
How Per- and Poly-Fluoroalkyl Substances Affect Gamete Viability and Fertilization Capability: Insights from the Literature



Abstract
1. Introduction
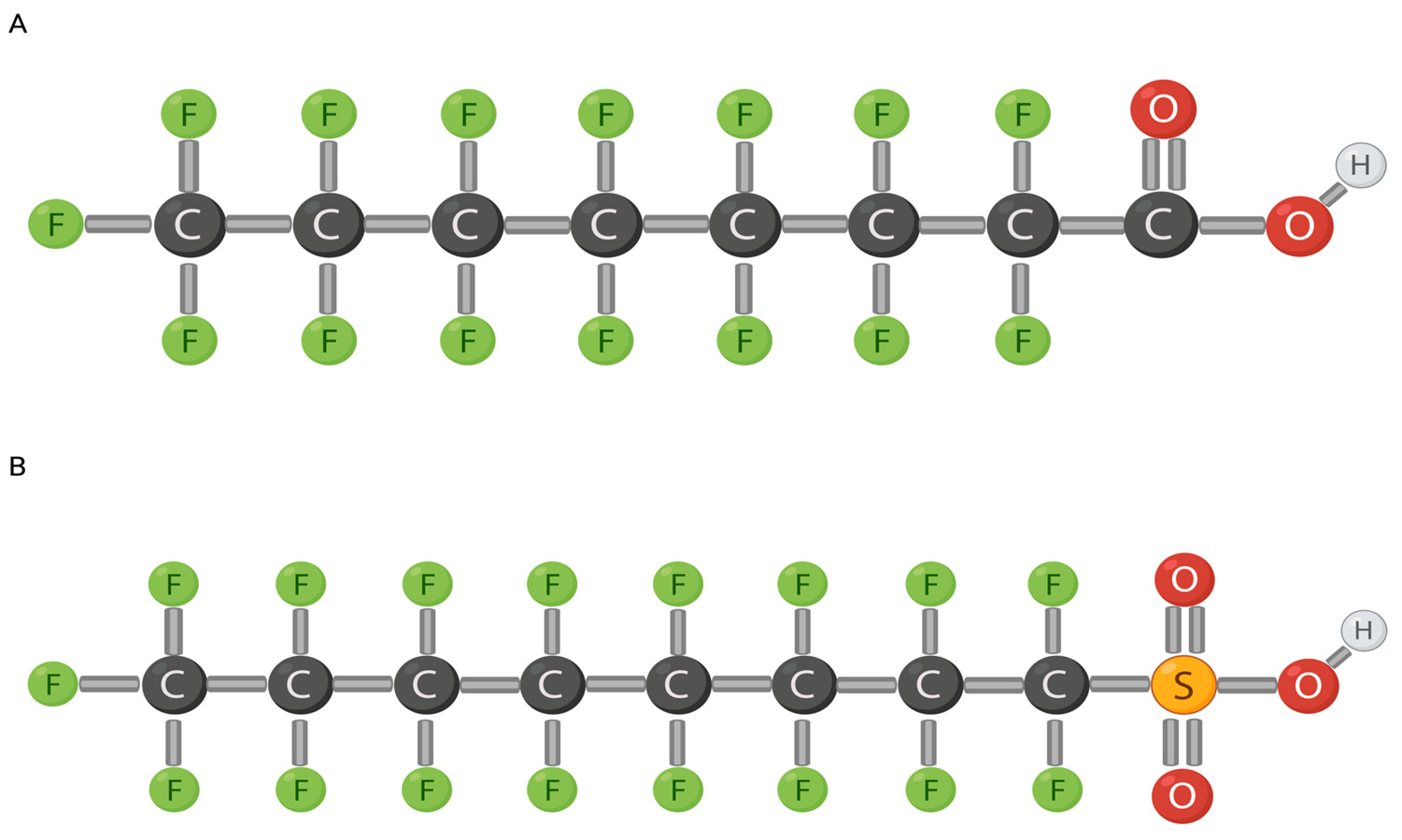
1.1. PFOA and PFOS and Their Mechanism of Action
1.2. Toxic Environmental Exposure
2. Methods
3. Effects on Reproductive Hormones
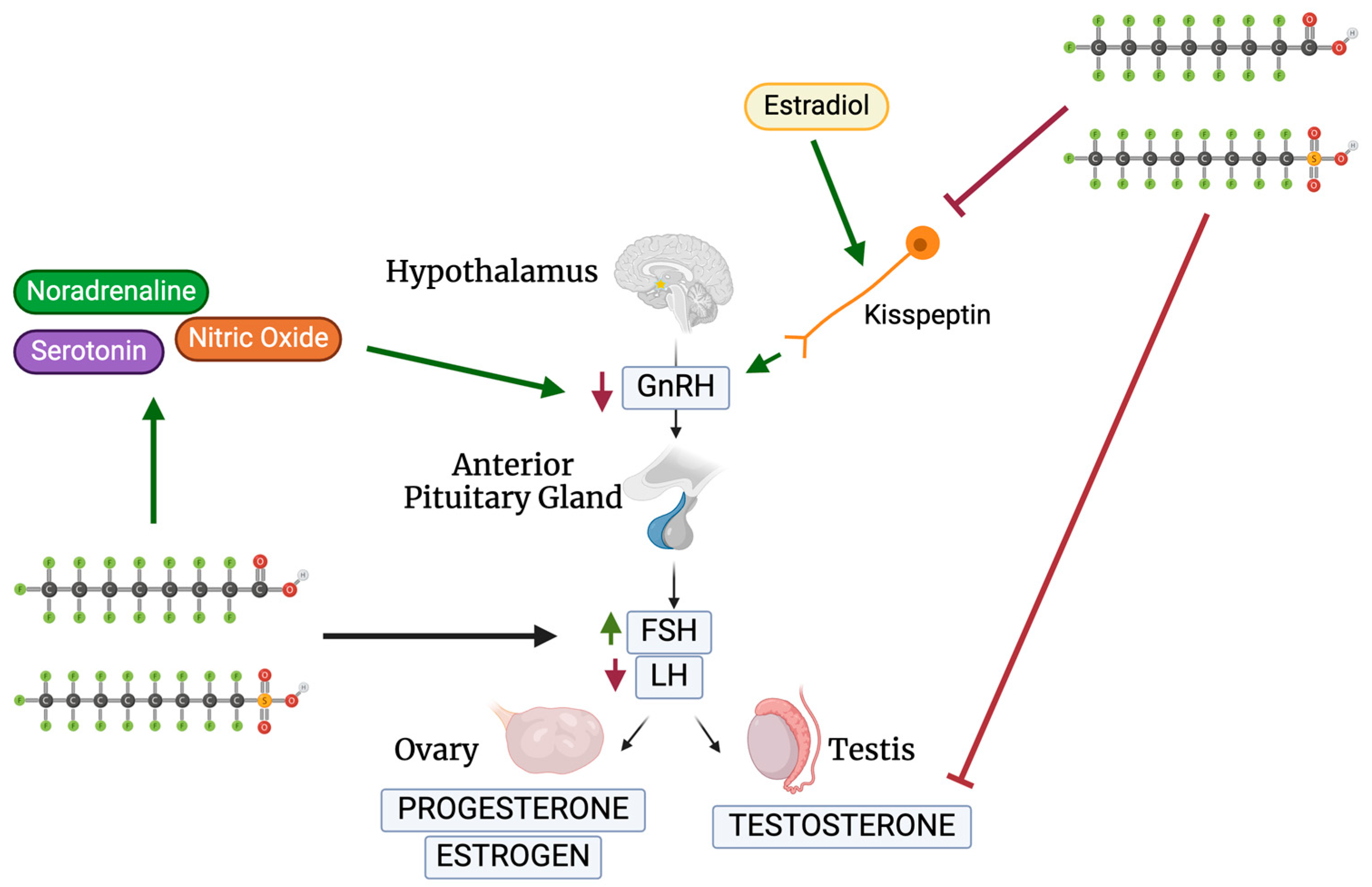
3.1. GnRH
3.2. FSH and LH
3.3. Testosterone Levels in Females
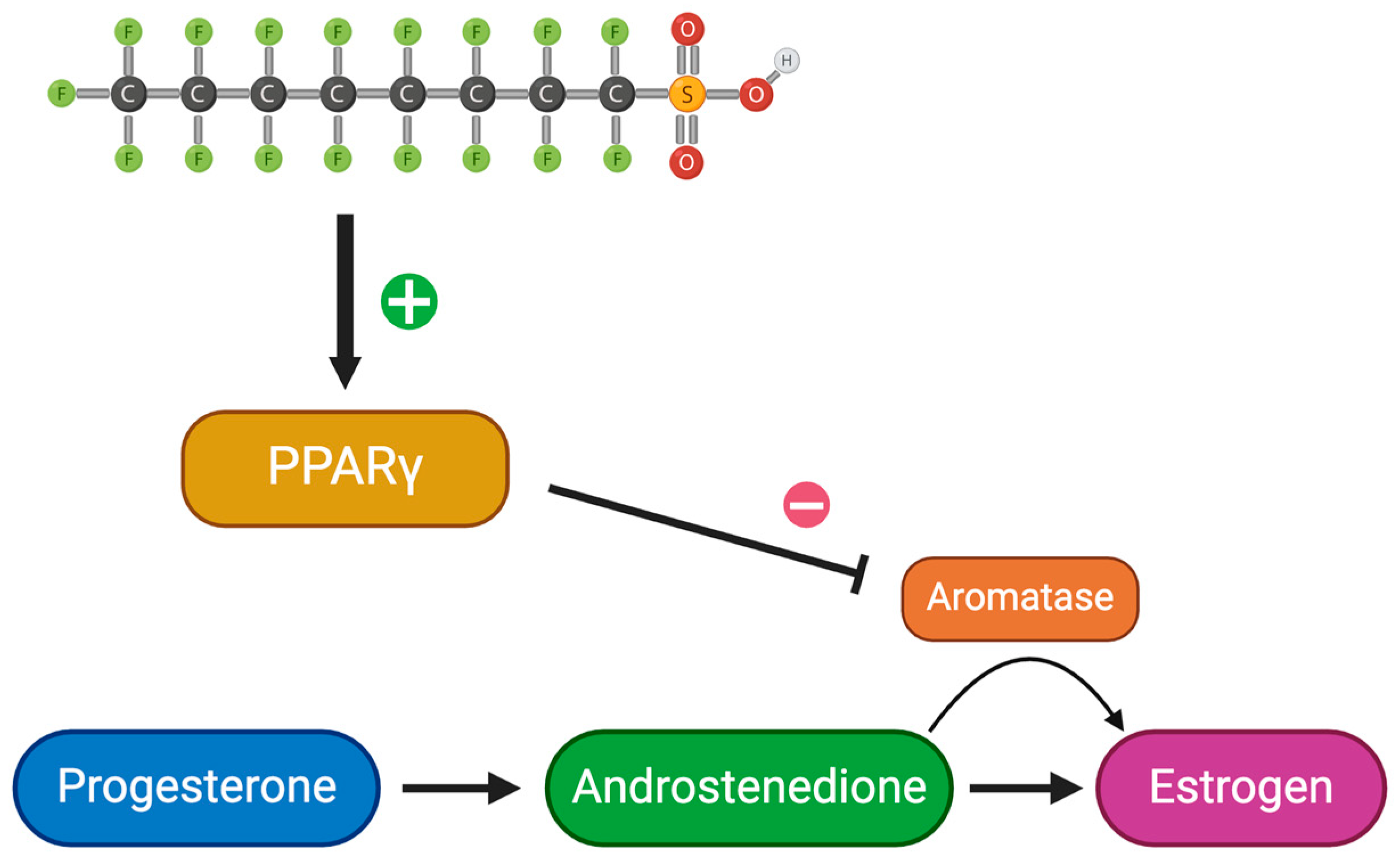
3.4. Cyp17 Gene Expression

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | In Vivo/In Vitro | Cell Type | Endpoint | Treatment Dose (mg/kg) | Effect Observed | Reference |
|---|---|---|---|---|---|---|
| Rattus rattus | in vivo | n/a | Detect a change in sexual maturation, looking at Kiss1, after PFAS exposure | 0.1, 1.0, and 10 mg/kg of PFOA or PFOS during PND1-5 | - 0.1 mg/kg: increased expression of Kiss1 - 10 mg/kg: decreased expression of Kiss1 | [39] |
| Rattus rattus | in vivo | n/a | Effects of PFOS exposure in GnRH | 0.5, 1.0, 3.0, 6.0 mg/kg/day for 28 days | - Decreased GnRH expression at 0.5, 1.0, and 3.0 mg/kg - Increased noradrenaline - GnRH receptor protein was inhibited at all doses | [43] |
| Rattus rattus | in vivo | n/a | Neuroendocrine effects of PFOS in regard to the estrous cycle | 1 and 10 mg/kg of PFOS for 2 weeks | - Induced norepinephrine in hypothalamus - Inhibited regular estrous cyclicity to 66% at 1 mg/kg and 42% at 10 mg/kg - Increased diestrus persistence from 8% to 33% | [44] |
| Mus musculus | in vivo | n/a | PFOS exposure effects on ovulation stages of mice | 10 mg/kg of PFOS for 14 days | - PFOS exposure decreased LH and FSH - LH elevated during diestrus - E2 and Kiss1 decreased | [46] |
| Sus scrofa | in vitro | Porcine theca and granulosa cells | PFAS’ effects on porcine ovarian cell steroidogenesis | 0.12, 1.2, 12, and 120 µM of PFOS and PFOA | PFOA: - PPARγ affects granulosa cells, −0.12 decreased P4 and E2 PFOS: - PPARα and PPARβ/δ affect theca cells - 1.2 µM+ increased P4 and decreased A4 | [47] |
| Danio Rerio | in vivo | n/a | Observe the implications of different genotypes of the Cyp17a1 gene | n/a | - Cyp17a1−/−: thinner efferent ducts and gonad degeneration - Decrease in spermatogenic cell count and sterility | [50] |
4. Effects on Female Gamete Development
4.1. Oocyte Competence
4.2. Oocyte Reserve
4.3. Chromosome Misalignment, F-Actin Organization, and Spindle Formation
| Species | Endpoint | Treatment Dose | Effect Observed | Reference |
|---|---|---|---|---|
| Mus musculus | PFOA effects on ROS levels in the ovary ex vivo | 28.2 and 112.8 μM of PFOA for 24 h | - Significant increase in fluorescence - 112.8 μM: same fluorescence as positive control | [51] |
| PFOA effects on GJIC | 2.5 mg/kg/daily for 3 days | - DMSO: high fluorescence - PFOA-COC: no fluorescence | ||
| Mus musculus | Impact of PFOS exposure on ovarian hormone production | 0.1 mg/kg for 6 months | - Inhibited ovarian hormones and follicular development | [56] |
| Mus musculus | Exposure to PFOA’s impact on the estrous cycle | Low dose, 1 mg/kg/d, and high dose, 5 mg/kg/d, of PFOA for 28 days | Low-dose group: - Significantly shorter estrus phase - Significant decrease in primordial follicles High dose: - Significantly extended metestrus and anestrus phases - Significant decrease in total ovarian follicle count Both groups: - Significantly smaller litter groups and ovaries | [52] |
| Unstable follicular ovulation and development following PFOA exposure | Low dose, 1 mg/kg/d, and high dose, 5 mg/kg/d, of PFOA for 28 days | - GVBD rate significantly reduced after 3 h - Oocytes in low-dose group could not resume meiosis |
| Species | Cell Type | Endpoint | Treatment Dose (µM) | Effect Observed | Reference |
|---|---|---|---|---|---|
| Mus musculus | Denuded murine oocytes | PFOA effects on mouse oocytes | 50, 100, and 150 μM of PFOA for 24 h | - 50 μM: no change - 150 μM: death of all oocytes | [51] |
| Observe apoptotic and necrotic effects in PFOA exposed oocytes | 28.2 and 112.8 μM of PFOA for 24 h | - Significant increase in apoptotic and necrotic oocytes in both exposed groups | |||
| Sus scrofa | Porcine oocytes | Effects of PFOS on the number of live oocytes after exposure | 12.5, 25, and 50 μM of PFOS of 44 h | From an initial 210 oocytes examined: - 25 μM: significant drop to 126 live oocytes - 50 μM: significant drop to 4 live oocytes | [53] |
| Effects of PFOS on the number of mature oocytes after exposure | 12.5, 25, and 50 μM of PFOS for 44 h | Control: 13 GV, 21 MI, and 67 MII 50 μM: 67 GV, 26 MI, and 5 MII | |||
| Mus musculus | Murine oocytes | Impact of PFAS exposure on embryo quality | 600 μM of PFOS and 600 μM of PFOA | - Lower GVBD and PBE rate - Cleavage rates of PFOS exposed oocytes were significantly lower than control rates (62.2%) | [20] |
| PFAS exposure effects on oocyte maturation | 600 μM PFOS | - Increased PB size - Significant increase in spindle migration |
5. Effects on Male Gamete and Fertilization Capability
5.1. Sperm Viability
| Species | In Vivo/In Vitro | Cell Type | Endpoint | Treatment Dose | Effect Observed | Reference |
|---|---|---|---|---|---|---|
| Animal Studies | ||||||
| Rattus rattus | in vivo | n/a | PFOS and PFOA exposure effects in Leydig cells | 0, 5, and 20 mg/kg PFOS | - Leydig cell hyperplasia and adenomas resulted - Decreased testosterone resulted by inhibition of 3B-HSDH and 17BHSDH3 | [61] |
| Rattus rattus | in vivo | n/a | Effect of Leydig cell regeneration after PFOS exposure | 0.5, 10 mg/kg of PFOS for 56 days | 35 days post: no testosterone levels - Significant decrease Both groups: significantly low testosterone levels day 56 - Inhibited LC regeneration and proliferation - CYP11A1 was decreased at 5 and 10 mg/kg day 56 and 11B-HSD1 reduced at 10 mg/kg for both days 35 and 56 | [65] |
| Mus musculus | in vivo | n/a | Determine testicular effects of PFOS exposure | 0.5, 5, and 10 mg/kg of PFOS for 4 weeks | - Significant decrease in mRNA presence of StAR - 5 and 10 mg/kg: significant decrease in sperm count and testosterone levels - 10 mg/kg: Germ cell degeneration | [66] |
| in vitro | Murine Leydig cells | Proving testicular effects occur after PFOS exposure | 0, 15, and 30 μM PFOS | - 15 and 30 mg/kg: Significant decrease in testosterone and mRNA StAR | ||
| Human Studies | ||||||
| Homo sapiens | in vitro | Human spermatozoa | PFAS exposure impairs spermatozoa quality | Quartile score: Low group: 2–3 Intermediate: 4–6 High group: 7–8 | - High: significantly decreased average number of normal spermatozoa - High: 6.2 million normal -Low: 15.5 million | [67] |
5.2. Capacitation Reaction
5.3. Acrosomal Reaction
6. Effects on Embryo Pre-Implantation
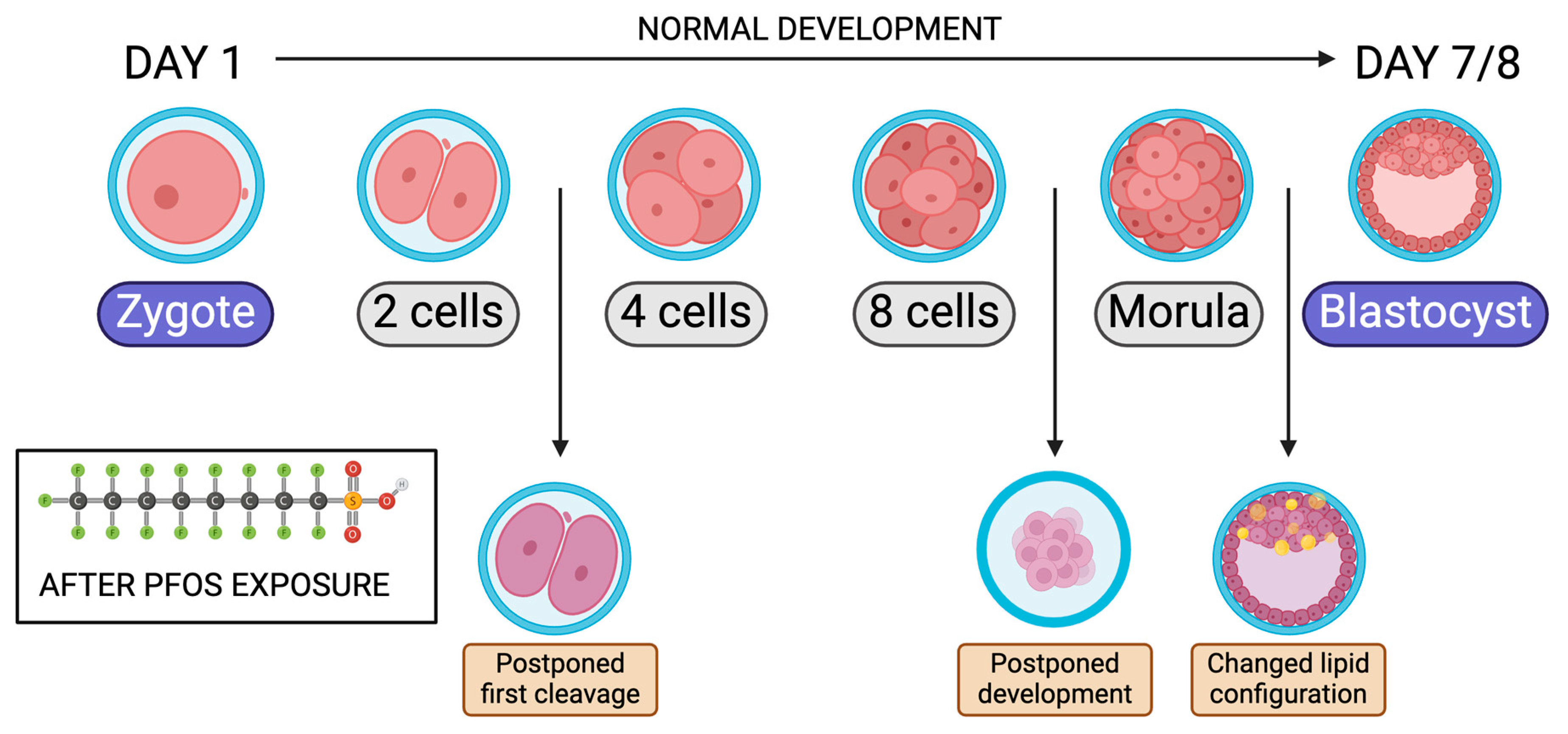
| Species | Cell Type | In Vivo/Vitro | Endpoint | Treatment Dose | Effect Observed | Reference |
|---|---|---|---|---|---|---|
| Animal Studies | ||||||
| Bos taurus | Bovine blastocysts | in vitro | Effects of PFNA on lipid accumulation in blastocysts | 10 and 0.1 µg/mL PFAS | - PFNA effects lipid metabolism in blastocysts - Cumulus cloud in 10 µg/mL group did not expand - Significant difference in lipid droplets | [77] |
| Bos taurus | Bovine blastocysts | in vitro | Effects of PFOS on bovine embryonic development | 2 ng/mL and 53 ng/mL of PFOS | - 53 ng/mL of PFOS significantly decreased cleavage past the 2-cell stage - Results of early delay were not noticeable in later embryonic development | [78] |
| Bos taurus | Bovine blastocysts | in vitro | Effects of PFAS on bovine COC fertilization and maturation | 0.01, 0.1, 1.0, 10, 20, 40, and 100 μg/mL PFAS before fertilization and cultured for 8 days | - 10, 40, and 100 μg/mL showed a decrease in passing the 2-cell stage - 40 µg/mL + decreased total cell count | [79] |
| Mus musculus | Murine zygotes | in vitro | Effects of PFOS exposure on ICM of pre-implantation embryo | 10 nM and 100 nM PFOS | - 100 nM: affects the blastocyst formation rate - Increased ROS and apoptosis - Impaired ICM in pre-implantation embryos | [80] |
| Human Studies | ||||||
| Homo sapiens | Human endometrial epithelial adenocarcinoma Ishikawa cells | in vitro | Effects of PFOA exposure on progesterone in endometrial cells | 0.5, 1, 1.8, 3, 5, 7, 8 mM of PFOA | - PFOA inhibits estrogen sulfotransferase genes - mRNA levels—of ITGB8, ALPPL2, and KLF5—are antagonized by PFOA, impairing embryo attachment - The endometrium responds more readily to PFOA rather than the hypothalamic–ovarian axis | [72] |
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Meegoda, J.N.; Kewalramani, J.A.; Li, B.; Marsh, R.W. A Review of the Applications, Environmental Release, and Remediation Technologies of Per- and Polyfluoroalkyl Substances. Int. J. Environ. Res. Public Health 2020, 17, 8117. [Google Scholar] [CrossRef] [PubMed]
- Rickard, B.P.; Rizvi, I.; Fenton, S.E. Per- and Poly-Fluoroalkyl Substances (PFAS) and Female Reproductive Outcomes: PFAS Elimination, Endocrine-Mediated Effects, and Disease. Toxicology 2022, 465, 153031. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.R.; White, N.; Bräunig, J.; Vijayasarathy, S.; Mueller, J.F.; Knox, C.L.; Harden, F.A.; Pacella, R.; Toms, L.-M.L. Per- and Poly-Fluoroalkyl Substances (PFASs) in Follicular Fluid from Women Experiencing Infertility in Australia. Environ. Res. 2020, 190, 109963. [Google Scholar] [CrossRef] [PubMed]
- Fei, C.; McLaughlin, J.K.; Tarone, R.E.; Olsen, J. Fetal Growth Indicators and Perfluorinated Chemicals: A Study in the Danish National Birth Cohort. Am. J. Epidemiol. 2008, 168, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Jane, L.; Espartero, L.; Yamada, M.; Ford, J.; Owens, G.; Prow, T.; Juhasz, A. Health-Related Toxicity of Emerging per- and Polyfluoroalkyl Substances: Comparison to Legacy PFOS and PFOA. Environ. Res. 2022, 212, 113431. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. Perfluorooctanesulfonic Acid. PubChem Compound Database. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Perfluorooctanesulfonic-acid (accessed on 1 May 2024).
- Ghisari, M.; Long, M.; Røge, D.M.; Olsen, J.; Bonefeld-Jørgensen, E.C. Polymorphism in Xenobiotic and Estrogen Metabolizing Genes, Exposure to Perfluorinated Compounds and Subsequent Breast Cancer Risk: A Nested Case-Control Study in the Danish National Birth Cohort. Environ. Res. 2017, 154, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Crone, B.C.; Speth, T.F.; Wahman, D.G.; Smith, S.J.; Abulikemu, G.; Kleiner, E.J.; Pressman, J.G. Occurrence of Per- and Polyfluoroalkyl Substances (PFAS) in Source Water and Their Treatment in Drinking Water. Crit. Rev. Environ. Sci. Technol. 2019, 49, 2359–2396. [Google Scholar] [CrossRef] [PubMed]
- Haug, M.; Dunder, L.; Lind, P.M.; Lind, L.; Salihovic, S. Associations of Perfluoroalkyl Substances (PFAS) with Lipid and Lipoprotein Profiles. J. Expo. Sci. Environ. Epidemiol. 2023, 33, 757–765. [Google Scholar] [CrossRef]
- Gaballah, S.; Swank, A.; Sobus, J.R.; Howey, X.M.; Schmid, J.; Catron, T.; McCord, J.; Hines, E.; Strynar, M.; Tal, T. Evaluation of Developmental Toxicity, Developmental Neurotoxicity, and Tissue Dose in Zebrafish Exposed to GenX and Other PFAS. Environ. Health Perspect. 2020, 128, 047005. [Google Scholar] [CrossRef]
- White, S.S.; Calafat, A.M.; Kuklenyik, Z.; Villanueva, L.; Zehr, R.D.; Helfant, L.; Strynar, M.J.; Lindstrom, A.B.; Thibodeaux, J.R.; Wood, C.; et al. Gestational PFOA Exposure of Mice Is Associated with Altered Mammary Gland Development in Dams and Female Offspring. Toxicol. Sci. 2006, 96, 133–144. [Google Scholar] [CrossRef]
- Oseguera-López, I.; Pérez-Cerezales, S.; Ortiz-Sánchez, P.B.; Mondragon-Payne, O.; Sánchez-Sánchez, R.; Jiménez-Morales, I.; Fierro, R.; González-Márquez, H. Perfluorooctane Sulfonate (PFOS) and Perfluorohexane Sulfonate (PFHxS) Alters Protein Phosphorylation, Increase ROS Levels and DNA Fragmentation during In Vitro Capacitation of Boar Spermatozoa. Animals 2020, 10, 1934. [Google Scholar] [CrossRef]
- Ding, N.; Harlow, S.D.; Randolph, J.F., Jr.; Loch-Caruso, R.; Park, S.K. Perfluoroalkyl and Polyfluoroalkyl Substances (PFAS) and Their Effects on the Ovary. Hum. Reprod. Update 2020, 26, 724–752. [Google Scholar] [CrossRef]
- White, S.S.; Stanko, J.P.; Kato, K.; Calafat, A.M.; Hines, E.P.; Fenton, S.E. Gestational and Chronic Low-Dose PFOA Exposures and Mammary Gland Growth and Differentiation in Three Generations of CD-1 Mice. Environ. Health Perspect. 2011, 119, 1070–1076. [Google Scholar] [CrossRef]
- Taibl, K.R.; Dunlop, A.L.; Barr, D.B.; Li, Y.-Y.; Eick, S.M.; Kannan, K.; Ryan, P.B.; Schroder, M.; Rushing, B.; Fennell, T.; et al. Newborn Metabolomic Signatures of Maternal Per- and Polyfluoroalkyl Substance Exposure and Reduced Length of Gestation. Nat. Commun. 2023, 14, 3120. [Google Scholar] [CrossRef]
- Hall, S.M.; Zhang, S.; Hoffman, K.; Miranda, M.L.; Stapleton, H.M. Concentrations of Per- and Polyfluoroalkyl Substances (PFAS) in Human Placental Tissues and Associations with Birth Outcomes. Chemosphere 2022, 295, 133873. [Google Scholar] [CrossRef]
- Ma, X.; Cui, L.; Chen, L.; Zhang, J.; Zhang, X.; Kang, Q.; Jin, F.; Ye, Y. Parental Plasma Concentrations of Perfluoroalkyl Substances and In Vitro Fertilization Outcomes. Environ. Pollut. 2021, 269, 116159. [Google Scholar] [CrossRef]
- Ortiz-Sánchez, P.B.; Roa-Espitia, A.L.; Fierro, R.; López-Torres, A.S.; Jiménez-Morales, I.; Oseguera-López, I.; Hernández-González, E.O.; González-Márquez, H. Perfluorooctane Sulfonate and Perfluorooctanoic Acid Induce Plasma Membrane Dysfunction in Boar Spermatozoa during in Vitro Capacitation. Reprod. Toxicol. 2022, 110, 85–96. [Google Scholar] [CrossRef]
- Björvang, R.D.; Hallberg, I.; Pikki, A.; Berglund, L.; Pedrelli, M.; Kiviranta, H.; Rantakokko, P.; Ruokojärvi, P.; Lindh, C.H.; Olovsson, M.; et al. Follicular Fluid and Blood Levels of Persistent Organic Pollutants and Reproductive Outcomes among Women Undergoing Assisted Reproductive Technologies. Environ. Res. 2022, 208, 112626. [Google Scholar] [CrossRef]
- Feng, J.; Soto-Moreno, E.J.; Prakash, A.; Balboula, A.Z.; Qiao, H. Adverse PFAS Effects on Mouse Oocyte in Vitro Maturation Are Associated with Carbon-chain Length and Inclusion of a Sulfonate Group. Cell Prolif. 2023, 56, e13353. [Google Scholar] [CrossRef]
- Governini, L.; Orvieto, R.; Guerranti, C.; Gambera, L.; De Leo, V.; Piomboni, P. The Impact of Environmental Exposure to Perfluorinated Compounds on Oocyte Fertilization Capacity. J. Assist. Reprod. Genet. 2011, 28, 415–418. [Google Scholar] [CrossRef]
- Hallberg, I.; Björvang, R.D.; Hadziosmanovic, N.; Koekkoekk, J.; Pikki, A.; Van Duursen, M.; Lenters, V.; Sjunnesson, Y.; Holte, J.; Berglund, L.; et al. Associations between Lifestyle Factors and Levels of Per- and Polyfluoroalkyl Substances (PFASs), Phthalates and Parabens in Follicular Fluid in Women Undergoing Fertility Treatment. J. Expo. Sci. Environ. Epidemiol. 2023, 33, 699–709. [Google Scholar] [CrossRef]
- Shen, H.; Gao, M.; Li, Q.; Sun, H.; Jiang, Y.; Liu, L.; Wu, J.; Yu, X.; Jia, T.; Xin, Y.; et al. Effect of PFOA Exposure on Diminished Ovarian Reserve and Its Metabolism. Reprod. Biol. Endocrinol. 2023, 21, 16. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, L.; Ducatman, A.; Deng, C.; Von Stackelberg, K.E.; Danford, C.J.; Zhang, X. Association of Per- and Polyfluoroalkyl Substance Exposure with Fatty Liver Disease Risk in US Adults. JHEP Rep. 2023, 5, 100694. [Google Scholar] [CrossRef]
- Björvang, R.D.; Damdimopoulou, P. Persistent Environmental Endocrine-Disrupting Chemicals in Ovarian Follicular Fluid and in Vitro Fertilization Treatment Outcome in Women. Upsala J. Med. Sci. 2020, 125, 85–94. [Google Scholar] [CrossRef]
- Brase, R.A.; Mullin, E.J.; Spink, D.C. Legacy and Emerging Per- and Polyfluoroalkyl Substances: Analytical Techniques, Environmental Fate, and Health Effects. Int. J. Mol. Sci. 2021, 22, 995. [Google Scholar] [CrossRef]
- Lin, C.; Nian, M.; Xie, M.; Zeng, L.; Qiu, W.; Zhang, J.; Yang, H. Associations of Per- and Polyfluoroalkyl Substances and Alternatives with Reproductive Hormones in Women of Childbearing Age. Int. J. Hyg. Environ. Health 2023, 250, 114158. [Google Scholar] [CrossRef]
- Behr, A.-C.; Plinsch, C.; Braeuning, A.; Buhrke, T. Activation of Human Nuclear Receptors by Perfluoroalkylated Substances (PFAS). Toxicol. Vitr. 2020, 62, 104700. [Google Scholar] [CrossRef]
- Government of California. Potential Designated Chemicals: Perfluoroalkyl and Polyfluoroalkyl Substances (PFASs). Meeting of the Scientific Guidance Panel. 2015. Available online: https://biomonitoring.ca.gov/sites/default/files/downloads/PotenDesigPFASs_031315.pdf (accessed on 25 April 2024).
- Knox, S.S.; Jackson, T.; Javins, B.; Frisbee, S.J.; Shankar, A.; Ducatman, A.M. Implications of Early Menopause in Women Exposed to Perfluorocarbons. J. Clin. Endocrinol. Metab. 2011, 96, 1747–1753. [Google Scholar] [CrossRef]
- Tarapore, P.; Ouyang, B. Perfluoroalkyl Chemicals and Male Reproductive Health: Do PFOA and PFOS Increase Risk for Male Infertility? Int. J. Environ. Res. Public Health 2021, 18, 3794. [Google Scholar] [CrossRef]
- Heffernan, A.L.; Cunningham, T.K.; Drage, D.S.; Aylward, L.L.; Thompson, K.; Vijayasarathy, S.; Mueller, J.F.; Atkin, S.L.; Sathyapalan, T. Perfluorinated Alkyl Acids in the Serum and Follicular Fluid of UK Women with and without Polycystic Ovarian Syndrome Undergoing Fertility Treatment and Associations with Hormonal and Metabolic Parameters. Int. J. Hyg. Environ. Health 2018, 221, 1068–1075. [Google Scholar] [CrossRef]
- Government of Canada. Perfluorooctane Sulfonate (PFOS) and Health. 2007. Available online: https://www.canada.ca/en/health-canada/services/environmental-workplace-health/reports-publications/environmental-contaminants/perfluorooctane-sulfonate-pfos-health.html (accessed on 1 May 2024).
- PFAS Testing and Concentrations to Inform Clinical Care of Exposed Patients. National Academies of Sciences, Engineering, and Medicine; Health and Medicine Division; Division on Earth and Life Studies; Board on Population Health and Public Health Practice; Board on Environmental Studies and Toxicology. 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK584705/ (accessed on 25 April 2024).
- Olsen, G.W.; Gilliland, F.D.; Burlew, M.M.; Burris, J.M.; Mandel, J.S.; Mandel, J.H. An Epidemiologic Investigation of Reproductive Hormones in Men with Occupational Exposure to Perfluorooctanoic Acid. J. Occup. Environ. Med. 1998, 40, 614–622. [Google Scholar] [CrossRef]
- Government of Canada. Per- and Polyfluoroalkyl Substances. 2007. Available online: https://www.canada.ca/en/health-canada/services/chemical-substances/other-chemical-substances-interest/per-polyfluoroalkyl-substances.html (accessed on 1 May 2024).
- Roepke, T.A.; Sadlier, N.C. Reproductive Toxicology: Impact of Endocrine Disruptors on Neurons Expressing GnRH or Kisspeptin and Pituitary Gonadotropins. Reproduction 2021, 162, F131–F145. [Google Scholar] [CrossRef]
- Sun, Z.; Wen, Y.; Wang, B.; Deng, S.; Zhang, F.; Fu, Z.; Yuan, Y.; Zhang, D. Toxic Effects of Per- and Polyfluoroalkyl Substances on Sperm: Epidemiological and Experimental Evidence. Front. Endocrinol. 2023, 14, 1114463. [Google Scholar] [CrossRef]
- Du, G.; Hu, J.; Huang, Z.; Yu, M.; Lu, C.; Wang, X.; Wu, D. Neonatal and Juvenile Exposure to Perfluorooctanoate (PFOA) and Perfluorooctane Sulfonate (PFOS): Advance Puberty Onset and Kisspeptin System Disturbance in Female Rats. Ecotoxicol. Environ. Saf. 2019, 167, 412–421. [Google Scholar] [CrossRef]
- Yeo, S.-H.; Colledge, W.H. The Role of Kiss1 Neurons As Integrators of Endocrine, Metabolic, and Environmental Factors in the Hypothalamic–Pituitary–Gonadal Axis. Front. Endocrinol. 2018, 9, 188. [Google Scholar] [CrossRef]
- López-Doval, S.; Salgado, R.; Pereiro, N.; Moyano, R.; Lafuente, A. Perfluorooctane Sulfonate Effects on the Reproductive Axis in Adult Male Rats. Environ. Res. 2014, 134, 158–168. [Google Scholar] [CrossRef]
- López-Doval, S.; Salgado, R.; Fernández-Pérez, B.; Lafuente, A. Possible Role of Serotonin and Neuropeptide Y on the Disruption of the Reproductive Axis Activity by Perfluorooctane Sulfonate. Toxicol. Lett. 2015, 233, 138–147. [Google Scholar] [CrossRef]
- López-Doval, S.; Salgado, R.; Lafuente, A. The Expression of Several Reproductive Hormone Receptors Can Be Modified by Perfluorooctane Sulfonate (PFOS) in Adult Male Rats. Chemosphere 2016, 155, 488–497. [Google Scholar] [CrossRef]
- Austin, M.E.; Kasturi, B.S.; Barber, M.; Kannan, K.; MohanKumar, P.S.; MohanKumar, S.M.J. Neuroendocrine Effects of Perfluorooctane Sulfonate in Rats. Environ. Health Perspect. 2003, 111, 1485–1489. [Google Scholar] [CrossRef]
- Rama Raju, G.; Chavan, R.; Deenadayal, M.; Gunasheela, D.; Gutgutia, R.; Haripriya, G.; Govindarajan, M.; Patel, N.; Patki, A. Luteinizing Hormone and Follicle Stimulating Hormone Synergy: A Review of Role in Controlled Ovarian Hyper-Stimulation. J. Hum. Reprod. Sci. 2013, 6, 227. [Google Scholar] [CrossRef]
- Wang, X.; Bai, Y.; Tang, C.; Cao, X.; Chang, F.; Chen, L. Impact of Perfluorooctane Sulfonate on Reproductive Ability of Female Mice through Suppression of Estrogen Receptor α-Activated Kisspeptin Neurons. Toxicol. Sci. 2018, 165, 475–486. [Google Scholar] [CrossRef]
- Chaparro-Ortega, A.; Betancourt, M.; Rosas, P.; Vázquez-Cuevas, F.G.; Chavira, R.; Bonilla, E.; Casas, E.; Ducolomb, Y. Endocrine Disruptor Effect of Perfluorooctane Sulfonic Acid (PFOS) and Perfluorooctanoic Acid (PFOA) on Porcine Ovarian Cell Steroidogenesis. Toxicol. Vitr. 2018, 46, 86–93. [Google Scholar] [CrossRef]
- Walters, K.A.; Handelsman, D.J. Role of Androgens in the Ovary. Mol. Cell. Endocrinol. 2018, 465, 36–47. [Google Scholar] [CrossRef]
- Dangudubiyyam, S.V.; Mishra, J.S.; Kumar, S. Perfluorooctane Sulfonic Acid Modulates Expression of Placental Steroidogenesis-Associated Genes and Hormone Levels in Pregnant Rats. Reprod. Toxicol. 2023, 118, 108390. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, X.; Liu, S.; Zhao, C.; Miao, Y.; Jin, L.; Wang, D.; Zhou, L. Cyp17a1 Is Required for Female Sex Determination and Male Fertility by Regulating Sex Steroid Biosynthesis in Fish. Endocrinology 2021, 162, bqab205. [Google Scholar] [CrossRef]
- López-Arellano, P.; López-Arellano, K.; Luna, J.; Flores, D.; Jiménez-Salazar, J.; Gavia, G.; Teteltitla, M.; Rodríguez, J.J.; Domínguez, A.; Casas, E.; et al. Perfluorooctanoic Acid Disrupts Gap Junction Intercellular Communication and Induces Reactive Oxygen Species Formation and Apoptosis in Mouse Ovaries. Environ. Toxicol. 2019, 34, 92–98. [Google Scholar] [CrossRef]
- Zhang, P.; Qi, C.; Ma, Z.; Wang, Y.; Zhang, L.; Hou, X. Perfluorooctanoic Acid Exposure in Vivo Perturbs Mitochondrial Metabolic during Oocyte Maturation. Environ. Toxicol. 2022, 37, 2965–2976. [Google Scholar] [CrossRef]
- Domínguez, A.; Salazar, Z.; Arenas, E.; Betancourt, M.; Ducolomb, Y.; González-Márquez, H.; Casas, E.; Teteltitla, M.; Bonilla, E. Effect of Perfluorooctane Sulfonate on Viability, Maturation and Gap Junctional Intercellular Communication of Porcine Oocytes in Vitro. Toxicol. Vitr. 2016, 35, 93–99. [Google Scholar] [CrossRef]
- Grazul-Bilska, A.T.; Vonnahme, K.A.; Bilski, J.J.; Borowczyk, E.; Soni, D.; Mikkelson, B.; Johnson, M.L.; Reynolds, L.P.; Redmer, D.A.; Caton, J.S. Expression of Gap Junctional Connexin Proteins in Ovine Fetal Ovaries: Effects of Maternal Diet. Domest. Anim. Endocrinol. 2011, 41, 185–194. [Google Scholar] [CrossRef]
- Bellavia, A.; Zou, R.; Björvang, R.D.; Roos, K.; Sjunnesson, Y.; Hallberg, I.; Holte, J.; Pikki, A.; Lenters, V.; Portengen, L.; et al. Association between Chemical Mixtures and Female Fertility in Women Undergoing Assisted Reproduction in Sweden and Estonia. Environ. Res. 2023, 216, 114447. [Google Scholar] [CrossRef]
- Feng, X.; Wang, X.; Cao, X.; Xia, Y.; Zhou, R.; Chen, L. Chronic Exposure of Female Mice to an Environmental Level of Perfluorooctane Sulfonate Suppresses Estrogen Synthesis Through Reduced Histone H3K14 Acetylation of the StAR Promoter Leading to Deficits in Follicular Development and Ovulation. Toxicol. Sci. 2015, 148, 368–379. [Google Scholar] [CrossRef]
- Yuan, Y.; Ding, X.; Cheng, Y.; Kang, H.; Luo, T.; Zhang, X.; Kuang, H.; Chen, Y.; Zeng, X.; Zhang, D. PFOA Evokes Extracellular Ca2+ Influx and Compromises Progesterone-Induced Response in Human Sperm. Chemosphere 2020, 241, 125074. [Google Scholar] [CrossRef]
- Pan, Y.; Cui, Q.; Wang, J.; Sheng, N.; Jing, J.; Yao, B.; Dai, J. Profiles of Emerging and Legacy Per-/Polyfluoroalkyl Substances in Matched Serum and Semen Samples: New Implications for Human Semen Quality. Environ. Health Perspect. 2019, 127, 127005. [Google Scholar] [CrossRef]
- Louis, G.M.B.; Chen, Z.; Schisterman, E.F.; Kim, S.; Sweeney, A.M.; Sundaram, R.; Lynch, C.D.; Gore-Langton, R.E.; Barr, D.B. Perfluorochemicals and Human Semen Quality: The LIFE Study. Environ. Health Perspect. 2015, 123, 57–63. [Google Scholar] [CrossRef]
- Shan, L.; Chai, Y.; Gao, T.; Li, K.; Yu, J.; Liang, F.; Ni, Y.; Sun, P. Perfluorooctane Sulfonate and Perfluorooctanoic Acid Inhibit Progesterone-Responsive Capacitation through cAMP/PKA Signaling Pathway and Induce DNA Damage in Human Sperm. Environ. Toxicol. Pharmacol. 2023, 100, 104165. [Google Scholar] [CrossRef]
- Zhao, B.; Li, L.; Liu, J.; Li, H.; Zhang, C.; Han, P.; Zhang, Y.; Yuan, X.; Ge, R.S.; Chu, Y. Exposure to Perfluorooctane Sulfonate In Utero Reduces Testosterone Production in Rat Fetal Leydig Cells. PLoS ONE 2014, 9, e78888. [Google Scholar] [CrossRef]
- Hagenäs, L.; Ritzén, E.M.; Plöen, L.; Hansson, V.; French, F.S.; Nayfeh, S.N. Sertoli Cell Origin of Testicular Androgen-Binding Protein (ABP). Mol. Cell. Endocrinol. 1975, 2, 339–350. [Google Scholar] [CrossRef]
- Labrie, F.; Luu-The, V.; Lin, S.; Simard, J.; Labrie, C.; El-Alfy, M.; Pelletier, G.; Belanger, A. Intracrinology: Role of the Family of 17 Beta-Hydroxysteroid Dehydrogenases in Human Physiology and Disease. J. Mol. Endocrinol. 2000, 25, 1–16. [Google Scholar] [CrossRef]
- Eggert, A.; Cisneros-Montalvo, S.; Anandan, S.; Musilli, S.; Stukenborg, J.-B.; Adamsson, A.; Nurmio, M.; Toppari, J. The Effects of Perfluorooctanoic Acid (PFOA) on Fetal and Adult Rat Testis. Reprod. Toxicol. 2019, 90, 68–76. [Google Scholar] [CrossRef]
- Mao, B.; Li, C.; Wen, Z.; Li, H.; Wang, Y.; Chen, L.; Lian, Q.; Ge, R.-S. Short-Term Perfluorooctane Sulfonate Exposure Impairs Leydig Cell Regeneration in the Adult Rat Testis via Targeting Hedgehog Signaling. Ecotoxicol. Environ. Saf. 2021, 214, 112121. [Google Scholar] [CrossRef]
- Qiu, L.; Wang, H.; Dong, T.; Huang, J.; Li, T.; Ren, H.; Wang, X.; Qu, J.; Wang, S. Perfluorooctane Sulfonate (PFOS) Disrupts Testosterone Biosynthesis via CREB/CRTC2/StAR Signaling Pathway in Leydig Cells. Toxicology 2021, 449, 152663. [Google Scholar] [CrossRef]
- Joensen, U.N.; Bossi, R.; Leffers, H.; Jensen, A.A.; Skakkebæk, N.E.; Jørgensen, N. Do Perfluoroalkyl Compounds Impair Human Semen Quality? Env. Health Perspect. 2009, 117, 923–927. [Google Scholar] [CrossRef]
- Esteves, S.C.; Verza, S.; Sharma, R.K.; Gosálvez, J.; Agarwal, A. Role and Significance of Sperm Function in Men with Unexplained Infertility. In Unexplained Infertility; Schattman, G.L., Esteves, S.C., Agarwal, A., Eds.; Springer: New York, NY, USA, 2015; pp. 91–119. [Google Scholar] [CrossRef]
- Rijsdijk, M. A Comparison of Three Spermatozoa Selection Techniques for Intracytoplasmic Sperm Injection (ICSI) using Swim-up, Cumulus Oophorus Model and PICSI® Dish. Doctoral Dissertation, Stellenbosch University, Stellenbosch, South Africa, 2015. [Google Scholar]
- Hirohashi, N.; Yanagimachi, R. Sperm Acrosome Reaction: Its Site and Role in Fertilization†. Biol. Reprod. 2018, 99, 127–133. [Google Scholar] [CrossRef]
- Brucker, C. The Human Sperm Acrosome Reaction: Physiology and Regulatory Mechanisms. An Update. Hum. Reprod. Update 1995, 1, 51–62. [Google Scholar] [CrossRef]
- Di Nisio, A.; Rocca, M.S.; Sabovic, I.; De Rocco Ponce, M.; Corsini, C.; Guidolin, D.; Zanon, C.; Acquasaliente, L.; Carosso, A.R.; De Toni, L.; et al. Perfluorooctanoic Acid Alters Progesterone Activity in Human Endometrial Cells and Induces Reproductive Alterations in Young Women. Chemosphere 2020, 242, 125208. [Google Scholar] [CrossRef]
- Cohen, N.J.; Yao, M.; Midya, V.; India-Aldana, S.; Mouzica, T.; Andra, S.S.; Narasimhan, S.; Meher, A.K.; Arora, M.; Chan, J.K.Y.; et al. Exposure to Perfluoroalkyl Substances and Women’s Fertility Outcomes in a Singaporean Population-Based Preconception Cohort. Sci. Total Environ. 2023, 873, 162267. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, W.; Wu, S.; Liang, F.; Li, Y.; Zhang, J.; Cui, L.; Feng, Y.; Wang, Y. Perfluoroalkyl Substances Exposure and Risk of Polycystic Ovarian Syndrome Related Infertility in Chinese Women. Environ. Pollut. 2019, 247, 824–831. [Google Scholar] [CrossRef]
- Vagi, S.J.; Azziz-Baumgartner, E.; Sjödin, A.; Calafat, A.M.; Dumesic, D.; Gonzalez, L.; Kato, K.; Silva, M.J.; Ye, X.; Azziz, R. Exploring the Potential Association between Brominated Diphenyl Ethers, Polychlorinated Biphenyls, Organochlorine Pesticides, Perfluorinated Compounds, Phthalates, and Bisphenol a in Polycystic Ovary Syndrome: A Case–Control Study. BMC Endocr. Disord. 2014, 14, 86. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, R.; Jin, F.; Lou, H.; Mao, Y.; Zhu, W.; Zhou, W.; Zhang, P.; Zhang, J. Perfluoroalkyl Substances and Endometriosis-Related Infertility in Chinese Women. Environ. Int. 2017, 102, 207–212. [Google Scholar] [CrossRef]
- Hallberg, I.; Kjellgren, J.; Persson, S.; Örn, S.; Sjunnesson, Y. Perfluorononanoic Acid (PFNA) Alters Lipid Accumulation in Bovine Blastocysts after Oocyte Exposure during in Vitro Maturation. Reprod. Toxicol. 2019, 84, 1–8. [Google Scholar] [CrossRef]
- Hallberg, I.; Persson, S.; Olovsson, M.; Sirard, M.-A.; Damdimopoulou, P.; Rüegg, J.; Sjunnesson, Y.C.B. Perfluorooctane Sulfonate (PFOS) Exposure of Bovine Oocytes Affects Early Embryonic Development at Human-Relevant Levels in an in Vitro Model. Toxicology 2021, 464, 153028. [Google Scholar] [CrossRef] [PubMed]
- Hallberg, I.; Persson, S.; Olovsson, M.; Moberg, M.; Ranefall, P.; Laskowski, D.; Damdimopoulou, P.; Sirard, M.-A.; Rüegg, J.; Sjunnesson, Y.C.B. Bovine Oocyte Exposure to Perfluorohexane Sulfonate (PFHxS) Induces Phenotypic, Transcriptomic, and DNA Methylation Changes in Resulting Embryos in Vitro. Reprod. Toxicol. 2022, 109, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Gao, M.; Cao, T.; Wang, J.; Luo, M.; Liu, S.; Zeng, X.; Huang, J. PFOS and F–53B Disrupted Inner Cell Mass Development in Mouse Preimplantation Embryo. Chemosphere 2024, 349, 140948. [Google Scholar] [CrossRef] [PubMed]
- Sawai, K. Roles of Cell Differentiation Factors in Preimplantation Development of Domestic Animals. J. Reprod. Dev. 2021, 67, 161–165. [Google Scholar] [CrossRef]
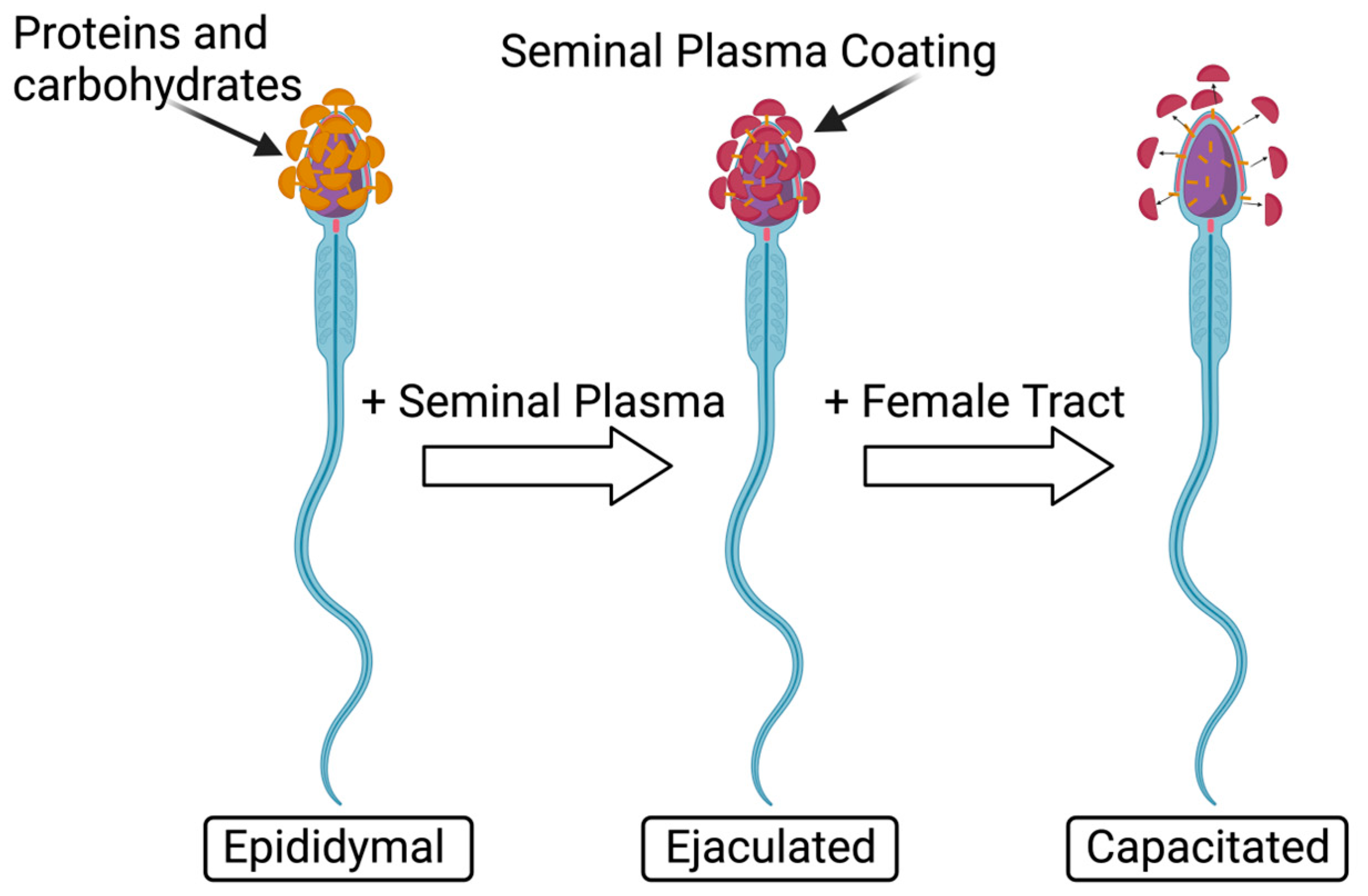
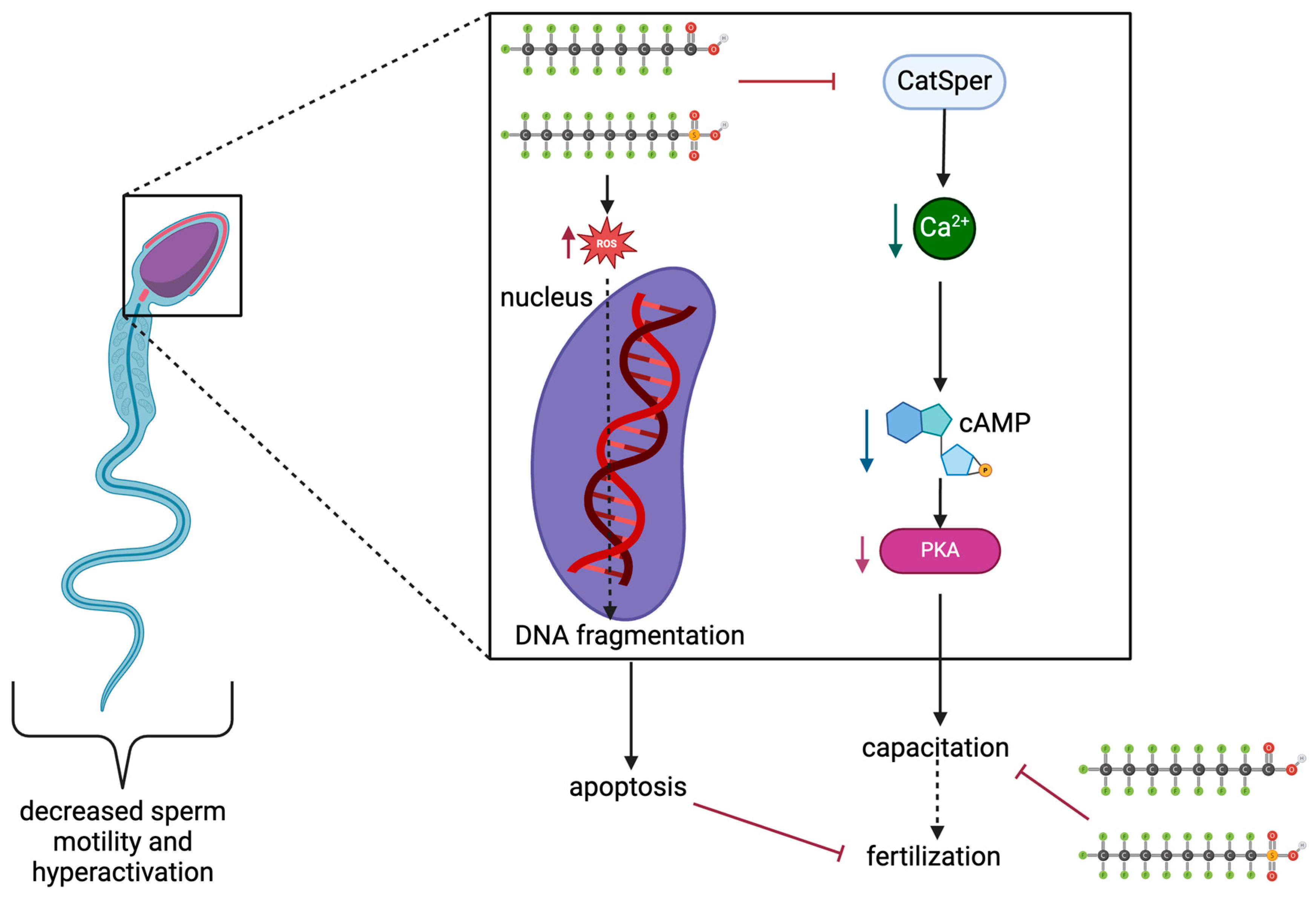

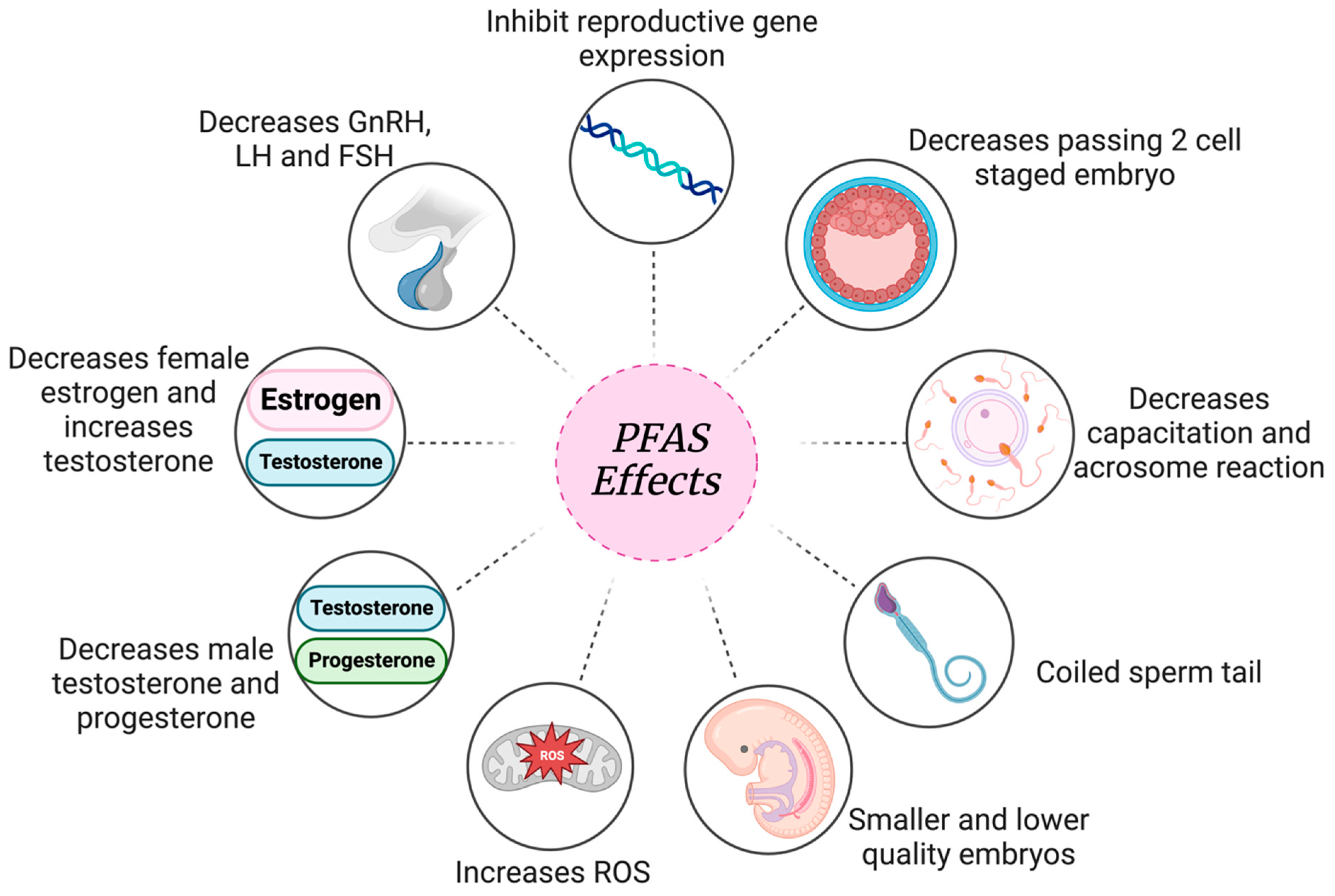
| Species | In Vivo/In Vitro | Cell Type | Endpoint | Treatment Dose (µM) | Effect Observed | Reference |
|---|---|---|---|---|---|---|
| Animal Studies | ||||||
| Sus scrofa | in vitro | Boar spermatozoa | Effects of PFOA and PFOS exposure on sperm motility and capacitation | 1000, 1500, 3000 μM for PFOS and 500, 1000, 1500, 2000, 2500 μM for PFOA | - 0 h PFOS: 20% in mortality - 4 h PFOS: 33% mortality - 1000 μM: significant decrease in capacitation - 1000 PFOS: 64% mortality - 3000 PFOS: 97% mortality - 1500 PFOA: 36% mortality - 2500 PFOA: 76.5% mortality - Capacitation levels of 64% significantly decreased to 36% for PFOS and 46% for PFOA | [18] |
| Test mechanism of capacitation inhibition after exposure to PFOS and PFOA | 1000, 1500, 3000 μM for PFOS and 500, 1000, 1500, 2000, 2500 μM for PFOA | - PFOS: significant increase in [Ca2+] at 0 h, slightly decreased between 1 and 2 h and significantly increased again at 3 h and 4 h - PFOA: increased [Ca2+] at 3 h | ||||
| Compare [Ca2+] levels with addition of ionophore and PFOA/PFOS exposure | 1000, 1500, 3000 μM for PFOS and 500, 1000, 1500, 2000, 2500 μM for PFOA | - Spermatozoa exposed to PFOA and PFOS did not respond to A23187 addition | ||||
| Pinpoint cause of increased calcium by looking at membrane potential fluctuation | 274 μM PFOS and 950 μM PFOA | - PFOS-treated samples did not repolarize after KCl and slightly responded to valinomycin - Impacted membrane function by 49% - PFOA samples had no response to valinomycin or KCl. Impacted membrane function by 47% | ||||
| Presence of cholesterol in PFAS exposed samples | 274 μM PFOS and 950 μM PFOA | - Significant decline in cholesterol in capacitated spermatozoa compared to non-capacitated - PFAS exposure caused similar results to non-capacitated spermatozoa | ||||
| Impact of ICC50 and LC50 of PFOA/PFOS on capacitation | ICC50 of PFOA: 1458 μM ICC50 of PFOS: 274 μM LC50 of PFOA: 1894 μM (half is 950 μM) LC50 of PFOS: 460 μM | - ICC50 PFOA: did not decrease capacitation - ICC50 of PFOS: significantly decreased capacitated spermatozoa by 43% - Half of LC50 PFOA significantly decreased capacitated spermatozoa by 28% | ||||
| Human Studies | ||||||
| Homo sapiens | in vitro | Human spermatozoa | PFAS’ effects on capacitation through cAMP/PKA mechanism and DNA damage | 10, 100, and 200 μM PFOS 10, 100, and 200 μM PFOA 100 μM PFOS + 100 μM PFOA | - Significantly decreased sperm motility and hyperactivation - Sperm motility patterns changed - Decreased protein phosphotyrosine, Ca2+ levels, cAMP levels (except 10 μM PFOA), and PKA - Significant increases in ROS for 100 and 200 μM PFOS and PFOA. - DNA fragmentation increased significantly - Significant increase in damaged sperm nuclei in 100 μM and 200 μM PFOA and PFOS groups | [60] |
| Species | In Vivo/In Vitro | Cell Type | Endpoint | Treatment Dose | Effect Observed | Reference |
|---|---|---|---|---|---|---|
| Animal Studies | ||||||
| Mus musculus | in vitro | Human spermatozoa | PFOA exposure impacts progesterone response in sperm | 0.25, 2.5, 25 μg/mL of PFOA | - Disturbed seminiferous tubules, as well as decreased sperm number, motility, testosterone, and progesterone levels | [57] |
| PFOA’s effects on sperm penetration | 0.25, 2.5, 25 μg/mL PFOA | - 25 µg/mL: significant decrease in sperm reaching penetration - 0.25 µg/mL: no change - All PFOA groups had a significant decrease after 10 μM P4 was added | ||||
| PFOA’s effects on acrosome reaction | 0.25, 2.5, 25 μg/mL PFOA | - No change until P4 was used to initiate acrosome reaction. All three doses showed significant inhibition of acrosome reaction - 25 µg/mL: significantly increased ROS | ||||
| Sus scrofa | in vitro | Boar spermatozoa | Effect of PFOA and PFOS exposure on acrosome reaction | 100, 200, 300 μM PFOS 150, 550, and 950 μM PFOA | - iAR reduced by 1.5% by PFOS and 18% by PFOA - PFOS dropped from 25% to 14, 11, and 1.5% (100, 200, and 300 μM) - PFOA dropped from 27% to 24, 23, and 18% (150, 550, and 950 μM) | [18] |
| Human Studies | ||||||
| Homo sapiens | in vitro | Human spermatozoa | Investigating presence of AR after PFAS exposure | 10, 100, and 200 μM PFOS 10, 100, and 200 μM PFOA 100 μM PFOS + 100 μM PFOA | - AR significantly decreased in the PFOS-exposed groups from 8.46% to 8.97, 7.68, and 6.44% (10, 100, and 300 μM) - For PFOA, significant decrease to 2.14 and 4.23 for 100 and 200 μM - Combined group had biggest effect to 5.32 | [60] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lockington, C.; Favetta, L.A. How Per- and Poly-Fluoroalkyl Substances Affect Gamete Viability and Fertilization Capability: Insights from the Literature. J. Xenobiot. 2024, 14, 651-678. https://doi.org/10.3390/jox14020038
Lockington C, Favetta LA. How Per- and Poly-Fluoroalkyl Substances Affect Gamete Viability and Fertilization Capability: Insights from the Literature. Journal of Xenobiotics. 2024; 14(2):651-678. https://doi.org/10.3390/jox14020038
Chicago/Turabian StyleLockington, Cielle, and Laura A. Favetta. 2024. "How Per- and Poly-Fluoroalkyl Substances Affect Gamete Viability and Fertilization Capability: Insights from the Literature" Journal of Xenobiotics 14, no. 2: 651-678. https://doi.org/10.3390/jox14020038
APA StyleLockington, C., & Favetta, L. A. (2024). How Per- and Poly-Fluoroalkyl Substances Affect Gamete Viability and Fertilization Capability: Insights from the Literature. Journal of Xenobiotics, 14(2), 651-678. https://doi.org/10.3390/jox14020038