
Influence of the Antimicrobial LL-37 Peptide on Legionella dumoffii Phospholipids Adsorbed at the Air–Liquid Interface

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Culture Conditions
2.2. Lipids Isolation
2.3. Langmuir Monolayer Studies
3. Results
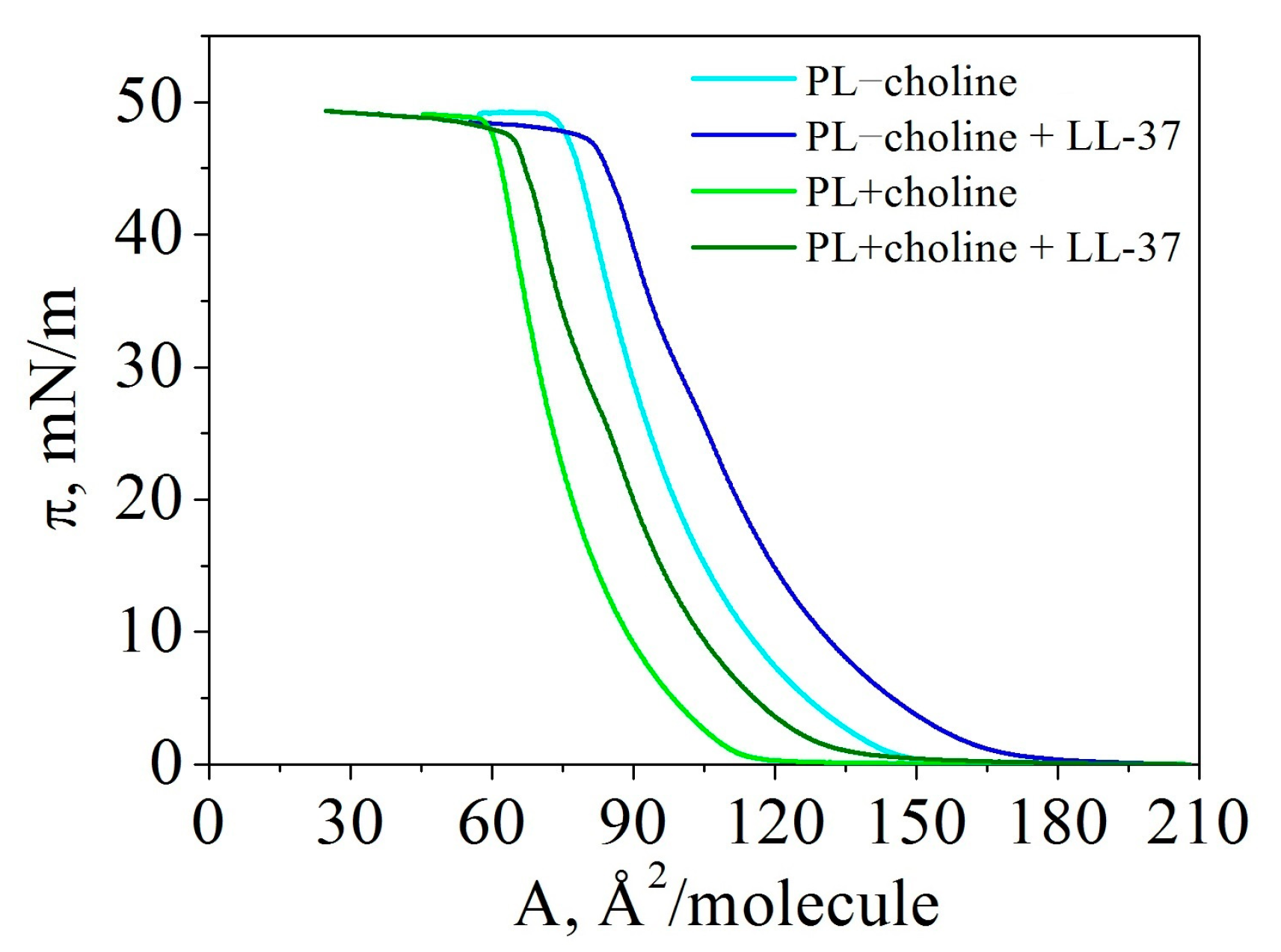
3.1. Isotherms
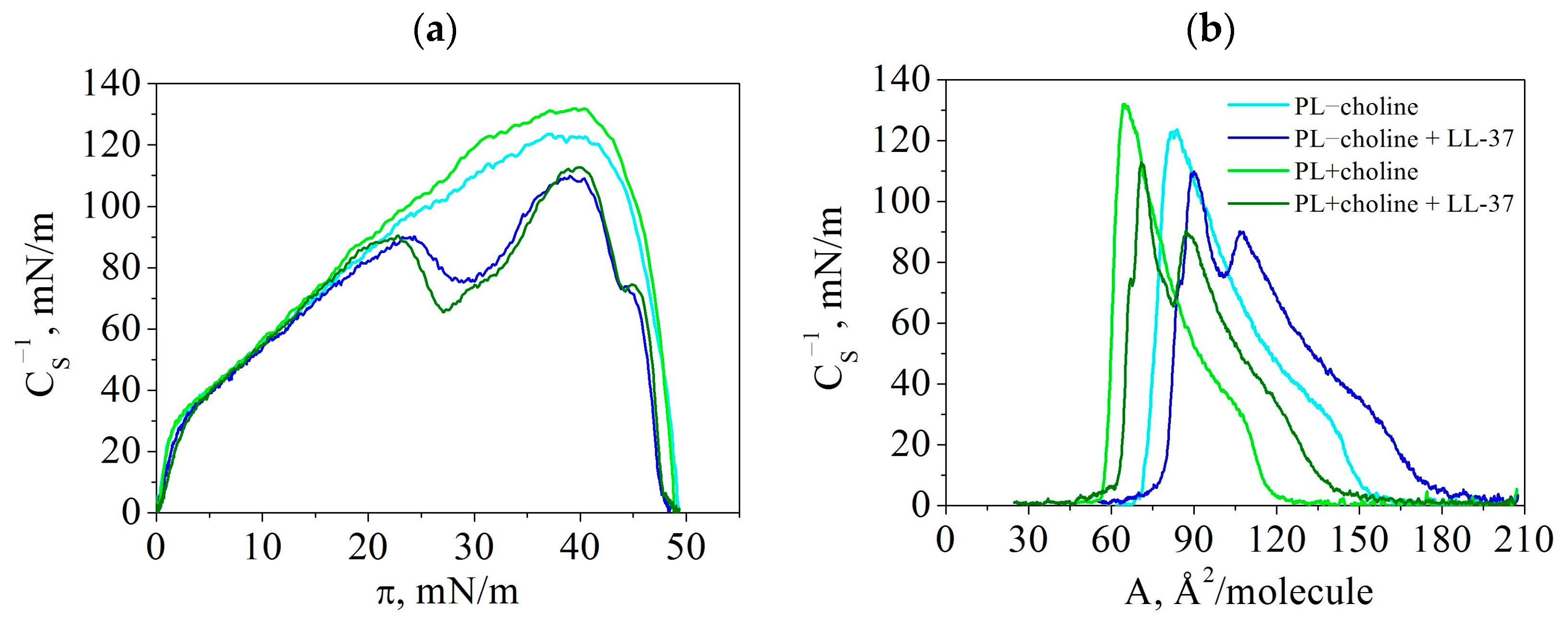
3.2. Compression Modulus (Elasticity)—
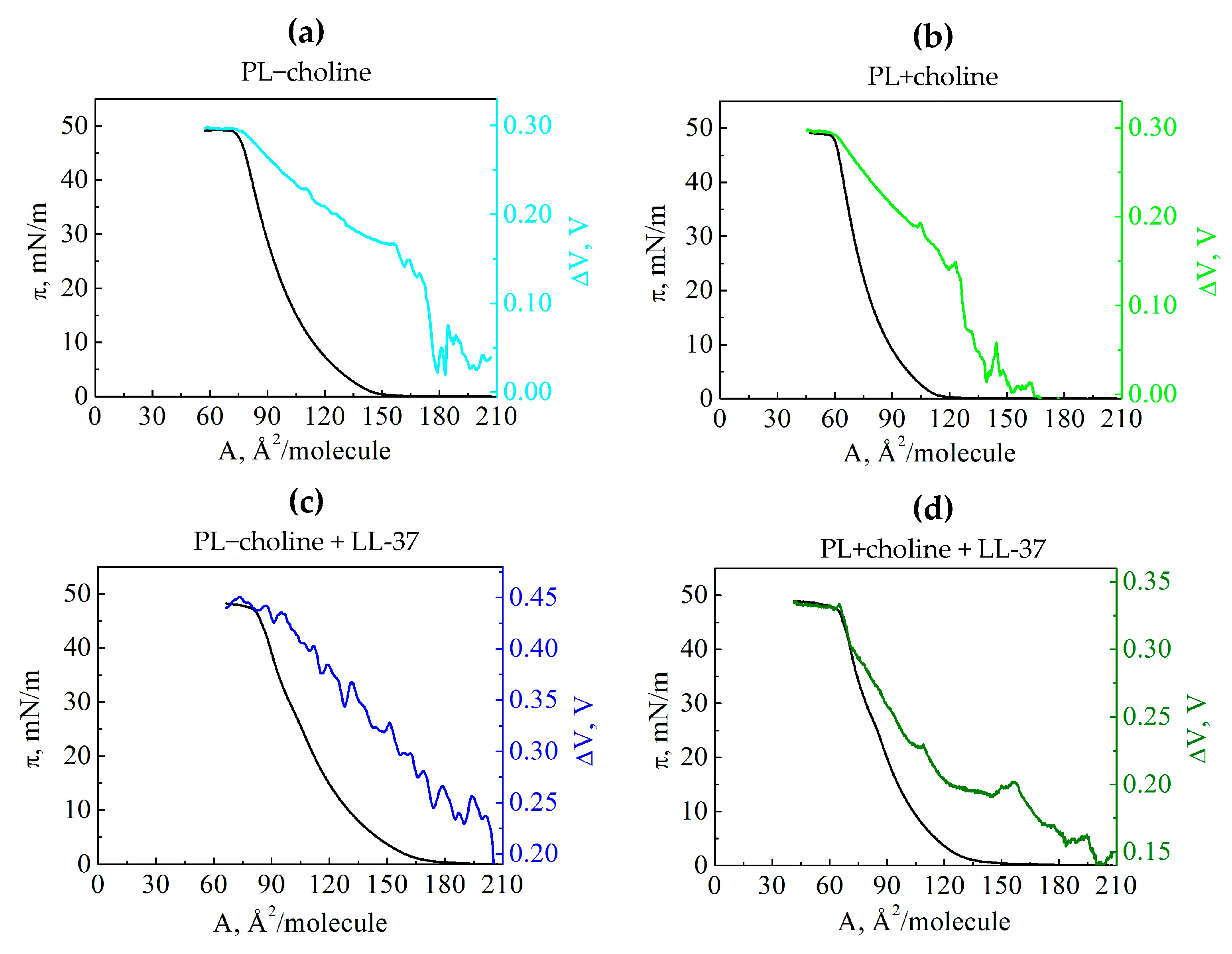
3.3. Surface Potential
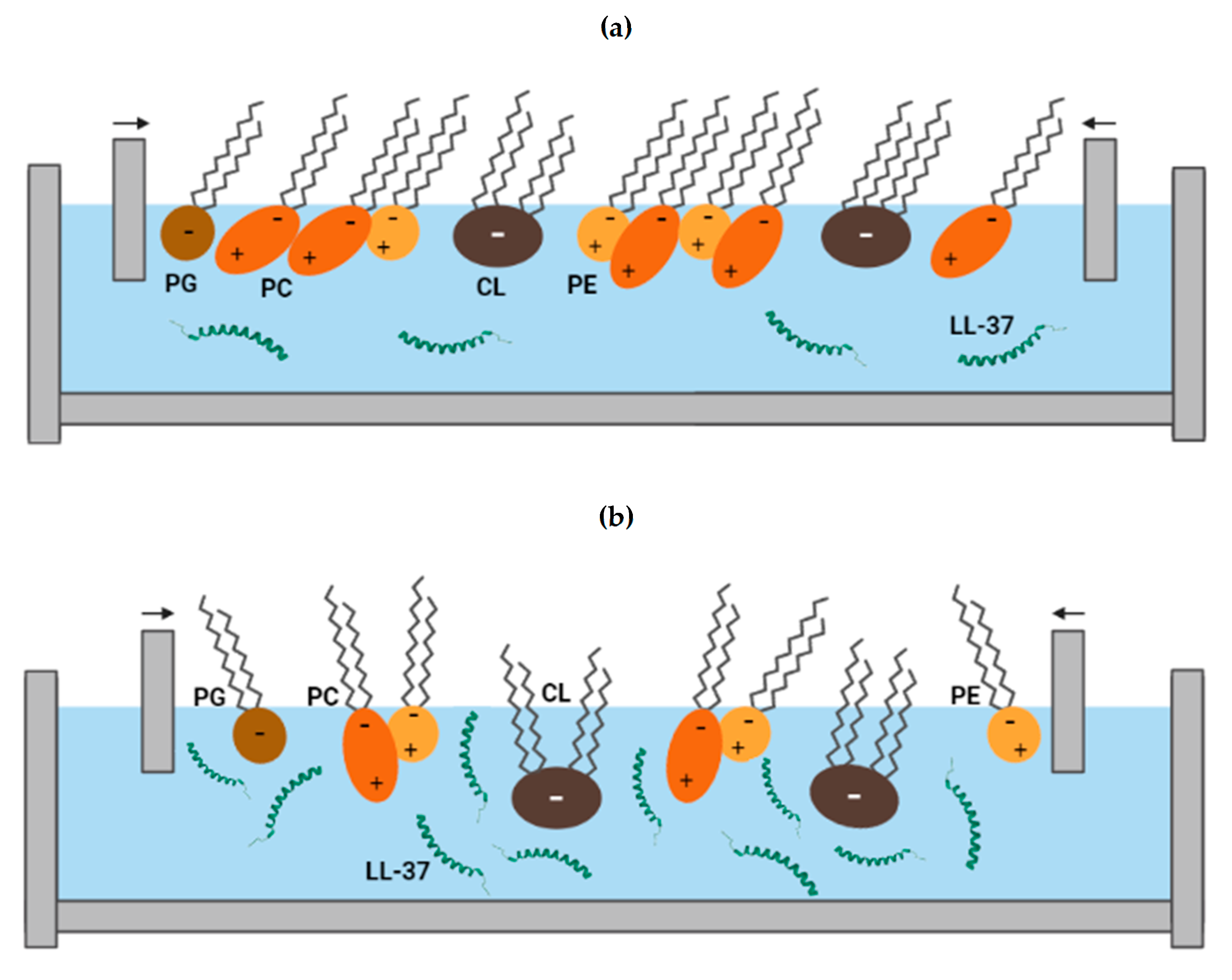
3.4. Surface Morphology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Correia, A.M.; Ferreira, J.S.; Borges, V.; Nunes, A.; Gomes, B.; Capucho, R.; Gonçalves, J.; Antunes, D.M.; Almeida, S.; Mendes, A.; et al. Probable person-to-person transmission of Legionnaires’ disease. N. Engl. J. Med. 2016, 374, 497–498. [Google Scholar] [CrossRef]
- Euzeby, J.P. List of Prokaryotic Names with Standing in Nomenclature Genus Legionella. Available online: https://www.bacterio.net/legionella.html (accessed on 27 February 2023).
- Miyashita, N.; Higa, F.; Aoki, Y.; Kikuchi, T.; Seki, M.; Tateda, K.; Maki, N.; Uchino, K.; Ogasawara, K.; Kiyota, H.; et al. Distribution of Legionella species and serogroups in patients with culture-confirmed Legionella pneumonia. J. Infect. Chemother. 2020, 26, 411–417. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, W.A.; Dhand, L.; Cianciotto, N.P. Infection of macrophage-like cells by Legionella species that have not been associated with disease. Infect. Immun. 1996, 64, 4381–4384. [Google Scholar] [CrossRef] [PubMed]
- Maruta, K.; Miyamoto, H.; Hamada, T.; Ogawa, M.; Taniguchi, H.; Yoshida, S. Entry and intracellular growth of Legionella dumoffii in alveolar epithelial cells. Am. J. Respir. Crit. Care Med. 1998, 157, 1967–1974. [Google Scholar] [CrossRef]
- Bruin, J.P.; Koshkalda, T.; IJzerman, E.P.F.; Luck, C.; Diederen, B.M.W.; Den Boer, J.W.; Mouton, J.W. Isolation of ciprofloxacin-resistant Legionella pneumophila in a patient with severe pneumonia. J. Antimicrob. Chemother. 2014, 69, 2869–2871. [Google Scholar] [CrossRef] [PubMed]
- Ridyard, K.E.; Overhage, J. The potential of human peptide LL-37 as an antimicrobial and anti-biofilm agent. Antibiotics 2021, 10, 650. [Google Scholar] [CrossRef] [PubMed]
- Dutta, I.; Ramesh, S.; Radebe, S.M.; Somboro, A.M.; de la Torre, B.G.; Kruger, H.G.; Essack, S.Y.; Albericio, F.; Govender, T. Optimized microwave assisted synthesis of LL-37, a cathelicidin human antimicrobial peptide. Int. J. Pept. Res. Ther. 2015, 21, 13–20. [Google Scholar] [CrossRef]
- Palusińska-Szysz, M.; Jurak, M.; Gisch, N.; Waldow, F.; Zehethofer, N.; Nehls, C.; Schwudke, D.; Koper, P.; Mazur, A. The human LL-37 peptide exerts antimicrobial activity against Legionella micdadei interacting with membrane phospholipids. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2022, 1867, 159138. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, J.W. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Davies, J.T.; Rideal, E.K. Interfacial Phenomena; Academic Press: New York, NY, USA, 1963. [Google Scholar]
- Adamson, A.W.; Gast, A.P. Physical Chemistry of Surfaces, 6th ed.; Wiley: New York, NY, USA, 1997. [Google Scholar]
- Dynarowicz-Łatka, P.; Milart, P. Synthesis and Langmuir monolayer characterisation of some nitro derivatives of polyphenyl carboxylic acids. J. Chem. Res. 2009, 2009, 225–228. [Google Scholar] [CrossRef]
- Chachaj-Brekiesz, A.; Kobierski, J.; Wnętrzak, A.; Dynarowicz-Łatka, P. Electrical properties of membrane phospholipids in Langmuir monolayers. Membranes 2021, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, H.; Ohki, S. Donnan potential and surface potential of a charged membrane. Biophys. J. 1985, 47, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Vogel, V.; Möbius, D. Local surface potentials and electric dipole moments of lipid monolayers: Contributions of the water/lipid and the lipid/air interfaces. J. Colloid Interface Sci. 1988, 126, 408–420. [Google Scholar] [CrossRef]
- Oliveira, O.N., Jr.; Bonardi, C. The surface potential of Langmuir monolayers revisited. Langmuir 1997, 13, 5920–5924. [Google Scholar] [CrossRef]
- Oliveira, O.N., Jr.; Riul, A., Jr.; Leite, V.B.P. Water at interfaces and its influence on the electrical properties of adsorbed films. Braz. J. Phys. 2004, 34, 73–83. [Google Scholar] [CrossRef]
- Palusińska-Szysz, M.; Zdybicka-Barabas, A.; Reszczyńska, E.; Luchowski, R.; Kania, M.; Gisch, N.; Waldow, F.; Mak, P.; Danikiewicz, W.; Gruszecki, W.I.; et al. The lipid composition of Legionella dumoffii membrane modulates the interaction with Galleria mellonella apolipophorin III. Biochim. Biophys. Acta 2016, 1861, 617–629. [Google Scholar] [CrossRef]
- Caillon, L.; Nieto, V.; Gehan, P.; Omrane, M.; Rodriguez, N.; Monticelli, L.; Thiam, A.R. Triacylglycerols sequester monotopic membrane proteins to lipid droplets. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Möhwald, H. Phospholipid and phospholipid-protein monolayers at the air/water interface. Annu. Rev. Phys. Chem. 1990, 41, 441–476. [Google Scholar] [CrossRef]
- Suetsugu, S.; Kurisu, S.; Takenawa, T. Dynamic shaping of cellular membranes by phospholipids and membrane-deforming proteins. Physiol. Rev. 2014, 94, 1219–1248. [Google Scholar] [CrossRef]
- Ridgway, N.D. Phospholipid synthesis in mammalian cells. In Biochemistry of Lipids, Lipoproteins and Membranes, 6th ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 209–236. [Google Scholar] [CrossRef]
- Leekumjorn, S.; Sum, A.K. Molecular simulation study of structural and dynamic properties of mixed DPPC/DPPE bilayers. Biophys. J. 2006, 90, 3951–3965. [Google Scholar] [CrossRef]
- Phan, M.D.; Shin, K. Effects of cardiolipin on membrane morphology: A Langmuir monolayer study. Biophys. J. 2015, 108, 1977–1986. [Google Scholar] [CrossRef] [PubMed]
- Epand, R.M.; Epand, R.F. Domains in bacterial membranes and the action of antimicrobial agents. Mol. Biosyst. 2009, 6, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Xhindoli, D.; Pacor, S.; Benincasa, M.; Scocchi, M.; Gennaro, R.; Tossi, A. The human cathelicidin LL-37—A pore-forming antibacterial peptide and host-cell modulator. Biochim. Biophys. Acta (BBA)-Biomembr. 2016, 1858, 546–566. [Google Scholar] [CrossRef]
- Neville, F.; Cahuzac, M.; Konovalov, O.; Ishitsuka, Y.; Lee, K.Y.C.; Kuzmenko, I.; Kale, G.M.; Gidalevitz, D. Lipid headgroup discrimination by antimicrobial peptide LL-37: Insight into mechanism of action. Biophys. J. 2006, 90, 1275–1287. [Google Scholar] [CrossRef] [PubMed]
- Sevcsik, E.; Pabst, G.; Richter, W.; Danner, S.; Amenitsch, H.; Lohner, K. Interaction of LL-37 with model membrane systems of different complexity: Influence of the lipid matrix. Biophys. J. 2008, 94, 4688–4699. [Google Scholar] [CrossRef] [PubMed]
- Shahmiri, M.; Enciso, M.; Adda, C.G.; Smith, B.J.; Perugini, M.A.; Mechler, A. Membrane core-specific antimicrobial action of cathelicidin LL-37 peptide switches between pore and nanofibre formation. Sci. Rep. 2016, 6, 38184. [Google Scholar] [CrossRef] [PubMed]
- Den Hertog, A.L.; van Marle, J.; van Veen, H.A.; van’t Hof, W.; Bolscher, J.G.M.; Veerman, E.C.I.; Amerongen, A.V.N. Candidacidal effects of two antimicrobial peptides: Histatin 5 causes small membrane defects, but LL-37 causes massive disruption of the cell membrane. Biochem. J. 2005, 388, 689–695. [Google Scholar] [CrossRef]
- Wildman, K.A.H.; Ramamoorthy, D.-K.L.A. Mechanism of lipid bilayer disruption by the human antimicrobial peptide, LL-37. Biochemistry 2003, 42, 6545–6558. [Google Scholar] [CrossRef]
- Febriza, A.; Hatta, M.; Natzir, R.; Kasim, V.N.A.; Idrus, H.H. Activity of antimicrobial peptide, cathelicidin, on bacterial infection. Open Biochem. J. 2019, 13, 45–53. [Google Scholar] [CrossRef]
- Ciumac, D.; Campbell, R.A.; Xu, H.; Clifton, L.A.; Hughes, A.V.; Webster, J.R.; Lu, J.R. Implications of lipid monolayer charge characteristics on their selective interactions with a short antimicrobial peptide. Colloids Surf. B Biointerfaces 2017, 150, 308–316. [Google Scholar] [CrossRef]
- Walker, L.R.; Marty, M.T. Lipid tails modulate antimicrobial peptide membrane incorporation and activity. Biochim. Biophys. Acta (BBA)-Biomembr. 2022, 1864, 183870. [Google Scholar] [CrossRef] [PubMed]
- Rashid, R.; Veleba, M.; Kline, K.A. Focal targeting of the bacterial envelope by antimicrobial peptides. Front. Cell Dev. Biol. 2016, 4, 55. [Google Scholar] [CrossRef] [PubMed]
- Sevcsik, E.; Pabst, G.; Jilek, A.; Lohner, K. How lipids influence the mode of action of membrane-active peptides. Biochim. Biophys. Acta (BBA)-Biomembr. 2007, 1768, 2586–2595. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monolayer | PL−choline | PL−choline + LL-37 | PL+choline | PL+choline + LL-37 |
| (/molecule) | 149.0 ± 2.0 | 175.0 ± 2.0 | 115.0 ± 2.0 | 147.0 ± 2.0 |
| (mN/m) | 49.2 ± 0.4 | 47.9 ± 0.7 | 48.8 ± 0.3 | 48.2 ± 0.1 |
| Monolayer | PL−choline | PL−choline + LL-37 | PL+choline | PL+choline + LL-37 |
| (mN/m) | 124 | 90 | 132 | 90 |
| (mN/m) | 37 | 24 | 39 | 23 |
| (/molecule) | 84 | 107 | 65 | 87 |
| (mN/m) | - | 110 | - | 113 |
| (mN/m) | - | 39 | - | 40 |
| (/molecule) | - | 90 | - | 71 |
| (mN/m) | - | 75 | - | 66 |
| (mN/m) | - | 29 | - | 27 |
| (/molecule) | - | 101 | - | 83 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pastuszak, K.; Kowalczyk, B.; Tarasiuk, J.; Jurak, M.; Palusińska-Szysz, M. Influence of the Antimicrobial LL-37 Peptide on Legionella dumoffii Phospholipids Adsorbed at the Air–Liquid Interface. Sustainability 2023, 15, 6670. https://doi.org/10.3390/su15086670
Pastuszak K, Kowalczyk B, Tarasiuk J, Jurak M, Palusińska-Szysz M. Influence of the Antimicrobial LL-37 Peptide on Legionella dumoffii Phospholipids Adsorbed at the Air–Liquid Interface. Sustainability. 2023; 15(8):6670. https://doi.org/10.3390/su15086670
Chicago/Turabian StylePastuszak, Katarzyna, Bożena Kowalczyk, Jacek Tarasiuk, Małgorzata Jurak, and Marta Palusińska-Szysz. 2023. "Influence of the Antimicrobial LL-37 Peptide on Legionella dumoffii Phospholipids Adsorbed at the Air–Liquid Interface" Sustainability 15, no. 8: 6670. https://doi.org/10.3390/su15086670
APA StylePastuszak, K., Kowalczyk, B., Tarasiuk, J., Jurak, M., & Palusińska-Szysz, M. (2023). Influence of the Antimicrobial LL-37 Peptide on Legionella dumoffii Phospholipids Adsorbed at the Air–Liquid Interface. Sustainability, 15(8), 6670. https://doi.org/10.3390/su15086670