
What Is the Impact of Microplastics and Lipid Regulators on Marine Meiofauna? Case Study of Polyvinyl Chloride, Atorvastatin, and Simvastatin

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Material and Methods
2.1. Collecting Site and Sediment Manipulation
2.2. Sediment Contamination and Experiment Set-Up
2.3. Meiofauna Analyses
- The shape of tails, according to Thistle et al. [45] as follows: conical (co), short/round (s/r), clavate/conico-cylindrical (cla), and elongated/filiform (e/f).
- The shape of amphideal fovea, according to Semprucci et al. [29], as follows: spiral (Sp), circular (Cr), indistinct (Id), and pocket (Pk).
- The life history (c–p scores), according to Bongers et al. [46] ranked from 1 to 5.
- The length of adults, according to Schratzberger et al. [47] divided in the following size-classes: 1–2 mm, 2–4 mm, and ˃4 mm. The body-size was measured using a Nikon DS-Fi2 camera coupled with a Nikon microscope (Image Software NIS Elements Analysis Version 4.0 Nikon 4.00.07ebuild 787e64 bit).
2.4. StatisticalAanalyses
3. Results
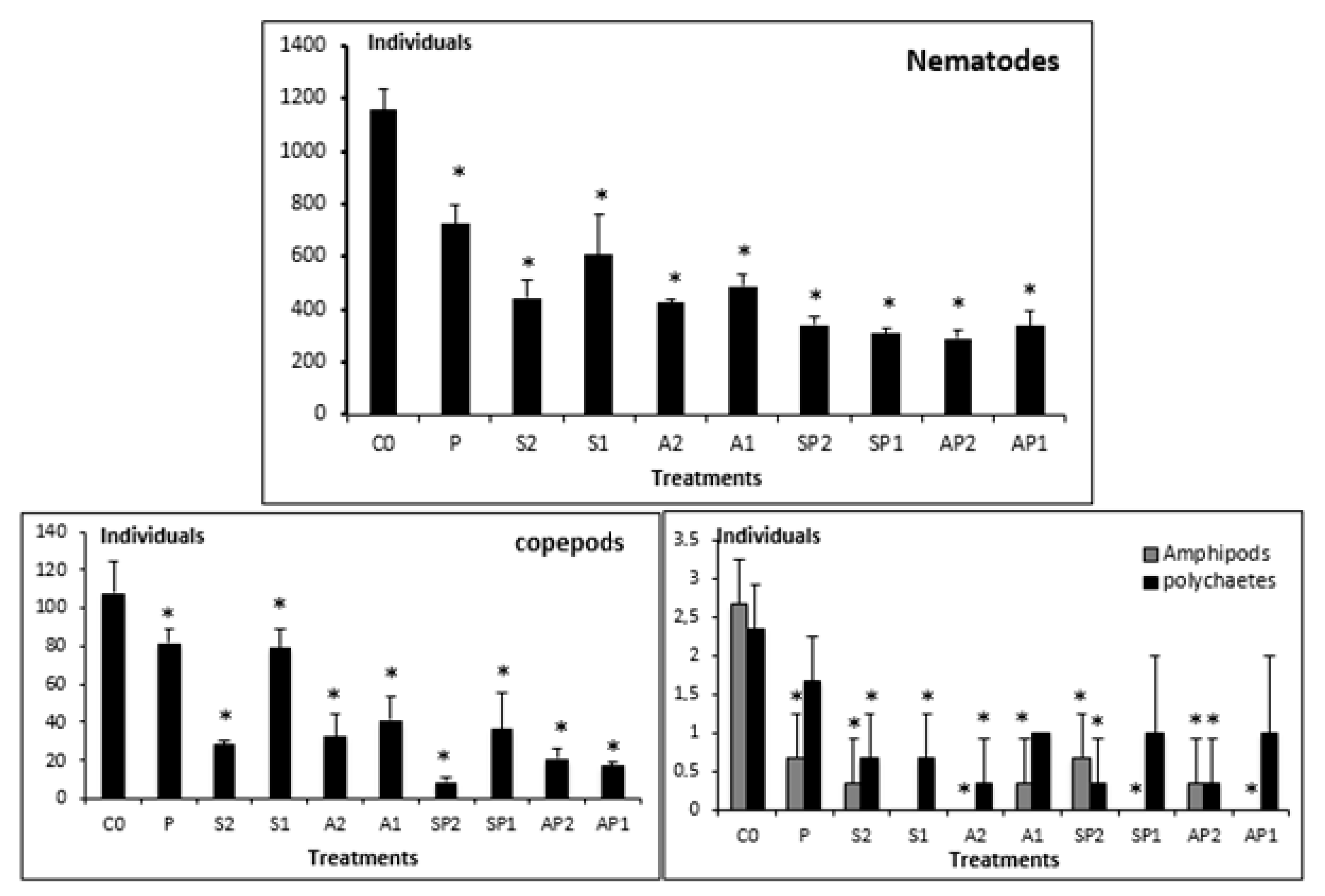
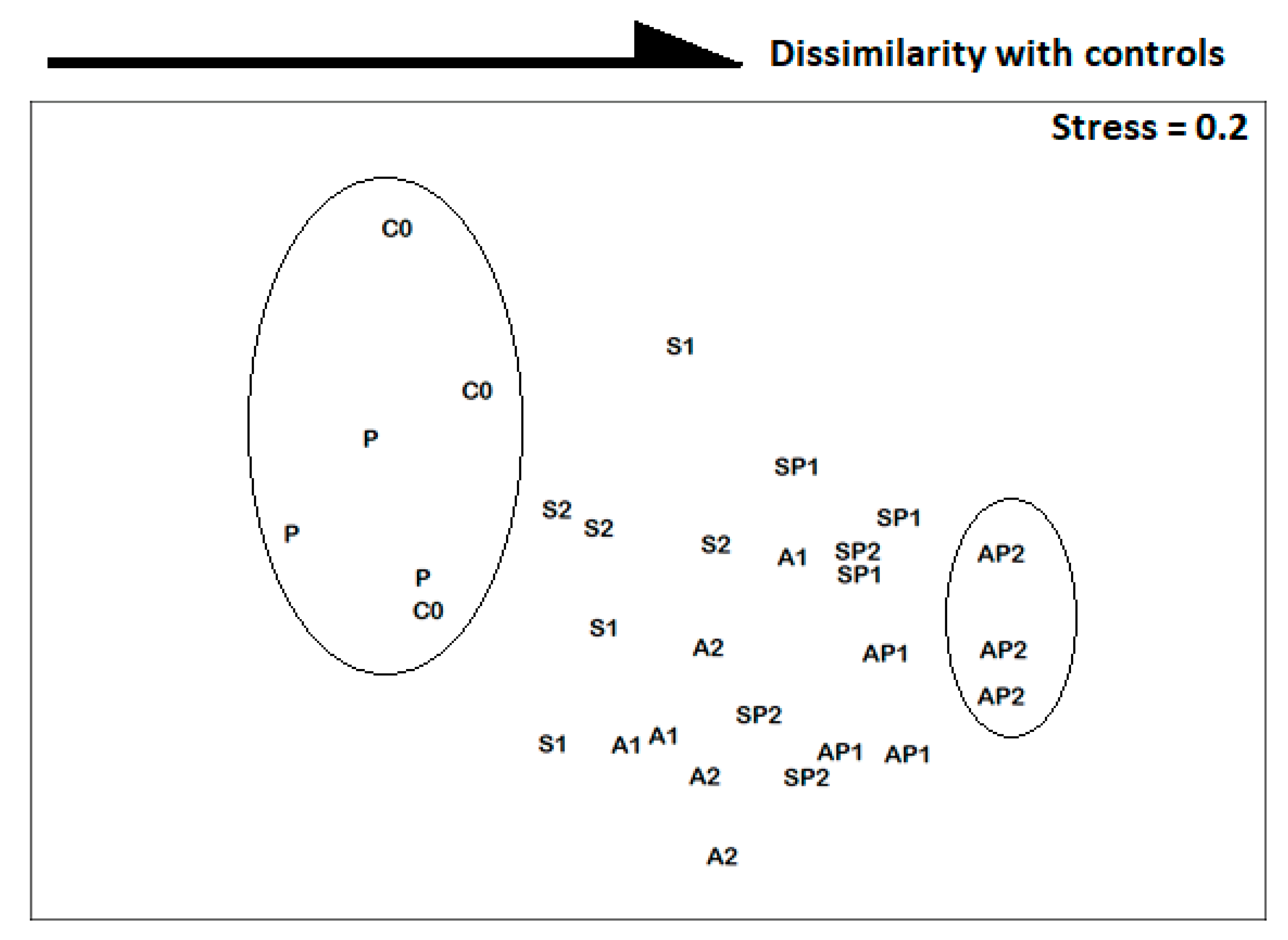
3.1. Meiofaunistic Abundances
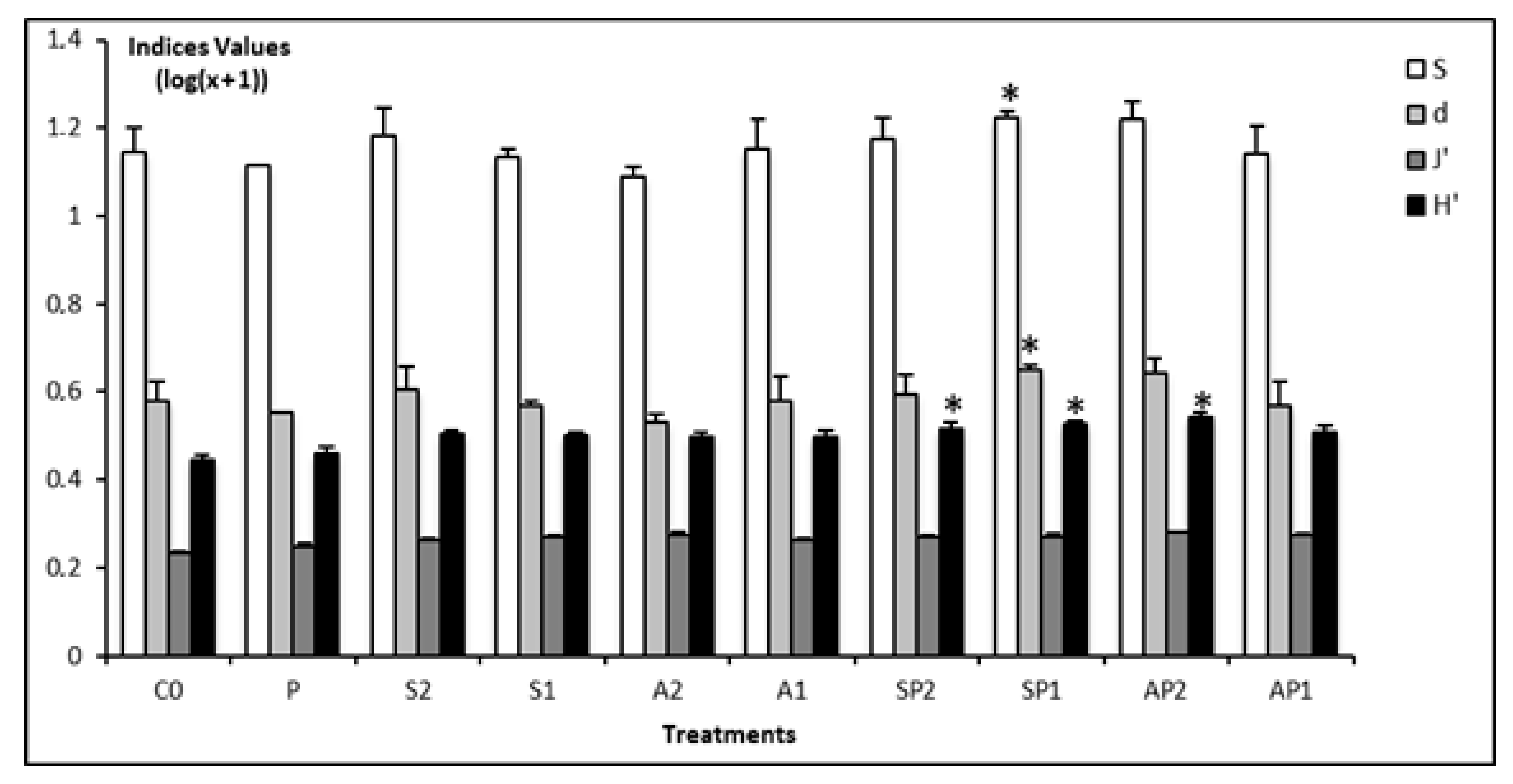
3.2. Taxonomic Diversity of Nematodes
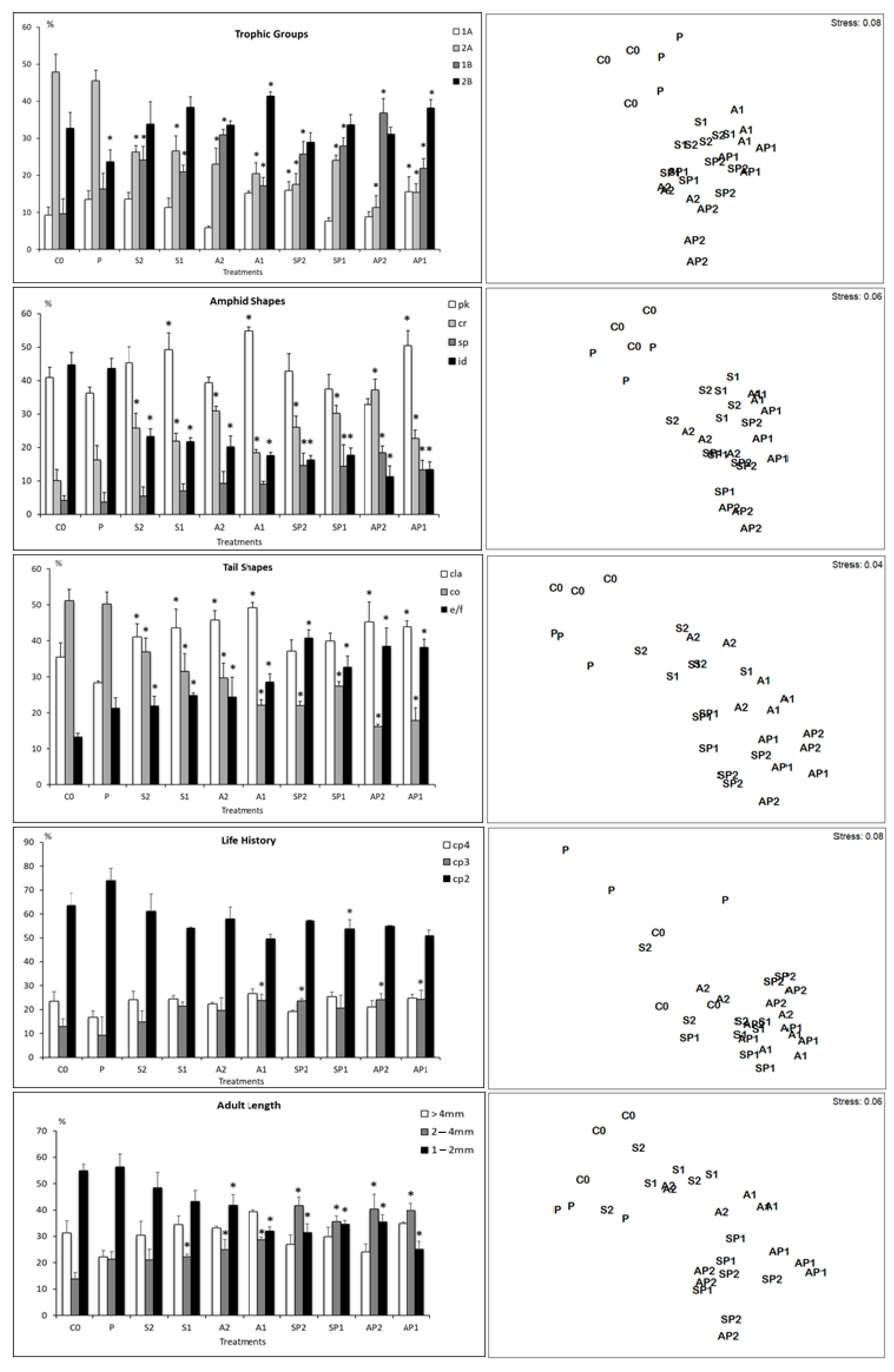
3.3. Functional Diversity of Nematodes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Selamoglu, M. Importance of the cold chain logistics in the marketing process of aquatic products: An update study. J. Surv. Fish. Sci. 2021, 8, 25–29. [Google Scholar]
- Cunningham, V.L.; Buzby, M.; Hutchinson, T.; Mastrocco, F.; Parke, N.; Roden, N. Effects of human pharmaceuticals on aquatic life: Next steps. Environ. Sci. Technol. 2006, 40, 3457–3462. [Google Scholar] [CrossRef] [Green Version]
- Gore, M.; Sadosky, A.; Leslie, D.; Sheehan, A.H. Selecting an appropriate medication for treating neuropathic pain in patients with diabetes: A study using the UK and Germany Mediplus databases. Pain Pr. 2008, 8, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Grimmsmann, T.; Schwabe, U.; Himmel, W. The influence of hospitalisation on drug prescription in primary care–a large-scale follow-up study. Eur. J. Clin. Pharmacol. 2007, 63, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Ghoshdastidar, A.J.; Fox, S.; Tong, A.Z. The presence of the top prescribed pharmaceuticals in treated sewage effluents and receiving waters in Southwest Nova Scotia, Canada. Environ. Sci. Pollut. Res. 2015, 22, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, A.; Pineda, M.; Venkata, K. Comprehension of top 200 prescribed drugs in the US as a resource for pharmacy teaching, training and practice. Pharmacy 2018, 6, 43. [Google Scholar] [CrossRef] [Green Version]
- Mc Namara, K.; Alzubaidi, H.; Jackson, J.K. Cardiovascular disease as a leading cause of death: How are pharmacists getting involved? Integr. Pharm. Res. Pract. 2019, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wilson, S.; Simari, R.D.; Best, P.J.; Peterson, T.E.; Lerman, L.O.; Avirman, M.; Nath, K.A.; Holmes, D.R., Jr.; Lerman, A. Simvastatin preserves coronary endothelial function in hypercholesterolemia in the absence of lipid lowering. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Dussault, E.B.; Balakrishnan, V.K.; Sverko, E.D.; Solomon, K.R.; Sibley, P.K. Toxicity of human pharmaceuticals and personal care products to benthic invertebrates. Environ. Toxicol. Chem. 2008, 27, 425–432. [Google Scholar] [CrossRef]
- Key, P.B.; Hoguet, J.; Reed, L.A.; Chung, K.W.; Fulton, M.H. Effects of the Statin Antihyperlipidemic Agent Simvastatin on Grass Shrimp, Palaemonetes pugio. Environ. Toxicol. 2008, 23, 153–160. [Google Scholar] [CrossRef]
- Chen, X.Q.; Cho, S.J.; Li, Y.; Venkatesh, S. Prediction of Aqueous Solubility of Organic Compounds Using a Quantitative Structure–Property Relationship. J. Pharm. Sci. 2002, 91, 1838–1852. [Google Scholar] [CrossRef] [PubMed]
- Estimation Program Interface (EPI) Suite. Available online: https://www2.epa.gov/tsca-screening-tools (accessed on 7 November 2020).
- Strungaru, S.A.; Jijie, R.; Nicoara, M.; Plavan, G.; Faggio, C. Micro-(nano) plastics in freshwater ecosystems: Abundance, toxicological impact and quantification methodology. TrAC Trends Anal. Chem. 2019, 110, 116–128. [Google Scholar] [CrossRef]
- Tagg, A.S.; Sapp, M.; Harrison, J.P.; Ojeda, J.J. Identification and quantification of microplastics in wastewater using focal plane array-based reflectance micro-FT-IR imaging. Anal. Chem. 2015, 87, 6032–6040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Souza Machado, A.A.; Kloas, W.; Zarfl, C.; Hempel, S.; Rillig, M.C. Microplastics as an emerging threat to terrestrial ecosystems. Glob. Chang. Biol. 2018, 24, 1405–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eerkes-Medrano, D.; Thompson, R.C.; Aldridge, D.C. Microplastics in freshwater systems: A review of the emerging threats, identification of knowledge gaps and prioritisation of research needs. Water. Res. 2015, 75, 63–82. [Google Scholar] [CrossRef]
- Aytan, U.; Valente, A.; Senturk, Y.; Usta, R.; Esensoy Sahin, F.B.; Mazlum, R.E.; Agirbas, E. First evaluation of neustonic microplastics in Black Sea waters. Mar. Environ. Res. 2016, 119, 22–30. [Google Scholar] [CrossRef]
- Andrady, A.L. The plastic in microplastics: A review. Mar. Pollut. Bull. 2017, 119, 12–22. [Google Scholar] [CrossRef]
- Öztekin, A.; Bat, L. Microlitter Pollution in Sea Water: A Preliminary Study from Sinop Sarikum Coast of the Southern Black Sea. Turk. J. Fish. Aquat. Sci. 2017, 17, 1431–1440. [Google Scholar] [CrossRef]
- Silva, A.B.; Bastos, A.S.; Justino, C.I.L.; da-Costa, J.P.; Duarte, A.C.; Rocha-Santos, T.A.P. Microplastics in the environment: Challenges in analytical chemistry-a review. Anal. Chim. Acta. 2018, 1017, 1–19. [Google Scholar] [CrossRef]
- Heskett, M.; Takada, H.; Yamashita, R.; Yuyama, M.; Ito, M.; Geok, Y.B.; Ogata, Y.; Kwan, C.; Heckhausen, A.; Taylor, H.; et al. Measurement of persistent organic pollutants (POPs) in plastic resin pellets from remote islands: Toward establishment of background concentrations for international pellet watch. Mar. Pollut. Bull. 2012, 64, 445–448. [Google Scholar] [CrossRef] [PubMed]
- Jasna, M.L.; Jelena, L.; Pero, T.; Varezić, D.B.; Šiljić, J.; Pribudić, J. Levels of trace metals on microplastic particles in beach sediments of the island of Vis, Adriatic Sea, Croatia. Mar. Pollut. Bull. 2018, 137, 231–236. [Google Scholar]
- Hale, R.C.; Seeley, M.E.; La Guardia, M.J.; Mai, L.; Zeng, E.Y. A global perspective on microplastics. J. Geophys. Res. Ocean. 2020, 125. [Google Scholar] [CrossRef]
- Yu, Q.; Hu, X.; Yang, B.; Zhang, G.; Wang, J.; Ling, W. Distribution, abundance and risks of microplastics in the environment. Chemosphere 2020, 249, 126059. [Google Scholar] [CrossRef] [PubMed]
- Warwick, R.M.; Price, R. Ecological and metabolic studies on free-living nematodes from an estuarine sand flat. Estuar. Coast. Mar. Sci. 1979, 9, 257–271. [Google Scholar] [CrossRef]
- Guo, Y.; Somerfield, P.J.; Warwick, R.M.; Zhang, Z. Large-scale patterns in the community structure and biodiversity of free living nematodes in the Bohai Sea, China. J. Mar. Biol. Assoc. U.K. 2001, 81, 755–763. [Google Scholar] [CrossRef] [Green Version]
- Balsamo, M.; Albertelli, G.; Ceccherelli, V.U.; Coccionia, R.; Colangeloc, M.A.; Curini-Gallettid, M.; Danovaroe, R.; D’Addabbof, R.; De Leonardisf, C.; Fabianob, M.; et al. Meiofauna of the Adriatic Sea: Present knowledge and future perspectives. Chem. Ecol. 2010, 26, 45–63. [Google Scholar] [CrossRef]
- Moreno, M.; Semprucci, F.; Vezzulli, L.; Balsamo, M.; Fabiano, M.; Albertelli, G. The use of nematodes in assessing ecological quality status in the Mediterranean coastal ecosystems. Ecol. Indic. 2011, 11, 328–336. [Google Scholar] [CrossRef]
- Semprucci, F.; Balsamo, M.; Appolloni, L.; Sandulli, R. Assessment of ecological quality status along the Apulian coasts (Eastern Mediterranean Sea) based on meiobenthic and nematode assemblages. Mar. Biodiver. 2018, 48, 105–115. [Google Scholar] [CrossRef]
- Hedfi, A.; Ben Ali, M.; Nourel-Deen, A.; Albogami, B.; Hassan, M.; Saif, T.; Boufahja, F. Distribution and Abundance of Benthic Meiofauna in the Eastern Red Sea Coasts (Jeddah, Saudi Arabia). ARRB 2018, 26, 1–12. [Google Scholar] [CrossRef]
- Mahmoudi, E.; Essid, N.; Beyrem, H.; Hedfi, A.; Boufahja, F.; Vtiello, P.; Aissa, P. Individual and combined effects of lead and zinc on a free-living marine nematode community: Results from microcosm experiments. J. Exp. Mar. Bio. Ecol. 2007, 343, 217–226. [Google Scholar] [CrossRef]
- Hedfi, A.; Mahmoudi, E.; Boufahja, F.; Beyrem, H.; Aïssa, P. Effects of increasing levels of nickel contamination on structure of offshore nematode communities in experimental microcosms. Bull. Environ. Contam. Toxicol. 2007, 79, 345–349. [Google Scholar] [CrossRef]
- Hedfi, A.; Boufahja, F.; Ben Ali, M.; Aïssa, P.; Mahmoudi, E.; Beyrem, H. Do trace metals (chromium, copper and nickel) influence toxicity of diesel fuel for free-living marine nematodes? Environ. Sci. Pollut. Res. 2013, 20, 3760–3770. [Google Scholar] [CrossRef]
- Hedfi, A. Les Nématodes Libres, Outils Communautaires de Biosurveillance du Milieu Marin: Étude In Situ et en Microcosme. Ph.D. Thesis, Carthage University, Bizerte, Tunisia, 2008. [Google Scholar]
- Schratzberger, M.; Whomersley, P.; Warr, K.; Bolam, S.G.; Rees, H.L. Colonisation of various types of sediment by estuarine nematodes via lateral infaunal migration: A laboratory study. Mar. Biol. 2004, 145, 69–78. [Google Scholar] [CrossRef]
- Wakkaf, T.; Allouche, M.; Harrath, A.H.; Mansour, L.; Alwasel, S.; Ansari, K.G.M.T.; Beyrem, H.; Sellami, B.; Boufahja, F. The individual and combined effects of cadmium, polyvinyl chloride (PVC) microplastics and their polyalkylamines modified forms on meiobenthic features in a microcosm. Environ. Pollut. 2020, 266, 115263. [Google Scholar] [CrossRef]
- Wieser, W. Benthic studies in buzzards bay. II. The meiofauna. Limnol. Oceanogr. 1960, 5, 121–137. [Google Scholar] [CrossRef]
- Elarbaoui, S.; Richard, M.; Boufahja, F.; Mahmoudi, E.; Thomas-Guyonc, H. Effect of crude oil exposure and dispersant application on meiofauna: An intertidal mesocosm experiment. Environ. Sci. Process. Impacts. 2015, 17, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Seinhorst, J.W. A rapid method for the transfer of nematodes from fixative to anhydrous glycerine. Nematologica. 1959, 4, 67–69. [Google Scholar] [CrossRef] [Green Version]
- Platt, H.M.; Warwick, R.M.; Furstenberg, J.P. Free-living marine nematodes. Part, I. British Enoploids. S. Afr. J. Zool. 1985, 20, 177. [Google Scholar] [CrossRef]
- Platt, H.M.; Warwick, R.M. Free-Living Marine Nematodes. Part II. British Chromadorids. Synopsis of the British Fauna (New Series No. 38); E.J. Brill/W.Backhuys: London, UK, 1988. [Google Scholar]
- Warwick, R.M.; Platt, H.M.; Somerfield, P.J. Free-Living Marine Nematodes. Part III. British Monohysterids. Synopsis of British Fauna (New Series No. 53); Field Studies Council: London, UK, 1998. [Google Scholar]
- Bezerra, T.N.; Decraemer, W.; Eisendle-Flockner, U.; Hodda, M.; Holovachov, O.; Leduc, D.; Miljutin, D.; Mokievsky, V.; Santiago, P.R.; Sharma, J.; et al. Nemys: World Database of Nematodes. 2020. Available online: http://nemys.ugent.be (accessed on 20 February 2021).
- Wieser, W. Die Beziehung zwischen Mundhöhlengestalt, Ernäh rungsweiseund Vorkommen bei freilebenden marinen Nematoden. Arkiv. För. Zoologi. 1953, 2, 439–484. [Google Scholar]
- Thistle, D.; Lambshead, P.J.D.; Sherman, K.M. Nematode tail-shape groups respond to environmental differences in the deep-sea. Vie Milieu. 1995, 45, 107–115. [Google Scholar]
- Bongers, T.; de Goede, R.G.M.; Korthals, G.W.; Yeates, G.W. An update to the cprating of nematode genera can be found in proposed changes of c-p classification for nematodes. Russ. J. Nematol. 1995, 3, 61–62. [Google Scholar]
- Schratzberger, M.; Warr, K.; Rogers, S.I. Functional diversity of nematode communities in the southwestern North Sea. Mar. Env. Res. 2007, 63, 368–389. [Google Scholar] [CrossRef] [Green Version]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMER-E., Ltd., Plymouth Marine Laboratory: London, UK, 2001. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v5: User Manual/Tutorial; PRIMER-E: London, UK, 2001. [Google Scholar]
- Allouche, M.; Nasri, A.; Harrath, A.H.; Mansour, L.; Alwasel, S.; Beyrem, H.; Plăvan, G.; Rohal-Lupher, M.; Boufahja, F. Meiobenthic nematode Oncholaimus campylocercoides as a model in laboratory studies: Selection, culture, and fluorescence microscopy After exposure to phenanthrene and chrysene. Environ. Sci. Pollut. Res. 2021, 28, 21303–21313. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.-S.; Metcalfe, C.D. Determination of cholesterol-lowering statin drugs in aqueous samples using liquid chromatography–electrospray ionization tandem mass spectrometry. J. Chromatogr. A 2003, 998, 133–141. [Google Scholar] [CrossRef]
- Dahl, U.; Gorokhova, E.; Breitholtz, M. Application of growth-related sublethal endpoints in ecotoxicological assessments using a harpacticoid copepod. Aquat. Toxicol. 2006, 77, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Kirstein, I.; Kirmizi, S.; Wichels, A.; Garin-Fernandez, A.; Erler, R.; Löder, M.; Gerdts, G. Dangerous hitchhikers? Evidence for potentially pathogenic Vibrio spp. on microplastic particles. Mar. Environ. Res. 2016, 120, 1–8. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Functional Traits | Treatments | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| c-p | Tl | Am | FG | Adult Length | C0 | P | S2 | S1 | A2 | A1 | SP2 | SP1 | AP2 | AP1 | |
| Anticoma acuminata | 2 | e/f | pk | 1A | 2–4 mm | 0.91 ± 0.78 | 0.46 ± 0.8 | 0.43 ± 0.75 | 0.47 ± 0.81 | 0.42 ± 0.72 | 1.63 ± 0.7 | 0.96 ± 0.83 | 0.44 ± 0.76 | 1.25 ± 1.27 | |
| Anticoma eberthi | 2 | e/f | pk | 1A | 2–4 mm | 8.27 ± 1.15 | 13.01 ± 1.37 | 13.55 ± 1.85 | 10.9 ± 1.85 | 5.35 ± 1.09 | 14.75 ± 1.06 | 14.28 ± 1.78 | 6.74 ± 0.88 | 8.32 ± 1.93 | 14.29 ± 3.02 |
| Calomicrolaimus honestus | 3 | co | sp | 2A | 1–2 mm | 2.75 ± 1.34 | 1.38 ± 1.37 | 1.26 ± 1.24 | 3.94 ± 2.3 | 1.8 ± 0.79 | 2.06 ± 1.46 | 1.63 ± 0.7 | 3.37 ± 0.85 | 1.75 ± 0.76 | 1.2 ± 1.19 |
| Chromadora sp. | 2 | co | id | 2A | 1–2 mm | 1.83 ± 0.75 | 4.19 ± 0.09 | 0.86 ± 0.75 | 0.84 ± 1.48 | 0.45 ± 0.78 | 1.22 ± 1.22 | 0.97 ± 0.84 | 0.43 ± 0.75 | ||
| Daptonema trabeculosum | 2 | cla | cr | 1B | 1–2 mm | 1.38 ± 1.35 | 2.79 ± 0.06 | 5.08 ± 1.26 | 1.75 ± 1.98 | 8.13 ± 1.72 | 4.92 ± 1.22 | 2.44 ± 1.21 | 3.85 ± 1.69 | 8.79 ± 2.12 | 6.1 ± 1.18 |
| Marylynnia puncticaudata | 3 | e/f | sp | 2A | 2–4 mm | 0.48 ± 0.82 | 0.91 ± 1.58 | 2.09 ± 1.39 | 2.65 ± 2.3 | 6.59 ± 4.48 | 5.72 ± 1.81 | 11.84 ± 1.94 | 6.74 ± 5.5 | 11. 83 ± 1.16 | 8.89 ± 2.29 |
| Metalinhomoeus numidicus | 2 | e/f | cr | 1B | 2–4 mm | 2.34 ± 1.68 | 5.58 ± 1.35 | 2.96 ± 0.65 | 6.99 ± 2.72 | 11.68 ± 0.94 | 5.72 ± 1.81 | 11.02 ± 1.16 | 14.42 ± 2.9 | 10.95 ± 1.41 | 10.54 ± 1.6 |
| Enoplolaimus littoralis | 3 | e/f | id | 2B | 2–4 mm | 0.45 ± 0.78 | 0.41 ± 0.71 | 0.43 ± 0.74 | 0.83 ± 1.07 | 1.63 ± 0.7 | 1.91 ± 2.18 | 3.94 ± 1.26 | 1.62 ± 0.68 | ||
| Metoncholaimus pristiurus | 3 | cla | pk | 2B | >4 mm | 8.31 ± 1.43 | 6.51 ± 2.1 | 8.03 ± 1.74 | 10.02 ± 1.8 | 10.76 ± 1.13 | 13.52 ± 1.07 | 8.17 ± 3.08 | 6.25 ± 1.64 | 6.15 ± 2.05 | 11.79 ± 3.02 |
| Odontophora villoti | 2 | co | cr | 1B | 2–4 mm | 0.45 ± 0.78 | 0.43 ± 0.75 | 0.48 ± 0.82 | |||||||
| Oncholaimus campylocercoides | 4 | cla | pk | 2B | >4 mm | 23.08 ± 2.28 | 15.84 ± 3.08 | 22.46 ± 4.06 | 24.44 ± 1.5 | 22.43 ± 0.6 | 25.82 ± 1.27 | 18.77 ± 0.65 | 23.56 ± 2.26 | 18 ± 1.74 | 23.18 ± 1.3 |
| Paracomesoma dubium | 2 | cla | sp | 2A | 1–2 mm | 0.45 ± 0.78 | 0.46 ± 0.82 | 0.41 ± 0.71 | 0.43 ± 0.74 | 0.92 ± 0.8 | 0.42 ± 0.72 | 0.81 ± 1.41 | 2.4 ± 0.83 | 1.75 ± 0.76 | 1.62 ± 0.68 |
| Paramonohystera wieseri | 2 | cla | cr | 1B | 1–2 mm | 1.4 ± 1.42 | 2.32 ± 1.6 | 4.28 ± 2.71 | 6.99 ± 2.72 | 3.53 ± 3.39 | 3.28 ± 0.73 | 6.55 ± 4.99 | 3.36 ± 1.66 | 7.02 ± 0.85 | 1.2 ± 2.09 |
| Paramonohystera pilosa | 2 | cla | cr | 1B | 1–2 mm | 0.46 ± 0.79 | 0.46 ± 0.8 | 0.43 ± 0.75 | 1.23 ± 1.22 | 0.41 ± 0.71 | 2.2 ± 0.78 | ||||
| Prochromadorella longicaudata | 2 | co | id | 2A | 1–2 mm | 42.38 ± 0.9 | 39.53 ± 1.85 | 22.06 ± 2.33 | 20.54 ± 1.21 | 19.82 ± 2.96 | 16.8 ± 0.48 | 13.47 ± 1.22 | 14.9 ± 0.71 | 7.01 ± 1.45 | 11.79 ± 3.02 |
| Spirinia gerlachi | 3 | co | cr | 2A | 1–2 mm | 0.46 ± 0.79 | 1.7 ± 0.75 | 0.88 ± 0.76 | 1.24 ± 1.25 | 0.41 ± 0.71 | 2.39 ± 1.64 | 0.43 ± 0.75 | 0.82 ± 0.71 | ||
| Steineria sp. | 2 | cla | cr | 1B | 1–2 mm | 0.46 ± 0.79 | 0.41 ± 0.71 | 0.48 ± 0.84 | 1.31 ± 1.32 | ||||||
| Synonchiella edax | 4 | e/f | sp | 2B | 2–4 mm | 0.46 ± 0.79 | 0.93 ± 0.81 | 1.7 ± 0.75 | 0.82 ± 0.71 | 0.41 ± 0.71 | 1.93 ± 0.84 | 3.07 ± 1.52 | 1.65 ± 1.43 | ||
| Theristus flevensis | 2 | co | cr | 1B | 1–2 mm | 3.24 ± 0.9 | 5.13 ± 2.2 | 11.03 ± 1.57 | 4.83 ± 2.1 | 7.65 ± 2.23 | 2.04 ± 0.69 | 5.31 ± 1.43 | 5.29 ± 1.69 | 6.6 ± 2.37 | 4.06 ± 0.62 |
| Viscosia cobbi | 3 | e/f | pk | 2B | 1–2 mm | 0.46 ± 0.79 | 0.48 ± 0.82 | 1.28 ± 1.3 | 3.5 ± 0.79 | 0.43 ± 0.74 | 0.42 ± 0.7 | ||||
| C0 vs. P | C0 vs. S2 | C0 vs. S1 | C0 vs. A2 | C0 vs. A1 | C0 vs. SP2 | C0 vs. SP1 | C0 vs. AP2 | C0 vs. AP1 | |
|---|---|---|---|---|---|---|---|---|---|
| Average dissimilarity (%) | 20.04 | 29.04 | 28.67 | 33.11 | 34.17 | 38.66 | 35.35 | 47.55 | 39.79 |
| R-statistics | 0.422 | 0.704 | 0.593 | 0.741 | 0.852 | 0.852 | 0.852 | 1 | 1 |
| p-value | 0.029 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 |
| Species | Oncholaimus campylocercoides 18.47% − | Prochromadorella longicaudata 30.37% − | Prochromadorella longicaudata 35.16% − | Prochromadorella longicaudata 32.88% − | Prochromadorella longicaudata 32.36% − | Prochromadorella longicaudata 33.01% − | Prochromadorella longicaudata 40.58% − | Prochromadorella longicaudata 35.91% − | Prochromadorella longicaudata 34.19% − |
| Anticoma eberthi 11.59% + | Theristus flevensis 14.44% + | Paramonohysteria weiseri 10.15% + | Metalinhomoeus numidicus 14.4% + | Anticoma eberthi 11.44% + | Marylynnia puncticaudata 15.67% + | Metalinhomoeus numidicus 16.62% + | Marylynnia puncticaudata 12.28% + | Metalinhomoeus numidicus 11.36% + | |
| Metoncholaimus pristiurus 11.58% − | Anticoma eberthi 10.65% + | Metalinhomoeus numidicus 8.58% + | Daptonema trabeculosum 10.34% + | Metoncholaimus pristiurus 9.51% + | Metalinhomoeus numidicus 12.3% + | Marylynnia puncticaudata 8.65% + | Metalinhomoeus numidicus 9.43% + | Marylynnia puncticaudata 11.35% + | |
| Prochromadorella longicaudata 8.8% − | Daptonema trabeculosum 6.84% + | Viscosia cobbi 5.48% + | Marylynnia puncticaudata 9.49% + | Oncholaimus campylocercoides 8.23% + | Daptonema trabeculosum 3.34% + | Daptonema trabeculosum 8.05% + | Anticoma eberthi 9.28% + | ||
| Theristus flevensis 8.45% + | Oncholaimus campylocercoides 6.34% + | Anticoma eberthi 5.48% + | Marylynnia puncticaudata 8.23% + | ||||||
| Metalinhomoeus numidicus 8.02% + | |||||||||
| Feeding groups | 12.36% | 23.49% | 21.34% | 27.36% | 27.36% | 30.11% | 25.22% | 36.53% | 31.82% |
| 2B − | 2A − | 2A− | 2A− | 2A− | 2A− | 2A− | 2A− | 2A− | |
| 1B + | 2B+ | ||||||||
| Tail shape | 9.09% | 14.77% | 20.16% | 21.75% | 30.58% | 30.74% | 23.93% | 35.74% | 35.22% |
| e/f + | co − | co − | co − | co − | e/f + | co − | co − | co − | |
| Amphid shape | 8.50% | 23.02% | 23.78% | 26.79% | 29.29% | 30.75% | 31.62% | 40.69% | 33.91% |
| cr + | id − | id − | id − | id − | id − | id − | id − | id − | |
| pk − | cr + | pk + | |||||||
| Adult length | 10.02% | 9.02% | 12.09% | 13.74% | 23.64% | 26.69% | 23.76% | 25.52% | 30.91% |
| >4mm + | 2–4mm + | 2–4mm + | 1–2mm − | 1–2mm − | 2–4mm + | 1–2mm − | 2–4mm + | 2–4mm + | |
| c-p score | 11.86% | 7.13% | 9.58% | 7.52% | 13.25% | 9.96% | 11.53% | 10.71% | 11.87% |
| cp2 + | cp2 + | cp3 + | cp3 + | cp3 + | cp3 + | cp2 − | cp3 + | cp3+ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Allouche, M.; Ishak, S.; Nasri, A.; Hedfi, A.; Ben Ali, M.; Albogami, B.; Almalki, M.; Pacioglu, O.; Boufahja, F. What Is the Impact of Microplastics and Lipid Regulators on Marine Meiofauna? Case Study of Polyvinyl Chloride, Atorvastatin, and Simvastatin. Sustainability 2021, 13, 13190. https://doi.org/10.3390/su132313190
Allouche M, Ishak S, Nasri A, Hedfi A, Ben Ali M, Albogami B, Almalki M, Pacioglu O, Boufahja F. What Is the Impact of Microplastics and Lipid Regulators on Marine Meiofauna? Case Study of Polyvinyl Chloride, Atorvastatin, and Simvastatin. Sustainability. 2021; 13(23):13190. https://doi.org/10.3390/su132313190
Chicago/Turabian StyleAllouche, Mohamed, Sahar Ishak, Ahmed Nasri, Amor Hedfi, Manel Ben Ali, Bander Albogami, Mohammed Almalki, Octavian Pacioglu, and Fehmi Boufahja. 2021. "What Is the Impact of Microplastics and Lipid Regulators on Marine Meiofauna? Case Study of Polyvinyl Chloride, Atorvastatin, and Simvastatin" Sustainability 13, no. 23: 13190. https://doi.org/10.3390/su132313190
APA StyleAllouche, M., Ishak, S., Nasri, A., Hedfi, A., Ben Ali, M., Albogami, B., Almalki, M., Pacioglu, O., & Boufahja, F. (2021). What Is the Impact of Microplastics and Lipid Regulators on Marine Meiofauna? Case Study of Polyvinyl Chloride, Atorvastatin, and Simvastatin. Sustainability, 13(23), 13190. https://doi.org/10.3390/su132313190