
Lights on for Autism: Exploring Photobiomodulation as an Effective Therapeutic Option

{kind=link}
Abstract
1. Introduction
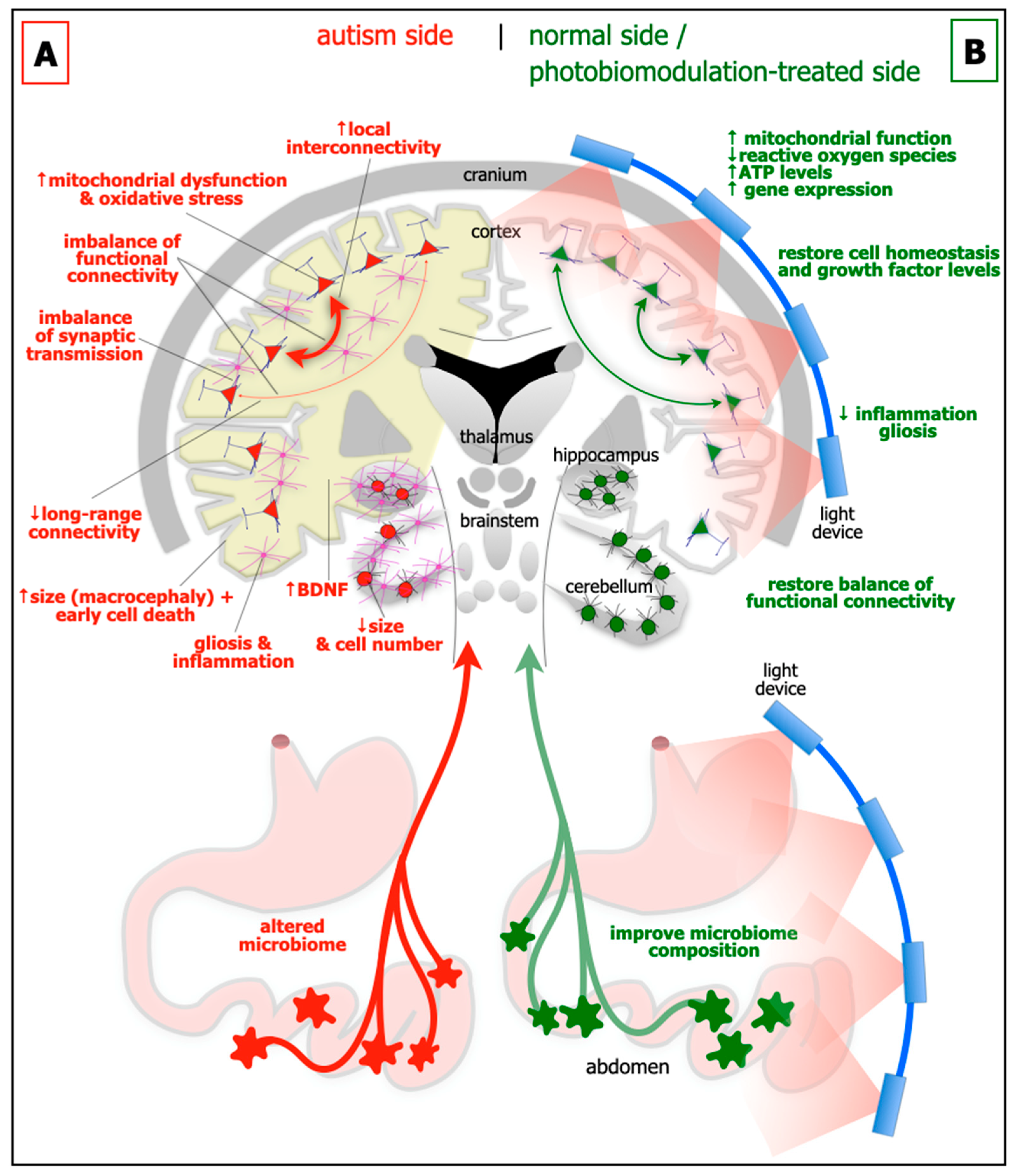
2. The Mechanisms
3. Current Treatments
4. Photobiomodulation: The Light
5. Effect of Photobiomodulation in Autism
6. A Working Hypothesis
- (1)
- improving the behaviour and abnormal neural circuitry in the brain; we suggest that photobiomodulation will induce a more balanced pattern of functional connectivity between different regions of the brain;
- (2)
- reducing cell death, mitochondrial dysfunction and oxidative stress, gliosis and inflammation in the brain; we propose that photobiomodulation will restore normal cell homeostasis;
- (3)
- altering the composition of the microbiome and thence brain neural circuitry and thus behaviour; we suggest that microbial activity will be restored towards “normal” levels and that this will lead to an improvement in brain function.
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fuentes, J.; Hervás, A.; Howlin, P. (ESCAP ASD Working Party) ESCAP Practice Guidance for Autism: A Summary of Evidence-Based Recommendations for Diagnosis and Treatment. Eur. Child. Adolesc. Psychiatry 2021, 30, 961–984. [Google Scholar] [CrossRef] [PubMed]
- Crawley, J.N. Translational Animal Models of Autism and Neurodevelopmental Disorders. Dialogues Clin. Neurosci. 2012, 14, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Won, H.; Mah, W.; Kim, E. Autism Spectrum Disorder Causes, Mechanisms, and Treatments: Focus on Neuronal Synapses. Front. Mol. Neurosci. 2013, 6, 19. [Google Scholar] [CrossRef]
- Mabunga, D.F.N.; Gonzales, E.L.T.; Kim, J.; Kim, K.C.; Shin, C.Y. Exploring the Validity of Valproic Acid Animal Model of Autism. Exp. Neurobiol. 2015, 24, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Chaliha, D.; Albrecht, M.; Vaccarezza, M.; Takechi, R.; Lam, V.; Al-Salami, H.; Mamo, J. A Systematic Review of the Valproic-Acid-Induced Rodent Model of Autism. Dev. Neurosci. 2020, 42, 12–48. [Google Scholar] [CrossRef]
- Hashem, S.; Nisar, S.; Bhat, A.A.; Yadav, S.K.; Azeem, M.W.; Bagga, P.; Fakhro, K.; Reddy, R.; Frenneaux, M.P.; Haris, M. Genetics of Structural and Functional Brain Changes in Autism Spectrum Disorder. Transl. Psychiatry 2020, 10, 229. [Google Scholar] [CrossRef]
- Garcia-Gutierrez, E.; Narbad, A.; Rodríguez, J.M. Autism Spectrum Disorder Associated With Gut Microbiota at Immune, Metabolomic, and Neuroactive Level. Front. Neurosci. 2020, 14, 578666. [Google Scholar] [CrossRef]
- Qi, Z.; Lyu, M.; Yang, L.; Yuan, H.; Cao, Y.; Zhai, L.; Dang, W.; Liu, J.; Yang, F.; Li, Y. A Novel and Reliable Rat Model of Autism. Front. Psychiatry 2021, 12, 549810. [Google Scholar] [CrossRef]
- Taleb, A.; Lin, W.; Xu, X.; Zhang, G.; Zhou, Q.-G.; Naveed, M.; Meng, F.; Fukunaga, K.; Han, F. Emerging Mechanisms of Valproic Acid-Induced Neurotoxic Events in Autism and Its Implications for Pharmacological Treatment. Biomed. Pharmacother. 2021, 137, 111322. [Google Scholar] [CrossRef]
- Hamblin, M.R. Shining Light on the Head: Photobiomodulation for Brain Disorders. BBA Clin. 2016, 6, 113–124. [Google Scholar] [CrossRef]
- Courchesne, E.; Campbell, K.; Solso, S. Brain Growth across the Life Span in Autism: Age-Specific Changes in Anatomical Pathology. Brain Res. 2011, 1380, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Alberts, I.; Li, X. The Apoptotic Perspective of Autism. Int. J. Dev. Neurosci. 2014, 36, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Courchesne, E. Abnormal Early Brain Development in Autism. Mol. Psychiatry 2002, 7 (Suppl. 2), S21–S23. [Google Scholar] [CrossRef]
- Sparks, B.F.; Friedman, S.D.; Shaw, D.W.; Aylward, E.H.; Echelard, D.; Artru, A.A.; Maravilla, K.R.; Giedd, J.N.; Munson, J.; Dawson, G.; et al. Brain Structural Abnormalities in Young Children with Autism Spectrum Disorder. Neurology 2002, 59, 184–192. [Google Scholar] [CrossRef]
- Falcone, C.; Mevises, N.-Y.; Hong, T.; Dufour, B.; Chen, X.; Noctor, S.C.; Martínez Cerdeño, V. Neuronal and Glial Cell Number Is Altered in a Cortical Layer-Specific Manner in Autism. Autism 2021, 25, 2238–2253. [Google Scholar] [CrossRef]
- Aylward, E.H.; Minshew, N.J.; Goldstein, G.; Honeycutt, N.A.; Augustine, A.M.; Yates, K.O.; Barta, P.E.; Pearlson, G.D. MRI Volumes of Amygdala and Hippocampus in Non-Mentally Retarded Autistic Adolescents and Adults. Neurology 1999, 53, 2145–2150. [Google Scholar] [CrossRef] [PubMed]
- Schumann, C.M.; Amaral, D.G. Stereological Analysis of Amygdala Neuron Number in Autism. J. Neurosci. 2006, 26, 7674–7679. [Google Scholar] [CrossRef] [PubMed]
- May, K.E.; Kana, R.K. Frontoparietal Network in Executive Functioning in Autism Spectrum Disorder. Autism Res. 2020, 13, 1762–1777. [Google Scholar] [CrossRef]
- Kim, D.; Lee, J.Y.; Jeong, B.C.; Ahn, J.; Kim, J.I.; Lee, E.S.; Kim, H.; Lee, H.J.; Han, C.E. Overconnectivity of the Right Heschl’s and Inferior Temporal Gyrus Correlates with Symptom Severity in Preschoolers with Autism Spectrum Disorder. Autism Res. 2021, 14, 2314–2329. [Google Scholar] [CrossRef]
- Herbert, M.R. Large Brains in Autism: The Challenge of Pervasive Abnormality. Neuroscientist 2005, 11, 417–440. [Google Scholar] [CrossRef]
- Purcell, A.E.; Jeon, O.H.; Zimmerman, A.W.; Blue, M.E.; Pevsner, J. Postmortem Brain Abnormalities of the Glutamate Neurotransmitter System in Autism. Neurology 2001, 57, 1618–1628. [Google Scholar] [CrossRef] [PubMed]
- Fatemi, S.H. Co-Occurrence of Neurodevelopmental Genes in Etiopathogenesis of Autism and Schizophrenia. Schizophr. Res. 2010, 118, 303–304. [Google Scholar] [CrossRef] [PubMed]
- Cook, E.H.J.; Courchesne, R.; Lord, C.; Cox, N.J.; Yan, S.; Lincoln, A.; Haas, R.; Courchesne, E.; Leventhal, B.L. Evidence of Linkage between the Serotonin Transporter and Autistic Disorder. Mol. Psychiatry 1997, 2, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Gzielo, K.; Nikiforuk, A. Astroglia in Autism Spectrum Disorder. IJMS 2021, 22, 11544. [Google Scholar] [CrossRef]
- Wen, Y.; Yao, Y. Autism Spectrum Disorders: The Mitochondria Connection. In Autism Spectrum Disorders; Exon Brisbane Australia: Brisbane City, Australia, 2021; ISBN 978-0-645-00178-5. [Google Scholar]
- Miyazaki, K.; Narita, N.; Sakuta, R.; Miyahara, T.; Naruse, H.; Okado, N.; Narita, M. Serum Neurotrophin Concentrations in Autism and Mental Retardation: A Pilot Study. Brain Dev. 2004, 26, 292–295. [Google Scholar] [CrossRef]
- Yeom, C.-W.; Park, Y.-J.; Choi, S.-W.; Bhang, S.-Y. Association of Peripheral BDNF Level with Cognition, Attention and Behavior in Preschool Children. Child. Adolesc. Psychiatry Ment. Health 2016, 10, 10. [Google Scholar] [CrossRef]
- Saghazadeh, A.; Rezaei, N. Brain-Derived Neurotrophic Factor Levels in Autism: A Systematic Review and Meta-Analysis. J. Autism Dev. Disord. 2017, 47, 1018–1029. [Google Scholar] [CrossRef]
- Tan, Q.; Orsso, C.E.; Deehan, E.C.; Kung, J.Y.; Tun, H.M.; Wine, E.; Madsen, K.L.; Zwaigenbaum, L.; Haqq, A.M. Probiotics, Prebiotics, Synbiotics, and Fecal Microbiota Transplantation in the Treatment of Behavioral Symptoms of Autism Spectrum Disorder: A Systematic Review. Autism Res. 2021, 14, 1820–1836. [Google Scholar] [CrossRef]
- Sharma, S.R.; Gonda, X.; Tarazi, F.I. Autism Spectrum Disorder: Classification, Diagnosis and Therapy. Pharmacol. Ther. 2018, 190, 91–104. [Google Scholar] [CrossRef]
- Genovese, A.; Butler, M.G. Clinical Assessment, Genetics, and Treatment Approaches in Autism Spectrum Disorder (ASD). IJMS 2020, 21, 4726. [Google Scholar] [CrossRef]
- Lai, M.-C.; Anagnostou, E.; Wiznitzer, M.; Allison, C.; Baron-Cohen, S. Evidence-Based Support for Autistic People across the Lifespan: Maximising Potential, Minimising Barriers, and Optimising the Person–Environment Fit. Lancet Neurol. 2020, 19, 434–451. [Google Scholar] [CrossRef]
- Lord, C.; Brugha, T.S.; Charman, T.; Cusack, J.; Dumas, G.; Frazier, T.; Jones, E.J.H.; Jones, R.M.; Pickles, A.; State, M.W.; et al. Autism Spectrum Disorder. Nat. Rev. Dis. Prim. 2020, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Shoaib, A.; Cepeda, M.S.; Murray, G.; Ochs-Ross, R. Autism: Comorbidities and Treatment Patterns in the Real World, a Retrospective Cohort Study Among Children, Adolescents and Adults Newly Diagnosed with Autism. J. Autism Dev. Disord. 2022, 52, 4311–4320. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.S.; Nasir, M.; Farhat, L.C.; Kook, M.; Artukoglu, B.B.; Bloch, M.H. Meta-Analysis: Pharmacologic Treatment of Restricted and Repetitive Behaviors in Autism Spectrum Disorders. J. Am. Acad. Child Adolesc. Psychiatry 2021, 60, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Mitrofanis, J. Run in the Light: Exploring Exercise and Photobiomodulation in Parkinson’s Disease; Morgan & Claypool Publishers: San Rafael, CA, USA, 2019; ISBN 978-1-64327-720-2. [Google Scholar]
- Hamblin, M.R. Could Photobiomodulation Treat Autism Spectrum Disorder? Photobiomodul. Photomed. Laser Surg. 2022, 40, 367–369. [Google Scholar] [CrossRef]
- Bicknell, B.; Liebert, A.; Johnstone, D.; Kiat, H. Photobiomodulation of the Microbiome: Implications for Metabolic and Inflammatory Diseases. Lasers Med. Sci. 2019, 34, 317–327. [Google Scholar] [CrossRef]
- Blivet, G.; Meunier, J.; Roman, F.J.; Touchon, J. Neuroprotective Effect of a New Photobiomodulation Technique against Aβ25–35 Peptide–Induced Toxicity in Mice: Novel Hypothesis for Therapeutic Approach of Alzheimer’s Disease Suggested. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2018, 4, 54–63. [Google Scholar] [CrossRef]
- Liebert, A.; Bicknell, B.; Johnstone, D.M.; Gordon, L.C.; Kiat, H.; Hamblin, M.R. “Photobiomics”: Can Light, Including Photobiomodulation, Alter the Microbiome? Photobiomodul. Photomed. Laser Surg. 2019, 37, 681–693. [Google Scholar] [CrossRef]
- Hamblin, M.R.; Liebert, A. Photobiomodulation Therapy Mechanisms beyond Cytochrome c Oxidase. Photobiomodul. Photomed. Laser Surg. 2022, 40, 75–77. [Google Scholar] [CrossRef]
- Vargas, E.; Barrett, D.W.; Saucedo, C.L.; Huang, L.-D.; Abraham, J.A.; Tanaka, H.; Haley, A.P.; Gonzalez-Lima, F. Beneficial Neurocognitive Effects of Transcranial Laser in Older Adults. Lasers Med. Sci. 2017, 32, 1153–1162. [Google Scholar] [CrossRef]
- Wang, X.; Tian, F.; Reddy, D.D.; Nalawade, S.S.; Barrett, D.W.; Gonzalez-Lima, F.; Liu, H. Up-Regulation of Cerebral Cytochrome-c-Oxidase and Hemodynamics by Transcranial Infrared Laser Stimulation: A Broadband near-Infrared Spectroscopy Study. J. Cereb. Blood Flow Metab. 2017, 37, 3789–3802. [Google Scholar] [CrossRef] [PubMed]
- Jahan, A.; Nazari, M.A.; Mahmoudi, J.; Salehpour, F.; Salimi, M.M. Transcranial Near-Infrared Photobiomodulation Could Modulate Brain Electrophysiological Features and Attentional Performance in Healthy Young Adults. Lasers Med. Sci. 2019, 34, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Zomorrodi, R.; Loheswaran, G.; Pushparaj, A.; Lim, L. Pulsed Near Infrared Transcranial and Intranasal Photobiomodulation Significantly Modulates Neural Oscillations: A Pilot Exploratory Study. Sci. Rep. 2019, 9, 6309. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.-C.; Fang, W.; Chang, Y.-C.; Chang, W.-D.; Wu, J.-H. Effect of Near-Infrared Pulsed Light on the Human Brain Using Electroencephalography. Evid. Based Complement. Altern. Med. 2021, 2021, 6693916. [Google Scholar] [CrossRef] [PubMed]
- Saucedo, C.L.; Courtois, E.C.; Wade, Z.S.; Kelley, M.N.; Kheradbin, N.; Barrett, D.W.; Gonzalez-Lima, F. Transcranial Laser Stimulation: Mitochondrial and Cerebrovascular Effects in Younger and Older Healthy Adults. Brain Stimul. 2021, 14, 440–449. [Google Scholar] [CrossRef]
- Konstantinović, L.M.; Jelić, M.B.; Jeremić, A.; Stevanović, V.B.; Milanović, S.D.; Filipović, S.R. Transcranial Application of Near-Infrared Low-Level Laser Can Modulate Cortical Excitability. Lasers Surg. Med. 2013, 45, 648–653. [Google Scholar] [CrossRef]
- Chow, R.T.; Armati, P.J. Photobiomodulation: Implications for Anesthesia and Pain Relief. Photomed. Laser Surg. 2016, 34, 599–609. [Google Scholar] [CrossRef]
- El Khoury, H.; Mitrofanis, J.; Henderson, L.A. Exploring the Effects of Near Infrared Light on Resting and Evoked Brain Activity in Humans Using Magnetic Resonance Imaging. Neuroscience 2019, 422, 161–171. [Google Scholar] [CrossRef]
- Fekete, Z.; Horváth, Á.C.; Zátonyi, A. Infrared Neuromodulation: A Neuroengineering Perspective. J. Neural Eng. 2020, 17, 051003. [Google Scholar] [CrossRef]
- Naeser, M.A.; Ho, M.D.; Martin, P.I.; Hamblin, M.R.; Koo, B.-B. Increased Functional Connectivity within Intrinsic Neural Networks in Chronic Stroke Following Treatment with Red/Near-Infrared Transcranial Photobiomodulation: Case Series with Improved Naming in Aphasia. Photobiomodul. Photomed. Laser Surg. 2020. [Google Scholar] [CrossRef]
- Mitrofanis, J.; Henderson, L.A. How and Why Does Photobiomodulation Change Brain Activity? Neural Regen. Res. 2020, 15, 2243–2244. [Google Scholar] [CrossRef] [PubMed]
- Chao, L.L. Effects of Home Photobiomodulation Treatments on Cognitive and Behavioral Function, Cerebral Perfusion, and Resting-State Functional Connectivity in Patients with Dementia: A Pilot Trial. Photobiomodul. Photomed. Laser Surg. 2019, 37, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Ceranoglu, T.A.; Cassano, P.; Hoskova, B.; Green, A.; Dallenbach, N.; DiSalvo, M.; Biederman, J.; Joshi, G. Transcranial Photobiomodulation in Adults with High-Functioning Autism Spectrum Disorder: Positive Findings from a Proof-of-Concept Study. Photobiomodul. Photomed. Laser Surg. 2022, 40, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Leisman, G.; Machado, C.; Machado, Y.; Chinchilla-Acosta, M. Effects of Low-Level Laser Therapy in Autism Spectrum Disorder. Adv. Exp. Med. Biol. 2018, 1116, 111–130. [Google Scholar] [CrossRef] [PubMed]
- Machado, C.; Machado, Y.; Chinchilla, M.; Machado, Y. Follow-Up Assessment of Autistic Children 6 Months after Finishing Low Lever Laser Therapy. Internet J. Neurol. 2019, 21, 1–8. [Google Scholar]
- Machado, C.; Machado, Y.; Chinchilla, M.; Machado, Y. Twelve Months Follow-up Comparison between the Autistic Children vs. Intial Placebo (Treated) Groups. Internet J. Neurol. 2019, 21, 2. [Google Scholar]
- Surapaty, I.A.; Simadibrata, C.; Rejeki, E.S.; Mangunatmadja, I. Laser Acupuncture Effects on Speech and Social Interaction in Patients with Autism Spectrum Disorder. Med. Acupunct. 2020, 32, 300–309. [Google Scholar] [CrossRef]
- Knyazkova, A.; Shulmina, P.; Samarinova, A.; Kistenev, Y.; Borisov, A. Electroencephalography Registration of Laser Acupuncture Action on Children with Autism Disorder. In Proceedings of the 13th International Joint Conference on Biomedical Engineering Systems and Technologies; SCITEPRESS—Science and Technology Publications, Valletta, Malta, 24 February 2020; pp. 387–394. [Google Scholar]
- Khongrum, J.; Wattanathorn, J. Laser Acupuncture Improves Behavioral Disorders and Brain Oxidative Stress Status in the Valproic Acid Rat Model of Autism. J. Acupunct. Meridian Stud. 2015, 8, 183–191. [Google Scholar] [CrossRef]
- Khongrum, J.; Wattanathorn, J. Laser Acupuncture at HT7 Improves the Cerebellar Disorders in Valproic Acid-Rat Model of Autism. J. Acupunct. Meridian Stud. 2017, 10, 231–239. [Google Scholar] [CrossRef]
- Ergaz, Z.; Weinstein-Fudim, L.; Ornoy, A. Genetic and non-genetic animal models for autism spectrum disorders (ASD). Reprod. Toxicol. 2016, 64, 116–140. [Google Scholar] [CrossRef]
- Saltmarche, A.E.; Naeser, M.A.; Ho, K.F.; Hamblin, M.R.; Lim, L. Significant Improvement in Cognition in Mild to Moderately Severe Dementia Cases Treated with Transcranial Plus Intranasal Photobiomodulation: Case Series Report. Photomed. Laser Surg. 2017, 35, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, C.; El Khoury, H.; Hamilton, D.; Nicklason, F.; Mitrofanis, J. The “Buckets”: The Use of Red and Infrared Light Helmets in Parkinson’s Disease Patients. Photobiomodul. Photomed. Laser Surg. 2019, 37, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, C.; Hamilton, D.; Nicklason, F.; El Massri, N.; Mitrofanis, J. Exploring the Use of Transcranial Photobiomodulation in Parkinson’s Disease Patients. Neural Regen. Res. 2018, 13, 1738–1740. [Google Scholar] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamilton, C.; Liebert, A.; Pang, V.; Magistretti, P.; Mitrofanis, J. Lights on for Autism: Exploring Photobiomodulation as an Effective Therapeutic Option. Neurol. Int. 2022, 14, 884-893. https://doi.org/10.3390/neurolint14040071
Hamilton C, Liebert A, Pang V, Magistretti P, Mitrofanis J. Lights on for Autism: Exploring Photobiomodulation as an Effective Therapeutic Option. Neurology International. 2022; 14(4):884-893. https://doi.org/10.3390/neurolint14040071
Chicago/Turabian StyleHamilton, Catherine, Ann Liebert, Vincent Pang, Pierre Magistretti, and John Mitrofanis. 2022. "Lights on for Autism: Exploring Photobiomodulation as an Effective Therapeutic Option" Neurology International 14, no. 4: 884-893. https://doi.org/10.3390/neurolint14040071
APA StyleHamilton, C., Liebert, A., Pang, V., Magistretti, P., & Mitrofanis, J. (2022). Lights on for Autism: Exploring Photobiomodulation as an Effective Therapeutic Option. Neurology International, 14(4), 884-893. https://doi.org/10.3390/neurolint14040071