Vegetation Abundance and Health Mapping Over Southwestern Antarctica Based on WorldView-2 Data and a Modified Spectral Mixture Analysis



Abstract
1. Introduction
2. Study Area and Datasets
2.1. Study Area
2.2. Remote Sensing Data
2.3. Field Measurement Data
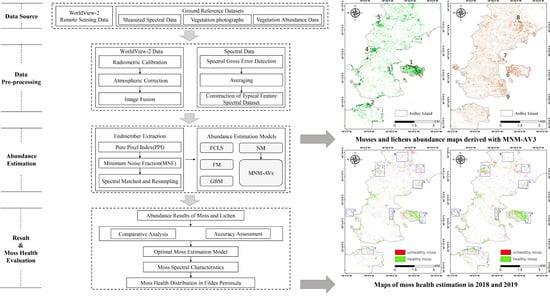
3. Methodology
3.1. Data Preprocessing
3.2. Endmember Extraction
3.3. Vegetation Abundance Estimation
3.3.1. The Linear Mixture Model
3.3.2. Nonlinear Mixture Models
- (a)
- Nascimento’s model (NM)
- (b)
- Fan’s model (FM)
- (c)
- Generalized bilinear model (GBM)
3.3.3. Modified Nascimento’s Models for Antarctic Vegetated Areas (MNM-AVs)
3.4. Moss Health Evaluation
4. Results
4.1. Estimation of Vegetation Abundance
4.2. Evaluation of Moss Health Status
4.3. Uncertainties and Limitations
5. Discussions
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A






References
- Longton, R.E. Biology of Polar Bryophytes and Lichens; Cambridge University Press: Cambridge, UK, 1988; p. 391. [Google Scholar] [CrossRef]
- Bokhorst, S.; Huiskes, A.; Convey, P.; Aerts, R. The effect of environmental change on vascular plant and cryptogam communities from the Falkland Islands and the Maritime Antarctic. BMC Ecol. 2007, 7, 15. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.-I.; Kim, H.-C.; Kim, S.-I.; Hong, S.G. Vegetation abundance on the Barton Peninsula, Antarctica: Estimation from high-resolution satellite images. Polar Biol. 2014, 37, 1579–1588. [Google Scholar] [CrossRef]
- Kim, J.; Ahn, I.-Y.; Hong, S.G.; Andreev, M.; Lim, K.-M.; Oh, M.; Koh, Y.; Hur, J.-S. Lichen flora around the Korean Antarctic Scientific Station, King George Island. Antarct. J. Microbiol. 2006, 44, 480–491. [Google Scholar]
- Lee, J.S.; Lee, H.K.; Hur, J.S.; Andreev, M.; Hong, S.G. Diversity of the lichenized fungi in King George Island, Antarctica, revealed by phylogenetic analysis of partial large subunit rDNA sequences. J. Microbiol. Biotechnol. 2008, 18, 1016–1023. [Google Scholar] [PubMed]
- Shen, Y.P.; Wang, Y.G. Key findings and assessment results of IPCC WGI Fifth assessment report. J. Glaciol. Geocryol. 2013, 35, 1068–1076. [Google Scholar]
- Bergstrom, D.M.; Turner, P.A.M.; Scott, J.; Copson, G.; Shaw, J. Restricted plant species on sub-Antarctic Macquarie and Heard Islands. Polar Biol. 2005, 29, 532–539. [Google Scholar] [CrossRef]
- Convey, P. Antarctic terrestrial biodiversity in a changing world. Polar Biol. 2011, 34, 1629–1641. [Google Scholar] [CrossRef]
- Sancho, L.G.; Pintado, A. Evidence of high annual growth rate for lichens in the maritime Antarctic. Polar Biol. 2004, 27, 312–319. [Google Scholar] [CrossRef]
- Torres-Mellado, G.; Jana, R.; Casanova-Katny, M. Antarctic hairgrass expansion in the South Shetland archipelago and Antarctic Peninsula revisited. Polar Biol. 2011, 34, 1679. [Google Scholar] [CrossRef]
- Green, T.A.; Sancho, L.G.; Pintado, A.; Schroeter, B. Functional and spatial pressures on terrestrial vegetation in Antarctica forced by global warming. Polar Biol. 2011, 34, 1643–1656. [Google Scholar] [CrossRef]
- Doran, P.T.; Priscu, J.C.; Lyons, W.B.; Walsh, J.E.; Fountain, A.G.; McKnight, D.M.; Moorhead, D.L.; Virginia, R.A.; Wall, D.H.; Clow, G.D. Antarctic climate cooling and terrestrial ecosystem response. Nature 2002, 415, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.; Barrand, N.E.; Bracegirdle, T.J.; Convey, P.; Hodgson, D.A.; Jarvis, M.; Jenkins, A.; Marshall, G.; Meredith, M.P.; Roscoe, H. Antarctic climate change and the environment: An update. Polar Rec. 2014, 50, 237–259. [Google Scholar] [CrossRef]
- Cannone, N.; Dalle Fratte, M.; Convey, P.; Worland, M.R.; Guglielmin, M. Ecology of moss banks on Signy Island (maritime Antarctic). Bot. J. Linnean Soc. 2017, 184, 518–533. [Google Scholar] [CrossRef]
- Robinson, S.A.; Wasley, J.; Tobin, A.K. Living on the edge–plants and global change in continental and maritime Antarctica. Glob. Chang. Biol. 2003, 9, 1681–1717. [Google Scholar] [CrossRef]
- Furmańczyk, K.; Ochyra, R. Plant communities of the Admiralty Bay region (King George Island, South Shetland Islands, Antarctic) I. Jasnorzewski Gardens. Pol. Polar Res. 1982, 3, 25–39. [Google Scholar]
- Poole, I.; Hunt, R.J.; Cantrill, D.J. A Fossil Wood Flora from King George Island: Ecological Implications for an Antarctic Eocene Vegetation. Ann. Bot. 2001, 88, 33–54. [Google Scholar] [CrossRef]
- Kim, J.H.; Ahn, I.-Y.; Lee, K.S.; Chung, H.; Choi, H.-G. Vegetation of Barton Peninsula in the neighbourhood of King Sejong Station (King George Island, maritime Antarctic). Polar Biol. 2007, 30, 903–916. [Google Scholar] [CrossRef]
- De, F.; Victoria, F.; Pereira, A.; Costa, D. Composition and distribution of moss formations in the ice-free areas adjoining the Arctowski region, Admiralty Bay, King George Island, Antarctica. Iheringia Ser. Bot. 2008, 64, 81–91. [Google Scholar]
- Fretwell, P.T.; Convey, P.; Fleming, A.H.; Peat, H.J.; Hughes, K.A. Detecting and mapping vegetation distribution on the Antarctic Peninsula from remote sensing data. Polar Biol. 2011, 34, 273–281. [Google Scholar] [CrossRef]
- Casanovas, P.; Black, M.; Fretwell, P.; Convey, P. Mapping lichen distribution on the Antarctic Peninsula using remote sensing, lichen spectra and photographic documentation by citizen scientists. Polar Res. 2015, 34, 25633. [Google Scholar] [CrossRef]
- Casanovas, P.; Lynch, H.J.; Fagan, W.F.; Naveen, R. Understanding lichen diversity on the Antarctic Peninsula using parataxonomic units as a surrogate for species richness. Ecology 2013, 94, 2110. [Google Scholar] [CrossRef]
- Sotille, M.E.; Bremer, U.F.; Vieira, G.; Velho, L.F.; Petsch, C.; Simões, J.C. Evaluation of UAV and satellite-derived NDVI to map maritime Antarctic vegetation. Appl. Geogr. 2020, 125, 102322. [Google Scholar] [CrossRef]
- Raynolds, M.K.; Walker, D.A.; Maier, H.A. NDVI patterns and phytomass distribution in the circumpolar Arctic. Remote Sens. Environ. 2006, 102, 271–281. [Google Scholar] [CrossRef]
- Kim, S.-H.; Hong, C.-H. Antarctic land-cover classification using IKONOS and Hyperion data at Terra Nova Bay. Int. J. Remote Sens. 2012, 33, 7151–7164. [Google Scholar] [CrossRef]
- Vieira, G.; Mora, C.; Pina, P.; Schaefer, C.E.R. A proxy for snow cover and winter ground surface cooling: Mapping Usnea sp. communities using high resolution remote sensing imagery (Maritime Antarctica). Geomorphology 2014, 225, 69–75. [Google Scholar] [CrossRef]
- Power, S.N.; Salvatore, M.R.; Sokol, E.R.; Stanish, L.F.; Barrett, J.E. Estimating microbial mat biomass in the McMurdo Dry Valleys, Antarctica using satellite imagery and ground surveys. Polar Biol. 2020, 43, 1753–1767. [Google Scholar] [CrossRef]
- Levy, J.; Cary, S.C.; Joy, K.; Lee, C.K. Detection and community-level identification of microbial mats in the McMurdo Dry Valleys using drone-based hyperspectral reflectance imaging. Antarct. Sci. 2020, 32, 367–381. [Google Scholar] [CrossRef]
- Jawak, S.D.; Luis, A.J.; Fretwell, P.T.; Convey, P.; Durairajan, U.A. Semiautomated Detection and Mapping of Vegetation Distribution in the Antarctic Environment Using Spatial-Spectral Characteristics of WorldView-2 Imagery. Remote Sens. 2019, 11, 1909. [Google Scholar] [CrossRef]
- Jawak, S.D.; Udhayaraj, A.; Luis, A.J. Geospatial mapping of vegetation in the Antarctic environment using very high-resolution WorldView-2 imagery. In Proceedings of the Land Surface and Cryosphere Remote Sensing III, New Delhi, India, 5 May 2016. [Google Scholar]
- Jawak, S.D.; Luis, A.J. A spectral index ratio-based Antarctic land-cover mapping using hyperspatial 8-band WorldView-2 imagery. Polar Sci. 2013, 7, 18–38. [Google Scholar] [CrossRef]
- Wahba, G. Support vector machines, reproducing kernel Hilbert spaces and the randomized GACV. In Advances in Kernel Methods-Support Vector Learning; Schölkopf, B., Burges, C.J.C., Smola, A.J., Eds.; MIT: Cambridge, MA, USA, 1998; pp. 125–143. [Google Scholar]
- Ichoku, C.; Karnieli, A. A review of mixture modeling techniques for sub-pixel land cover estimation. Remote Sens. Rev. 1996, 13, 161–186. [Google Scholar] [CrossRef]
- Théau, J.; Peddle, D.R.; Duguay, C.R. Mapping lichen in a caribou habitat of Northern Quebec, Canada, using an enhancement_classification method and spectral mixture analysis. Remote Sens. Environ. 2005, 94, 232–243. [Google Scholar] [CrossRef]
- Morison, M.; Cloutis, E.; Mann, P. Spectral unmixing of multiple lichen species and underlying substrate. Int. J. Remote Sens. 2014, 35, 478–492. [Google Scholar] [CrossRef]
- Zhang, J.; Rivard, B.; Sánchez-Azofeifa, A. Spectral unmixing of normalized reflectance data for the deconvolution of lichen and rock mixtures. Remote Sens. Environ. 2005, 95, 57–66. [Google Scholar] [CrossRef]
- Calviño-Cancela, M.; Martín-Herrero, J. Spectral Discrimination of Vegetation Classes in Ice-Free Areas of Antarctica. Remote Sens. 2016, 8, 856. [Google Scholar]
- Davidson, S.J.; Santos, M.J.; Sloan, V.L.; Watts, J.D.; Phoenix, G.K.; Oechel, W.C.; Zona, D. Mapping Arctic Tundra Vegetation Communities Using Field Spectroscopy and Multispectral Satellite Data in North Alaska, USA. Remote Sens. 2016, 8, 978. [Google Scholar] [CrossRef]
- Øvstedal, D.; Lewis Smith, R. Lichens of Antarctica and South Georgia: A Guide to Their Identification and Ecology; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Bartak, M.; Váczi, P.; Stachoň, Z.; Kubešová, S. Vegetation mapping of moss-dominated areas of northern part of James Ross Island (Antarctica) and a suggestion of protective measures. Czech Polar Rep. 2015, 5, 75–87. [Google Scholar] [CrossRef]
- Robinson, S.A.; King, D.H.; Bramley-Alves, J.; Waterman, M.J.; Ashcroft, M.B.; Wasley, J.; Turnbull, J.D.; Miller, R.E.; Ryan-Colton, E.; Benny, T.; et al. Rapid change in East Antarctic terrestrial vegetation in response to regional drying. Nat. Clim. Chang. 2018, 8, 879–884. [Google Scholar] [CrossRef]
- King, D.H.; Wasley, J.; Ashcroft, M.B.; Ryan-Colton, E.; Lucieer, A.; Chisholm, L.A.; Robinson, S.A. Semi-Automated Analysis of Digital Photographs for Monitoring East Antarctic Vegetation. Front Plant Sci. 2020, 11, 766. [Google Scholar] [CrossRef]
- Malenovský, Z.; Turnbull, J.D.; Lucieer, A.; Robinson, S.A. Antarctic moss stress assessment based on chlorophyll content and leaf density retrieved from imaging spectroscopy data. New Phytol. 2015, 208, 608–624. [Google Scholar] [CrossRef]
- Turner, D.J.; Malenovský, Z.; Lucieer, A.; Turnbull, J.D.; Robinson, S.A. Optimizing Spectral and Spatial Resolutions of Unmanned Aerial System Imaging Sensors for Monitoring Antarctic Vegetation. IEEE J. Stars 2019, 12, 3813–3825. [Google Scholar] [CrossRef]
- Pressel, S. The illustrated moss flora of Antarctica. Ann. Bot. 2009, 104, vi–vii. [Google Scholar] [CrossRef][Green Version]
- Øvstedal, D.; Smith, R. Further additions to the lichen flora of Antarctica and South Georgia. Nova Hedwig. 2009, 88, 157–168. [Google Scholar] [CrossRef]
- Olech, M. Plant Communities on King George Island. In Geoecology of Antarctic Ice-Free Coastal Landscapes; Beyer, L., Bölter, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; Volume 154, pp. 215–231. [Google Scholar]
- Andrade, A.M.D.; Michel, R.F.M.; Bremer, U.F.; Schaefer, C.E.G.R.; Simões, J.C. Relationship between solar radiation and surface distribution of vegetation in Fildes Peninsula and Ardley Island, Maritime Antarctica. Int. J. Remote Sens. 2018, 39, 2238–2254. [Google Scholar] [CrossRef]
- Xiao, C.S.; Liu, D.L.; Zhou, P.J. The Study of Total Number of Bacteria and Fungi isolated from the Fildes Peninsula in Antarctic. Antarct. Res. 1994, 6, 67–72. [Google Scholar]
- Shen, J.; Xu, R.M.; Zhou, G.F.; Wu, B.L.; Huang, F.P. Research on the structure and relationship of terrestrial, freshwater, intertidal and shallow sea ecosystems in Fildes Peninsula, Antarctic. Chin. J. Polar Res. 1999, 11, 23–35. [Google Scholar]
- Pina, P.; Vieira, G.; Bandeira, L.; Mora, C. Accurate determination of surface reference data in digital photographs in ice-free surfaces of Maritime Antarctica. Sci. Total Environ. 2016, 573, 290–302. [Google Scholar] [CrossRef]
- Lee, Y.I.; Lim, H.S.; Yoon, H.I. Geochemistry of soils of King George Island, South Shetland Islands, West Antarctica: Implications for pedogenesis in cold polar regions. Geochim. Cosmochim. Acta 2004, 68, 4319–4333. [Google Scholar] [CrossRef]
- Michel, R.F.M.; Schaefer, C.E.G.R.; López-Martínez, J.; Simas, F.N.B.; Haus, N.W.; Serrano, E.; Bockheim, J.G. Soils and landforms from Fildes Peninsula and Ardley Island, Maritime Antarctica. Geomorphology 2014, 225, 76–86. [Google Scholar] [CrossRef]
- Digital Globe. Radiometric Use of World View-2 Imagery. 2012. Available online: www.digitalglobe.com/downloads/Radiometric_Use_of_WorldView-2_Imagery.pdf (accessed on 11 May 2019).
- Rees, W.G.; Tutubalina, O.V.; Golubeva, E.I. Reflectance spectra of subarctic lichens between 400 and 2400 nm. Remote Sens. Environ. 2004, 90, 281–292. [Google Scholar] [CrossRef]
- Jensen, J.R. Introductory Digital Image Processing: A REMOTE Sensing Perspective, 3rd ed.; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2005; pp. 431–465. [Google Scholar]
- Adams, J.B.; Smith, M.O.; Johnson, P.E. Spectral mixture modeling: A new analysis of rock and soil types at the Viking Lander 1 Site. J. Geophys. Res. Sol. Earth 1986, 91, 8098–8112. [Google Scholar] [CrossRef]
- Nascimento, J.M.P.; Bioucas-Dias, J.M. Nonlinear mixture model for hyperspectral unmixing. In Proceedings of the Image and Signal Processing for Remote Sensing XV, Berlin, Germany, 28 September 2009; p. 74770I. [Google Scholar]
- Fan, W.; Hu, B.; Miller, J.; Li, M. Comparative study between a new nonlinear model and common linear model for analysing laboratory simulated-forest hyperspectral data. Int. J. Remote Sens. 2009, 30, 2951–2962. [Google Scholar] [CrossRef]
- Altmann, Y.; Dobigeon, N.; Tourneret, J. Bilinear models for nonlinear unmixing of hyperspectral images. In Proceedings of the 2011 3rd Workshop on Hyperspectral Image and Signal Processing: Evolution in Remote Sensing (WHISPERS), Lisbon, Portugal, 6–9 June 2011. [Google Scholar]
- Halimi, A.; Altmann, Y.; Dobigeon, N.; Tourneret, J. Nonlinear Unmixing of Hyperspectral Images Using a Generalized Bilinear Model. IEEE Trans. Geosci. Remote Sens. 2011, 49, 4153–4162. [Google Scholar] [CrossRef]
- Somers, B.; Cools, K.; Delalieux, S.; Stuckens, J.; Van der Zande, D.; Verstraeten, W.W.; Coppin, P. Nonlinear Hyperspectral Mixture Analysis for tree cover estimates in orchards. Remote Sens. Environ. 2009, 113, 1183–1193. [Google Scholar] [CrossRef]
- Dunn, J.L.; Robinson, S.A. Ultraviolet B screening potential is higher in two cosmopolitan moss species than in a co-occurring Antarctic endemic moss: Implications of continuing ozone depletion. Global Chang. Biol. 2006, 12, 2282–2296. [Google Scholar] [CrossRef]
- Waterman, M.J.; Bramley-Alves, J.; Miller, R.E.; Keller, P.A.; Robinson, S.A. Photoprotection enhanced by red cell wall pigments in three East Antarctic mosses. Biol. Res. 2018, 51, 49. [Google Scholar] [CrossRef]
- Selkirk, P.M.; Skotnicki, M.L. Measurement of moss growth in continental Antarctica. Polar Biol. 2007, 30, 407–413. [Google Scholar] [CrossRef]
- Pang, X.P.; Li, Y.H. Eco-environmental spatial characteristics of Fildes Peninsula based on TuPu models. Adv. Polar Sci. 2012, 23, 155–162. [Google Scholar] [CrossRef]
- Miranda, V.; Pina, P.; Heleno, S.; Vieira, G.; Mora, C.; Schaefer, C.E. Monitoring recent changes of vegetation in Fildes Peninsula (King George Island, Antarctica) through satellite imagery guided by UAV surveys. Sci. Total Environ. 2020, 704, 135295. [Google Scholar] [CrossRef]
- Lucieer, A.; Turner, D.; King, D.H.; Robinson, S.A. Using an Unmanned Aerial Vehicle (UAV) to capture micro-topography of Antarctic moss beds. Int. J. Appl. Earth Obs. 2014, 27, 53–62. [Google Scholar] [CrossRef]
- May, J.L.; Parker, T.; Unger, S.; Oberbauer, S.F. Short term changes in moisture content drive strong changes in Normalized Difference Vegetation Index and gross primary productivity in four Arctic moss communities. Remote Sens. Environ. 2018, 212, 114–120. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bands | Spectral Range (nm) | Characteristics |
|---|---|---|
| Band 1 Coastline | 400–450 | lichen absorption |
| Band 2 Blue | 450–510 | lichen reflection |
| Band 3 Green | 510–580 | moss health status |
| Band 4 Yellow | 582–625 | snow reflection |
| Band 5 Red | 620–690 | vegetation absorption |
| Band 6 RedEdge | 705–745 | moss health status |
| Band 7 NIR1 | 770–895 | vegetation reflection |
| Band 8 NIR2 | 860–1040 | vegetation reflection |
| Model | Type | Assumptions |
|---|---|---|
| Fully Constrained Least Square (FCLS) | Linear | Assumes no interaction between the objects |
| Nascimento’s Model (NM) | Nonlinear | Considers the secondary interactions between objects |
| Fan’s Model (FM) | Nonlinear | Assume that the secondary scattering effects of two endmembers in the pixel are directly related to their abundance |
| Generalized Bilinear Model (GBM) | Nonlinear | Considers the energy consumption in the transmission process by adding adjustment coefficient |
| Modified Nascimento’s Models for Antarctic Vegetated areas (MNM-AV) 1-3 | Nonlinear | Redistributes the virtual abundance considering the actual situation in AP |
| Model | FCLS | NM | FM | GBM | MNM-AV1 | MNM-AV2 | MNM-AV3 |
|---|---|---|---|---|---|---|---|
| R2 | 0.750 | 0.660 | 0.803 | 0.825 | 0.786 | 0.788 | 0.782 |
| RMSE | 0.212 | 0.403 | 0.243 | 0.231 | 0.250 | 0.215 | 0.202 |
| Model | FCLS | NM | FM | GBM | MNM-AV1 | MNM-AV2 | MNM-AV3 |
|---|---|---|---|---|---|---|---|
| R2 | 0.557 | 0.670 | 0.726 | 0.713 | 0.673 | 0.723 | 0.692 |
| RMSE | 0.322 | 0.374 | 0.282 | 0.272 | 0.306 | 0.237 | 0.212 |
| Class | Unhealthy (pixel) | Non-moss | Healthy (pixel) | Commission (%) | Omission (%) | Produced Accuracy (%) | User Accuracy (%) |
|---|---|---|---|---|---|---|---|
| Unhealthy | 6 | 0 | 0 | 0.00 | 40 | 60 | 100 |
| Non-moss | 1 | 10 | 5 | 37.50 | 9.09 | 90.91 | 62.50 |
| Healthy | 3 | 1 | 24 | 14.28 | 17.24 | 82.76 | 85.72 |
| Total | 10 | 11 | 29 | — | — | — | — |
| Overall Accuracy: 80.00% | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Wu, W.; Li, X.; Xu, X.; Li, J. Vegetation Abundance and Health Mapping Over Southwestern Antarctica Based on WorldView-2 Data and a Modified Spectral Mixture Analysis. Remote Sens. 2021, 13, 166. https://doi.org/10.3390/rs13020166
Sun X, Wu W, Li X, Xu X, Li J. Vegetation Abundance and Health Mapping Over Southwestern Antarctica Based on WorldView-2 Data and a Modified Spectral Mixture Analysis. Remote Sensing. 2021; 13(2):166. https://doi.org/10.3390/rs13020166
Chicago/Turabian StyleSun, Xiaohui, Wenjin Wu, Xinwu Li, Xiyan Xu, and Jinfeng Li. 2021. "Vegetation Abundance and Health Mapping Over Southwestern Antarctica Based on WorldView-2 Data and a Modified Spectral Mixture Analysis" Remote Sensing 13, no. 2: 166. https://doi.org/10.3390/rs13020166
APA StyleSun, X., Wu, W., Li, X., Xu, X., & Li, J. (2021). Vegetation Abundance and Health Mapping Over Southwestern Antarctica Based on WorldView-2 Data and a Modified Spectral Mixture Analysis. Remote Sensing, 13(2), 166. https://doi.org/10.3390/rs13020166