
Dietary Behavior of Drosophila melanogaster Fed with Genetically-Modified Corn or Roundup®



Abstract
1. Introduction
Specific Hypotheses
2. Experimental Section
2.1. Fly Maintenance
2.2. Assay 1: Food Preference—Medium with GMO Corn
2.3. Assay 2: Food Preference—Sucrose with Roundup
2.4. Assay 3: Sucrose Consumption after Roundup Exposure
3. Results
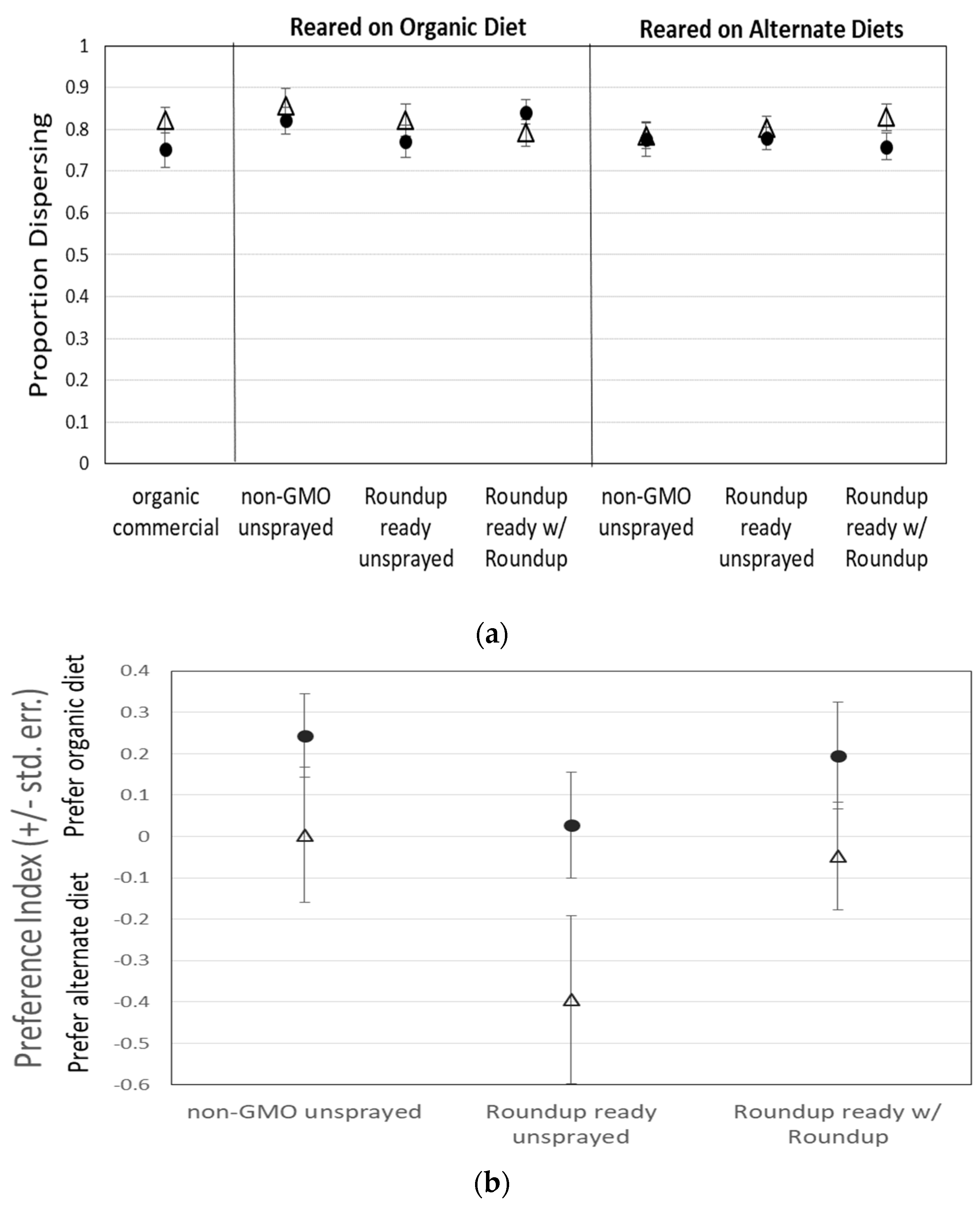
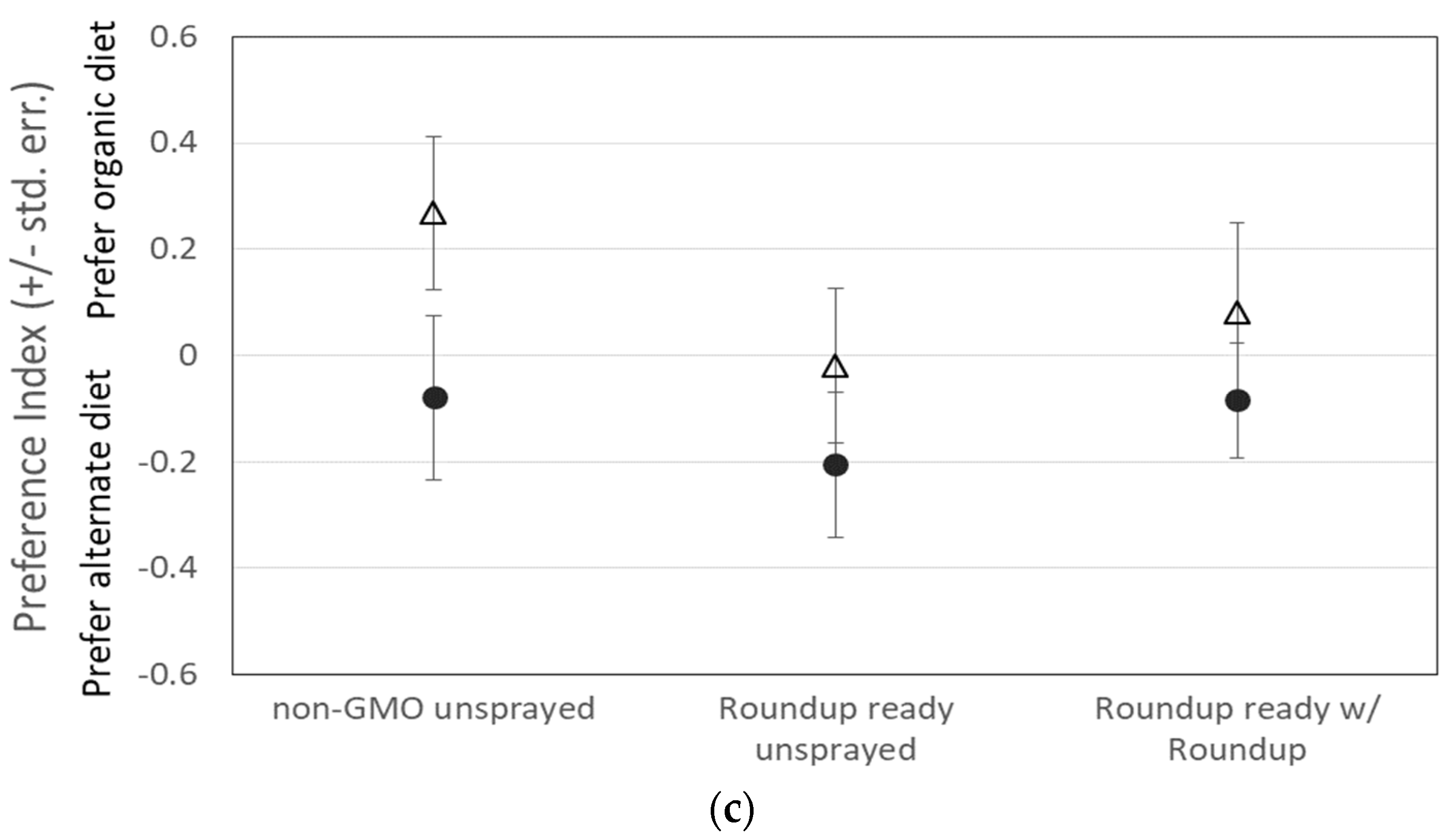
3.1. Assay 1: Food Preference—Medium with GMO Corn
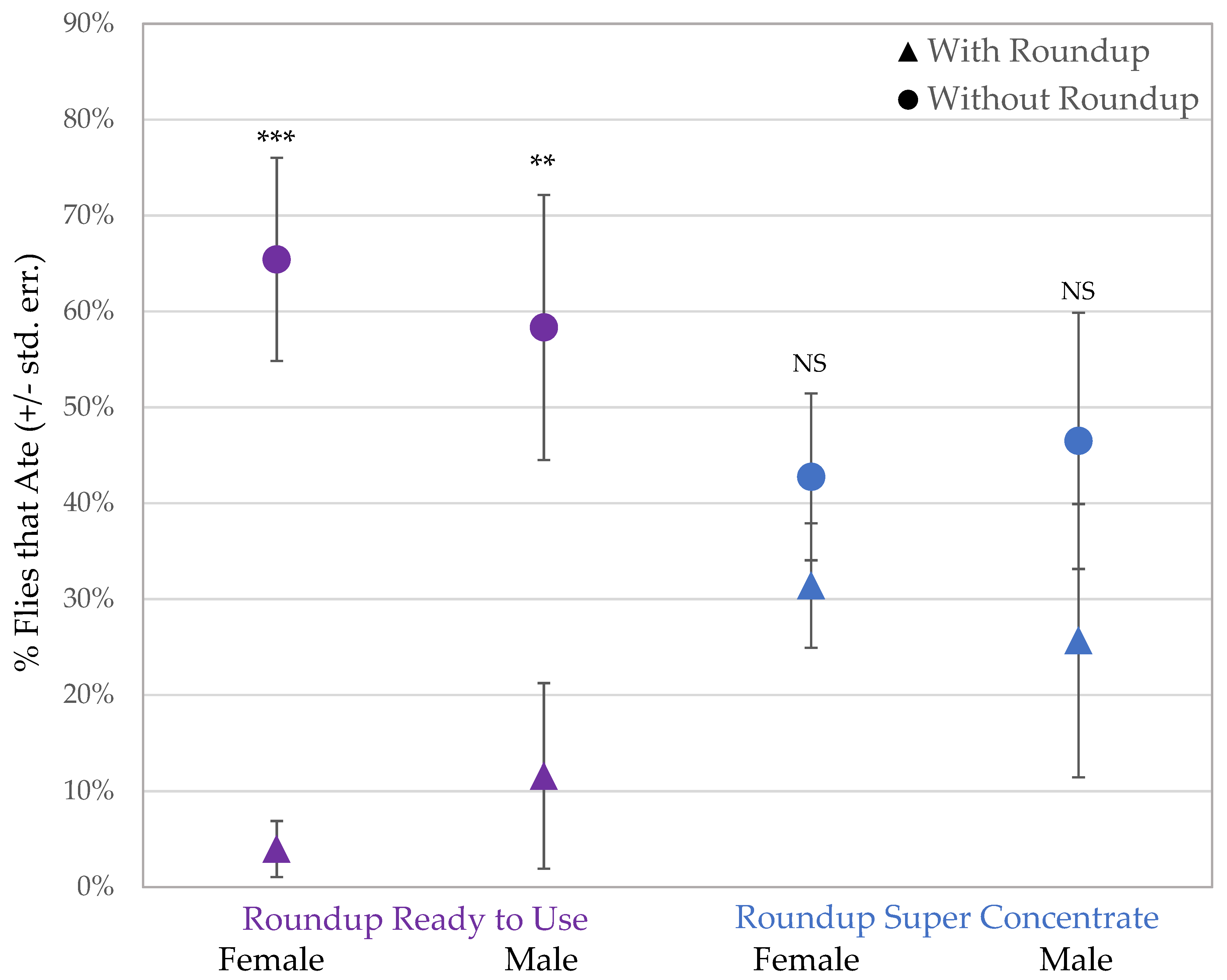
3.2. Assay 2: Consumption of Sucrose with Roundup
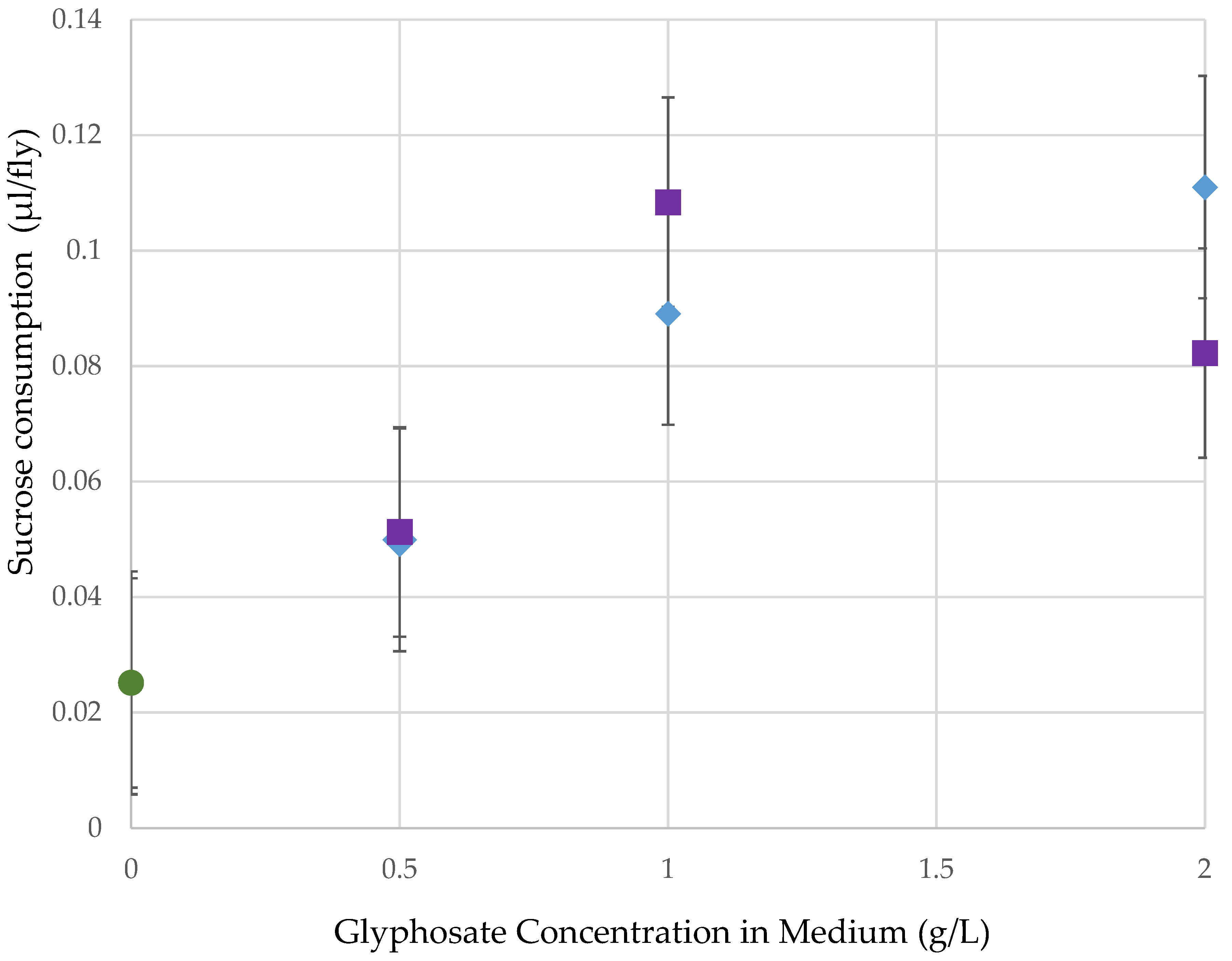
3.3. Assay 3: Sucrose Consumption after Roundup Exposure
4. Discussion
Future Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- USDA. Available online: https://www.ams.usda.gov/grades-standards/organic-standards (accessed on 1 March 2019).
- Brantsaeter, A.L.; Ydersbond, T.A.; Hoppin, J.A.; Haugen, M.; Meltzer, H.M. Organic Food in the Diet: Exposure and Health Implications. Annu. Rev. Public Health 2017, 38, 295–313. [Google Scholar] [CrossRef]
- Bøhn, T.; Cuhra, M.; Traavik, T.; Sanden, M.; Fagan, J.; Primicerio, R. Compositional differences in soybeans on the market: Glyphosate accumulates in Roundup Ready GM soybeans. Food Chem. 2014, 153, 207–215. [Google Scholar] [CrossRef]
- Barański, M.; Rempelos, L.; Iversen, P.O.; Leifert, C. Effects of organic food consumption on human health; the jury is still out! Food Nutr. Res. 2017, 61, 1287333. [Google Scholar] [CrossRef]
- Crinnion, W.J. Organic foods contain higher levels of certain nutrients, lower levels of pesticides, and may provide health benefits for the consumer. Altern. Med. Rev. 2010, 15, 4–12. [Google Scholar]
- Tsui, M.T.; Chu, L.M. Aquatic toxicity of glyphosate-based formulations: Comparison between different organisms and the effects of environmental factors. Chemosphere 2003, 52, 1189–1197. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, D.; Yang, S. Effect of organic and conventional rearing system on the mineral content of pork. Meat Sci. 2016, 118, 103–107. [Google Scholar] [CrossRef]
- Brandt, K.; Mølgaard, J.P. Organic agriculture: Does it enhance or reduce the nutritional value of plant foods? J. Sci. Food Agric. 2001, 81, 924–931. [Google Scholar] [CrossRef]
- Elmore, R.W.; Roeth, F.W.; Nelson, L.A.; Shapiro, C.A.; Klein, R.N.; Knezevic, S.Z.; Martin, A. Glyphosate-resistant soybean cultivar yields compared with sister lines. Agron. J. 2001, 93, 408–412. [Google Scholar] [CrossRef]
- Rótolo, G.C.; Francis, C.; Craviotto, R.M.; Ulgiati, S. Environmental assessment of maize production alternatives: Traditional, intensive and GMO-based cropping patterns. Ecol. Indic. 2015, 57, 48–60. [Google Scholar] [CrossRef]
- Benbrook, C.M. Impacts of genetically engineered crops on pesticide use in the US—The first sixteen years. Environ. Sci. Eur. 2012, 24, 24. [Google Scholar] [CrossRef]
- Iwase, H.; Ekser, B.; Hara, H.; Phelps, C.; Ayares, D.; Cooper, D.K.; Ezzelarab, M.B. Regulation of human platelet aggregation by genetically modified pig endothelial cells and thrombin inhibition. Xenotransplantation 2014, 21, 72–83. [Google Scholar] [CrossRef]
- Hasković, E.; Pekić, M.; Fočak, M.; Suljević, D.; Mešalić, L. Effects of Glyphosate on Enzyme Activity and Serum Glucose in Rats Rattus norvegicus. Acta Vet. 2016, 66, 214–221. [Google Scholar] [CrossRef]
- Mesnage, R.; Arno, M.; Séralini, G.E.; Antoniou, M.N. Transcriptome and metabolome analysis of liver and kidneys of rats chronically fed NK603 Roundup-tolerant genetically modified maize. Environ. Sci. Eur. 2017, 29, 6. [Google Scholar] [CrossRef]
- Schnell, J.; Steele, M.; Bean, J.; Neuspiel, M.; Girard, C.; Dormann, N.; Pearson, C.; Savoie, A.; Bourbonnière, L.; Macdonald, P. A comparative analysis of insertional effects in genetically engineered plants: Considerations for pre-market assessments. Transgenic Res. 2015, 24, 1–17. [Google Scholar] [CrossRef]
- Wehtje, G.; Altland, J.E.; Gilliam, C.H. Interaction of glyphosate and diquat in ready-to-use weed control products. Weed Technol. 2008, 22, 472–476. [Google Scholar] [CrossRef]
- Amrhein, N.; Deus, B.; Gehrke, P.; Steinrücken, H.C. The site of the inhibition of the shikimate pathway by glyphosate: II. Interference of glyphosate with chorismate formation in vivo and in vitro. Plant Physiol. 1980, 66, 830–834. [Google Scholar] [CrossRef] [PubMed]
- Dai, P.; Hu, P.; Tang, J.; Li, Y.; Li, C. Effect of glyphosate on reproductive organs in male rat. Acta Histochem. 2016, 118, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Dallegrave, E.; Mantese, F.D.; Oliveira, R.T.; Andrade, A.J.; Dalsenter, P.R.; Langeloh, A. Pre-and postnatal toxicity of the commercial glyphosate formulation in Wistar rats. Arch. Toxicol. 2007, 81, 665–673. [Google Scholar] [CrossRef]
- Gasnier, C.; Dumont, C.; Benachour, N.; Clair, E.; Chagnon, M.C.; Séralini, G.E. Glyphosate-based herbicides are toxic and endocrine disruptors in human cell lines. Toxicology 2009, 262, 184–191. [Google Scholar] [CrossRef]
- Aguiar, L.M.; Figueira, F.H.; Gottschalk, M.S.; Rosa, C.E. Glyphosate-based herbicide exposure causes antioxidant defense responses in the fruit fly Drosophila melanogaster. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2016, 185–186, 94–101. [Google Scholar] [CrossRef]
- Larsen, A.E.; Gaines, S.D.; Deschênes, O. Agricultural pesticide use and adverse birth outcomes in the San Joaquin Valley of California. Nat. Commun. 2017, 8, 302. [Google Scholar] [CrossRef] [PubMed]
- Milesi, M.M.; Lorenz, V.; Pacini, G.; Repetti, M.R.; Demonte, L.D.; Varayoud, J.; Luque, E.H. Perinatal exposure to a glyphosate-based herbicide impairs female reproductive outcomes and induces second-generation adverse effects in Wistar rats. Arch. Toxicol. 2018, 92, 2629–2643. [Google Scholar] [CrossRef] [PubMed]
- Muller, K.; Herrera, K.; Talyn, B.; Melchiorre, E. Toxicological effects of Roundup® on Drosophila melanogaster reproduction. Toxics 2021, 9, 161. [Google Scholar] [CrossRef]
- Aitbali, Y.; Ba-M'hamed, S.; Elhidar, N.; Nafis, A.; Soraa, N.; Bennis, M. Glyphosate based-herbicide exposure affects gut microbiota, anxiety and depression-like behaviors in mice. Neurotoxicol. Teratol. 2018, 67, 44–49. [Google Scholar] [CrossRef]
- Hietanen, E.; Linnainmaa, K.; Vainio, H. Effects of phenoxyherbicides and glyphosate on the hepatic and intestinal biotransformation activities in the rat. Basic Clin. Pharmacol. Toxicol. 1983, 53, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Shehata, A.A.; Kühnert, M.; Haufe, S.; Krüger, M. Neutralization of the antimicrobial effect of glyphosate by humic acid in vitro. Chemosphere 2014, 104, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Jayasumana, C.; Gunatilake, S.; Senanayake, P. Glyphosate, hard water and nephrotoxic metals: Are they the culprits behind the epidemic of chronic kidney disease of unknown etiology in Sri Lanka? Int. J. Environ. Res. Public Health 2014, 11, 2125–2147. [Google Scholar] [CrossRef]
- Mesnage, R.; Arno, M.; Costanzo, M.; Malatesta, M.; Séralini, G.E.; Antoniou, M.N. Transcriptome profile analysis reflects rat liver and kidney damage following chronic ultra-low dose Roundup exposure. Environ. Health 2015, 14, 70. [Google Scholar] [CrossRef]
- Roy, N.M.; Carneiro, B.; Ochs, J. Glyphosate induces neurotoxicity in zebrafish. Environ. Toxicol. Pharmacol. 2016, 42, 45–54. [Google Scholar] [CrossRef]
- Shaw, W. Elevated urinary glyphosate and clostridia metabolites with altered dopamine metabolism in triplets with autistic spectrum disorder or suspected seizure disorder: A case study. Integr. Med. A Clin. J. 2017, 16, 50. [Google Scholar]
- Woźniak, E.; Sicińska, P.; Muyh7ichałowicz, J.; Woźniak, K.; Reszka, E.; Huras, B.; Zakrzewski, J.; Bukowska, B. The mechanism of DNA damage induced by Roundup 360 PLUS, glyphosate and AMPA in human peripheral blood mononuclear cells-genotoxic risk assessment. Food Chem. Toxicol. 2018, 120, 510–522. [Google Scholar] [CrossRef]
- Fritschi, L.; McLaughlin, J.; Sergi, C.M.; Calaf, G.M.; Le Curieux, F.; Forastiere, F.; Kromhout, H.; Egeghy, P.; Jahnke, G.D.; Jameson, C.W.; et al. Carcinogenicity of tetrachlorvinphos, parathion, malathion, diazinon, and glyphosate. Red 2015, 114, 70134–70138. [Google Scholar]
- Krüger, M.; Schrödl, W.; Pedersen, I. Detection of Glyphosate in Malformed Piglets. J. Environ. Anal. Toxicol. 2014, 4, 2161-0525. [Google Scholar] [CrossRef]
- Malhotra, R.C.; Ghia, D.K.; Cordato, D.J.; Beran, R.G. Glyphosate–surfactant herbicide-induced reversible encephalopathy. J. Clin. Neurosci. 2010, 17, 1472–1473. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.G.; Telles, L.F.; Hess, R.A.; Mahecha, G.A.; Oliveira, C.A. Effects of the herbicide Roundup on the epididymal region of drakes Anas platyrhynchos. Reprod. Toxicol. 2007, 23, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Murphy, D.; Rowlands, H. Glyphosate: Unsafe on Any Plate. Food Democracy Now; 14 November 2016. Available online: https://s3.amazonaws.com/media.fooddemocracynow.org/images/FDN_Glyphosate_FoodTesting_Report_p2016.pdf (accessed on 5 December 2021).
- Morjan, W.E.; Pedigo, L.P.; Lewis, L.C. Fungicidal effects of glyphosate and glyphosate formulations on four species of entomopathogenic fungi. Environ. Entomol. 2002, 31, 1206–1212. [Google Scholar] [CrossRef]
- Mesnage, R.; Bernay, B.; Séralini, G.E. Ethoxylated adjuvants of glyphosate-based herbicides are active principles of human cell toxicity. Toxicology 2013, 313, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Defarge, N.; De Vendômois, J.S.; Séralini, G.E. Toxicity of formulants and heavy metals in glyphosate-based herbicides and other pesticides. Toxicol. Rep. 2018, 5, 156–163. [Google Scholar] [CrossRef]
- Székács, A.; Darvas, B. Forty years with glyphosate. Herbic.-Prop. Synth. Control Weeds 2012, 14, 247–284. [Google Scholar]
- Bridi, D.; Afltenhofen, S.; Gonzalez, J.B.; Reolon, G.K.; Bonan, C.D. Glyphosate and Roundup® alter morphology and behavior in zebrafish. Toxicology 2017, 392, 32–39. [Google Scholar] [CrossRef]
- Brovini, E.M.; Cardoso, S.J.; Quadra, G.R.; Vilas-Boas, J.A.; Paranaíba, J.R.; de Oliveira Pereira, R.; Mendonça, R.F. Glyphosate concentrations in global freshwaters: Are aquatic organisms at risk? Environ. Sci. Pollut. Res. 2021, 28, 60635–60648. [Google Scholar] [CrossRef]
- Mesnage, R.; Moesch, C.; Grand, R.; Lauthier, G.; Vendômois, J.; Gress, S.; Séralini, G. Glyphosate exposure in a farmer’s family. J. Environ. Prot. 2012, 9, 1001–1003. [Google Scholar] [CrossRef]
- Parvez, S.; Gerona, R.R.; Proctor, C.; Friesen, M.; Ashby, J.L.; Reiter, J.L.; Lui, Z.; Winchester, P.D. Glyphosate exposure in pregnancy and shortened gestational length: A prospective Indiana birth cohort study. Environ. Health 2018, 17, 23. [Google Scholar] [CrossRef] [PubMed]
- Hardell, L.; Eriksson, M. A case–control study of non-Hodgkin lymphoma and exposure to pesticides. Cancer 1999, 85, 1353–1360. [Google Scholar] [CrossRef]
- Schinasi, L.; Leon, M. Non-Hodgkin Lymphoma and Occupational Exposure to Agricultural Pesticide Chemical Groups and Active Ingredients: A Systematic Review and Meta-Analysis. Int. J. Environ. Res. Public Health 2014, 11, 4449–4527. [Google Scholar] [CrossRef]
- Krüger, M.; Schledorn, P.; Schrödl, W.; Hoppe, H.W.; Lutz, W.; Shehata, A.A. Detection of glyphosate residues in animals and humans. J. Environ. Anal. Toxicol. 2014, 4, 1–5. [Google Scholar]
- Fagan, J.; Bohlen, L.; Patton, S.; Klein, K. Organic diet intervention significantly reduces urinary glyphosate levels in U.S. children and adults. Environ. Res. 2020, 189, 109898. [Google Scholar] [CrossRef]
- Amrein, H.; Thorne, N. Gustatory perception and behavior in Drosophila melanogaster. Curr. Biol. 2005, 15, R673–R684. [Google Scholar] [CrossRef] [PubMed]
- Talyn, B.; Lemon, R.; Badoella, M.; Melchiorre, D.; Villalobos, M.; Elias, R.; Muller, K.; Santos, M.; Melchiorre, E. Roundup®, but not Roundup-Ready® corn, increases mortality of Drosophila melanogaster. Toxics 2019, 7, 38. [Google Scholar] [CrossRef]
- Ge, X.; Hannan, F.; Xie, Z.; Feng, C.; Tully, T.; Zhou, H.; Xie, Z.; Zhong, Y. Notch signaling in Drosophila long-term memory formation. Proc. Natl. Acad. Sci. USA 2004, 101, 10172–10176. [Google Scholar] [CrossRef]
- Suh, G.S.; de Leon, S.B.T.; Tanimoto, H.; Fiala, A.; Benzer, S.; Anderson, D.J. Light activation of an innate olfactory avoidance response in Drosophila. Curr. Biol. 2007, 17, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Toshima, N.; Hara, C.; Scholz, C.J.; Tanimura, T. Genetic variation in food choice behaviour of amino acid-deprived Drosophila. J. Insect Physiol. 2014, 69, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Russell, C.; Wessnitzer, J.; Young, J.M.; Armstrong, J.D.; Webb, B. Dietary salt levels affect salt preference and learning in larval Drosophila. PLoS ONE 2011, 6, e20100. [Google Scholar] [CrossRef] [PubMed]
- Tully, T.; Cambiazo, V.; Kruse, L. Memory through metamorphosis in normal and mutant Drosophila. J. Neurosci. 1994, 14, 68–74. [Google Scholar] [CrossRef]
- Wong, R.; Piper, M.D.; Blanc, E.; Partridge, L. Pitfalls of measuring feeding rate in the fruit fly Drosophila melanogaster. Nat. Methods 2008, 5, 214. [Google Scholar] [CrossRef]
- Diegelmann, S.; Jansen, A.; Jois, S.; Kastenholz, K.; Escarcena, L.V.; Strudthoff, N.; Scholz, H. The CApillary FEeder assay measures food intake in Drosophila melanogaster. J. Vis. Exp. JoVE 2017, 121, e55024. [Google Scholar] [CrossRef]
- Mandel, S.J.; Shoaf, M.L.; Braco, J.T.; Silver, W.L.; Johnson, E.C. Behavioral Aversion to AITC Requires Both Painless and dTRPA1 in Drosophila. Front. Neural Circuits 2018, 12, 45. [Google Scholar] [CrossRef]
- Magwere, T.; Chapman, T.; Partridge, L. Sex differences in the effect of dietary restriction on life span and mortality rates in female and male Drosophila melanogaster. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2004, 59, B3–B9. [Google Scholar] [CrossRef]
- De Vendômois, J.S.; Roullier, F.; Cellier, D.; Séralini, G.E. A comparison of the effects of three GM corn varieties on mammalian health. Int. J. Biol. Sci. 2009, 5, 706. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Okasha, E.F. Effect of genetically modified corn on the jejunal mucosa of adult male albino rat. Exp. Toxicol. Pathol. 2016, 68, 579–588. [Google Scholar] [CrossRef]
- Leaper, C.; Holloway, P.J. Adjuvants and glyphosate activity. Pest Manag. Sci. 2000, 56, 313–319. [Google Scholar] [CrossRef]
- Wehtje, G.; Gilliam, C.H.; Marble, S.C. Postemergence weed control with glyphosate plus flumioxazin combinations. Weed Technol. 2010, 24, 356–360. [Google Scholar] [CrossRef]
- Stamps, J.A.; Yang, L.H.; Morales, V.M.; Boundy-Mills, K.L. Drosophila regulate yeast density and increase yeast community similarity in a natural substrate. PLoS ONE 2012, 7, e42238. [Google Scholar] [CrossRef] [PubMed]
- Ringo, J.; Talyn, B.; Brannan, M. Effects of precocene and low protein diet on reproductive behavior in Drosophila melanogaster (Diptera: Drosophilidae). Ann. Entomol. Soc. Am. 2005, 98, 601–607. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Multivariate | Dispersal Index | Preference Index | |||||
|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | ||
| Flies reared on organic diet | Overall | 1.5732 | NS | 0.9438 | NS | 2.2213 | NS |
| Diet | 1.9016 | NS | 0.8211 | NS | 3.0717 | NS | |
| Sex | 5.1857 | 0.0078 | 1.0682 | NS | 8.8508 | 0.0039 | |
| Interaction | 0.6494 | NS | 1.1123 | NS | 0.1697 | NS | |
| Flies reared on treatment diet | Overall | 1.1721 | NS | 0.6488 | NS | 1.731 | NS |
| Diet | 1.0932 | NS | 0.0594 | NS | 2.2143 | NS | |
| Sex | 2.3102 | NS | 3.3639 | NS | 0.8941 | NS | |
| Interaction | 0.9749 | NS | 0.4277 | NS | 1.6823 | NS | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elias, R.; Talyn, B.; Melchiorre, E. Dietary Behavior of Drosophila melanogaster Fed with Genetically-Modified Corn or Roundup®. J. Xenobiot. 2021, 11, 215-227. https://doi.org/10.3390/jox11040014
Elias R, Talyn B, Melchiorre E. Dietary Behavior of Drosophila melanogaster Fed with Genetically-Modified Corn or Roundup®. Journal of Xenobiotics. 2021; 11(4):215-227. https://doi.org/10.3390/jox11040014
Chicago/Turabian StyleElias, Raquel, Becky Talyn, and Erik Melchiorre. 2021. "Dietary Behavior of Drosophila melanogaster Fed with Genetically-Modified Corn or Roundup®" Journal of Xenobiotics 11, no. 4: 215-227. https://doi.org/10.3390/jox11040014
APA StyleElias, R., Talyn, B., & Melchiorre, E. (2021). Dietary Behavior of Drosophila melanogaster Fed with Genetically-Modified Corn or Roundup®. Journal of Xenobiotics, 11(4), 215-227. https://doi.org/10.3390/jox11040014