
Genomic and Phenotypic Characterization of a Novel Virulent Strain of Cyvirus cyprinidallo2 Originating from an Outbreak in The Netherlands

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Handling
2.2. In Vitro Methods
2.2.1. CyHV-2 PCR Detection
2.2.2. Cells and Virus
2.2.3. Initial In Vitro Culture and Characterization of the CyHV-2 Isolates
2.2.4. Plaque Purification and Amplification
2.2.5. Viral DNA Extraction and Restriction Fragment Length Polymorphism (RFLP) Analysis
2.2.6. Genome Sequencing, Assembly, Phylogenetic and Mutation Analysis
2.2.7. Viral Growth Assay
2.2.8. Viral Plaque Size Assay
2.3. In Vivo
2.3.1. Fish
2.3.2. Infection of Fish with CyHV-2
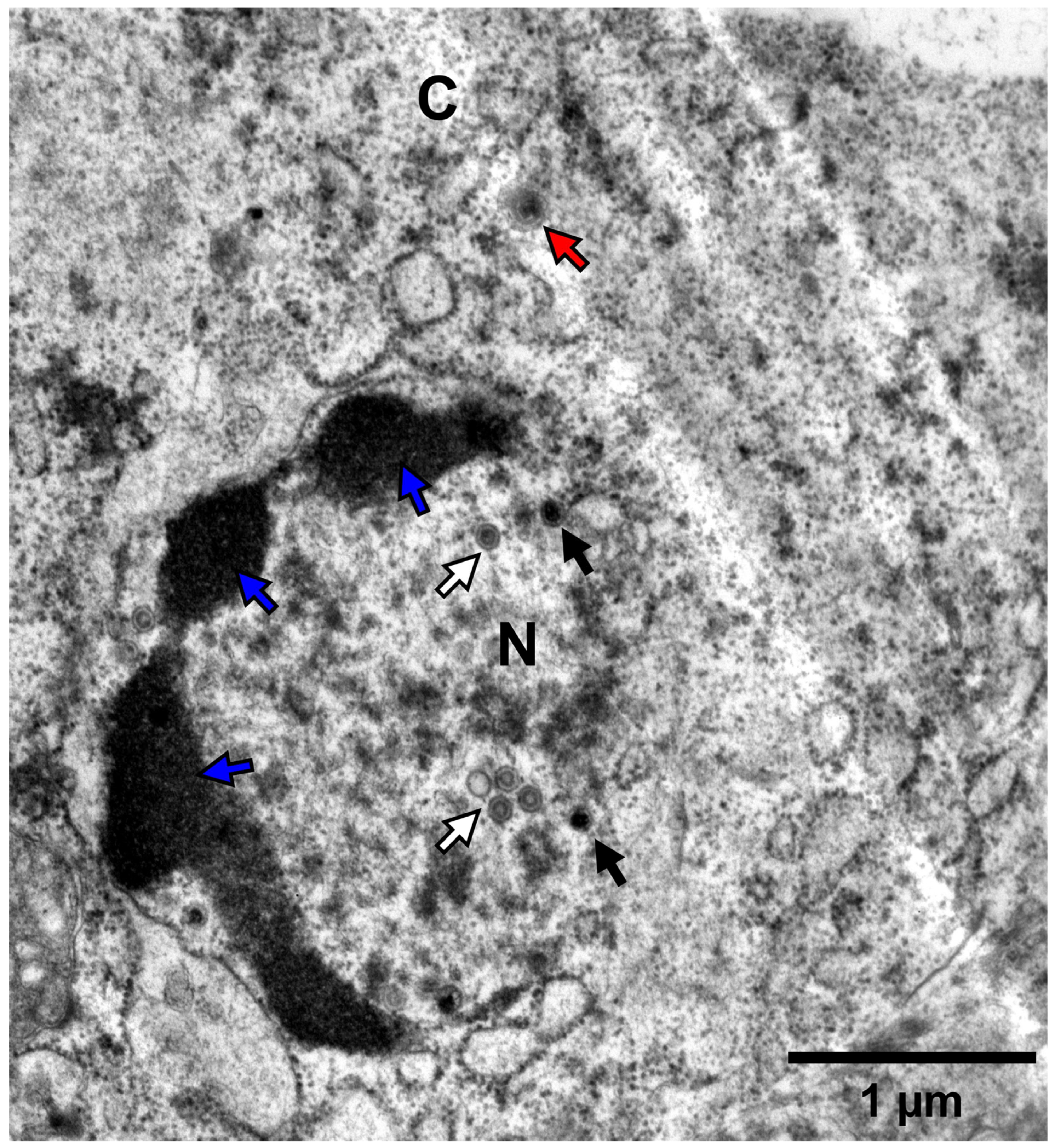
2.3.3. Transmission Electron Microscopy
2.4. Statistical Analysis
2.5. Ethics Statement
3. Results
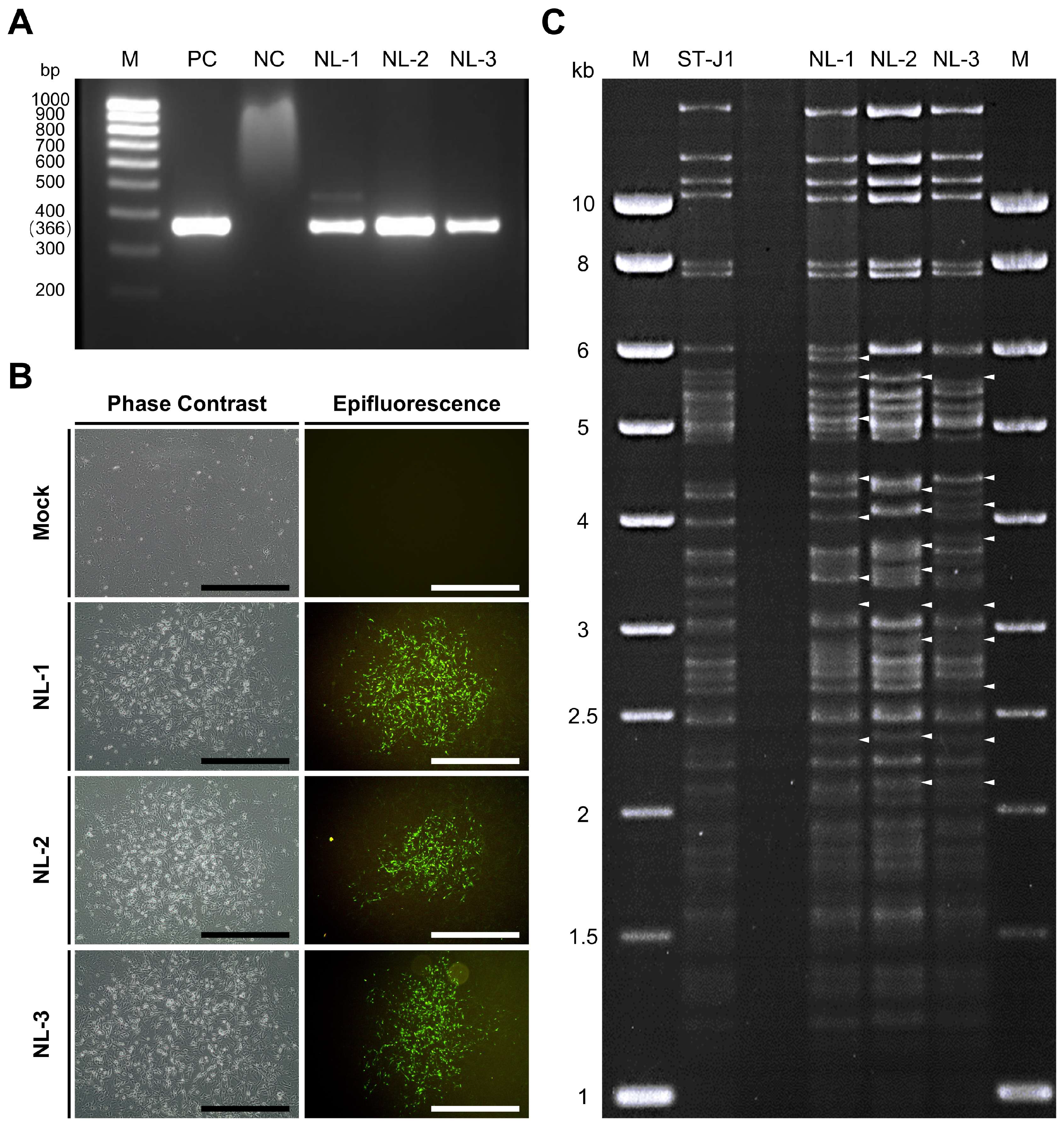
3.1. Confirmation of CyHV-2 Presence in Pooled Organ Samples
3.2. Virus Isolation and Amplification
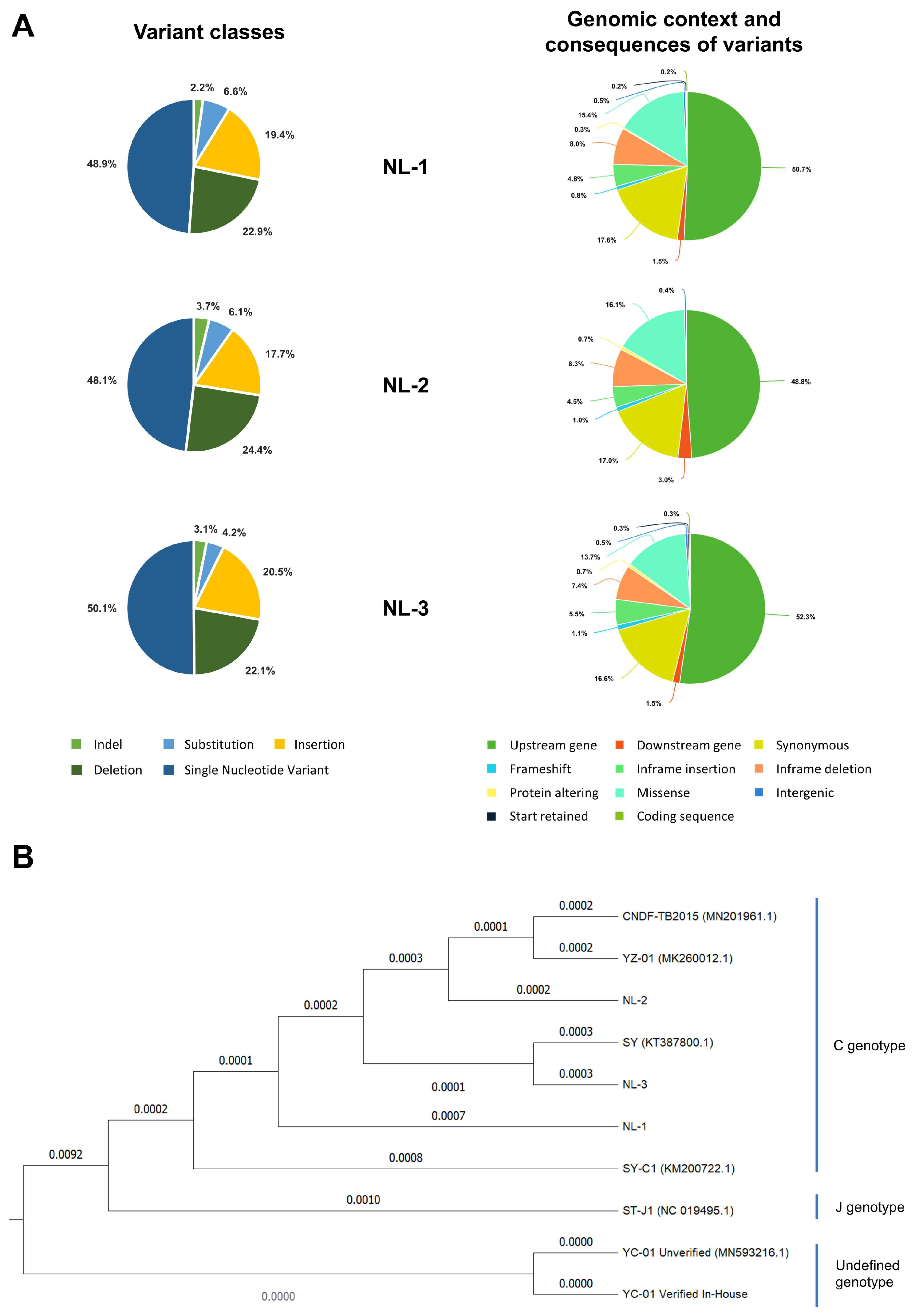
3.3. Genomic and Phylogenetic Analysis
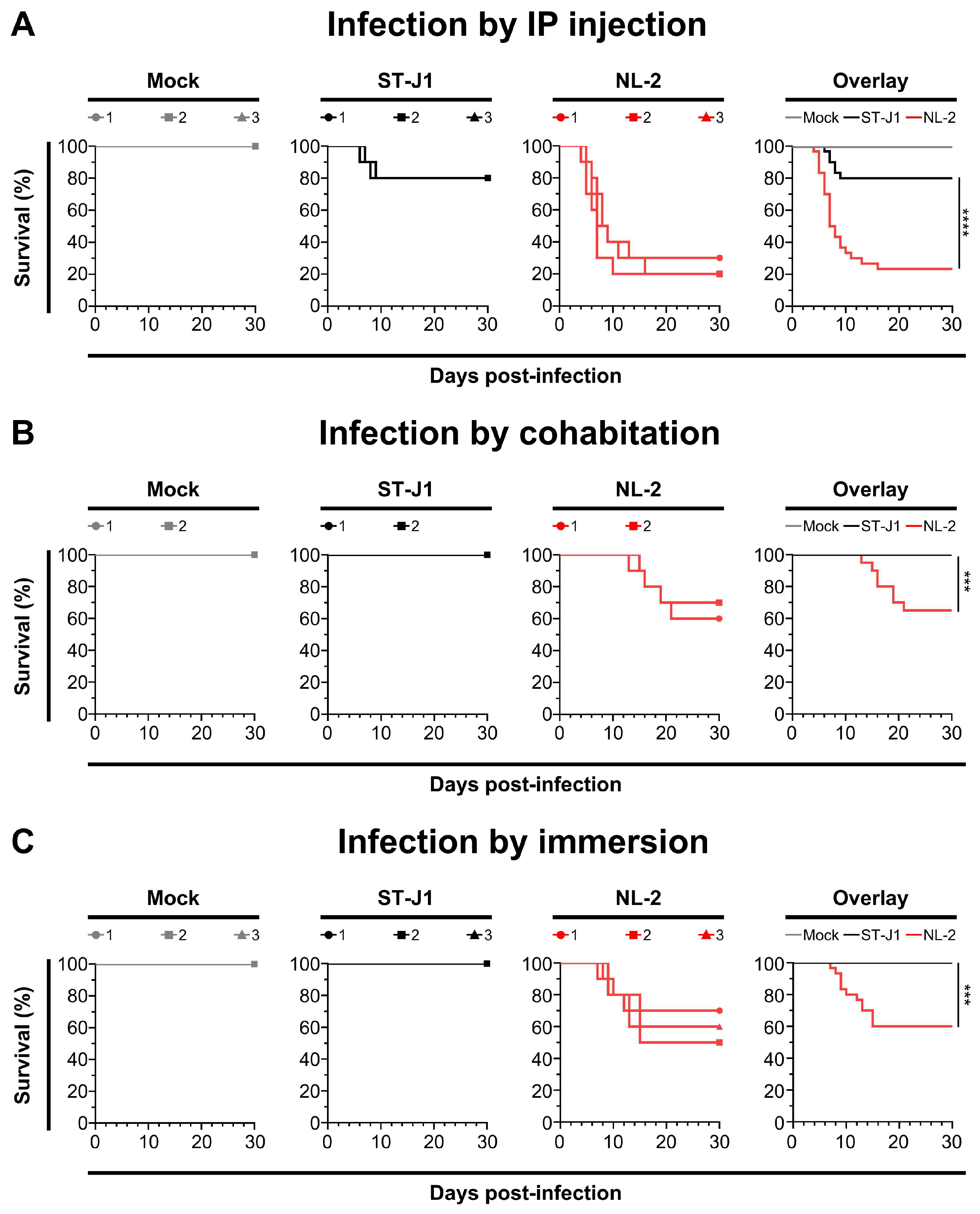
3.4. Pathogenicity of the Three Isolates
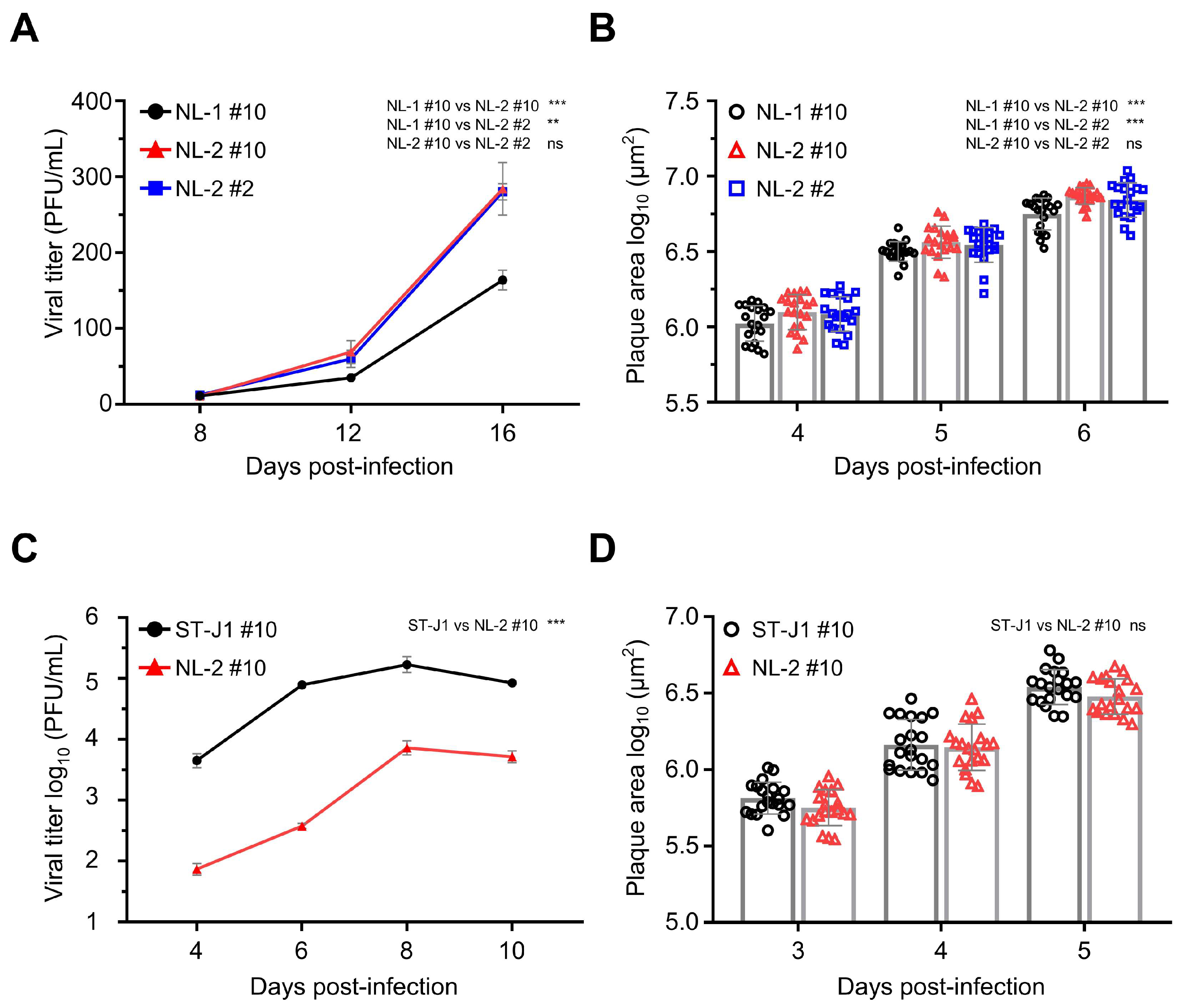
3.5. In Vitro Growth Kinetics Comparison
3.6. Comparison of In Vivo Pathogenicity of CyHV-2 NL-2 Strain and Reference Strain ST-J1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanson, L.A.; Doszpoly, A.; van Beurden, S.; de Oliveira Viadanna, P.H.; Waltzek, T. Chapter 8—Alloherpesviruses of Fish. In Aquaculture Virology, 2nd ed.; Kibenge, F.S.B., Godoy, M.G., Eds.; Academic Press: Cambridge, MA, USA, 2024; pp. 165–189. ISBN 978-0-323-91169-6. [Google Scholar]
- Boutier, M.; Morvan, L.; Delrez, N.; Origgi, F.; Doszpoly, A.; Vanderplasschen, A. Fish and Amphibian Alloherpesviruses (Herpesviridae). In Encyclopedia of Virology, 4th ed.; Bamford, D.H., Zuckerman, M., Eds.; Academic Press: Oxford, UK, 2021; pp. 306–315. ISBN 978-0-12-814516-6. [Google Scholar]
- Pradeepkiran, J.A. Aquaculture Role in Global Food Security with Nutritional Value: A Review. Transl. Anim. Sci. 2019, 3, 903–910. [Google Scholar] [CrossRef] [PubMed]
- ICTV. Available online: https://ictv.global/taxonomy (accessed on 5 September 2024).
- Donohoe, O.; Zhang, H.; Delrez, N.; Gao, Y.; Suárez, N.M.; Davison, A.J.; Vanderplasschen, A. Genomes of Anguillid Herpesvirus 1 Strains Reveal Evolutionary Disparities and Low Genetic Diversity in the Genus Cyprinivirus. Microorganisms 2021, 9, 998. [Google Scholar] [CrossRef] [PubMed]
- Streiff, C.; He, B.; Morvan, L.; Zhang, H.; Delrez, N.; Fourrier, M.; Manfroid, I.; Suárez, N.M.; Betoulle, S.; Davison, A.J.; et al. Susceptibility and Permissivity of Zebrafish (Danio rerio) Larvae to Cypriniviruses. Viruses 2023, 15, 768. [Google Scholar] [CrossRef] [PubMed]
- Davison, A.J.; Kurobe, T.; Gatherer, D.; Cunningham, C.; Korf, I.; Fukuda, H.; Hedrick, R.P.; Waltzek, T.B. Comparative Genomics of Carp Herpesviruses. J. Virol. 2013, 87, 2908–2922. [Google Scholar] [CrossRef]
- Padhiary, S.; Paul, A.; Tripathy, D.K.; Nayak, D.; Mohanty, J.; Pillai, D.; Rejish Kumar, V.J.; Krishnan, R.; Haridas, L.; Abdul Majeed, S.; et al. Current understanding of Cyprinid herpesvirus 2, and prospects in its management. Indian J. Anim. Health 2023, 62, 49–60. [Google Scholar] [CrossRef]
- Jung, S.J.; Miyazaki, T. Herpesviral Haematopoietic Necrosis of Goldfish, Carassius auratus (L.). J. Fish Dis. 1995, 18, 211–220. [Google Scholar] [CrossRef]
- Thangaraj, R.S.; Nithianantham, S.R.; Dharmaratnam, A.; Kumar, R.; Pradhan, P.K.; Thangalazhy Gopakumar, S.; Sood, N. Cyprinid Herpesvirus-2 (CyHV-2): A Comprehensive Review. Rev. Aquac. 2021, 13, 796–821. [Google Scholar] [CrossRef]
- Zhang, H.; Sridhar, A.; Delrez, N.; He, B.; Fourny, S.; Gao, Y.; Donohoe, O.; Vanderplasschen, A.F.C. Development Using Bioluminescence Imaging of a Recombinant Anguillid Herpesvirus 1 Vaccine Candidate Associated with Normal Replication In Vitro but Abortive Infection In Vivo. Vaccines 2024, 12, 1423. [Google Scholar] [CrossRef]
- Boutier, M.; Ronsmans, M.; Ouyang, P.; Fournier, G.; Reschner, A.; Rakus, K.; Wilkie, G.S.; Farnir, F.; Bayrou, C.; Lieffrig, F.; et al. Rational Development of an Attenuated Recombinant Cyprinid Herpesvirus 3 Vaccine Using Prokaryotic Mutagenesis and In Vivo Bioluminescent Imaging. PLoS Pathog. 2015, 11, e1004690. [Google Scholar] [CrossRef]
- Saito, H.; Okamura, T.; Shibata, T.; Kato, G.; Sano, M. Development of a Live Attenuated Vaccine Candidate against Herpesviral Hematopoietic Necrosis of Goldfish. Aquaculture 2022, 552, 737974. [Google Scholar] [CrossRef]
- Sun, Y.; Xu, C.; Wang, H.; Qiao, G.; Wang, Z.; Li, Z.; Li, Q.; Wei, C. An Attenuated Strain of Cyprinid Herpesvirus 2 as a Vaccine Candidate against Herpesviral Hematopoietic Necrosis Disease in Gibel Carp, Carassius auratus gibelio. Fish Shellfish Immunol. 2023, 138, 108826. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Sridhar, A.; Streiff, C.; Deketelaere, C.; Zhang, H.; Gao, Y.; Hu, Y.; Pirotte, S.; Delrez, N.; Davison, A.J.; et al. In Vivo Imaging Sheds Light on the Susceptibility and Permissivity of Carassius auratus to Cyprinid Herpesvirus 2 According to Developmental Stage. Viruses 2023, 15, 1746. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Kurita, J.; Haenen, O.L.M. Importation of CyHV-2-Infected Goldfish into the Netherlands. Dis. Aquat. Org. 2017, 126, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Kurobe, T.; Kurita, J.; Haenen, O.; Voorbergen-Laarman, M.; Ito, T. Mass Mortality Events Associated with Cyprinid Herpesvirus 2 (CyHV-2) Infection in Wild Prussian Carp Carassius gibelio in the Netherlands, and Molecular Biology of Virus Strains. J. Fish Dis. 2024, 47, e13868. [Google Scholar] [CrossRef]
- Waltzek, T.B.; Kurobe, T.; Goodwin, A.E.; Hedrick, R.P. Development of a Polymerase Chain Reaction Assay to Detect Cyprinid Herpesvirus 2 in Goldfish. J. Aquat. Anim. Health 2009, 21, 60–67. [Google Scholar] [CrossRef]
- Shibata, T.; Nanjo, A.; Saito, M.; Yoshii, K.; Ito, T.; Nakanishi, T.; Sakamoto, T.; Sano, M. In Vitro Characteristics of Cyprinid Herpesvirus 2: Effect of Kidney Extract Supplementation on Growth. Dis. Aquat. Org. 2015, 115, 223–232. [Google Scholar] [CrossRef]
- BBTools. Available online: https://jgi.doe.gov/data-and-tools/software-tools/bbtools/ (accessed on 5 October 2022).
- Antipov, D.; Korobeynikov, A.; McLean, J.S.; Pevzner, P.A. hybridSPAdes: An Algorithm for Hybrid Assembly of Short and Long Reads. Bioinformatics 2016, 32, 1009–1015. [Google Scholar] [CrossRef]
- Mikheenko, A.; Prjibelski, A.; Saveliev, V.; Antipov, D.; Gurevich, A. Versatile Genome Assembly Evaluation with QUAST-LG. Bioinformatics 2018, 34, i142–i150. [Google Scholar] [CrossRef]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive Visualization of de Novo Genome Assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef]
- SnapGene LLC. SnapGene | Software for Everyday Molecular Biology. Available online: https://www.snapgene.com/ (accessed on 7 May 2024).
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, Interactive Sequence Choice and Visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Complete Genomes: Alloherpesviridae. Available online: https://www.ncbi.nlm.nih.gov/genomes/GenomesGroup.cgi?taxid=548682 (accessed on 7 August 2024).
- Viral Genomes. Available online: https://www.ncbi.nlm.nih.gov/genome/viruses/ (accessed on 7 August 2024).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Lindenbaum, P. JVarkit: Java-Based Utilities for Bioinformatics. 2015. Available online: https://github.com/lindenb/jvarkit (accessed on 1 April 2025).
- MsaToVcf. Available online: http://lindenb.github.io/jvarkit/MsaToVcf.html (accessed on 27 June 2024).
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-Level Expression Analysis of RNA-Seq Experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve Years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative Genomics Viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef]
- Geneious | Bioinformatics Software. Available online: https://www.geneious.com/ (accessed on 4 July 2024).
- Baldwin, S.; Revanna, R.; Thomson, S.; Pither-Joyce, M.; Wright, K.; Crowhurst, R.; Fiers, M.; Chen, L.; Macknight, R.; McCallum, J.A. A Toolkit for Bulk PCR-Based Marker Design from next-Generation Sequence Data: Application for Development of a Framework Linkage Map in Bulb Onion (Allium cepa L.). BMC Genom. 2012, 13, 637. [Google Scholar] [CrossRef]
- McCallum, J. Cfljam/Galaxy-Pcr-Markers 2022. Available online: https://github.com/cfljam/galaxy-pcr-markers (accessed on 1 April 2025).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- John, F.; Sanford, W. An R Companion to Applied Regression; SAGE Publications, Inc.: Los Angeles, CA, USA, 2018; ISBN 978-1-5443-3647-3. [Google Scholar]
- Lenth, R.V. Least-Squares Means: The R Package Lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef]
- Li, L.; Luo, Y.; Gao, Z.; Huang, J.; Zheng, X.; Nie, H.; Zhang, J.; Lin, L.; Yuan, J. Molecular Characterisation and Prevalence of a New Genotype of Cyprinid Herpesvirus 2 in Mainland China. Can. J. Microbiol. 2015, 61, 381–387. [Google Scholar] [CrossRef]
- Wang, F.; Xu, Y.; Zhou, Y.; Ding, C.; Duan, H. Isolation and Characterization of a Cyprinid Herpesvirus Strain YZ01 from Apparently Healthy Goldfish after Rising Water Temperature. bioRxiv 2021. [Google Scholar] [CrossRef]
- Yang, J.; Xiao, S.; Lu, L.; Wang, H.; Jiang, Y. Genomic and Molecular Characterization of a Cyprinid Herpesvirus 2 YC-01 Strain Isolated from Gibel Carp. Heliyon 2024, 10, e32811. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, K.R.; Bateman, K.; Bayley, A.; Feist, S.W.; Hulland, J.; Longshaw, C.; Stone, D.; Woolford, G.; Way, K. Isolation of a Cyprinid Herpesvirus 2 from Goldfish, Carassius auratus (L.), in the UK. J. Fish Dis. 2007, 30, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.Z.; Lin, L.; Liu, Y.; Wu, Z.X.; Gu, Z.M.; Li, L.J.; Yuan, J.F. Haematopoietic Necrosis of Cultured Prussian Carp, Carassius Gibelio (Bloch), Associated with Cyprinid Herpesvirus 2. J. Fish Dis. 2013, 36, 1035–1039. [Google Scholar] [CrossRef]
- Stephens, F.J.; Raidal, S.R.; Jones, B. Haematopoietic Necrosis in a Goldfish (Carassius auratus) Associated with an Agent Morphologically Similar to Herpesvirus. Aust. Vet. J. 2004, 82, 167–169. [Google Scholar] [CrossRef]
- Adamek, M.; Hellmann, J.; Jung-Schroers, V.; Teitge, F.; Steinhagen, D. CyHV-2 Transmission in Traded Goldfish Stocks in Germany—A Case Study. J. Fish Dis. 2018, 41, 401–404. [Google Scholar] [CrossRef]
- Boitard, P.-M.; Baud, M.; Labrut, S.; de Boisséson, C.; Jamin, M.; Bigarré, L. First Detection of Cyprinid Herpesvirus 2 (CyHV-2) in Goldfish (Carassius auratus) in France. J. Fish Dis. 2016, 39, 673–680. [Google Scholar] [CrossRef]
- Daněk, T.; Kalous, L.; Vesel, T.; Krásová, E.; Reschová, S.; Rylková, K.; Kulich, P.; L, M.P.; Pokorová, D.; Knytl, M. Massive Mortality of Prussian Carp Carassius gibelio in the Upper Elbe Basin Associated with Herpesviral Hematopoietic Necrosis (CyHV-2). Dis. Aquat. Organ. 2012, 102, 87–95. [Google Scholar] [CrossRef]
- Doszpoly, A.; Benkő, M.; Csaba, G.; Dán, Á.; Lang, M.; Harrach, B. Introduction of the Family Alloherpesviridae: The First Molecular Detection of Herpesviruses of Cyprinid Fish in Hungary. Magy. Allatorvosok Lapja 2011, 133, 174–181. [Google Scholar]
- Giovannini, S.; Bergmann, S.M.; Keeling, C.; Lany, C.; Schütze, H.; Schmidt-Posthaus, H. Herpesviral Hematopoietic Necrosis in Goldfish in Switzerland: Early Lesions in Clinically Normal Goldfish (Carassius auratus). Vet. Pathol. 2016, 53, 847–852. [Google Scholar] [CrossRef]
- Haenen, O.; Way, K.; Gorgoglione, B.; Ito, T.; Paley, R.; Bigarré, L.; Waltzek, T. Novel Viral Infections Threatening Cyprinid Fish. Bull.-Eur. Assoc. Fish Pathol. 2016, 36, 11–23. [Google Scholar]
- Schiphouwer, M.; Kessel, N.; Matthews, J.; Leuven, R.S.E.W.; Koppel, S.; Kranenbarg, J.; Haenen, O.; Lenders, H.; Nagelkerke, L.; Van der Velde, G.; et al. Risk Analysis of Exotic Fish Species Included in the Dutch Fisheries Act and Their Hybrids; Nederlands Expertise Centrum Exoten (NEC-E): Nijmegen, The Netherlands, 2014; Available online: https://edepot.wur.nl/295342 (accessed on 1 April 2025).
- Ito, T.; Kurita, J.; Ozaki, A.; Sano, M.; Fukuda, H.; Ototake, M. Growth of Cyprinid Herpesvirus 2 (CyHV-2) in Cell Culture and Experimental Infection of Goldfish Carassius auratus. Dis. Aquat. Org. 2013, 105, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Xue, M.; Xu, C.; Jiang, N.; Luo, X.; Li, Y.; Fan, Y.; Meng, Y.; Liu, W.; Zeng, L.; et al. First Report of Cyprinid Herpesvirus 2 Isolated from the Golden Crucian Carp in China. Aquaculture 2022, 558, 738361. [Google Scholar] [CrossRef]
- Xu, J.; Zeng, L.; Zhang, H.; Zhou, Y.; Ma, J.; Fan, Y. Cyprinid Herpesvirus 2 Infection Emerged in Cultured Gibel Carp, Carassius auratus Gibelio in China. Vet. Microbiol. 2013, 166, 138–144. [Google Scholar] [CrossRef]
- Jing, H.; Gao, L.; Zhang, M.; Wang, N.; Lin, X.; Zhang, L.; Wu, S. Establishment from the Snout and Kidney of Goldfish, Carassius auratus, of Two New Cell Lines and Their Susceptibility to Infectious Pancreatic Necrosis Virus. Fish Physiol. Biochem. 2016, 42, 303–311. [Google Scholar] [CrossRef]
- Gao, Y.; Sridhar, A.; Bernard, N.; He, B.; Zhang, H.; Pirotte, S.; Desmecht, S.; Vancsok, C.; Boutier, M.; Suárez, N.M.; et al. Virus-Induced Interference as a Means for Accelerating Fitness-Based Selection of Cyprinid Herpesvirus 3 Single Nucleotide Variants in Vitro and in Vivo. Virus Evol. 2023, 9, vead003. [Google Scholar] [CrossRef]
- Gao, Y.; Suárez, N.M.; Wilkie, G.S.; Dong, C.; Bergmann, S.; Lee, P.-Y.A.; Davison, A.J.; Vanderplasschen, A.F.C.; Boutier, M. Genomic and Biologic Comparisons of Cyprinid Herpesvirus 3 Strains. Vet. Res. 2018, 49, 40. [Google Scholar] [CrossRef]
- Ahmad, I.; Wilson, D.W. HSV-1 Cytoplasmic Envelopment and Egress. Int. J. Mol. Sci. 2020, 21, 5969. [Google Scholar] [CrossRef]
- Jambunathan, N.; Clark, C.M.; Musarrat, F.; Chouljenko, V.N.; Rudd, J.; Kousoulas, K.G. Two Sides to Every Story: Herpes Simplex Type-1 Viral Glycoproteins gB, gD, gH/gL, gK, and Cellular Receptors Function as Key Players in Membrane Fusion. Viruses 2021, 13, 1849. [Google Scholar] [CrossRef]
- Al Rumaih, Z.; Tuazon Kels, M.J.; Ng, E.; Pandey, P.; Pontejo, S.M.; Alejo, A.; Alcamí, A.; Chaudhri, G.; Karupiah, G. Poxvirus-Encoded TNF Receptor Homolog Dampens Inflammation and Protects from Uncontrolled Lung Pathology during Respiratory Infection. Proc. Natl. Acad. Sci. USA 2020, 117, 26885–26894. [Google Scholar] [CrossRef]
- Janeway, C.A., Jr.; Travers, P.; Walport, M.; Shlomchik, M.J. Induced Innate Responses to Infection. In Immunobiology: The Immune System in Health and Disease, 5th ed.; Garland Science: New York, NY, USA, 2001. [Google Scholar]
- Netea, M.G.; Kullberg, B.J.; Van der Meer, J.W.M. Circulating Cytokines as Mediators of Fever. Clin. Infect. Dis. 2000, 31, S178–S184. [Google Scholar] [CrossRef]
- Rahman, M.M.; Lucas, A.R.; McFadden, G. Viral TNF Inhibitors as Potential Therapeutics. In Madame Curie Bioscience Database [Internet]; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Rakus, K.; Ronsmans, M.; Forlenza, M.; Boutier, M.; Piazzon, M.C.; Jazowiecka-Rakus, J.; Gatherer, D.; Athanasiadis, A.; Farnir, F.; Davison, A.J.; et al. Conserved Fever Pathways across Vertebrates: A Herpesvirus Expressed Decoy TNF-α Receptor Delays Behavioral Fever in Fish. Cell Host Microbe 2017, 21, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Chai, W.; Qi, L.; Zhang, Y.; Hong, M.; Jin, L.; Li, L.; Yuan, J. Evaluation of Cyprinid Herpesvirus 2 Latency and Reactivation in Carassius gibel. Microorganisms 2020, 8, 445. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.-G.; Xie, J.; Chen, K.; Bing, X. Pathogenicity and Biological Characteristics of CyHV-2. Fish Pathol. 2015, 35, 85–93. [Google Scholar]
- Ouyang, P.; Zhou, Y.; Wang, K.; Geng, Y.; Lai, W.; Huang, X.; Chen, D.; Guo, H.; Fang, J.; Chen, Z.; et al. First Report of Cyprinid Herpesvirus 2 Outbreak in Cultured Gibel Carp, Carassius auratus Gibelio at Low Temperature. J. World Aquac. Soc. 2020, 51, 1208–1220. [Google Scholar] [CrossRef]
- Wen, J.; Xu, Y.; Su, M.; Lu, L.; Wang, H. Susceptibility of Goldfish to Cyprinid Herpesvirus 2 (CyHV-2) SH01 Isolated from Cultured Crucian Carp. Viruses 2021, 13, 1761. [Google Scholar] [CrossRef]
- Løkka, G.; Gamil, A.A.A.; Evensen, Ø.; Kortner, T.M. Establishment of an In Vitro Model to Study Viral Infections of the Fish Intestinal Epithelium. Cells 2023, 12, 1531. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NL-1 | NL-2 | NL-3 | |||||
|---|---|---|---|---|---|---|---|
| Number | Proportion | Number | Proportion | Number | Proportion | ||
| Variants Class | Indel | 13 | 2.2% | 27 | 3.7% | 23 | 3.1% |
| Substitution | 40 | 6.6% | 45 | 6.1% | 31 | 4.2% | |
| Insertion | 117 | 19.4% | 130 | 17.7% | 153 | 20.5% | |
| Deletion | 138 | 22.9% | 179 | 24.4% | 165 | 22.1% | |
| SNV | 295 | 48.9% | 353 | 48.1% | 374 | 50.1% | |
| Consequences | Stop gained | 0 | 0% | 0 | 0% | 1 | 0.1% |
| Frameshift * | 5 | 0.8% | 7 | 1% | 8 | 1.1% | |
| Inframe insertion | 29 | 4.8% | 33 | 4.5% | 41 | 5.5% | |
| Inframe deletion | 48 | 8% | 61 | 8.3% | 55 | 7.4% | |
| Protein altering | 2 | 0.3% | 5 | 0.7% | 5 | 0.7% | |
| Missenses | 93 | 15.4% | 118 | 16.1% | 102 | 13.7 | |
| Start retained | 1 | 0.2% | 0 | 0% | 2 | 0.3% | |
| Stop retained | 0 | 0% | 1 | 0.1% | 1 | 0.1% | |
| Synonymous | 106 | 17.6% | 125 | 17% | 124 | 16.6% | |
| Coding sequence | 1 | 0.2% | 1 | 0.1% | 2 | 0.3% | |
| Upstream gene | 306 | 50.7% | 358 | 48.8% | 390 | 52.3% | |
| Downstream gene | 9 | 1.5% | 22 | 3% | 11 | 1.5% | |
| Intergenic | 3 | 0.5% | 3 | 0.4% | 4 | 0.5% | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, B.; Sridhar, A.; Thiry, M.; Haenen, O.; Vanderplasschen, A.F.C.; Donohoe, O. Genomic and Phenotypic Characterization of a Novel Virulent Strain of Cyvirus cyprinidallo2 Originating from an Outbreak in The Netherlands. Viruses 2025, 17, 658. https://doi.org/10.3390/v17050658
He B, Sridhar A, Thiry M, Haenen O, Vanderplasschen AFC, Donohoe O. Genomic and Phenotypic Characterization of a Novel Virulent Strain of Cyvirus cyprinidallo2 Originating from an Outbreak in The Netherlands. Viruses. 2025; 17(5):658. https://doi.org/10.3390/v17050658
Chicago/Turabian StyleHe, Bo, Arun Sridhar, Marc Thiry, Olga Haenen, Alain F. C. Vanderplasschen, and Owen Donohoe. 2025. "Genomic and Phenotypic Characterization of a Novel Virulent Strain of Cyvirus cyprinidallo2 Originating from an Outbreak in The Netherlands" Viruses 17, no. 5: 658. https://doi.org/10.3390/v17050658
APA StyleHe, B., Sridhar, A., Thiry, M., Haenen, O., Vanderplasschen, A. F. C., & Donohoe, O. (2025). Genomic and Phenotypic Characterization of a Novel Virulent Strain of Cyvirus cyprinidallo2 Originating from an Outbreak in The Netherlands. Viruses, 17(5), 658. https://doi.org/10.3390/v17050658