



Insect Pest Pheromone Lures May Enhance the Activity of Insectivorous Bats in Mediterranean Vineyards and Apple Orchards



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
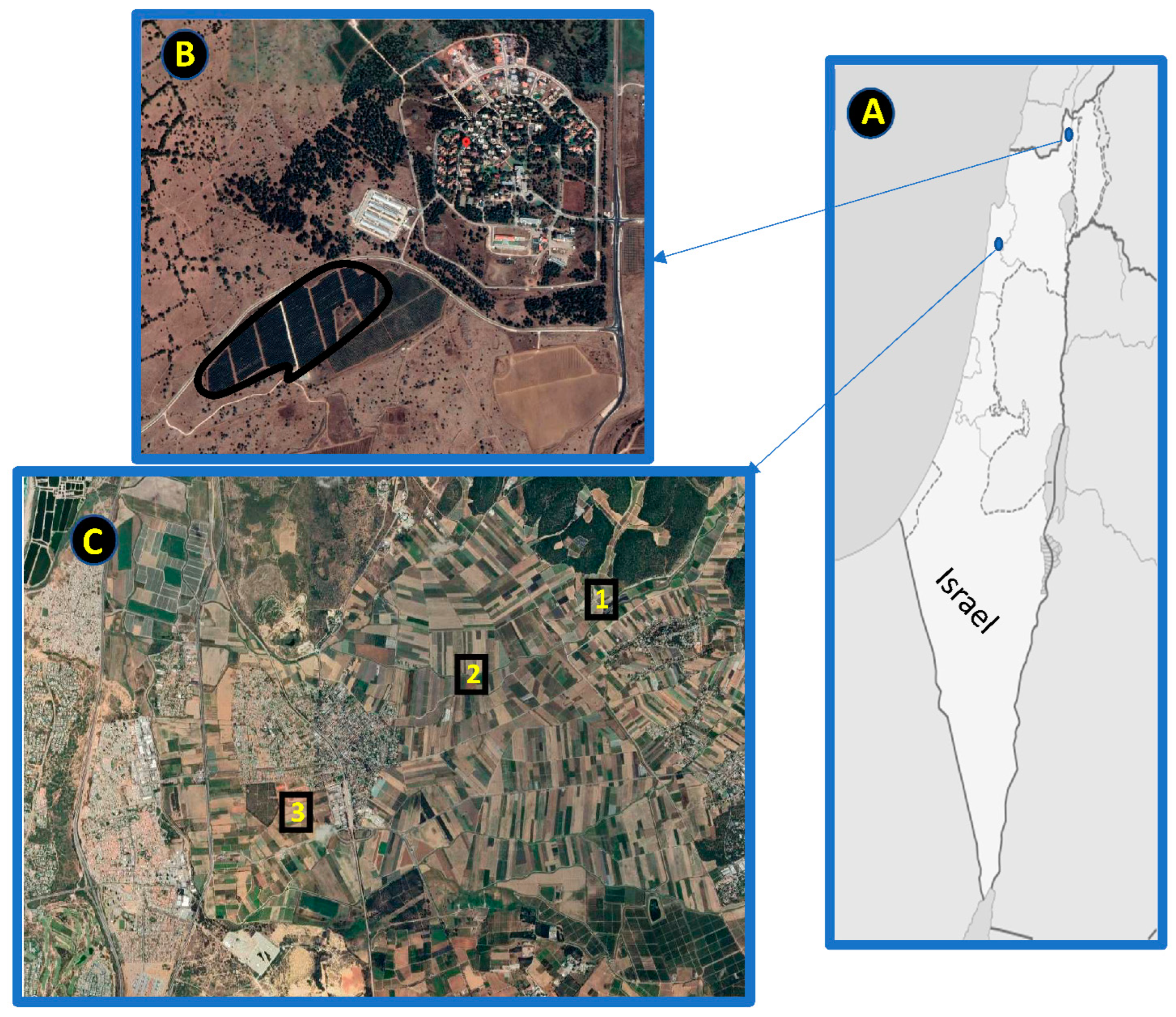
2.1. Study Areas and Bat Diversity
2.2. Experimental Design
2.3. Acoustic Bat Monitoring
2.4. Data Analysis
3. Results
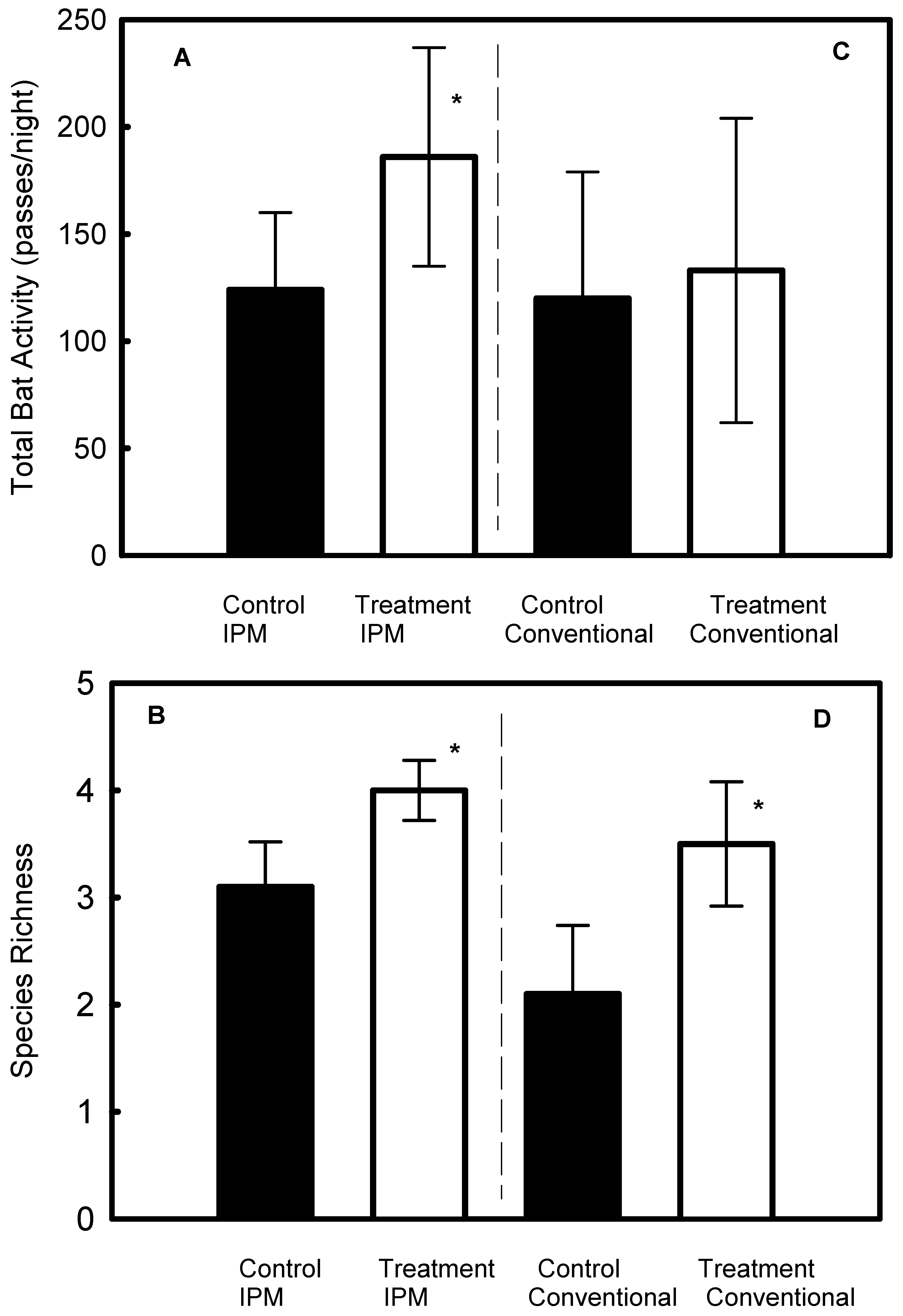
3.1. Effect of Pheromone Treatment in IPM and Conventional Vineyards
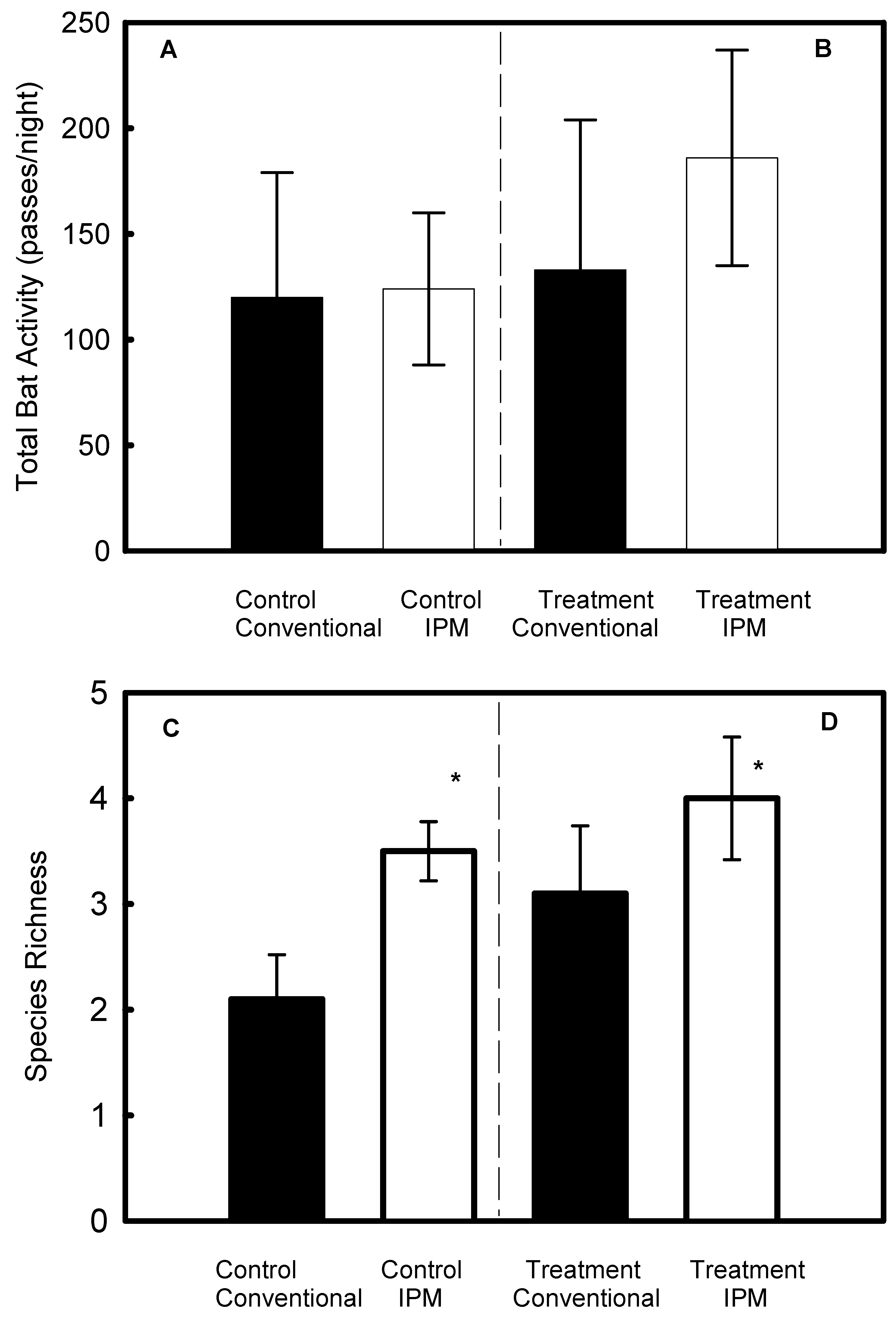
3.2. Comparison between IPM Vineyards and Conventional Vineyards
3.3. Community Structure
3.4. Apple Orchards
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671. [Google Scholar] [CrossRef] [PubMed]
- Kogan, M.; Bajwa, W.I. Integrated pest management: A global reality? Ann. Soc. Entomol. Bras. 1999, 28, 01–25. [Google Scholar] [CrossRef]
- Burkholder, W.E.; Ma, M. Pheromones for monitoring and control of stored-product insects. Ann. Rev. Entomol. 1985, 30, 257–272. [Google Scholar] [CrossRef]
- Witzgall, P.; Kirsch, P.; Cork, A. Sex pheromones and their impact on pest management. J. Chem. Ecol. 2010, 36, 80–100. [Google Scholar] [CrossRef]
- Stenberg, J.A. A conceptual framework for integrated pest management. Trends. Plant Sci. 2017, 22, 759–769. [Google Scholar] [CrossRef]
- Cardé, R.T.; Minks, A.K. Insect Pheromone Research: New Directions; Chapman and Hall: New York, NY, USA, 1977. [Google Scholar]
- Landolt, P.J. Sex attractant and aggregation pheromones of male phytophagous insects. Am. Entomol. 1997, 43, 12–22. [Google Scholar] [CrossRef]
- Rydell, J. Exploitation of insects around streetlamps by bats in Sweden. Func. Ecol. 1992, 6, 744–750. [Google Scholar] [CrossRef]
- Charbonnier, Y.; Barbaro, L.; Theillout, A.; Jactel, H. Numerical and functional responses of forest bats to a major insect pest in pine plantations. PLoS ONE 2014, 9, e109488. [Google Scholar] [CrossRef]
- Kunz, T.H.; Braun de Torrez, E.; Bauer, D.; Lobova, T.; Fleming, T.H. Ecosystem services provided by bats. Ann. N. Y. Acad. Sci. 2011, 1223, 1–38. [Google Scholar] [CrossRef]
- Cleveland, C.J.; Betke, M.; Federico, P.; Frank, J.D.; Hallam, T.G.; Horn, J.; Lopez, J.D., Jr; McCracken, G.F.; Medellín, R.A.; Moreno-Valdez, A.; et al. Economic value of the pest control service provided by Brazilian free-tailed bats in south-central Texas. Front Ecol. Environ. 2006, 4, 238–243. [Google Scholar] [CrossRef]
- Maine, J.J.; Boyles, J.G. Bats initiate vital agroecological interactions in corn. Proc. Nat. Acad. Sci. USA 2015, 112, 12438–12443. [Google Scholar] [CrossRef] [PubMed]
- Cohen, Y.; Bar-David, S.; Nielsen, M.; Bohmann, K.; Korine, C. An appetite for pests: Synanthropic insectivorous bats exploit cotton pest irruptions and consume various deleterious arthropods. Mol. Ecol. 2020, 29, 1185–1198. [Google Scholar] [CrossRef] [PubMed]
- Kolkert, H.; Smith, R.; Rader, R.; Reid, N. Insectivorous bats provide significant economic value to the Australian cotton industry. Ecosyst. Serv. 2021, 49, 101280. [Google Scholar] [CrossRef]
- Puig-Montserrat, X.; Flaquer, C.; Gómez-Aguilera, N.; Burgas, A.; Mas, M.; Tuneu, C.; López-Baucells, A. Bats actively prey on mosquitoes and other deleterious insects in rice paddies: Potential impact on human health and agriculture. Pest Manag. Sci. 2020, 76, 3759–3769. [Google Scholar] [CrossRef] [PubMed]
- Brown, V.A.; de Torrez, E.B.; McCracken, G.F. Crop pests eaten by bats in organic pecan orchards. Crop Prot. 2015, 67, 66–71. [Google Scholar] [CrossRef]
- Taylor, P.J.; Grass, I.; Alberts, A.J.; Joubert, E.; Tscharntke, T. Economic value of bat predation services—A review and new estimates from macadamia orchards. Ecosyst. Serv. 2018, 30, 372–381. [Google Scholar] [CrossRef]
- Williams-Guillén, K.; Perfecto, I.; Vandermeer, J. Bats limit insects in a neotropical agroforestry system. Science 2008, 320, 70. [Google Scholar] [CrossRef]
- Froidevaux, J.S.; Louboutin, B.; Jones, G. Does organic farming enhance biodiversity in Mediterranean vineyards? A case study with bats and arachnids. Agric. Ecosyst. Environ. 2017, 249, 112–122. [Google Scholar] [CrossRef]
- Rodríguez-San Pedro, A.; Allendes, J.L.; Beltrán, C.A.; Chaperon, P.N.; Saldarriaga-Córdoba, M.M.; Silva, A.X.; Grez, A.A. Quantifying ecological and economic value of pest control services provided by bats in a vineyard landscape of central Chile. Agric. Ecosyst. Environ. 2020, 302, 107063. [Google Scholar] [CrossRef]
- Baroja, U.; Garin, I.; Vallejo, N.; Aihartza, J.; Rebelo, H.; Goiti, U. Bats actively track and prey on grape pest populations. Ecol. Indic. 2021, 126, 107718. [Google Scholar] [CrossRef]
- Jay, M.; de Roincé, C.B.; Ricard, J.M.; Garcin, A.; Mandrin, J.F.; Lavigne, C.; Bouvier, J.C.; Tupinier, Y.; Puechmaille, S. Functional biodiversity in apple orchards: Are the bats eating the pests? Infos-Ctifl 2012, 86, 28–34. [Google Scholar]
- Wickramasinghe, L.P.; Harris, S.; Jones, G.; Vaughan, N. Bat activity and species richness on organic and conventional farms: Impact of agricultural intensification. J. App. Ecol. 2003, 40, 984–993. [Google Scholar] [CrossRef]
- Puig-Montserrat, X.; Torre, I.; López-Baucells, A.; Guerrieri, E.; Monti, M.M.; Ràfols-García, R.; Ferrer, X.; Gisbert, D.; Flaquer, C. Pest control service provided by bats in Mediterranean rice paddies: Linking agroecosystems structure to ecological functions. Mamm. Biol. 2015, 80, 237–245. [Google Scholar] [CrossRef]
- Krings, C.H.; Darras, K.; Hass, A.; Batáry, P.; Fabian, Y. Not only hedgerows, but also flower fields can enhance bat activity in intensively used agricultural landscapes. Basic Appl. Ecol. 2022, 63, 23–35. [Google Scholar] [CrossRef]
- Russo, D.; Bosso, L.; Ancillotto, L. Novel perspectives on bat insectivory highlight the value of this ecosystem service in farmland: Research frontiers and management implications. Agric. Ecosyst. Environ. 2018, 266, 31–38. [Google Scholar] [CrossRef]
- Bayat, S.; Geiser, F.; Kristiansen, P.; Wilson, S.C. Organic contaminants in bats: Trends and new issues. Environ. Int. 2014, 63, 40–52. [Google Scholar] [CrossRef]
- Park, K.J. Mitigating the impacts of agriculture on biodiversity: Bats and their potential role as bioindicators. Mamm. Biol. 2015, 80, 191–204. [Google Scholar] [CrossRef]
- Williams-Guillén, K.; Olimpi, E.; Maas, B.; Taylor, P.J.; Arlettaz, R. Bats in the anthropogenic matrix: Challenges and opportunities for the conservation of Chiroptera and their ecosystem services in agricultural landscapes. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C.C., Kingston, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 151–186. [Google Scholar]
- Kahnonitch, I.; Lubin, Y.; Korine, C. Insectivorous bats in semi-arid agroecosystems− effects on foraging activity and implications for insect pest control. Agric. Ecosyst. Environ. 2018, 261, 80–92. [Google Scholar] [CrossRef]
- Pasek, J.E. Influence of wind and windbreaks on local dispersal of insects. Agric. Ecosyst. Environ. 1988, 2, 539–554. [Google Scholar] [CrossRef]
- Boughey, K.L.; Lake, I.R.; Haysom, K.A.; Dolman, P.M. Improving the biodiversity benefits of hedgerows: How physical characteristics and the proximity of foraging habitat affect the use of linear features by bats. Biol. Cons. 2011, 144, 1790–1798. [Google Scholar] [CrossRef]
- Angell, R.L.; Langton, S.D.; MacDonald, M.A.; Skates, J.; Haysom, K.A. The effect of a Welsh agri-environment scheme on bat activity: A large-scale study. Agric. Ecosyst. Environ. 2019, 275, 32–41. [Google Scholar] [CrossRef]
- Froidevaux, J.S.; Boughey, K.L.; Hawkins, C.L.; Broyles, M.; Jones, G. Managing hedgerows for nocturnal wildlife: Do bats and their insect prey benefit from targeted agri-environment schemes? J. Appl. Ecol. 2019, 56, 1610–1623. [Google Scholar] [CrossRef]
- Korine, C.; Niv, A.; Axelrod, M.; Dahan, T. Species richness and activity of insectivorous bats in cotton fields in semi-arid and mesic Mediterranean agroecosystems. Mamm. Biol. 2020, 100, 3–80. [Google Scholar] [CrossRef]
- Tuttle, S.R.; Chambers, C.L.; Theimer, T.C. Potential effects of livestock water-troughs modifications on bats in Northern Arizona. Wildl. Soc. Bull. 2006, 34, 602–608. [Google Scholar] [CrossRef]
- Korine, C.; Adams, R.; Russo, D.; Fisher-Phelps, M.; Jacobs, D. Bats and water: Anthropogenic alterations threaten global bat populations. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C.C., Kingston, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 215–241. [Google Scholar]
- Yom-Tov, Y.; Kadmon, R. Analysis of the Distribution of Insectivorous Bats in Israel. Divers. Distrib. 1998, 4, 63–70. [Google Scholar] [CrossRef]
- Dolev, A.; Perevolotsky, A. Vertebrates in Israel: The Red Book; Israel Nature and Parks Authority: Jerusalem, Israel, 2004. [Google Scholar]
- Jacobson, M. Insect Sex Pheromones; Elsevier: Cambridge, UK, 2012. [Google Scholar]
- Cvikel, N.; Berg, K.E.; Levin, E.; Hurme, E.; Borissov, I.; Boonman, A.; Amichai, E.; Yovel, Y. Bats aggregate to improve prey search but might be impaired when their density becomes too high. Curr. Biol. 2015, 25, 206–211. [Google Scholar] [CrossRef]
- Harari, A.R.; Zahavi, T.; Gordon, D.; Anshelevich, L.; Harel, M.; Ovadia, S.; Dunkelblum, E. Pest management programmes in vineyards using male mating disruption. Pest Manag. Sci. Former. Pestic. Sci. 2007, 63, 769–775. [Google Scholar] [CrossRef]
- Gordon, D.; Zahavi, T.; Anshelevich, L.; Harel, M.; Ovadia, S.; Dunkelblum, E.; Harari, A.R. Mating disruption of Lobesia botrana (Lepidoptera: Tortricidae): Effect of pheromone formulations and concentrations. J. Econ. Entomol. 2005, 98, 135–142. [Google Scholar] [CrossRef]
- Ju, D.; Mota-Sanchez, D.; Fuentes-Contreras, E.; Zhang, Y.L.; Wang, X.Q.; Yang, X.Q. Insecticide resistance in the Cydia pomonella (L.): Global status, mechanisms, and research directions. Pestic. Biochem. Physiol. 2021, 178, 104925. [Google Scholar] [CrossRef]
- Gavara, A.; Navarro-Llopis, V.; Primo, J.; Vacas, S. Influence of weather conditions on Lobesia botrana (Lepidoptera: Tortricidae) mating disruption dispensers’ emission rates and efficacy. Crop Protec. 2022, 155, 105926. [Google Scholar] [CrossRef]
- Fenton, M.B. A technique for monitoring bat activity with results obtained from different environments in southern Ontario. Can. J. Zool. 1970, 48, 847–851. [Google Scholar] [CrossRef]
- Benda, P.; Dietz, C.; Andreas, M.; Hotovy, J.; Lucan, R.K.; Maltby, A.; Meakin, K.; Truscott, J.; Vallo, P. Bats (Mammalia Chiroptera) of the Eastern Mediterranean and Middle East. Part 6. Bats of Sinai (Egypt) with some taxonomic, ecologic and echolocation data on this fauna. Acta Soc. Zool. Bohem. 2008, 72, 1–103. [Google Scholar]
- Russo, D.; Jones, G. Identification of twenty-two bat species (Mammalia: Chiroptera) from Italy by analysis of time-expanded recordings of echolocation calls. J. Zool. 2002, 258, 91–103. [Google Scholar] [CrossRef]
- Hackett, T.D.; Holderied, M.W.; Korine, C. Echolocation call description of 15 species of Middle-Eastern desert dwelling insectivorous bats. Bioacoustics 2017, 26, 217–235. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Wagner, H. Vegan: Community Ecology Package, R Package Version 2.4-1; 2016. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 1 December 2021).
- Anderson, M.J. Permutational multivariate analysis of variance (PERMANOVA). Wiley Statsref: Statistics Reference Online. 2015. Available online: https://onlinelibrary.wiley.com/doi/full/10.1002/9781118445112.stat07841 (accessed on 1 December 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 1 December 2021).
- McCracken, G.F.; Westbrook, J.K.; Brown, V.A.; Eldridge, M.; Federico, P.; Kunz, T.H. Bats track and exploit changes in insect pest populations. PLoS ONE 2012, 7, e43839. [Google Scholar] [CrossRef]
- Blažek, J.; Konečný, A.; Bartonička, T. Bat aggregational response to pest caterpillar emergence. Sci. Rep. 2021, 11, 13634. [Google Scholar] [CrossRef]
- Welter, S.; Pickel, C.; Millar, J.; Cave, F.; Van Steenwyk, R.; Dunley, J. Pheromone mating disruption offers selective management options for key pests. Calif. Agric. 2005, 59, 16–22. [Google Scholar] [CrossRef]
- Ayelo, P.M.; Pirk, C.W.; Yusuf, A.A.; Chailleux, A.; Mohamed, S.A.; Deletre, E. Exploring the kairomone-based foraging behaviour of natural enemies to enhance biological control: A review. Front. Ecol. Evol. 2021, 9, 143. [Google Scholar]
- Dicke, M.; Sabelis, M.W. Infochemical terminology: Based on cost-benefit analysis rather than origin of compounds? Func. Ecol. 1988, 2, 131–139. [Google Scholar] [CrossRef]
- Teeling, E.C.; Jones, G.; Rossiter, S.J. Phylogeny, genes, and hearing: Implications for the evolution of echolocation in bats. In Bat Bioacoustics; Fenton, M.B., Grinnell, A.D., Popper, A.N., Fay, R.R., Eds.; Springer: New York, NY, USA, 2016; pp. 25–54. [Google Scholar]
- Brokaw, A.F.; Smotherman, M. Role of ecology in shaping external nasal morphology in bats and implications for olfactory tracking. PLoS ONE 2020, 15, e0226689. [Google Scholar] [CrossRef]
- Razgour, O.; Korine, C.; Saltz, D. Pond characteristics as determinants of species diversity and community composition in desert bats. Anim. Conserv. 2010, 13, 505–513. [Google Scholar] [CrossRef]
- Bommarco, R.; Miranda, F.; Bylund, H.; Björkman, C. Insecticides suppress natural enemies and increase pest damage in cabbage. J. Econ. Entomol. 2011, 104, 782–791. [Google Scholar] [CrossRef] [PubMed]
- El-Wakeil, N.; Gaafar, N.; Sallam, A.; Volkmar, C. Side effects of insecticides on natural enemies and possibility of their integration in plant protection strategies. In Insecticides: Development of Safer and More Effective Technologies Agricultural and Biological Sciences; Trdan, S., Ed.; Tech Open Access Publisher: Rijeka, Croatia, 2013; pp. 1–56. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korine, C.; Cohen, Y.; Kahnonitch, I. Insect Pest Pheromone Lures May Enhance the Activity of Insectivorous Bats in Mediterranean Vineyards and Apple Orchards. Sustainability 2022, 14, 16566. https://doi.org/10.3390/su142416566
Korine C, Cohen Y, Kahnonitch I. Insect Pest Pheromone Lures May Enhance the Activity of Insectivorous Bats in Mediterranean Vineyards and Apple Orchards. Sustainability. 2022; 14(24):16566. https://doi.org/10.3390/su142416566
Chicago/Turabian StyleKorine, Carmi, Yuval Cohen, and Idan Kahnonitch. 2022. "Insect Pest Pheromone Lures May Enhance the Activity of Insectivorous Bats in Mediterranean Vineyards and Apple Orchards" Sustainability 14, no. 24: 16566. https://doi.org/10.3390/su142416566
APA StyleKorine, C., Cohen, Y., & Kahnonitch, I. (2022). Insect Pest Pheromone Lures May Enhance the Activity of Insectivorous Bats in Mediterranean Vineyards and Apple Orchards. Sustainability, 14(24), 16566. https://doi.org/10.3390/su142416566