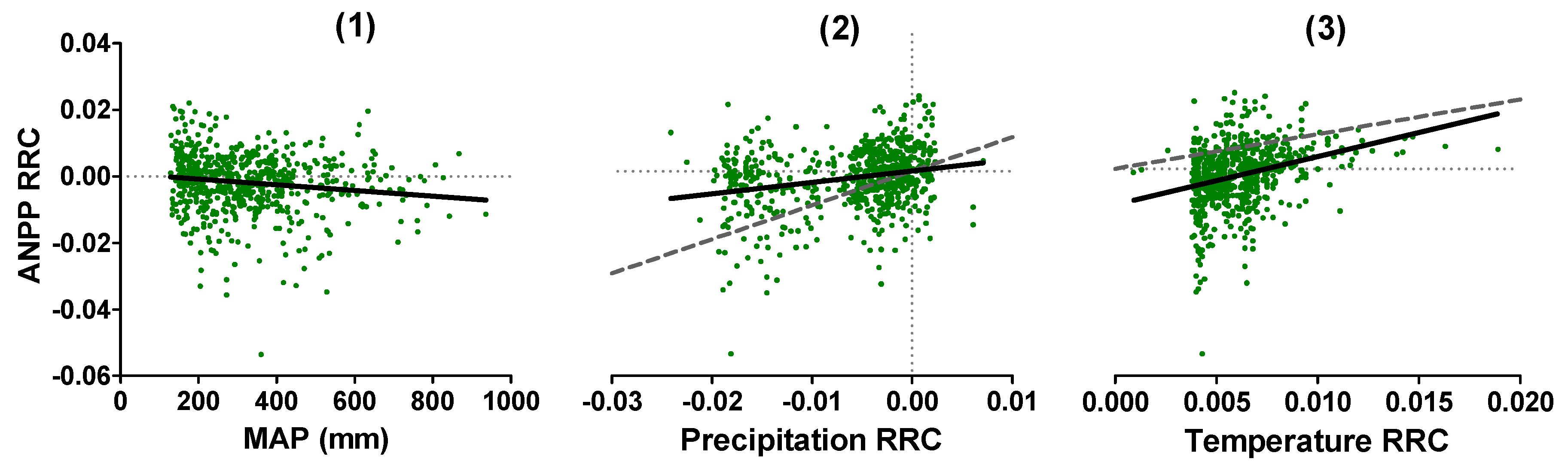
3.1.3. ANPP Trends Relationship with Environmental Controls
3.2. Impact on Livestock Carrying Capacity of Meadows
3.3. Trend Syndromes in Meadow Areas
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Xiaoyan, B.; Suocheng, D.; Wenbao, M.; Fujia, L. Spatial-temporal change of carbon storage and sink of wetland ecosystem in arid regions, Ningxia Plain. Atmos. Environ. 2019, 204, 89–101. [Google Scholar] [CrossRef]
- Fang, Q.; Wang, G.; Liu, T.; Xue, B.L.; Yinglan, A. Controls of carbon flux in a semi-arid grassland ecosystem experiencing wetland loss: Vegetation patterns and environmental variables. Agric. For. Meteorol. 2018, 259, 196–210. [Google Scholar] [CrossRef]
- UNCCD. Elaboration of an International Convention to Combat Desertification in Countries Experiencing Serious Drought and/or Desertification, Particularly in Africa. 1994, Volume 24, pp. 1–58. Available online: https://wedocs.unep.org/handle/20.500.11822/27569;jsessionid=3C6D21F808FF14535A97268CAD716845 (accessed on 7 May 2023).
- Fluet-Chouinard, E.; Stocker, B.D.; Zhang, Z.; Malhotra, A.; Melton, J.R.; Poulter, B.; Kaplan, J.O.; Goldewijk, K.K.; Siebert, S.; Minayeva, T.; et al. Extensive global wetland loss over the past three centuries. Nature 2023, 614, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Brinson, M.M.; Malvárez, A.I. Temperate freshwater wetlands: Types, status, and threats. Environ. Conserv. 2002, 29, 115–133. [Google Scholar] [CrossRef]
- Mazzoni, E.; Rabassa, J. Types and internal hydro-geomorphologic variability of mallines (wet-meadows) of Patagonia: Emphasis on volcanic plateaus. J. S. Am. Earth Sci. 2013, 46, 170–182. [Google Scholar] [CrossRef]
- Collantes, M.B.; Anchorena, J.; Stoffella, S.; Escartín, C.; Rauber, R. Wetlands of the magellanic steppe (Tierra del Fuego, Argentina). Folia Geobot. 2009, 44, 227–245. [Google Scholar] [CrossRef]
- Oliva, G.; García, G.; Ferrante, D.; Massara, V.; Rimoldi, P.; Díaz, B.; Paredes, P. Estado de los Recursos Naturales de la Patagonia Sur Centro Regional Patagonia; Instituto Nacional de Tecnología Agropecuaria: EEA Santa Cruz, Argentina, 2017. [Google Scholar] [CrossRef]
- Chimner, R.a.; Bonvissuto, G.L.; Cremona, M.V.; Gaitan, J.J.; Lopez, C.R. Ecohydrological conditions of wetlands along a precipitation gradient in Patagonia, Argentina. Ecol. Austral 2011, 21, 329–337. [Google Scholar]
- Paruelo, J.M.; Golluscio, R.A.; Guerschman, J.P.; Cesa, A.; Jouve, V.V.; Garbulsky, M.F. Regional scale relationships between ecosystem structure and functioning: The case of the Patagonian steppes. Glob. Ecol. Biogeogr. 2004, 13, 385–395. [Google Scholar] [CrossRef]
- Buono, G.; Oesterheld, M.; Nakamatsu, V.; Paruelo, J.M. Spatial and temporal variation of primary production of Patagonian wet meadows. J. Arid Environ. 2010, 74, 1257–1261. [Google Scholar] [CrossRef]
- Irisarri, J.G.N.; Oesterheld, M.; Paruelo, J.M.; Texeira, M.A. Patterns and controls of above-ground net primary production in meadows of Patagonia. A remote sensing approach. J. Veg. Sci. 2012, 23, 114–126. [Google Scholar] [CrossRef]
- Raffaele, E. Susceptibility of a Patagonian mallín flooded meadow to invasion by exotic species. Biol. Invasions 2004, 6, 473–481. [Google Scholar] [CrossRef]
- Navarro, M.; Navarro, C.; Barrios, R.; Calamari, N.; Dieta, V.; Martínez García, G.; Iturralde Elortegui, M.; Kurtz, D.; Michard, N.; Paredes, P.; et al. Mapa de Distribución Potencial de Humedales en Argentina. Informe Técnico. 2022. Available online: https://intahumedales.users.earthengine.app/view/mapahumedalesargentina (accessed on 20 April 2023).
- Bonvissuto, G.; Somlo, R.; Ayesa, J.; Lanciotti, M.L.; Moricz de Tecso, E. La condición de los mallines del área ecológica sierras y mesetas de Patagonia. Rev. Argentina Prod. Anim. 1992, 12, 391–400. [Google Scholar]
- Bruce, A.; Dufilho, A.C. Los mallines en Patagonia: Una perspectiva histórico cultural de los recursos naturales. Mundo Agrar. 2002, 2. Available online: http://www.scielo.org.ar/scielo.php?pid=S1515-59942002000100005&script=sci_arttext&tlng=en (accessed on 7 May 2023).
- Collantes, M.B.; Faggi, A.M. Los Humedales del Sur de Sudamérica. Tópicos Sobre Humedales Subtropicales y Templados de Sudamérica; Unesco: Montevideo, Uruguay, 1999. [Google Scholar]
- Easdale, M.; Gaitan, J. Relación entre la superficie y clase de mallines y la composición de la estructura ganadera en establecimientos del noroeste de la Patagonia. Rev. Argent. Prod. Anim. 2010, 30, 69–80. [Google Scholar]
- Golluscio, R.A.; Deregibus, V.A.; Paruelo, J.M. Sustainability and range management in the Patagonian steppes. Ecol. Austral 1998, 8, 265–284. [Google Scholar]
- Irisarri, J.G.N.; Texeira, M.; Oesterheld, M.; Verón, S.R.; Della Nave, F.; Paruelo, J.M. Discriminating the biophysical signal from human-induced effects on long-term primary production dynamics. The case of Patagonia. Glob. Chang. Biol. 2021, 27, 4381–4391. [Google Scholar] [CrossRef] [PubMed]
- Oliva, G.; Gaitan, J.; Ferrante, D. Humans Cause Deserts: Evidence of Irreversible Changes in Argentinian Patagonia Rangelands. In The End of Desertification? Springer: Berlin/Heidelberg, Germany, 2016; pp. 363–386. [Google Scholar] [CrossRef]
- Zhao, M.; Running, S.W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef]
- García Martínez, G.C.; Ciari, G.; Gaitan, J.; Caruso, C.; Nagahama, N.; Opazo, W.; Nakamatsu, V.; Lloyd, C.; Cotut, C.; Irisarri, J.G.N.; et al. Diagnóstico de la evolución del clima y los pastizales naturales en el noroeste de la provincia de Chubut en el período 2000-2014. Agri. Sci. 2017, 34, 59–69. [Google Scholar]
- Paruelo, J.M.; Sala, O.E. Water Losses in the Patagonian Steppe: A Modelling Approach. Ecology 1995, 76, 510–520. [Google Scholar] [CrossRef]
- Bandieri, L.M.; Fernández, R.J.; Bisigato, A.J. Risks of Neglecting Phenology When Assessing Climatic Controls of Primary Production. Ecosystems 2020, 23, 164–174. [Google Scholar] [CrossRef]
- Spinoni, J.; Barbosa, P.; De Jager, A.; McCormick, N.; Naumann, G.; Vogt, J.V.; Magni, D.; Masante, D.; Mazzeschi, M. A new global database of meteorological drought events from 1951 to 2016. J. Hydrol. Reg. Stud. 2019, 22, 100593. [Google Scholar] [CrossRef]
- Irisarri, J.G.N.; Cipriotti, P.A.; Texeira, M.; Curcio, M.H. Trends in ANPP Response to Temperature in Wetland Meadows across a Subcontinental Gradient in Patagonia. Meteorology 2022, 2, 15. [Google Scholar] [CrossRef]
- Oliva, G.; Paredes, P.; Ferrante, D.; Cepeda, C.; Rabinovich, J. Remotely sensed primary productivity shows that domestic and native herbivores combined are overgrazing Patagonia. J. Appl. Ecol. 2019, 56, 13408. [Google Scholar] [CrossRef]
- Boggio, F.; Cremona, M.V.; Aramayo, M.V.D.L.; Girardin, L.; Raffo, F.; Fariña, C.M.; Enriquez, A.S. Guía para el curso: “Restauración y Mejoramiento de Mallines Mediante obras de Redistribución del agua de Escurrimiento” Ediciones INTA 2019. Available online: http://hdl.handle.net/20.500.12123/6529 (accessed on 20 April 2023).
- Gandullo, R.; Schmid, P.; Peña, O. Dinámica de la vegetación de los humedales del Parque Nacional Laguna Blanca (Neuquén, Argentina). Propuesta de un modelo de estados y transiciones. Rev. Fac. Agron. UNLPam 2011, 20, 43–62. [Google Scholar]
- Aguiar, M.; Paruelo, J. Impacto humano sobre los ecosistemas: El caso de la desertificación. Cienchoy 2003, 13, 48–59. [Google Scholar]
- Collantes, M.B.; Escartín, C.; Braun, K.; Cingolani, A.; Anchorena, J. Grazing and grazing exclusion along a resource gradient in magellanic meadows of tierra del fuego. Rangel. Ecol. Manag. 2013, 66, 688–699. [Google Scholar] [CrossRef]
- Scheffer, M.; Bascompte, J.; Brock, W.A.; Brovkin, V.; Carpenter, S.R.; Dakos, V.; Held, H.; Van Nes, E.H.; Rietkerk, M.; Sugihara, G. Early-warning signals for critical transitions. Nature 2009, 461, 53–59. [Google Scholar] [CrossRef]
- Scheffer, M. Foreseeing tipping points. Nature 2010, 467, 411–412. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Beltrán, A.; Jobbágy, E.G.; Sala, O.E.; Golluscio, R.A. The climate of Patagonia: General patterns and controls on biotic processes. Ecol. Austral 1998, 8, 85–102. [Google Scholar]
- Jobbágy, E.; Paruelo, J.M.; León, R.J.C. Estimación del régimen de precipitación a partir de la distancia a la cordillera en el noroeste de la Patagonia. Ecol. Austral 1995, 5, 47–53. [Google Scholar]
- León, R.J.C.; Bran, D.; Collantes, M.; Paruelo, J.M.; Soriano, A. Grandes unidades de vegetacion de la Patagonia extra andina. Ecol. Austral 1998, 8, 125–144. [Google Scholar]
- Oyarzabal, M.; Clavijo, J.; Oakley, L.; Biganzoli, F.; Tognetti, P.; Barberis, I.; Maturo, H.M.; Aragón, R.; Campanello, P.I.; Prado, D.; et al. Unidades de vegetación de la Argentina. Ecol. Austral 2018, 28, 040–063. [Google Scholar] [CrossRef]
- IDE Chile. Pisos Vegetacionales de Luebert y Pliscoff. Available online: https://www.ide.cl/index.php/flora-y-fauna/item/1524-pisos-vegetacionales-luebert-pliscoff-2017 (accessed on 9 February 2023).
- Merlo, L.R. Variación Espacial y Temporal de la Producción Primaria Neta Aérea de Mallines de la Patagonia. Master’s Thesis, Universidad de Buenos Aires, Facultad de Agronomía, Buenos Aires, Argentina, 2017. [Google Scholar]
- Crego, R.D.; Didier, K.A.; Nielsen, C.K. Modeling meadow distribution for conservation action in arid and semi-arid Patagonia, Argentina. J. Arid Environ. 2014, 102, 68–75. [Google Scholar] [CrossRef]
- Mazzoni, E.; Vázquez, M. Evaluación de pastizales húmedos para un aprovechamiento sustentable en la cuenca del río Gallegos (Provincia de Santa Cruz, Argentina). In Proceedings of the VIII Encuentro Latinoam, Geógrafos, Santiago, Chile, 4–10 March 2001; pp. 175–182. [Google Scholar]
- Lopez, C.R.; Gaitan, J.J.; Siffredi, G.L.; Ayesa, J.a; Lagorio, P. a Desarrollo de un sistema de informacion geografico (SIG) como herramienta para la planificacion manejo del pasroreo en mallines del Dpto. de Pilcaniyeu, Rio Negro. Revisita Cient. Agropecu. 2005, 9, 163–171. [Google Scholar]
- Utrilla, V.; Ferrante, D.; Peri, P.; Kofalt, J.C.; Humano, G. Efecto de la dinámica hídrica edáfica y ambiental sobre la productividad y calidad forrajera de mallines en la Patagonia Austral. Informe técnico final. EEA INTA Santa Cruz 2008, 31. [Google Scholar]
- Filipová, L.; Hédl, R.; Dančák, M. Magellanic Wetlands: More than Moor. Folia Geobot. 2013, 48, 163–188. [Google Scholar] [CrossRef]
- Utrilla, V. Monitoreo de indicadores de degradación en mallines bajo pastoreo ovino en el Sur de Santa Cruz. Mayéutica 2014, 40, 31. [Google Scholar] [CrossRef]
- Vargas, P.P.; Mazzoni, E. Caracterización de la Composición Florística y Productividad Primaria del Mallín Pali Aike, Patagonia Austral Argentina. 2014. Available online: https://redargentinadegeografiafisica.files.wordpress.com/2014/04/trabajo-vargas-y-mazzoni.pdf (accessed on 10 March 2023).
- Grima, D.; Vazquez, M.; Diez, P.; Investigadores, D.; Austral, P.; Acad, U.; Laboratorio, G. Composición Florística De Pequeñas Areas De Mallines Con Distintas Exposición Y Pendiente. Inf. Científicos Técnicos-UNPA 2015, 7, 144–161. [Google Scholar] [CrossRef]
- Gaitan, J.; Bran, D.; Raffo, F.; Ayesa, J. Evaluación y cartografía de mallines de las zonas de Loncopué y Chos Malal, provincia del Neuquén. Comun. Técnica 2015, 131, 41. [Google Scholar]
- Irisarri, J.G.N.; Oyarzabal, M.; Arocena, D.; Vassallo, M.; Oesterheld, M. Focus: Software de Gestión de Información Satelital Para Observar Recursos Naturales (Versión 2018); LART, IFEVA, Universidad de Buenos Aires, CONICET, Facultad de Agronomía: Buenos Aires, Argentina, 2018. [Google Scholar]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Monteith, J.L. Climate and the Efficiency of Crop Production in Britain. Philos. Trans. R. Soc. B Biol. Sci. 1977, 281, 277–294. [Google Scholar] [CrossRef]
- Grossi Gallegos, H. Distribución espacial de la radiación fotosintéticamente activa en Argentina. Meteorologica 2004, 29, 27–36. [Google Scholar]
- Baldassini, P.; Irisarri, G.; Oyarzábal, M.; Paruelo, J.M. Eficiencia en el uso de la Radiación y Controles de la Productividad de las Estepas Patagónicas; Reunión Argentina de Ecología: Luján, Argentina, 2012. [Google Scholar]
- Chapin, F.S.; Matson, P.A.; Mooney, H.A. Principles of Terrestrial Ecosystem Ecology; Springer: Berlin/Heidelberg, Germany, 2002; ISBN 9781441995032. [Google Scholar]
- Grigera, G.; Oesterheld, M. Variability of radiation use efficiency in mixed pastures under varying resource availability, defoliation and time scale. Grassl. Sci. 2021, 67, 156–166. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Dobrowski, S.Z.; Parks, S.A.; Hegewisch, K.C. TerraClimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958–2015. Sci. Data 2018, 5, 191. [Google Scholar] [CrossRef] [PubMed]
- Irisarri, J.G.N.; Oesterheld, M. Temporal variation of stocking rate and primary production in the face of drought and land use change. Agric. Syst. 2020, 178, 102750. [Google Scholar] [CrossRef]
- Revelle, W. Psych: Procedures for Psychological, Psychometric, and Personality Research. Evanston, Illinois: Northwestern University. 2018. Available online: https://cran.r-project.org/web/packages/psych/index.html (accessed on 20 April 2023).
- ARC (Agricultural Research Council). The Nutrient Requirement of Ruminant Livestock; England Commonwealth Agricultural Bureaux: Farnham Royal, UK, 1980. [Google Scholar]
- Wickham, H.; François, R.; Henry, L.; Müller, K.; Vaughan, D. dplyr: A Grammar of Data Manipulation. Available online: https://dplyr.tidyverse.org/authors.html (accessed on 9 February 2023).
- Wickham, H.; Girlich, M. Tidyr: Tidy Messy Data. Available online: https://tidyr.tidyverse.org/ (accessed on 9 February 2023).
- Oliva, G.; Gaitan, J. Positive Changes in Regional Vegetation Cover in Patagonia Shown by MARAS Monitoring System. Int. Grassl. Congr. Proc. 2021. Available online: https://uknowledge.uky.edu/igc/24/1/38/ (accessed on 20 April 2023).
- Enriquez, A.S.; Chimner, R.A.; Cremona, M.V.; Diehl, P.; Bonvissuto, G.L. Grazing intensity levels influence C reservoirs of wet and mesic meadows along a precipitation gradient in Northern Patagonia. Wetl. Ecol. Manag. 2015, 23, 439–451. [Google Scholar] [CrossRef]
- Blackburn, H.D.; Kothmann, M.M. Modelling diet selection and intake for grazing herbivores. Ecol. Modell. 1991, 57, 145–163. [Google Scholar] [CrossRef]
- Kätterer, T.; Bolinder, M.A.; Andrén, O.; Kirchmann, H.; Menichetti, L. Roots contribute more to refractory soil organic matter than above-ground crop residues, as revealed by a long-term field experiment. Agric. Ecosyst. Environ. 2011, 141, 184–192. [Google Scholar] [CrossRef]
- Instituto Nacional de Estadísticas y Censos (INDEC) de la República Argentina Censo Nacional Agropecuario 2002. Available online: https://datos.gob.ar/dataset/agroindustria-censo---ganaderia-cna-02 (accessed on 10 February 2023).
- Instituto Nacional de Estadística y Censos (INDEC) de la República Argentina Censo Nacional Agropecuario 2018. Available online: https://www.indec.gob.ar/indec/web/Nivel4-Tema-3-8-87 (accessed on 10 February 2023).
- Verón, S.R.; Paruelo, J.M. Desertification alters the response of vegetation to changes in precipitation. J. Appl. Ecol. 2010, 47, 1233–1241. [Google Scholar] [CrossRef]
- Prince, S.D.; Wessels, K.J.; Tucker, C.J.; Nicholson, S.E. Desertification in the Sahel: A reinterpretation of a reinterpretation. Glob. Chang. Biol. 2007, 13, 1308–1313. [Google Scholar] [CrossRef]
- Verón, S.R.; Paruelo, J.M.; Oesterheld, M. Assessing desertification. J. Arid Environ. 2006, 66, 751–763. [Google Scholar] [CrossRef]
- Easdale, M.H.; Fariña, C.; Hara, S.; Pérez León, N.; Umaña, F.; Tittonell, P.; Bruzzone, O. Trend-cycles of vegetation dynamics as a tool for land degradation assessment and monitoring. Ecol. Indic. 2019, 107, 105545. [Google Scholar] [CrossRef]
- Westoby, M.; Walker, B.; Noy-Meir, I. Opportunistic management for rangelands not at equilibrium. Rangel. Ecol. Manag. J. Range Manag. Arch. 1989, 42, 266–274. [Google Scholar] [CrossRef]
- Enriquez, A.S.; Cremona, M.V. Particulate organic carbon is a sensitive indicator of soil degradation related to overgrazing in Patagonian wet and mesic meadows. Wetl. Ecol. Manag. 2018, 26, 345–357. [Google Scholar] [CrossRef]
- Montgomery, D.R. Soil erosion and agricultural sustainability. Proc. Natl. Acad. Sci. USA 2007, 104, 13268–13272. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, H.F. Mallines de ambiente árido. Pradera salina y estepa arbustiva graminosa en el NO del Chubut. In Secuencias de Deterioro de Distintos Ambientes Patagónicos. Su Caracterarterización Mediante el Modelo de Estados y Transiciones; Paruelo, J.M., Bertiller, M.B., Schichter, T.M., Coronato, F.R., Eds.; Convenio Argentino-Alemán de Cooperación Técnica, Instituto Nacional de Tecnología Agropecuaria (INTA), Deutsche Gesellschaftfür Technische Zusammenarbeit (INTA-GTZ) (Ludepa-SMR): Buenos Aires, Argentina, 1993; pp. 31–39. [Google Scholar]
- Enriquez, A.S.; Cremona, M.V. El rol de los suelos en la restauracion ecológica. Ed. Univ. Nac. Comahue 2018, 37–58. Available online: https://repositorio.inta.gob.ar/handle/20.500.12123/5065 (accessed on 20 April 2023).
- Epele, L.B.; Grech, M.G.; Manzo, L.M.; Macchi, P.A.; Hermoso, V.; Miserendino, M.L.; Bonada, N.; Cañedo-Argüelles, M. Identifying high priority conservation areas for Patagonian wetlands biodiversity. Biodivers. Conserv. 2021, 30, 1359–1374. [Google Scholar] [CrossRef]
- Oliva, G.; Dos Santos, E.; Sofía, O.; Umaña, F.; García Martínez, G.; Caruso, C.; Cariac, G.; Echevarría, D.; Fantozzi, A.; Butti, L.; et al. The MARAS dataset, vegetation and soil characteristics of dryland rangelands across Patagonia. Sci. Data 2020, 7, 327. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curcio, M.; Irisarri, G.; García Martínez, G.; Oesterheld, M. Trends of Aboveground Net Primary Productivity of Patagonian Meadows, the Omitted Ecosystem in Desertification Studies. Remote Sens. 2023, 15, 2531. https://doi.org/10.3390/rs15102531
Curcio M, Irisarri G, García Martínez G, Oesterheld M. Trends of Aboveground Net Primary Productivity of Patagonian Meadows, the Omitted Ecosystem in Desertification Studies. Remote Sensing. 2023; 15(10):2531. https://doi.org/10.3390/rs15102531
Chicago/Turabian StyleCurcio, Matías, Gonzalo Irisarri, Guillermo García Martínez, and Martín Oesterheld. 2023. "Trends of Aboveground Net Primary Productivity of Patagonian Meadows, the Omitted Ecosystem in Desertification Studies" Remote Sensing 15, no. 10: 2531. https://doi.org/10.3390/rs15102531
APA StyleCurcio, M., Irisarri, G., García Martínez, G., & Oesterheld, M. (2023). Trends of Aboveground Net Primary Productivity of Patagonian Meadows, the Omitted Ecosystem in Desertification Studies. Remote Sensing, 15(10), 2531. https://doi.org/10.3390/rs15102531