

Changes in TRPV1 Expression as Well as Substance P and Vasoactive Intestinal Peptide Levels Are Associated with Recurrence of Pterygium


Abstract
1. Introduction
2. Results
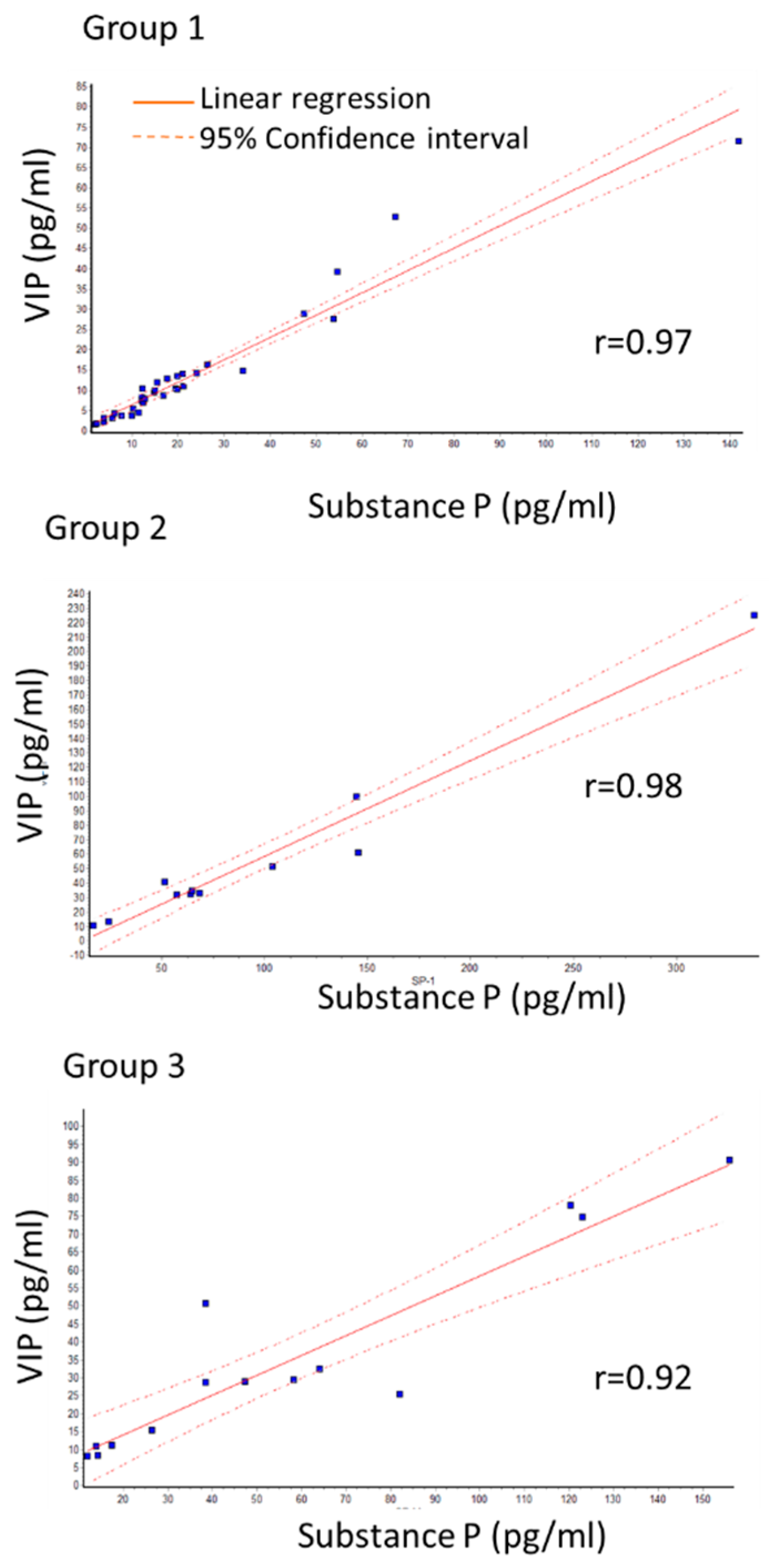
2.1. Changes in SP and VIP Levels
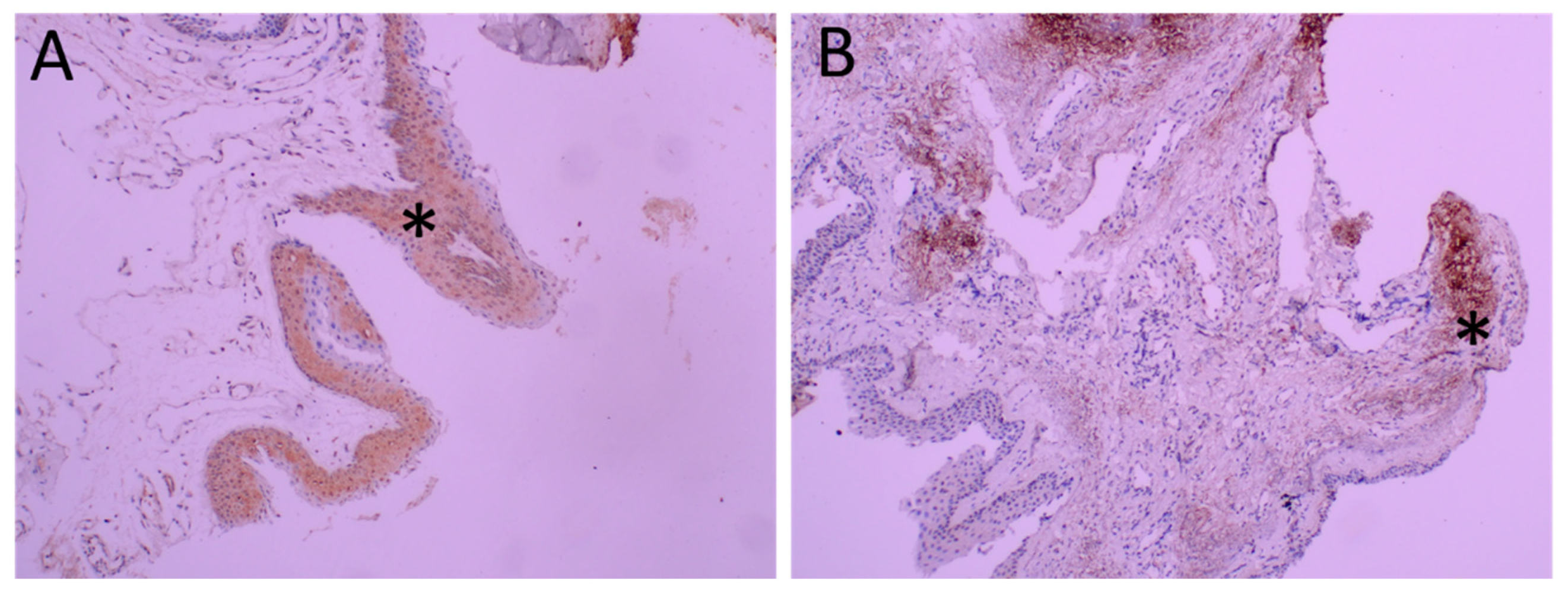
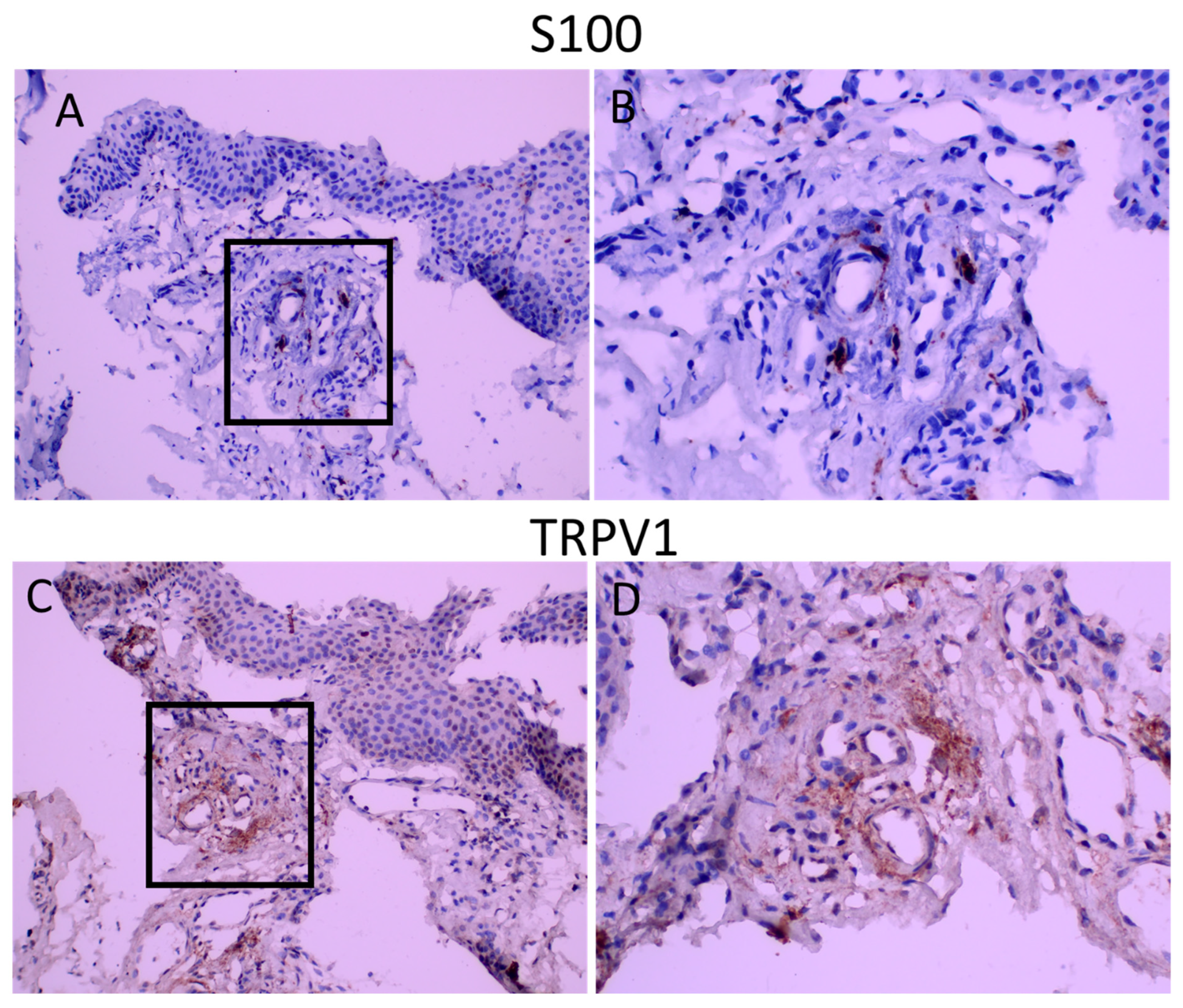
2.2. Changes in TRPV1 Expression
2.3. TRPV1 Expression in Epithelium and Stroma (Table 4)

{kind=link}
{kind=link}
{kind=link}
| 3–4+ | TRPV1 Distribution | TRPV1 Intensity | S100 Distribution (2–3+) | |
|---|---|---|---|---|
| Group 1 | Epithelium | 9/18 | 3/18 | |
| Stroma | 6/18 | 10/18 * | 12/18 (67%) | |
| Group 2 | Epithelium | 4/8 | 1/8 | |
| Stroma | 1/8 | 0/8 ** | 5/8 (62.5%) | |
| Group 3 | Epithelium | 4/9 | 0/9 | |
| Stroma | 1/9 | 3/9 | 4/9 (44%) |
2.4. TRPV1 Expression in Stroma (Table 4)
2.5. S100 Immunoreactivity
3. Discussion
4. Materials and Methods
4.1. Measurement of SP and VIP Levels
4.2. Histopathological Evaluation
4.3. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wong, W.W. A hypothesis on the pathogenesis of pterygiums. Ann. Ophthalmol. 1978, 10, 303–308. [Google Scholar] [PubMed]
- Chui, J.; Di Girolamo, N.; Wakefield, D.; Coroneo, M.T. The pathogenesis of pterygium: Current concepts and their therapeutic implications. Ocul. Surf. 2008, 6, 24–43. [Google Scholar] [CrossRef] [PubMed]
- Kria, L.; Ohira, A.; Amemiya, T. Immunohistochemical localization of basic fibroblast growth factor, platelet derived growth factor, transforming growth factor-beta and tumor necrosis factor-alpha in the pterygium. Acta Histochem. 1996, 98, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Kria, L.; Ohira, A.; Amemiya, T. Growth factors in cultured pterygium fibroblasts: Immunohistochemical and ELISA analysis. Graefes Arch. Clin. Exp. Ophthalmol. 1998, 236, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Di Girolamo, N.; Kumar, R.K.; Coroneo, M.T.; Wakefield, D. UVB-mediated induction of interleukin-6 and -8 in pterygia and cultured human pterygium epithelial cells. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3430–3437. [Google Scholar]
- Fernandes, M.; Sangwan, V.S.; Bansal, A.K.; Gangopadhyay, N.; Sridhar, M.S.; Garg, P.; Aasuri, M.K.; Nutheti, R.; Rao, G.N. Outcome of pterygium surgery: Analysis over 14 years. Eye 2005, 19, 1182–1190. [Google Scholar] [CrossRef]
- Muller, L.J.; Marfurt, C.F.; Kruse, F.; Tervo, T.M. Corneal nerves: Structure, contents and function. Exp. Eye Res. 2003, 76, 521–542. [Google Scholar] [CrossRef]
- Auran, J.D.; Koester, C.J.; Kleiman, N.J.; Rapaport, R.; Bomann, J.S.; Wirotsko, B.M.; Florakis, G.J.; Koniarek, J.P. Scanning slit confocal microscopic observation of cell morphology and movement within the normal human anterior cornea. Ophthalmology 1995, 102, 33–41. [Google Scholar] [CrossRef]
- Chui, J.; Di Girolamo, N.; Coroneo, M.T.; Wakefield, D. The role of substance P in the pathogenesis of pterygia. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4482–4489. [Google Scholar] [CrossRef]
- Fischer, M.J.M.; Ciotu, C.I.; Szallasi, A. The Mysteries of Capsaicin-Sensitive Afferents. Front. Physiol. 2020, 11, 554195. [Google Scholar] [CrossRef]
- Gallar, J.; Pozo, M.A.; Rebollo, I.; Belmonte, C. Effects of capsaicin on corneal wound healing. Investig. Ophthalmol. Vis. Sci. 1990, 31, 1968–1974. [Google Scholar]
- Araki, K.; Ohashi, Y.; Kinoshita, S.; Hayashi, K.; Kuwayama, Y.; Tano, Y. Epithelial wound healing in the denervated cornea. Curr. Eye Res. 1994, 13, 203–211. [Google Scholar] [CrossRef]
- Garcia-Hirschfeld, J.; Lopez-Briones, L.G.; Belmonte, C. Neurotrophic influences on corneal epithelial cells. Exp. Eye Res. 1994, 59, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Mikulec, A.A.; Tanelian, D.L. CGRP increases the rate of corneal re-epithelialization in an in vitro whole mount preparation. J. Ocul. Pharmacol. Ther. 1996, 12, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.J.; Marfurt, C.F.; McDermott, A.; Bentley, E.; Abrams, G.A.; Reid, T.W.; Campbell, S. Spontaneous chronic corneal epithelial defects (SCCED) in dogs: Clinical features, innervation, and effect of topical SP, with or without IGF-1. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2252–2261. [Google Scholar]
- Seifert, P.; Stuppi, S.; Spitznas, M.; Weihe, E. Differential distribution of neuronal markers and neuropeptides in the human lacrimal gland. Graefes Arch. Clin. Exp. Ophthalmol. 1996, 234, 232–240. [Google Scholar] [CrossRef]
- Satitpitakul, V.; Sun, Z.; Suri, K.; Amouzegar, A.; Katikireddy, K.R.; Jurkunas, U.V.; Kheirkhah, A.; Dana, R. Vasoactive Intestinal Peptide Promotes Corneal Allograft Survival. Am. J. Pathol. 2018, 188, 2016–2024. [Google Scholar] [CrossRef]
- Ladek, A.M.; Trost, A.; Bruckner, D.; Schroedl, F.; Kaser-Eichberger, A.; Lenzhofer, M.; Reitsamer, H.A.; Strohmaier, C.A. Immunohistochemical Characterization of Neurotransmitters in the Episcleral Circulation in Rats. Investig. Ophthalmol. Vis. Sci. 2019, 60, 3215–3220. [Google Scholar] [CrossRef]
- Nolano, M.; Provitera, V.; Caporaso, G.; Stancanelli, A.; Leandri, M.; Biasiotta, A.; Cruccu, G.; Santoro, L.; Truini, A. Cutaneous innervation of the human face as assessed by skin biopsy. J. Anat. 2013, 222, 161–169. [Google Scholar] [CrossRef]
- Assimakopoulou, M.; Pagoulatos, D.; Nterma, P.; Pharmakakis, N. Immunolocalization of cannabinoid receptor type 1 and CB2 cannabinoid receptors, and transient receptor potential vanilloid channels in pterygium. Mol. Med. Rep. 2017, 16, 5285–5293. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, J.; Liang, H.; Zhang, S.; Shao, C.; Fan, X.; Fu, Y. Corneal Reinnervation and Sensitivity Recovery after Pterygium Excision. J. Ophthalmol. 2020, 2020, 1349072. [Google Scholar] [CrossRef]
- Mergler, S.; Valtink, M.; Coulson-Thomas, V.J.; Lindemann, D.; Reinach, P.S.; Engelmann, K.; Pleyer, U. TRPV channels mediate temperature-sensing in human corneal endothelial cells. Exp. Eye Res. 2010, 90, 758–770. [Google Scholar] [CrossRef] [PubMed]
- Mergler, S.; Garreis, F.; Sahlmuller, M.; Reinach, P.S.; Paulsen, F.; Pleyer, U. Thermosensitive transient receptor potential channels in human corneal epithelial cells. J. Cell Physiol. 2011, 226, 1828–1842. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Wang, Z.; Yang, H.; Zhang, F.; Reinach, P.S. TRPV1 activation is required for hypertonicity-stimulated inflammatory cytokine release in human corneal epithelial cells. Investig. Ophthalmol. Vis. Sci. 2011, 52, 485–493. [Google Scholar] [CrossRef]
- Tan, D.T.; Chee, S.P.; Dear, K.B.; Lim, A.S. Effect of pterygium morphology on pterygium recurrence in a controlled trial comparing conjunctival autografting with bare sclera excision. Arch. Ophthalmol. 1997, 115, 1235–1240. [Google Scholar] [CrossRef]
- Tan, J.; Vollmer-Conna, U.; Tat, L.; Coroneo, M. Dry-Eye Disease in Recurrent Pterygium. Ophthalmic. Res. 2019, 61, 199–203. [Google Scholar] [CrossRef]
- Garreis, F.; Schroder, A.; Reinach, P.S.; Zoll, S.; Khajavi, N.; Dhandapani, P.; Lucius, A.; Pleyer, U.; Paulsen, F.; Mergler, S. Upregulation of Transient Receptor Potential Vanilloid Type-1 Channel Activity and Ca2+ Influx Dysfunction in Human Pterygial Cells. Investig. Ophthalmol. Vis. Sci. 2016, 57, 2564–2577. [Google Scholar] [CrossRef] [PubMed]
- Jiao, H.; Ivanusic, J.J.; McMenamin, P.G.; Chinnery, H.R. Distribution of Corneal TRPV1 and Its Association With Immune Cells During Homeostasis and Injury. Investig. Ophthalmol. Vis. Sci. 2021, 62, 6. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, C.; Aracil, A.; Acosta, M.C.; Luna, C.; Gallar, J. Nerves and sensations from the eye surface. Ocul. Surf. 2004, 2, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Erin, N.; Clawson, G.A. Parameters affecting substance P measurement in heart, lung, and skin. Biotechniques 2004, 37, 232–239. [Google Scholar] [CrossRef]
- Bignami, F.; Giacomini, C.; Lorusso, A.; Aramini, A.; Rama, P.; Ferrari, G. NK1 receptor antagonists as a new treatment for corneal neovascularization. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6783–6794. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.S.; Lee, J.; Lee, E.; Kwon, Y.S.; Lee, E.; Ahn, W.; Jiang, M.H.; Kim, J.C.; Son, Y. A new role of substance P as an injury-inducible messenger for mobilization of CD29(+) stromal-like cells. Nat. Med. 2009, 15, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Araki-Sasaki, K.; Aizawa, S.; Hiramoto, M.; Nakamura, M.; Iwase, O.; Nakata, K.; Sasaki, Y.; Mano, T.; Handa, H.; Tano, Y. Substance P-induced cadherin expression and its signal transduction in a cloned human corneal epithelial cell line. J. Cell Physiol. 2000, 182, 189–195. [Google Scholar] [CrossRef]
- Nishida, T.; Inui, M.; Nomizu, M. Peptide therapies for ocular surface disturbances based on fibronectin-integrin interactions. Prog. Retin. Eye Res. 2015, 47, 38–63. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Di, G.; Qi, X.; Qu, M.; Wang, Y.; Duan, H.; Danielson, P.; Xie, L.; Zhou, Q. Substance P promotes diabetic corneal epithelial wound healing through molecular mechanisms mediated via the neurokinin-1 receptor. Diabetes 2014, 63, 4262–4274. [Google Scholar] [CrossRef]
- Nakamura, M.; Ofuji, K.; Chikama, T.; Nishida, T. The NK1 receptor and its participation in the synergistic enhancement of corneal epithelial migration by substance P and insulin-like growth factor-1. Br. J. Pharmacol. 1997, 120, 547–552. [Google Scholar] [CrossRef]
- Seifert, P.; Spitznas, M. Vasoactive intestinal polypeptide (VIP) innervation of the human eyelid glands. Exp. Eye Res. 1999, 68, 685–692. [Google Scholar] [CrossRef]
- Cox, S.M.; Nichols, J.J. The neurobiology of the meibomian glands. Ocul. Surf. 2014, 12, 167–177. [Google Scholar] [CrossRef]
- Diebold, Y.; Rios, J.D.; Hodges, R.R.; Rawe, I.; Dartt, D.A. Presence of nerves and their receptors in mouse and human conjunctival goblet cells. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2270–2282. [Google Scholar]
- Villanueva-Romero, R.; Gutierrez-Canas, I.; Carrion, M.; Perez-Garcia, S.; Seoane, I.V.; Martinez, C.; Gomariz, R.P.; Juarranz, Y. The Anti-Inflammatory Mediator, Vasoactive Intestinal Peptide, Modulates the Differentiation and Function of Th Subsets in Rheumatoid Arthritis. J. Immunol. Res. 2018, 2018, 6043710. [Google Scholar] [CrossRef]
- He, Y.; Zang, N.; Liu, E.M. Vasoactive intestinal peptide: A potential target for antiviral therapy. Sheng Li Xue Bao 2022, 74, 419–433. [Google Scholar] [PubMed]
- Jansen, M.I.; Thomas Broome, S.; Castorina, A. Exploring the Pro-Phagocytic and Anti-Inflammatory Functions of PACAP and VIP in Microglia: Implications for Multiple Sclerosis. Int. J. Mol. Sci. 2022, 23, 4788. [Google Scholar] [CrossRef] [PubMed]
- Wilhelmsen, K.; Khakpour, S.; Tran, A.; Sheehan, K.; Schumacher, M.; Xu, F.; Hellman, J. The endocannabinoid/endovanilloid N-arachidonoyl dopamine (NADA) and synthetic cannabinoid WIN55,212-2 abate the inflammatory activation of human endothelial cells. J. Biol. Chem. 2014, 289, 13079–13100. [Google Scholar] [CrossRef] [PubMed]
- Joffre, J.; Wong, E.; Lawton, S.; Lloyd, E.; Nguyen, N.; Xu, F.; Sempio, C.; Kobzik, L.; Zlatanova, I.; Schumacher, M.; et al. N-Oleoyl dopamine induces IL-10 via central nervous system TRPV1 and improves endotoxemia and sepsis outcomes. J. Neuroinflammation 2022, 19, 118. [Google Scholar] [CrossRef]
- Erin, N.; Ulusoy, O. Differentiation of neuronal from non-neuronal Substance P. Regul. Pept. 2009, 152, 108–113. [Google Scholar] [CrossRef]



| Group 1 | Group 2 | Group 3 | |
|---|---|---|---|
| Gender (n) M/F | 18/15 | 7/4 | 14/6 |
| Age | 54.82 ± 5.48 | 52.36 ± 5.03 | 53.55 ± 2.99 |
| Pterygium vertical length | 3.77 ± 0.11 | 4.5 ± 0.3 | 4.02 ± 0.21 |
| Duration between recurrences (month) | 0 | 7.73±1.16 | 81 ± 17.8 * |
| Recurrence number | 0 | 1 | 2.3 ± 0.39 |
| Group 1 (n= 33) | Group 2 (n= 11) | Group 1 vs. Group 2 | Group 3 (n= 20) | Group 1 vs. Group 3 | |
|---|---|---|---|---|---|
| VIP | 13.59 (±2.63) | 57.38 (±18.31) | p < 0.001 | 35.60 (±6.4) | p < 0.05 |
| SP | 22.99 (±4.61) | 98.14 (±27.13) | p < 0.001 | 56.94 (±10.18) | p < 0.05 |
| Recurrence after Last Surgery (n) | SP | VIP | |
|---|---|---|---|
| 0–6 months | 11 | 112.53 ± 25.63 * | 57.37 ± 24.79 |
| After 6 months | 6 | 43.39 ± 9.75 | 41.83 ± 11.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
İlhan, H.D.; Ünal, B.; Ayaz, Y.; Erin, N. Changes in TRPV1 Expression as Well as Substance P and Vasoactive Intestinal Peptide Levels Are Associated with Recurrence of Pterygium. Int. J. Mol. Sci. 2022, 23, 15692. https://doi.org/10.3390/ijms232415692
İlhan HD, Ünal B, Ayaz Y, Erin N. Changes in TRPV1 Expression as Well as Substance P and Vasoactive Intestinal Peptide Levels Are Associated with Recurrence of Pterygium. International Journal of Molecular Sciences. 2022; 23(24):15692. https://doi.org/10.3390/ijms232415692
Chicago/Turabian Styleİlhan, Hatice Deniz, Betül Ünal, Yusuf Ayaz, and Nuray Erin. 2022. "Changes in TRPV1 Expression as Well as Substance P and Vasoactive Intestinal Peptide Levels Are Associated with Recurrence of Pterygium" International Journal of Molecular Sciences 23, no. 24: 15692. https://doi.org/10.3390/ijms232415692
APA Styleİlhan, H. D., Ünal, B., Ayaz, Y., & Erin, N. (2022). Changes in TRPV1 Expression as Well as Substance P and Vasoactive Intestinal Peptide Levels Are Associated with Recurrence of Pterygium. International Journal of Molecular Sciences, 23(24), 15692. https://doi.org/10.3390/ijms232415692