
Invasive Apple Snail Diets in Native vs. Non-Native Habitats Defined by SIAR (Stable Isotope Analysis in R)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Collection Sites
2.2. Pomacea canaliculata, Aquatic and Riparian Plants, and Detritus Collections
2.3. Planktonic Organism Collection
2.4. Stable Isotope Analysis (13C and 15N)
2.5. SIAR (Stable Isotope Analysis in R)
2.6. Plant Genetic Barcoding
Plant DNA Extraction for Uruguayan and Chinese Plant Species
2.7. Hawaiian Plant Species Identifications
2.8. Pomacea canaliculata Genetic Barcoding
2.9. Polymerase Chain Reaction (PCR) Amplification
2.10. Thermocycler
2.11. Pomacea canaliculata Agarose Gels
2.12. Plant Agarose Gels
3. Results
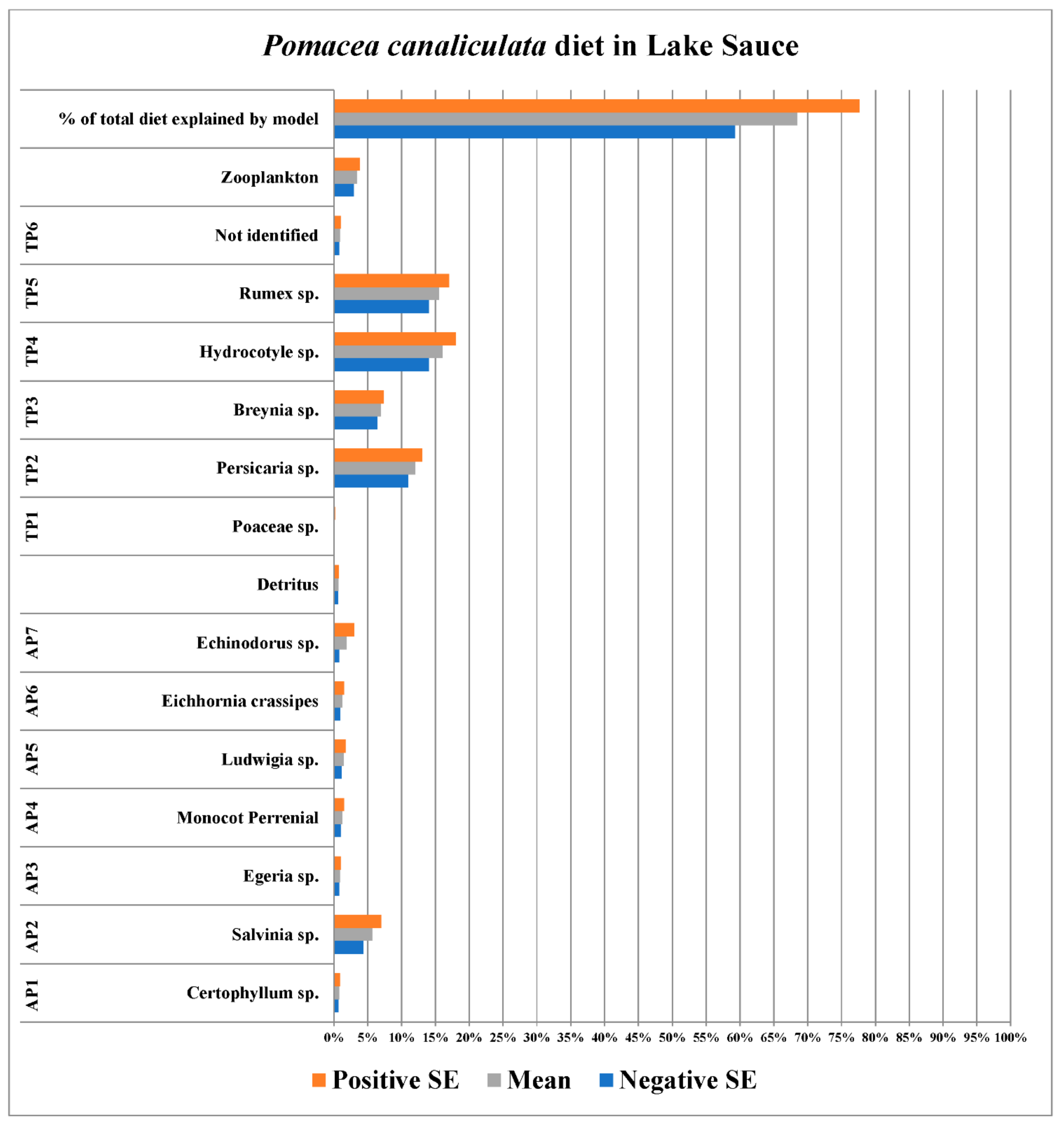
3.1. Lake Sauce (Maldonado, Uruguay)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plants/Plankton/ Detritus | Sample Size (n) | Species ID | Source of Identification |
|---|---|---|---|
| AP1 | 5 | Certophyllum sp. | Sequence Data |
| AP2 | 10 | Salvinia sp. | Sequence Data |
| AP3 | 10 | Egeria sp. | Sequence Data |
| AP4 | 10 | Monocot Perennial | Morphological Characteristics |
| AP5 | 10 | Ludwigia sp. | Sequence Data |
| AP6 | 10 | Eichhornia crassipes | Sequence Data |
| AP7 | 10 | Echinodorus sp. | Sequence Data |
| TP1 | 10 | Poaceae sp. | Sequence Data |
| TP2 | 5 | Persicaria sp. | Sequence Data |
| TP3 | 10 | Breynia sp. | Sequence Data |
| TP4 | 10 | Hydrocotyle sp. | Sequence Data |
| TP5 | 10 | Rumex sp. | Sequence Data |
| TP6 | 5 | Not Identified | N/A |
| Detritus | 5 | N/A | N/A |
| Zooplankton | 6 | N/A | N/A |

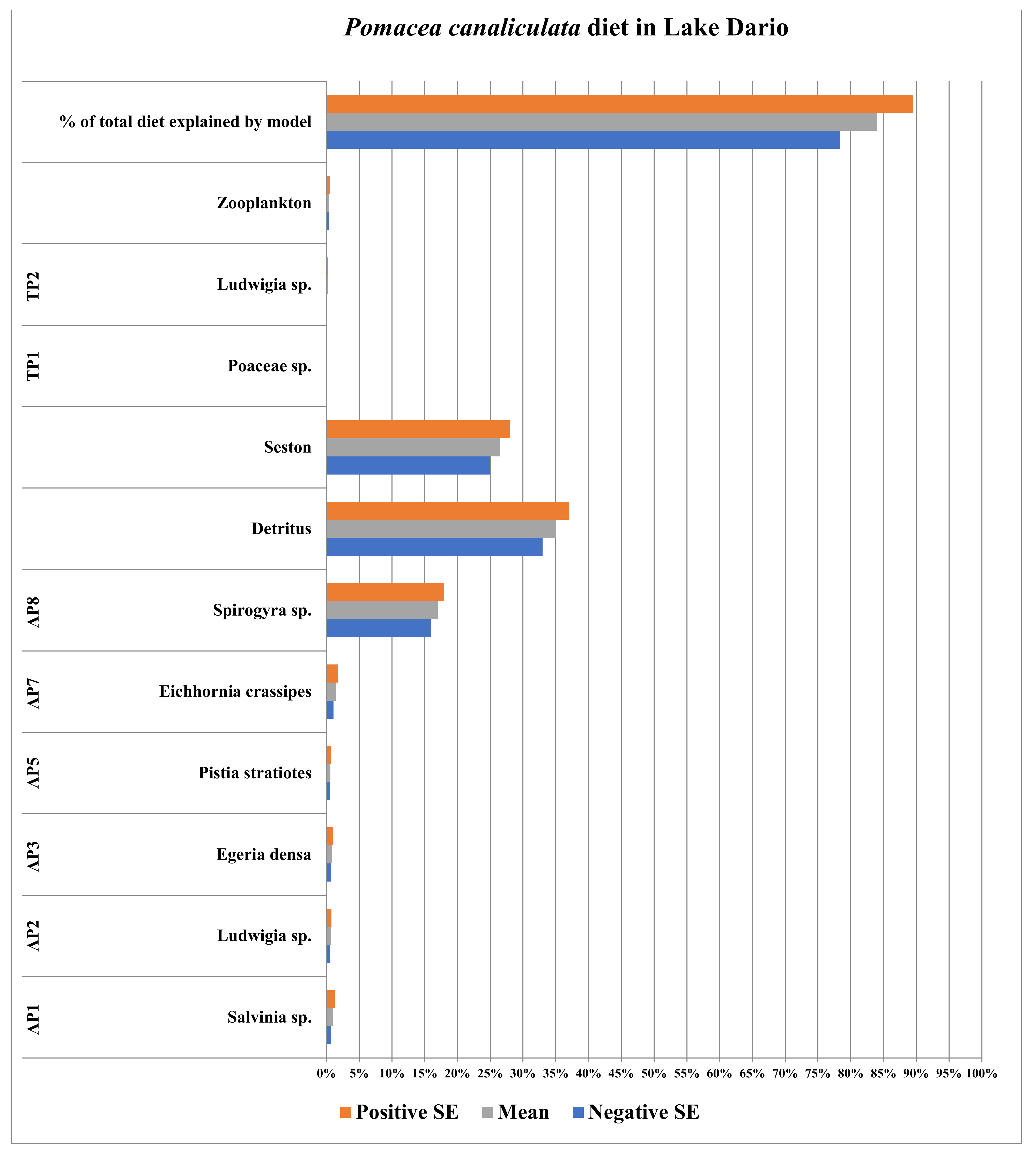
3.2. Lake Dario (Maldonado, Uruguay)
| Plants/Plankton/ Seston/Detritus | Sample Size (n) | Species Identification | Source of Identification |
|---|---|---|---|
| AP1 | 10 | Salvinia sp. | Sequence Data |
| AP2 | 10 | Ludwigia sp. | Sequence Data |
| AP3 | 10 | Egeria densa | Sequence Data |
| AP4 | 10 | Potamogeton sp. | Sequence Data |
| AP5 | 10 | Pistia stratiotes | Sequence Data |
| AP6 | 10 | Potamogeton sp. | Sequence Data |
| AP7 | 10 | Eichhornia Crassipes | Sequence Data |
| AP8 | 2 | Spirogyra sp. (Filamentous Alga) | Microscopy |
| TP1 | 5 | Poaceae sp. | Sequence Data |
| TP2 | 10 | Ludwigia sp. | Sequence Data |
| Detritus | 5 | N/A | N/A |
| Zooplankton | 3 | N/A | N/A |
| Seston | 4 | N/A | N/A |
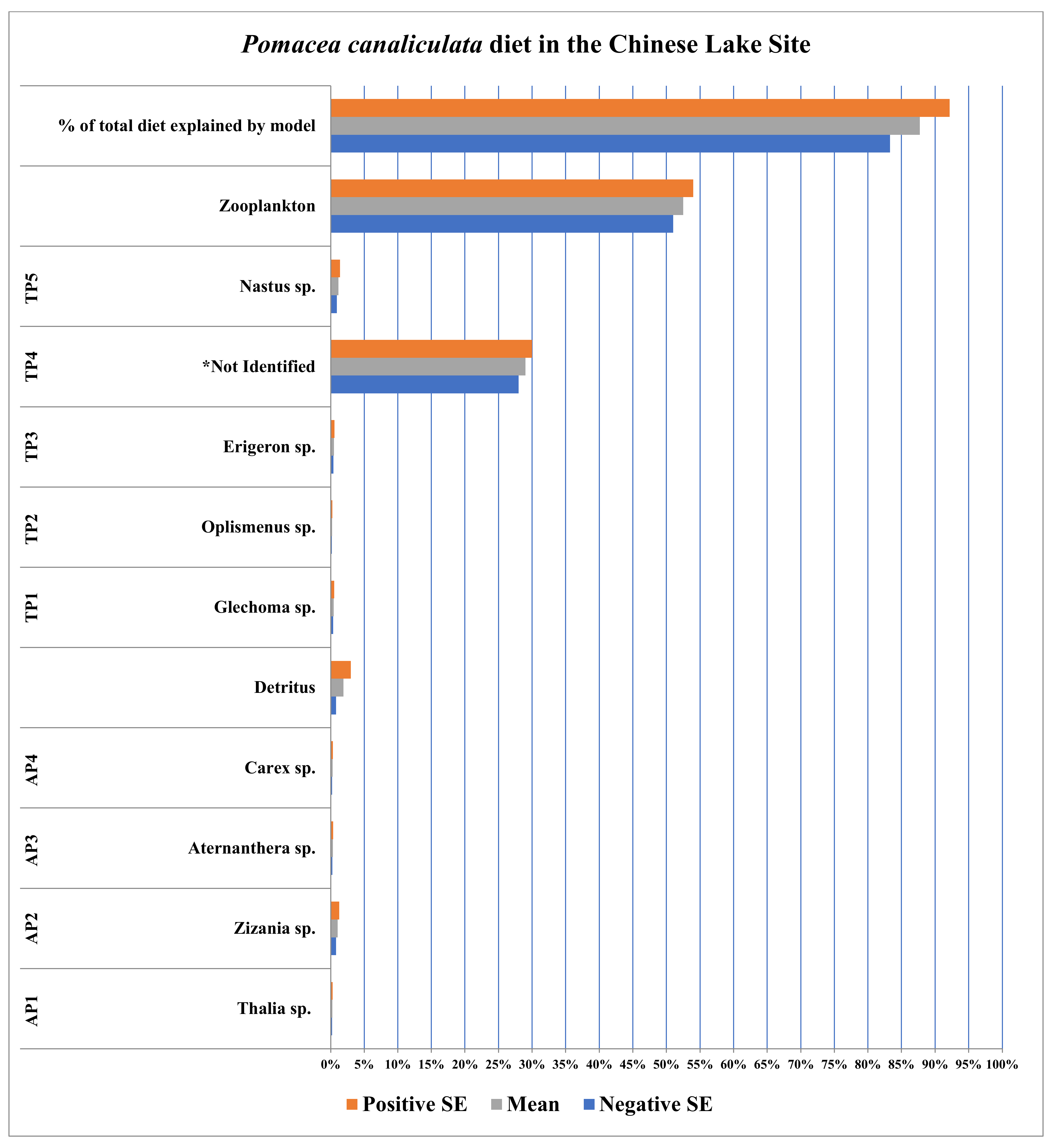
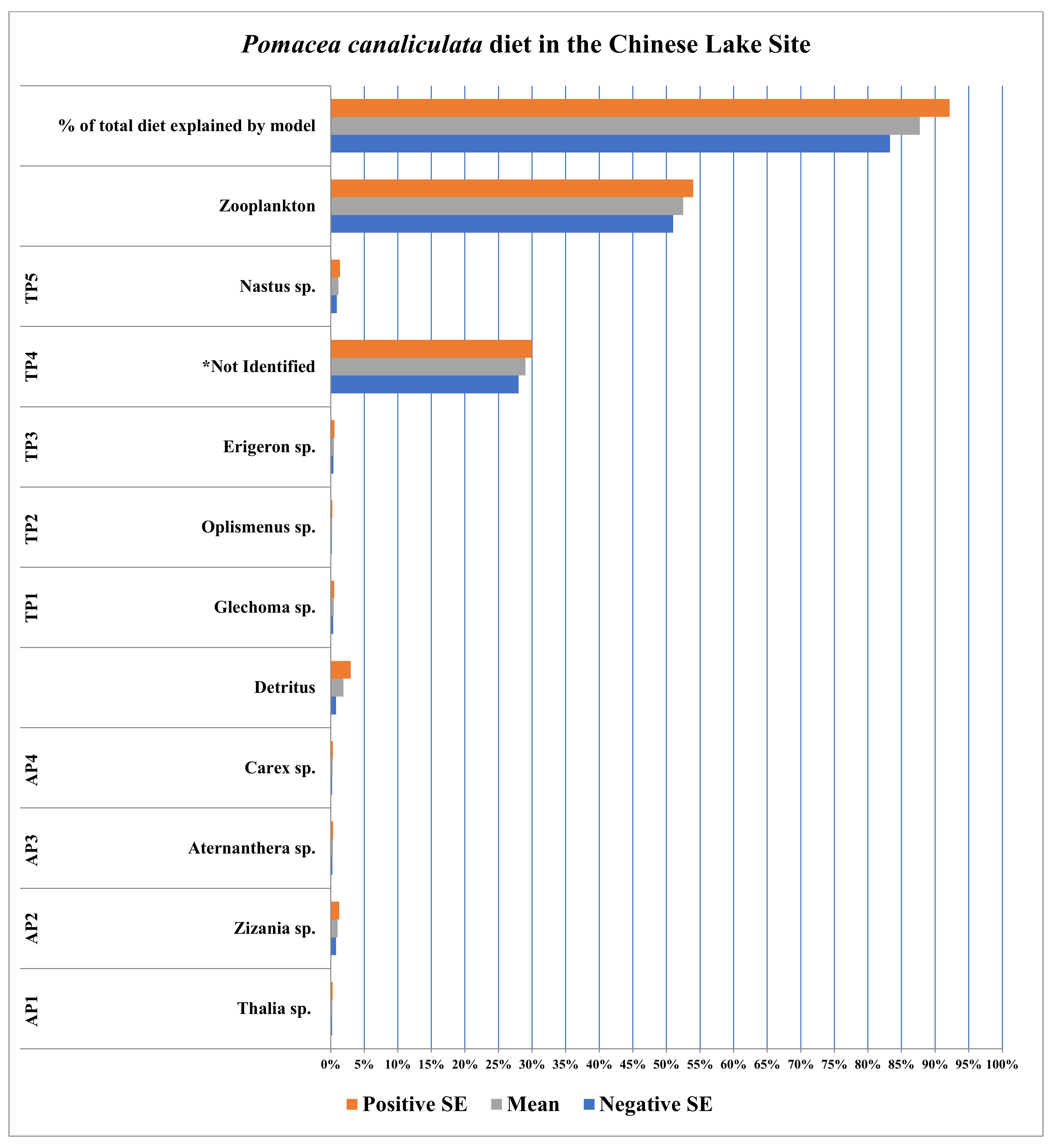
3.3. Chinese Lake Site (Hangzhou, Zhejiang, China)
| Plants/ Detritus | Sample Size (n) | Species ID | Source of Identification |
|---|---|---|---|
| AP 1 | 10 | Thalia sp. | Sequence Data |
| AP2 | 10 | Zizania sp. | Sequence Data |
| AP3 | 10 | Aternanthera sp. | Sequence Data |
| AP4 | 5 | Carex sp. | Sequence Data |
| TP1 | 5 | Glechoma sp. | Sequence Data |
| TP2 | 5 | Oplismenus sp. | Sequence Data |
| TP3 | 5 | Erigeron sp. | Sequence Data |
| TP4 | 5 | Not identified | N/A |
| TP5 | 5 | Nastus sp. | Sequence Data |
| Detritus | 5 | N/A | N/A |
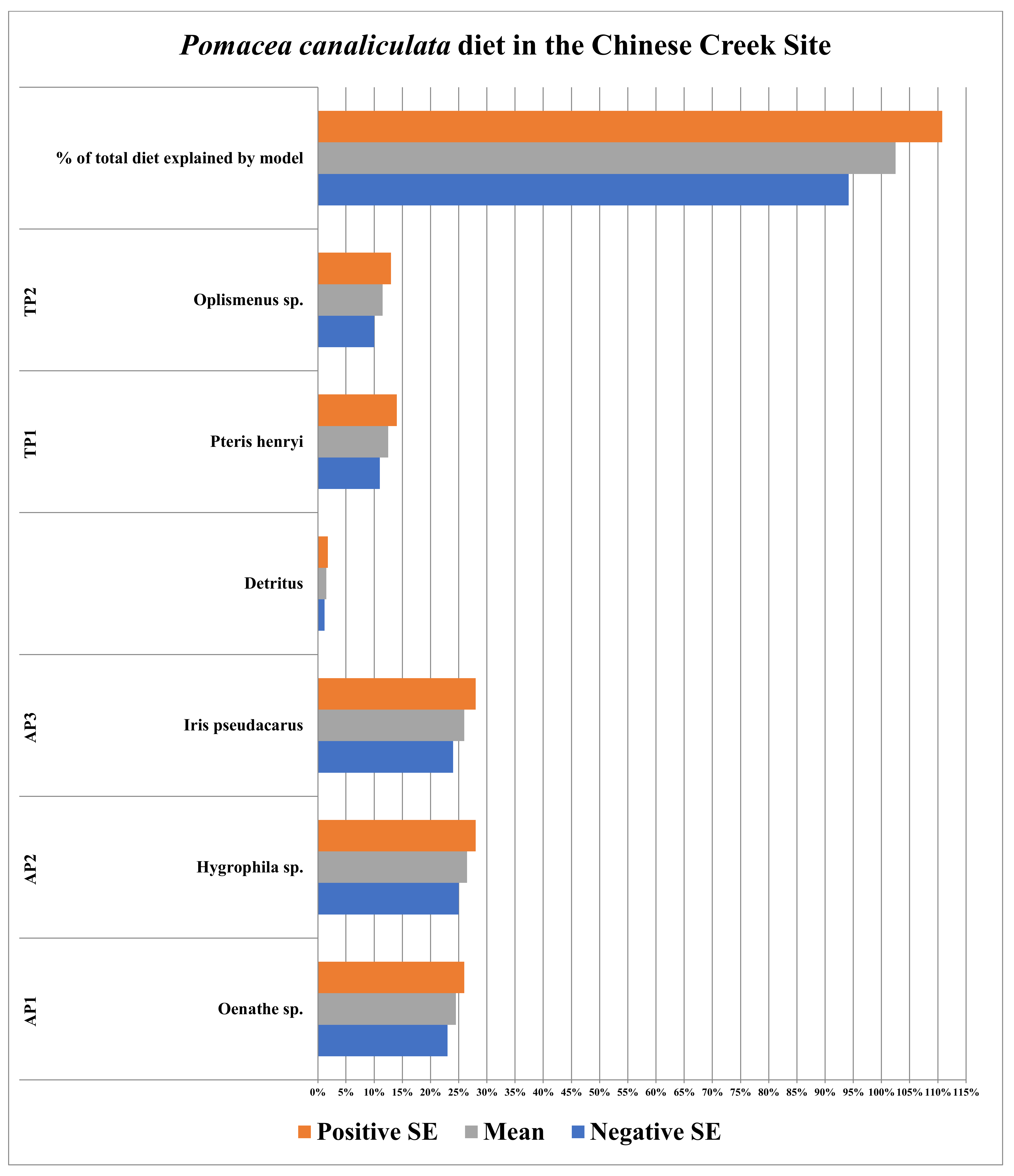
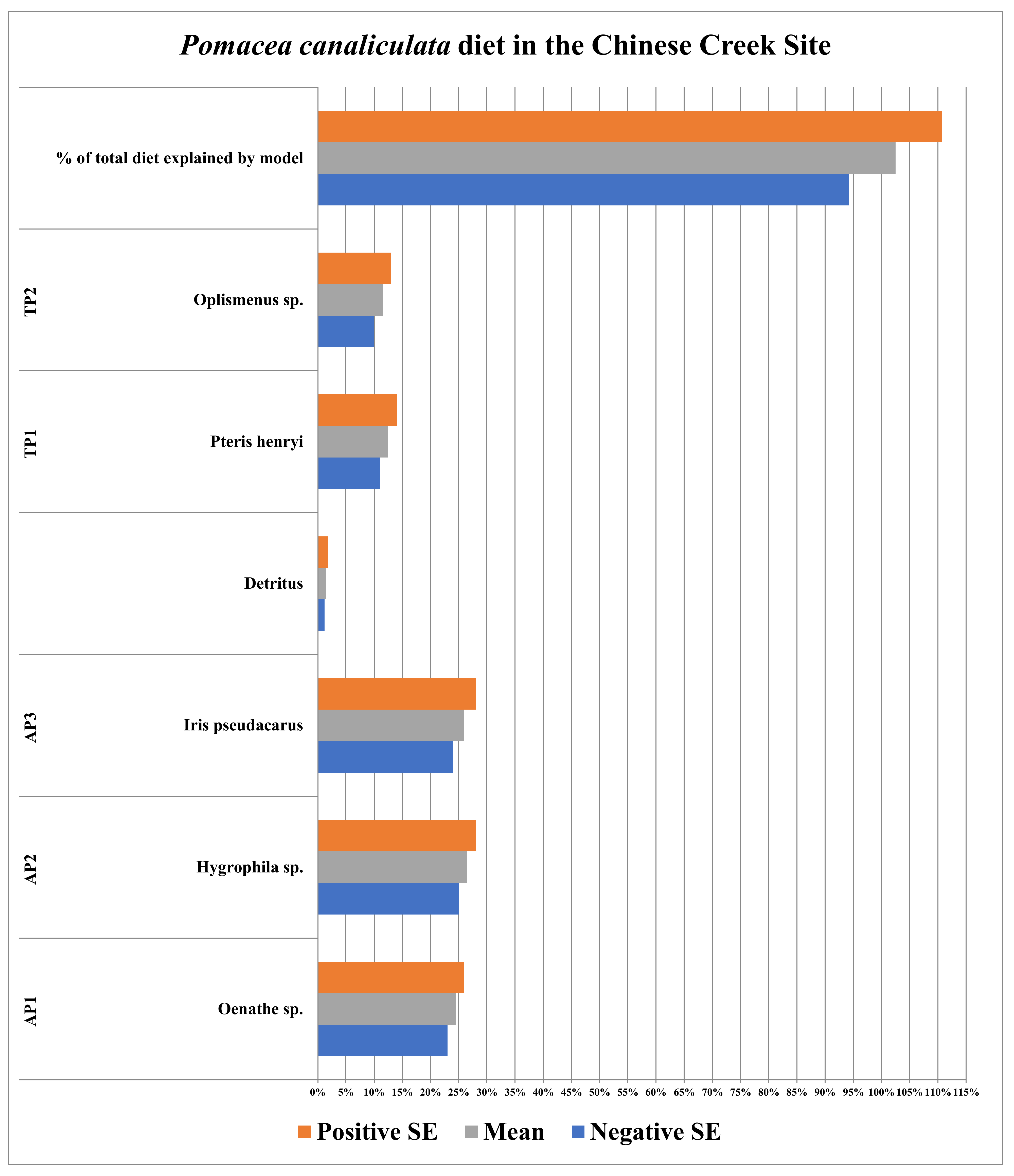
3.4. Chinese Creek Site (Hangzhou, China)
| Plants/Detritus | Sample Size (n) | Species | Source of Identification |
|---|---|---|---|
| AP1 | 10 | Oenathe sp. | Sequence Data |
| AP2 | 10 | Hygrophila sp. | Sequence Data |
| AP3 | 10 | Iris pseudacarus | Sequence Data |
| TP1 | 12 | Pteris henryi | Sequence Data |
| TP2 | 10 | Oplismenus sp. | Sequence Data |
| Detritus | 5 | N/A | N/A |
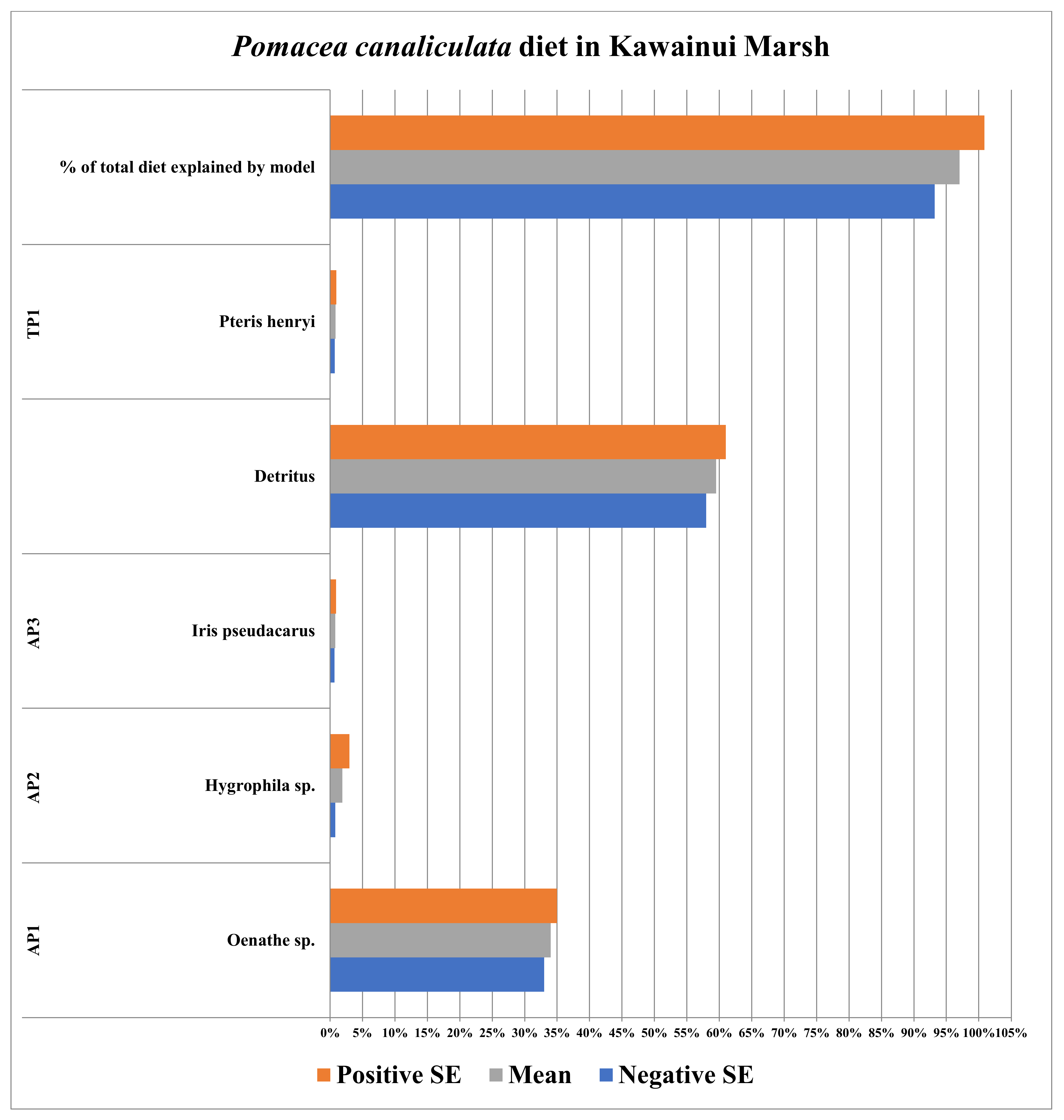
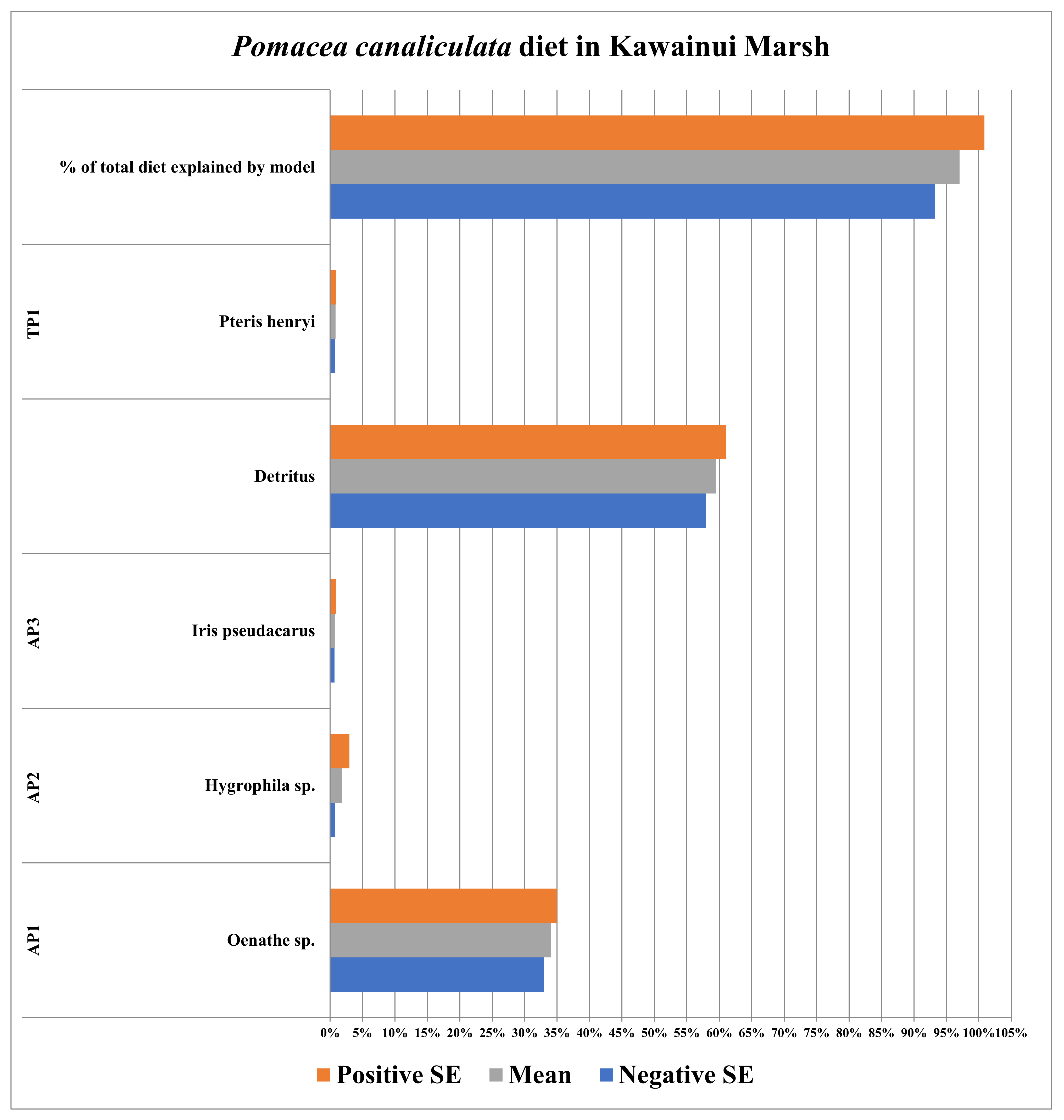
3.5. Kawainui Marsh (Oahu, HI, USA)
| Plants/Detritus | Sample Size (n) | Species | Source of Identification |
|---|---|---|---|
| AP1 | 5 | Bacopa monnieri | Field Identification |
| AP2 | 5 | Brahchiaria mutica | Field Identification |
| AP3 | 5 | Echinochloa crusgalli | Field Identification |
| TP1 | 5 | Cynodon dactylon | Field Identification |
| Detritus | 5 | N/A | N/A |
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Study Collection Site Coordinates
Appendix A.2. Respective Primers Used in This Study
- 1.
- rbcLaF (Bafeel et al., 2011; [60])
- 2.
- rbcLaR, (Bafeel et al., 2011; [60])
Appendix A.3. Specific Thermocycler Programs and Paired Primers Utilized in This Study
- (1)
- Pomacea canaliculata
| 5 min | 1 min | 1 min | 30 s | 30 s | 45 s | 10 min |
| 95 °C | 45 °C | 72 °C | 95 °C | 48 °C | 72 °C | 4 °C |
| 75 Cycles | ||||||
- (2)
- Plant Species
| 5 min | 1 min | 1 min | 30 s | 30 s | 45 s | 10 min |
| 95 °C | 45 °C | 72 °C | 95 °C | 48 °C | 72 °C | 4 °C |
| 50 Cycles | ||||||
Appendix A.4
| Sample | δ15N | δ13C |
|---|---|---|
| (‰, Air) | (‰, VPDB) | |
| Pomacea canaliculata | 5.0 | −27.0 |
| Pomacea canaliculata | 5.7 | −28.7 |
| Pomacea canaliculata | 4.3 | −25.7 |
| Pomacea canaliculata | 5.8 | −27.3 |
| Pomacea canaliculata | 5.6 | −28.1 |
| Pomacea canaliculata | 5.9 | −26.7 |
| Pomacea canaliculata | 5.0 | −26.7 |
| Pomacea canaliculata | 5.1 | −27.1 |
| Pomacea canaliculata | 5.2 | −27.8 |
| Pomacea canaliculata | 5.6 | −27.7 |
| Pomacea canaliculata | 5.7 | −27.5 |
| Pomacea canaliculata | 5.3 | −27.8 |
| Pomacea canaliculata | 5.5 | −27.7 |
| Pomacea canaliculata | 6.6 | −27.0 |
| Pomacea canaliculata | 5.4 | −27.4 |
| Pomacea canaliculata | 6.4 | −27.2 |
| Pomacea canaliculata | 5.2 | −27.8 |
| Pomacea canaliculata | 6.4 | −27.2 |
| Pomacea canaliculata | 5.3 | −27.6 |
| Pomacea canaliculata | 6.2 | −27.1 |
| Pomacea canaliculata | 5.7 | −27.0 |
| Pomacea canaliculata | 5.5 | −22.9 |
| Pomacea canaliculata | 5.0 | −27.6 |
| Pomacea canaliculata | 5.0 | −28.3 |
| Pomacea canaliculata | 4.2 | −26.8 |
| Pomacea canaliculata | 3.6 | −27.7 |
| Pomacea canaliculata | 4.7 | −26.9 |
| Pomacea canaliculata | 4.0 | −26.1 |
| Pomacea canaliculata | 5.2 | −26.8 |
| Pomacea canaliculata | 4.8 | −28.2 |
| Pomacea canaliculata | 5.7 | −27.1 |
| Pomacea canaliculata | 4.3 | −26.4 |
| Pomacea canaliculata | 4.0 | −28.5 |
| Pomacea canaliculata | 4.7 | −28.1 |
| Pomacea canaliculata | 3.3 | −28.6 |
| Pomacea canaliculata | 4.5 | −28.5 |
| Pomacea canaliculata | 4.2 | −27.5 |
| Pomacea canaliculata | 3.4 | −27.9 |
| Pomacea canaliculata | 5.1 | −28.3 |
| Pomacea canaliculata | 4.2 | −27.7 |
| Pomacea canaliculata | 5.2 | −28.2 |
| Pomacea canaliculata | 4.2 | −27.1 |
| Pomacea canaliculata | 5.4 | −27.2 |
| Pomacea canaliculata | 3.8 | −27.3 |
| Pomacea canaliculata | 5.3 | −27.8 |
| Pomacea canaliculata | 4.6 | −26.5 |
| Pomacea canaliculata | 4.2 | −21.3 |
| Pomacea canaliculata | 3.6 | −27.7 |
| Pomacea canaliculata | 2.9 | −28.0 |
| Pomacea canaliculata | 3.6 | −26.5 |
| Sample | δ15N | δ13C |
|---|---|---|
| (‰, Air) | (‰, VPDB) | |
| Pomacea canaliculata | 2.5 | −26.2 |
| Pomacea canaliculata | 2.5 | −26.0 |
| Pomacea canaliculata | 1.9 | −25.1 |
| Pomacea canaliculata | 2.9 | −26.4 |
| Pomacea canaliculata | 1.8 | −25.9 |
| Pomacea canaliculata | 1.4 | −25.3 |
| Pomacea canaliculata | 2.3 | −27.9 |
| Pomacea canaliculata | 2.8 | −27.0 |
| Pomacea canaliculata | 3.4 | −28.4 |
| Pomacea canaliculata | 0.3 | −25.9 |
| Pomacea canaliculata | 1.5 | −26.0 |
| Pomacea canaliculata | 1.0 | −24.4 |
| Pomacea canaliculata | 1.2 | −25.9 |
| Pomacea canaliculata | 1.0 | −27.0 |
| Pomacea canaliculata | 0.8 | −26.3 |
| Pomacea canaliculata | 1.5 | −28.7 |
| Pomacea canaliculata | 1.5 | −28.2 |
| Pomacea canaliculata | 1.8 | −28.5 |
| Pomacea canaliculata | 1.9 | −26.0 |
| Pomacea canaliculata | 1.1 | −22.3 |
| Pomacea canaliculata | 0.3 | −25.9 |
| Pomacea canaliculata | −0.1 | −27.1 |
| Pomacea canaliculata | 2.3 | −28.9 |
| Pomacea canaliculata | 2.1 | −28.2 |
| Pomacea canaliculata | 0.5 | −27.2 |
| Pomacea canaliculata | 1.9 | −28.5 |
| Pomacea canaliculata | 2.0 | −28.9 |
| Pomacea canaliculata | 0.4 | −27.1 |
| Pomacea canaliculata | 1.7 | −28.4 |
| Pomacea canaliculata | 1.9 | −28.9 |
| Pomacea canaliculata | 1.5 | −26.9 |
| Pomacea canaliculata | 1.5 | −28.1 |
| Pomacea canaliculata | 2.3 | −27.6 |
| Pomacea canaliculata | 1.1 | −28.3 |
| Pomacea canaliculata | 1.9 | −28.5 |
| Pomacea canaliculata | 1.9 | −28.1 |
| Pomacea canaliculata | 2.2 | −27.4 |
| Pomacea canaliculata | 1.5 | −27.4 |
| Pomacea canaliculata | 1.7 | −26.7 |
| Pomacea canaliculata | 2.5 | −28.8 |
| Pomacea canaliculata | 2.6 | −28.4 |
| Pomacea canaliculata | 3.2 | −24.5 |
| Pomacea canaliculata | 2.4 | −23.6 |
| Pomacea canaliculata | 1.3 | −26.2 |
| Pomacea canaliculata | 2.3 | −26.9 |
| Pomacea canaliculata | 2.9 | −27.6 |
| Pomacea canaliculata | 2.9 | −26.6 |
| Pomacea canaliculata | 2.1 | −26.6 |
| Pomacea canaliculata | 2.9 | −28.2 |
| Pomacea canaliculata | 2.7 | −28.5 |
| Pomacea canaliculata | 2.2 | −26.4 |
| Pomacea canaliculata | 2.6 | −27.1 |
| Pomacea canaliculata | 3.6 | −28.3 |
| Pomacea canaliculata | 2.4 | −25.9 |
| Pomacea canaliculata | 3.4 | −27.5 |
| Pomacea canaliculata | 2.6 | −26.6 |
| Pomacea canaliculata | 2.4 | −27.5 |
| Pomacea canaliculata | 2.3 | −27.6 |
| Pomacea canaliculata | 2.4 | −26.7 |
| Pomacea canaliculata | 2.9 | −26.8 |
| Pomacea canaliculata | 2.7 | −27.1 |
| Pomacea canaliculata | 3.0 | −26.1 |
| Pomacea canaliculata | 3.1 | −28.3 |
| Pomacea canaliculata | 4.2 | −28.6 |
| Sample | δ15N | δ13C |
|---|---|---|
| (‰, Air) | (‰, VPDB) | |
| Pomacea canaliculata | 3.2 | −28.9 |
| Pomacea canaliculata | 2.9 | −28.0 |
| Pomacea canaliculata | 3.6 | −27.6 |
| Pomacea canaliculata | 3.5 | −28.7 |
| Pomacea canaliculata | 4.1 | −25.0 |
| Pomacea canaliculata | 3.0 | −29.1 |
| Pomacea canaliculata | 2.8 | −29.5 |
| Pomacea canaliculata | 2.8 | −28.0 |
| Pomacea canaliculata | 4.1 | −29.3 |
| Pomacea canaliculata | 3.5 | −28.2 |
| Pomacea canaliculata | 3.8 | −28.7 |
| Pomacea canaliculata | 1.8 | −27.7 |
| Pomacea canaliculata | 3.4 | −28.4 |
| Pomacea canaliculata | 2.3 | −28.5 |
| Pomacea canaliculata | 3.4 | −28.4 |
| Pomacea canaliculata | 2.6 | −27.6 |
| Pomacea canaliculata | 3.1 | −28.3 |
| Pomacea canaliculata | 3.2 | −29.0 |
| Pomacea canaliculata | 3.5 | −26.8 |
| Pomacea canaliculata | 2.7 | −28.6 |
| Pomacea canaliculata | 2.9 | −27.5 |
| Pomacea canaliculata | 2.9 | −29.4 |
| Pomacea canaliculata | 3.3 | −26.8 |
| Pomacea canaliculata | 2.5 | −28.2 |
| Pomacea canaliculata | 2.7 | −28.1 |
| Pomacea canaliculata | 5.7 | −28.5 |
| Pomacea canaliculata | 4.7 | −28.2 |
| Pomacea canaliculata | 3.9 | −26.7 |
| Pomacea canaliculata | 4.5 | −28.6 |
| Pomacea canaliculata | 5.7 | −27.8 |
| Pomacea canaliculata | 4.7 | −27.7 |
| Pomacea canaliculata | 5.1 | −28.6 |
| Pomacea canaliculata | 5.4 | −27.7 |
| Pomacea canaliculata | 5.3 | −28.6 |
| Pomacea canaliculata | 4.4 | −27.8 |
| Pomacea canaliculata | 5.2 | −28.8 |
| Pomacea canaliculata | 4.8 | −28.2 |
| Pomacea canaliculata | 5.0 | −28.7 |
| Pomacea canaliculata | 4.5 | −27.9 |
| Pomacea canaliculata | 4.8 | −28.1 |
| Pomacea canaliculata | 4.6 | −27.6 |
| Pomacea canaliculata | 5.1 | −27.9 |
| Pomacea canaliculata | 4.5 | −27.9 |
| Pomacea canaliculata | 4.8 | −28.4 |
| Pomacea canaliculata | 5.5 | −28.1 |
| Pomacea canaliculata | 5.1 | −27.9 |
| Pomacea canaliculata | 5.2 | −28.7 |
| Pomacea canaliculata | 4.5 | −28.2 |
| Pomacea canaliculata | 4.1 | −28.4 |
| Pomacea canaliculata | 4.8 | −28.5 |
| Sample | δ15N | δ13C |
|---|---|---|
| (‰, Air) | (‰, VPDB) | |
| Pomacea canaliculata | 3.4 | −30.5 |
| Pomacea canaliculata | 5.9 | −30.9 |
| Pomacea canaliculata | 5.5 | −30.8 |
| Pomacea canaliculata | 5.2 | −30.7 |
| Pomacea canaliculata | 2.7 | −31.3 |
| Pomacea canaliculata | 6.6 | −30.3 |
| Pomacea canaliculata | 7.4 | −28.6 |
| Pomacea canaliculata | 3.7 | −31.2 |
| Pomacea canaliculata | 9.4 | −28.9 |
| Pomacea canaliculata | 4.8 | −30.2 |
| Pomacea canaliculata | 8.3 | −29.3 |
| Pomacea canaliculata | 7.1 | −29.8 |
| Pomacea canaliculata | 7.1 | −30.5 |
| Pomacea canaliculata | 3.7 | −30.9 |
| Pomacea canaliculata | 9.1 | −28.5 |
| Pomacea canaliculata | 9.8 | −28.4 |
| Pomacea canaliculata | 6.9 | −30.1 |
| Pomacea canaliculata | 6.0 | −30.1 |
| Sample | δ15N | δ13C |
|---|---|---|
| (‰, Air) | (‰, VPDB) | |
| Pomacea canaliculata | 8.5 | −18.0 |
| Pomacea canaliculata | 7.3 | −19.4 |
| Pomacea canaliculata | 7.5 | −19.8 |
| Pomacea canaliculata | 8.8 | −25.0 |
| Pomacea canaliculata | 5.5 | −24.9 |
| Pomacea canaliculata | 8.0 | −22.1 |
| Pomacea canaliculata | 9.8 | −18.2 |
| Pomacea canaliculata | 10.2 | −26.6 |
| Pomacea canaliculata | 9.5 | −23.9 |
| Pomacea canaliculata | 7.2 | −19.6 |
| Pomacea canaliculata | 8.3 | −20.1 |
| Pomacea canaliculata | 8.5 | −20.3 |
| Pomacea canaliculata | 7.5 | −16.5 |
| Pomacea canaliculata | 7.4 | −18.7 |
| Pomacea canaliculata | 7.5 | −22.9 |
| Pomacea canaliculata | 7.9 | −24.7 |
| Pomacea canaliculata | 8.5 | −25.7 |
Appendix A.5
| Plants /Plankton | Sample Size (n) | Species ID | Means of ID | δ15N | δ13C | ||
|---|---|---|---|---|---|---|---|
| Detritus /Phytoplankton | (‰, Air) | δ15N Std Dev. | (‰, VPDB) | δ13C Std Dev. | |||
| Aquatic Plant 1 | 5 | Certophyllum sp. | Sequence Data | 6.6 | 0.6 | −26.6 | 1.2 |
| Aquatic Plant 2 | 10 | Salvinia sp. | Sequence Data | 5.0 | 0.5 | −29.2 | 0.8 |
| Aquatic Plant 3 | 10 | Egeria sp. | Sequence Data | 5.9 | 1.1 | −26.7 | 0.8 |
| Aquatic Plant 4 | 10 | Monocot Perennial | Morphological Characteristics | 4.9 | 1.0 | −28.3 | 0.7 |
| Aquatic Plant 5 | 10 | Ludwigia sp. | Sequence Data | 5.2 | 0.8 | −28.2 | 0.5 |
| Aquatic Plant 6 | 10 | Eichhornia crassipes | Sequence Data | 5.5 | 0.3 | −27.9 | 0.3 |
| Aquatic Plant 7 | 10 | Echinodorus sp. | Sequence Data | 4.7 | 0.9 | −27.4 | 0.9 |
| Terrestrial Plant 1 | 10 | Poaceae sp. | Sequence Data | 5.6 | 2.4 | −12.4 | 0.9 |
| Terrestrial Plant 2 | 5 | Persicaria sp. | Sequence Data | 2.6 | 0.9 | −30.8 | 1.0 |
| Terrestrial Plant 3 | 10 | Breynia sp. | Sequence Data | 4.3 | 1.5 | −29.7 | 0.8 |
| Terrestrial Plant 4 | 10 | Hydrocotyle sp. | Sequence Data | 3.1 | 0.6 | −30.7 | 0.4 |
| Terrestrial Plant 5 | 10 | Rumex sp. | Sequence Data | 4.2 | 1.4 | −30.3 | 0.4 |
| Terrestrial Plant 6 | 5 | Not Identified | N/A | 5.5 | 1.2 | −27.4 | 0.5 |
| Zooplankton | 11 | N/A | N/A | 5.0 | 1.5 | −29.1 | 1.9 |
| Detritus | 5 | N/A | N/A | 2.83 | 0.8 | −26.5 | 0.7 |
| Phytoplankton | 3 | N/A | N/A | 4.7 | 0.9 | −28.3 | 0.4 |
| Plants/ Detritus | Sample Size (n) | Species | Means of ID | δ15N | δ13C | ||
|---|---|---|---|---|---|---|---|
| Zooplankton/ Seston | ID | (‰, Air) | δ15N Std Dev. | (‰, VPDB) | δ13C Std Dev. | ||
| Aquatic Plant 1 | 10 | Salvinia sp. | Sequence Data | 2.7 | 0.6 | −28.5 | 0.3 |
| Aquatic Plant 2 | 10 | Ludwigia sp. | Sequence Data | 3.6 | 2.3 | −27.7 | 0.6 |
| Aquatic Plant 3 | 10 | Egeria densa | Sequence Data | 3 | 0.6 | −29.4 | 1.3 |
| Aquatic Plant 4 | 10 | Potamogeton sp. | Sequence Data | −0.04 | 0.8 | −23.9 | 1.4 |
| Aquatic Plant 5 | 10 | Pistia stratiotes | Sequence Data | 4.3 | 0.5 | −29 | 0.5 |
| Aquatic Plant 6 | 10 | Potamogeton sp. | Sequence Data | 3 | 1.8 | −23.9 | 1.0 |
| Aquatic Plant 7 | 10 | Eichhornia crassipes | Sequence Data | 2.3 | 0.6 | −27.8 | 0.9 |
| Aquatic Plant 8 | 2 | Spirogyra sp. | Microscopy | 0.9 | 0.02 | −21.0 | 0.04 |
| (Alga) | |||||||
| Terrestrial Plant 1 | 5 | Poaceae sp. | Sequence Data | 7.9 | 0.36 | −13.2 | 0.1 |
| Terrestrial Plant 2 | 10 | Ludwigia sp. | Sequence Data | 6.0 | 1 | −29.5 | 0.7 |
| Detritus | 5 | N/A | N/A | 1.3 | 0.8 | −28.0 | 0.7 |
| Zooplankton | 3 | N/A | N/A | 3.4 | 0.02 | −25.4 | 1.4 |
| Seston | 3 | N/A | N/A | 1.7 | 0.2 | −30.5 | 0.1 |
| Plants/ | Sample Size (n) | Species ID | Means of ID | δ15N | δ13C | ||
|---|---|---|---|---|---|---|---|
| Detritus | (‰, Air) | δ15N Std Dev. | (‰, VPDB) | δ13C Std Dev. | |||
| Aquatic Plant 1 | 10 | Thalia sp. | Sequence Data | 5.0 | 0.8 | −30.6 | 0.9 |
| Aquatic Plant 2 | 10 | Zizania sp. | Sequence Data | 3.5 | 2.1 | −28.5 | 0.4 |
| Aquatic Plant 3 | 10 | Aternanthera sp. | Sequence Data | 5.6 | 0.2 | −28.5 | 0.7 |
| Aquatic Plant 4 | 5 | Carex sp. | Sequence Data | 3.3 | 1.0 | −31.5 | 0.7 |
| Terrestrial Plant 1 | 5 | Glechoma sp. | Sequence Data | −3.3 | 0.4 | −37.4 | 1.5 |
| Terrestrial Plant 2 | 5 | Oplismenus sp. | Sequence Data | 1.3 | 0.6 | −34.6 | 0.7 |
| Terrestrial Plant 3 | 5 | Erigeron sp. | Sequence Data | 0.3 | 0.5 | −33.3 | 0.7 |
| Terrestrial Plant 4 | 5 | Not identified | N/A | −0.5 | 0.6 | −31.1 | 0.6 |
| Terrestrial Plant 5 | 5 | Nastus sp. | Sequence Data | 1.4 | 0.3 | −30.6 | 0.2 |
| Detritus | 5 | N/A | N/A | 2.9 | 1.0 | −29.2 | 1.4 |
| Zooplankton | 4 | N/A | N/A | 3.3 | 0.4 | −26.8 | 0.2 |
| Plants/ | Sample Size (n) | Species | Means of ID | δ15N | δ13C | ||
|---|---|---|---|---|---|---|---|
| Detritus | (‰, Air) | δ15N Std Dev. | (‰, VPDB) | δ13C Std Dev. | |||
| Aquatic Plant 1 | 10 | Oenathe sp. | Sequence Data | 5.3 | 1.6 | −30.8 | 1.3 |
| Aquatic Plant 2 | 10 | Hygrophila sp. | Sequence Data | 4.8 | 1.8 | −31.5 | 1.0 |
| Aquatic Plant 3 | 10 | Iris pseudacarus | Sequence Data | 6.7 | 1.4 | −31.1 | 0.9 |
| Terrestrial Plant 1 | 12 | Pteris henryi | Sequence Data | 1.3 | 2.0 | −29.0 | 1.3 |
| Terrestrial Plant 2 | 10 | Oplismenus sp. | Sequence Data | −4.2 | 4.1 | −31.3 | 0.7 |
| Detritus | 5 | N/A | N/A | −0.1 | 3.3 | −21.0 | 2.4 |
| Plants/ | Sample Size (n) | Species ID | Means of ID | δ15N | δ13C | ||
|---|---|---|---|---|---|---|---|
| Detritus | (‰, Air) | δ15N Std Dev. | (‰, VPDB) | δ13C Std Dev. | |||
| Aquatic Plant 1 | 5 | Bacopa monnieri | Field ID Manual | 9.9 | 1.4 | −30.1 | 0.3 |
| Aquatic Plant 2 | 5 | Brahchiaria mutica | Field ID Manual | 7.5 | 0.3 | −12.4 | 0.3 |
| Aquatic Plant 3 | 5 | Echinochloa crusgalli | Field ID Manual | 8.9 | 1.0 | −12.3 | 0.3 |
| Terrestrial Plant 1 | 5 | Cynodon dactylon | Field ID Manual | 9.0 | 0.6 | −12.6 | 0.5 |
| Detritus | 5 | N/A | N/A | 3.4 | 1.4 | −16.2 | 1.6 |
References
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walther, G.R. Community and ecosystem responses to recent climate change. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2019–2024. [Google Scholar] [CrossRef] [PubMed]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Vilà, M.; Basnou, C.; Pyšek, P.; Josefsson, M.; Genovesi, P.; Gollasch, S.; Nentwig, W.; Olenin, S.; Roques, A.; Roy, D.; et al. How well do we understand the impacts of alien species on ecosystem services? A pan-European, cross-taxa assessment. Front. Ecol. Environ. 2010, 8, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Çinar, M.E.; Oztürk, B.; Grabowski, M.; Golani, D.; Cardoso, A.C. Impacts of invasive alien marine species on ecosystem services and biodiversity: A pan-European review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Baillie, J.E.; Hilton-Taylor, C.; Stuart, S.N. 2004 IUCN Red List of Threatened Species: A Global Species Assessment. International Union for Conservation of Nature (IUCN) 2004; IUCN: Glad, Switzerland; Cambridge, UK, 2004. [Google Scholar]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef]
- Hooper, D.U.; Adair, E.C.; Cardinale, B.J.; Byrnes, J.E.; Hungate, B.A.; Matulich, K.L.; Gonzalez, A.; Duffy, J.E.; Gamfeldt, L.; O’Connor, M.I. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 2012, 486, 105–109. [Google Scholar] [CrossRef]
- Boulton, A.J.; Ekebom, J.; Gislason, G.M. Integrating ecosystem services into conservation strategies for freshwater and marine habitats: A review. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 26, 963–985. [Google Scholar] [CrossRef] [Green Version]
- Ehrenfeld, J.G. Ecosystem consequences of biological invasions. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 59–80. [Google Scholar] [CrossRef] [Green Version]
- Strayer, D.L. Alien species in fresh waters: Ecological effects, interactions with other stressors, and prospects for the future. Freshw. Biol. 2010, 55, 152–174. [Google Scholar] [CrossRef]
- Vander Zanden, M.J.; Olden, J.D. A management framework for preventing the secondary spread of aquatic invasive species. Can. J. Fish. Aquat. Sci. 2008, 65, 1512–1522. [Google Scholar] [CrossRef]
- Keller, R.P.; Lodge, D.M.; Lewis, M.A.; Shogren, J.F. Bioeconomics of Invasive Species: Integrating Ecology, Economics, Policy, and Management, 1st ed.; Oxford University Press: New York, NY, USA, 2009; pp. 151–179. [Google Scholar]
- Clavero, M.; García-Berthou, E. Invasive species are a leading cause of animal extinctions. Trends Ecol. Evol. 2005, 20, 110. [Google Scholar] [CrossRef] [Green Version]
- Gurevitch, J.; Padilla, D.K. Are invasive species a major cause of extinctions? Trends Ecol. Evol. 2004, 19, 470–474. [Google Scholar] [CrossRef]
- Charles, H.; Dukes, J.S. Impacts of Invasive Species on Ecosystem Services. In Biological Invasions; Nentwig, W., Caldwell, M.M., Heldmaier, G., Jackson, R.B., Lange, O.L., Mooney, H.A., Schulze, E.-D., Sommer, U., Eds.; Ecological Studies Volume 193; Springer: Berlin/Heidelberg, Germany, 2008; pp. 217–237. [Google Scholar]
- Pejchar, L.; Mooney, H.A. Invasive species, ecosystem services and human well-being. Trends Ecol. Evol. 2009, 24, 497–504. [Google Scholar] [CrossRef]
- Didham, R.K.; Tylianakis, J.M.; Hutchison, M.A.; Ewers, R.M.; Gemmell, N.J. Are invasive species the drivers of ecological change? Trends Ecol. Evol. 2005, 20, 470–474. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species. In the Global Invasive Species Database; The Invasive Species Specialist Group (ISSG) a Specialist Group of the Species Survival Commission (SSC) of the World Conservation Union (IUCN): Auckland, New Zealand, 2010; Volume 1, pp. 1–12. [Google Scholar]
- Cowie, R.H. Apple snails (Ampullariidae) as agricultural pests: Their biology, impacts and management. In Molluscs as Crop Pests, 1st ed.; Barker, G.M., Ed.; CABI Publishing: New York, NY, USA, 2002; Volume 1, pp. 145–192. [Google Scholar]
- Levin, P. Statewide Strategic Control Plan for Apple Snail (Pomacea canaliculata) in Hawai‘i; The Hawaii Land Restoration Institute: Wailuku, HI, USA, 2006; pp. 1–20. [Google Scholar]
- Wong, P.K.; Liang, Y.A.N.; Liu, N.Y.; Qiu, J.W. Palatability of macrophytes to the invasive freshwater snail Pomacea canaliculata: Differential effects of multiple plant traits. Freshw. Biol. 2010, 55, 2023–2031. [Google Scholar] [CrossRef]
- Gilal, A.A.; Muhamad, R.; Omar, D.; Aziz, N.A.A.; Gnanasegaram, M. Foes can be Friends: Laboratory Trials on Invasive Apple Snails, Pomacea spp. Preference to Invasive Weed, Limnocharis flava (L.) Buchenau Compared to Rice, Oryza sativa L. Pak. J. Zool. 2016, 48, 673–679. [Google Scholar]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef]
- Carlsson, N.O.; Brönmark, C.; Hansson, L.A. Invading herbivory: The golden apple snail alters ecosystem functioning in Asian wetlands. Ecology 2004, 85, 1575–1580. [Google Scholar] [CrossRef] [Green Version]
- Fickbohm, S.S.; Zhu, W.X. Exotic purple loosestrife invasion of native cattail freshwater wetlands: Effects on organic matter distribution and soil nitrogen cycling. Appl. Soil Ecol. 2006, 32, 123–131. [Google Scholar] [CrossRef]
- Lv, S.; Zhang, Y.; Liu, H.X.; Hu, L.; Yang, K.; Steinmann, P.; Chen, Z.; Wang, L.Y.; Utzinger, J.; Zhou, X.N. Invasive snails and an emerging infectious disease: Results from the first national survey on Angiostrongylus cantonensis in China. PLoS Negl. Trop. Dis. 2009, 3, e368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, R.A. A Handbook of Global Freshwater Invasive Species, 1st ed.; Earthscan: New York, NY, USA, 2012; pp. 1–376. [Google Scholar]
- Wiryareja, S.; Tjoe-Awie, J.R. Golden apple snail: Its occurrence and importance in Suriname’s rice ecosystem. In Global Advances in Ecology and Management of Golden Apple Snails; PhilRice: Science City of Munoz, Philippines, 2006; pp. 337–342. [Google Scholar]
- Minson, D.J.; Ludlow, M.M.; Troughton, J.H. Differences in natural carbon isotope ratios of milk and hair from cattle grazing tropical and temperate pastures. Nature 1975, 256, 602. [Google Scholar] [CrossRef] [PubMed]
- Tieszen, L.L.; Boutton, T.W.; Tesdahl, K.G.; Slade, N.A. Fractionation and turnover of stable carbon isotopes in animal tissues: Implications for δ 13 C analysis of diet. Oecologia 1983, 57, 32–37. [Google Scholar] [CrossRef]
- Samelius, G.; Alisauskas, R.T.; Hobson, K.A.; Larivière, S. Prolonging the arctic pulse: Long-term exploitation of cached eggs by arctic foxes when lemmings are scarce. J. Anim. Ecol. 2007, 76, 873–880. [Google Scholar] [CrossRef]
- López Van Oosterom, M.V.; Ocon, C.S.; Ferreira, A.C.; Rodrigues Capítulo, A. The diet of Pomacea canaliculata (Gastropoda: Ampullariidae) in its native habitat based on gut content and stable isotopes analysis. Intropica 2016, 11, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Iglesias, C.; Meerhoff, M.; Johansson, L.S.; González-Bergonzoni, I.; Mazzeo, N.; Pacheco, J.P.; Teixeira-de Mello, F.; Goyenola, G.; Lauridsen, T.L.; Søndergaard, M.; et al. Stable isotope analysis confirms substantial differences between subtropical and temperate shallow Lake food webs. Hydrobiologia 2017, 784, 111–123. [Google Scholar] [CrossRef]
- Schimmelmann, A.; Albertino, A.; Sauer, P.E.; Qi, H.; Molinie, R.; Mesnard, F. Nicotine, acetanilide and urea multi-level 2H-, 13C-and 15N-abundance reference materials for continuous-flow isotope ratio mass spectrometry. Rapid Commun. Mass Spectrom. An. Int. J. Devoted Rapid Dissem. Up Min. Res. Mass Spectrom. 2009, 23, 3513–3521. [Google Scholar] [CrossRef]
- Parnell, A.C.; Inger, R.; Bearhop, S.; Jackson, A.L. Source partitioning using stable isotopes: Coping with too much variation. PLoS ONE 2010, 5, e9672. [Google Scholar] [CrossRef]
- Phillips, D.L.; Inger, R.; Bearhop, S.; Jackson, A.L.; Moore, J.W.; Parnell, A.C.; Semmens, B.X.; Ward, E.J. Best practices for use of stable isotope mixing models in food-web studies. Can. J. Zool. 2014, 92, 823–835. [Google Scholar] [CrossRef] [Green Version]
- Raw, J.L.; Perissinotto, R.; Miranda, N.A.F.; Peer, N. Feeding dynamics of Melanoides tuberculata (Müller, 1774). J. Molluscan Stud. 2016, 82, 328–335. [Google Scholar] [CrossRef] [Green Version]
- Vanderklift, M.A.; Ponsard, S. Sources of variation in consumer-diet δ 15 N enrichment: A meta-analysis. Oecologia 2003, 136, 169–182. [Google Scholar] [CrossRef]
- McCutchan, J.H., Jr.; Lewis, W.M., Jr.; Kendall, C.; McGrath, C.C. Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos 2003, 102, 378–390. [Google Scholar] [CrossRef]
- Erickson, T.A.; Puttock, C.F. Hawai’i Wetland Field Guide: An Ecological and Identification Guide to Wetlands and Wetland Plants of the Hawaiian Islands, 1st ed.; Bess Press: Honolulu, HI, USA, 2006; pp. 249–263. [Google Scholar]
- García-Robledo, C.; Erickson, D.L.; Staines, C.L.; Erwin, T.L.; Kress, W.J. Tropical plant–herbivore networks: Reconstructing species interactions using DNA barcodes. PLoS ONE 2013, 8, e52967. [Google Scholar] [CrossRef]
- Bourne, G.R. Differential snail-size predation by snail kites and limpkins. Oikos 1993, 68, 217–223. [Google Scholar] [CrossRef]
- Tanaka, M.O.; Souza, A.L.; Módena, É.S. Habitat structure effects on size selection of snail kites (Rostrhamus sociabilis) and limpkins (Aramus guarauna) when feeding on apple snails (Pomacea spp.). Acta Oecologica 2006, 30, 88–96. [Google Scholar] [CrossRef]
- Jackson, M.C.; Wasserman, R.J.; Grey, J.; Ricciardi, A.; Dick, J.T.; Alexander, M.E. Novel and disrupted trophic links following invasion in freshwater ecosystems. In Advances in Ecological Research; Academic Press: Cambridge, MA, USA, 2017; Volume 57, pp. 55–97. [Google Scholar]
- Pyke, G.H. Optimal foraging theory: A critical review. Annu. Rev. Ecol. Syst. 1984, 15, 523–575. [Google Scholar] [CrossRef]
- MacNeil, C.; Dick, J.T.; Johnson, M.P.; Hatcher, M.J.; Dunn, A.M. A species invasion mediated through habitat structure, intraguild predation, and parasitism. Limnol. Oceanogr. 2004, 49, 1848–1856. [Google Scholar] [CrossRef]
- Moyle, P.B.; Light, T. Biological invasions of fresh water: Empirical rules and assembly theory. Biol. Conserv. 1996, 78, 149–161. [Google Scholar] [CrossRef]
- Cucherousset, J.; Olden, J.D. Ecological impacts of nonnative freshwater fishes. Fisheries 2011, 36, 215–230. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Hill, A.M.; Lodge, D.M. Replacement of resident crayfishes by an exotic crayfish: The roles of competition and predation. Ecol. Appl. 1999, 9, 678–690. [Google Scholar] [CrossRef]
- James, J.; Thomas, J.R.; Ellis, A.; Young, K.A.; England, J.; Cable, J. Over-invasion in a freshwater ecosystem: Newly introduced virile crayfish (Orconectes virilis) outcompete established invasive signal crayfish (Pacifastacus leniusculus). Mar. Freshw. Behav. Physiol. 2016, 49, 9–18. [Google Scholar] [CrossRef]
- Covich, A.P. Dispersal-limited biodiversity of tropical insular streams. Pol. J. Ecol. 2006, 54, 523–547. [Google Scholar]
- Dick, J.T. Role of behaviour in biological invasions and species distributions; lessons from interactions between the invasive Gammarus pulex and the native G. duebeni (Crustacea: Amphipoda). Contrib. Zool. 2008, 77, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Westhoff, J.T.; Rabeni, C.F. Resource selection and space use of a native and an invasive crayfish: Evidence for competitive exclusion? Freshw. Sci. 2013, 32, 1383–1397. [Google Scholar] [CrossRef]
- Jackson, M.C.; Britton, J.R. Divergence in the trophic niche of sympatric freshwater invaders. Biol. Invasions 2014, 16, 1095–1103. [Google Scholar] [CrossRef]
- Brandon-Mong, G.J.; Gan, H.M.; Sing, K.W.; Lee, P.S.; Lim, P.E.; Wilson, J.J. DNA metabarcoding of insects and allies: An evaluation of primers and pipelines. Bull. Entomol. Res. 2015, 105, 717–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colgan, D.J.; Hutchings, P.A.; Brown, S. Phylogenetic relationships within the Terebellomorpha. J. Mar. Biol. Assoc. UK 2001, 81, 765–773. [Google Scholar] [CrossRef]
- Bafeel, S.O.; Arif, I.A.; Bakir, M.A.; Khan, H.A.; Al Farhan, A.H.; Al Homaidan, A.A.; Ahamed, A.; Thomas, J. Comparative evaluation of PCR success with universal primers of maturase K (matK) and ribulose-1, 5-bisphosphate carboxylase oxygenase large subunit (rbcL) for barcoding of some arid plants. Plant. Omics 2011, 4, 195–198. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scriber, K.E.; France, C.A.M.; Jackson, F.L.C. Invasive Apple Snail Diets in Native vs. Non-Native Habitats Defined by SIAR (Stable Isotope Analysis in R). Sustainability 2022, 14, 7108. https://doi.org/10.3390/su14127108
Scriber KE, France CAM, Jackson FLC. Invasive Apple Snail Diets in Native vs. Non-Native Habitats Defined by SIAR (Stable Isotope Analysis in R). Sustainability. 2022; 14(12):7108. https://doi.org/10.3390/su14127108
Chicago/Turabian StyleScriber, Kevin E., Christine A. M. France, and Fatimah L. C. Jackson. 2022. "Invasive Apple Snail Diets in Native vs. Non-Native Habitats Defined by SIAR (Stable Isotope Analysis in R)" Sustainability 14, no. 12: 7108. https://doi.org/10.3390/su14127108
APA StyleScriber, K. E., France, C. A. M., & Jackson, F. L. C. (2022). Invasive Apple Snail Diets in Native vs. Non-Native Habitats Defined by SIAR (Stable Isotope Analysis in R). Sustainability, 14(12), 7108. https://doi.org/10.3390/su14127108