
Reduced Binding between Omicron B.1.1.529 and the Human ACE2 Receptor in a Surrogate Virus Neutralization Test for SARS-CoV-2

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Serum Samples
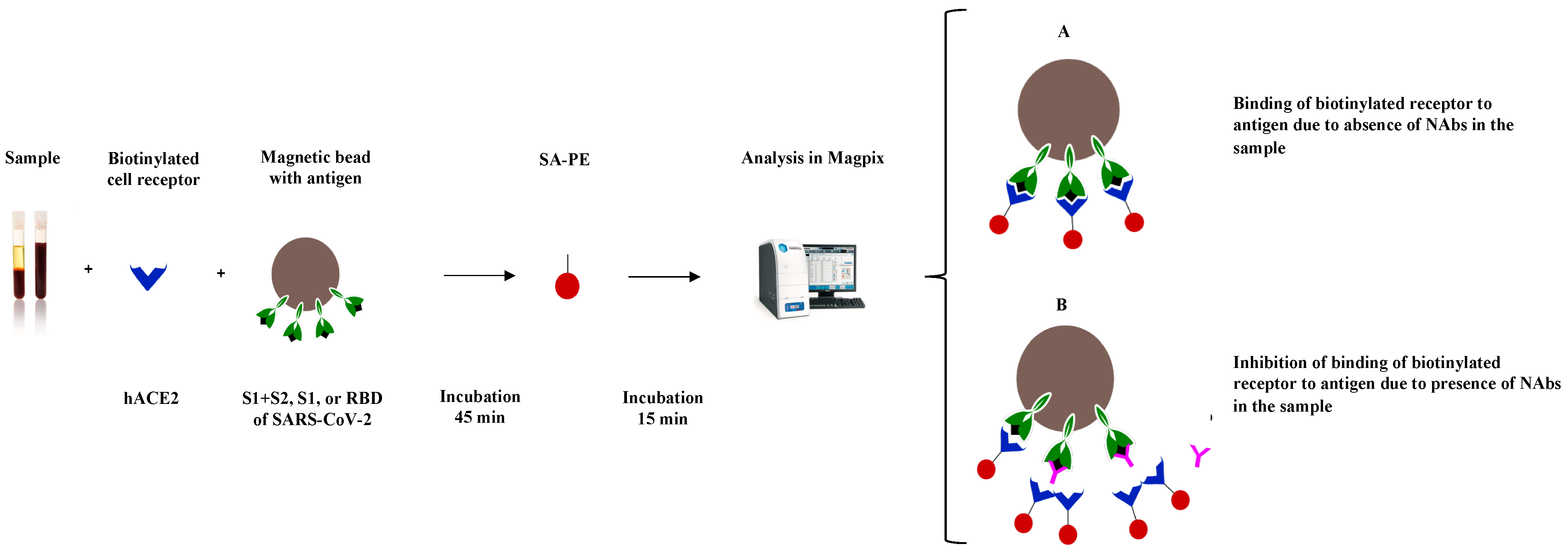
2.2. SARS-CoV-2 sVNT
2.2.1. Conjugation
2.2.2. Binding Inhibition
2.2.3. Assay Cut-Off
2.3. SARS-CoV-2 cVNT
2.3.1. Cells and Virus
2.3.2. Plaque Reduction Neutralization Test
2.4. Statistical Analyses and Data Visualization
3. Results
3.1. SARS-CoV-2 sVNT in Singleplex Format
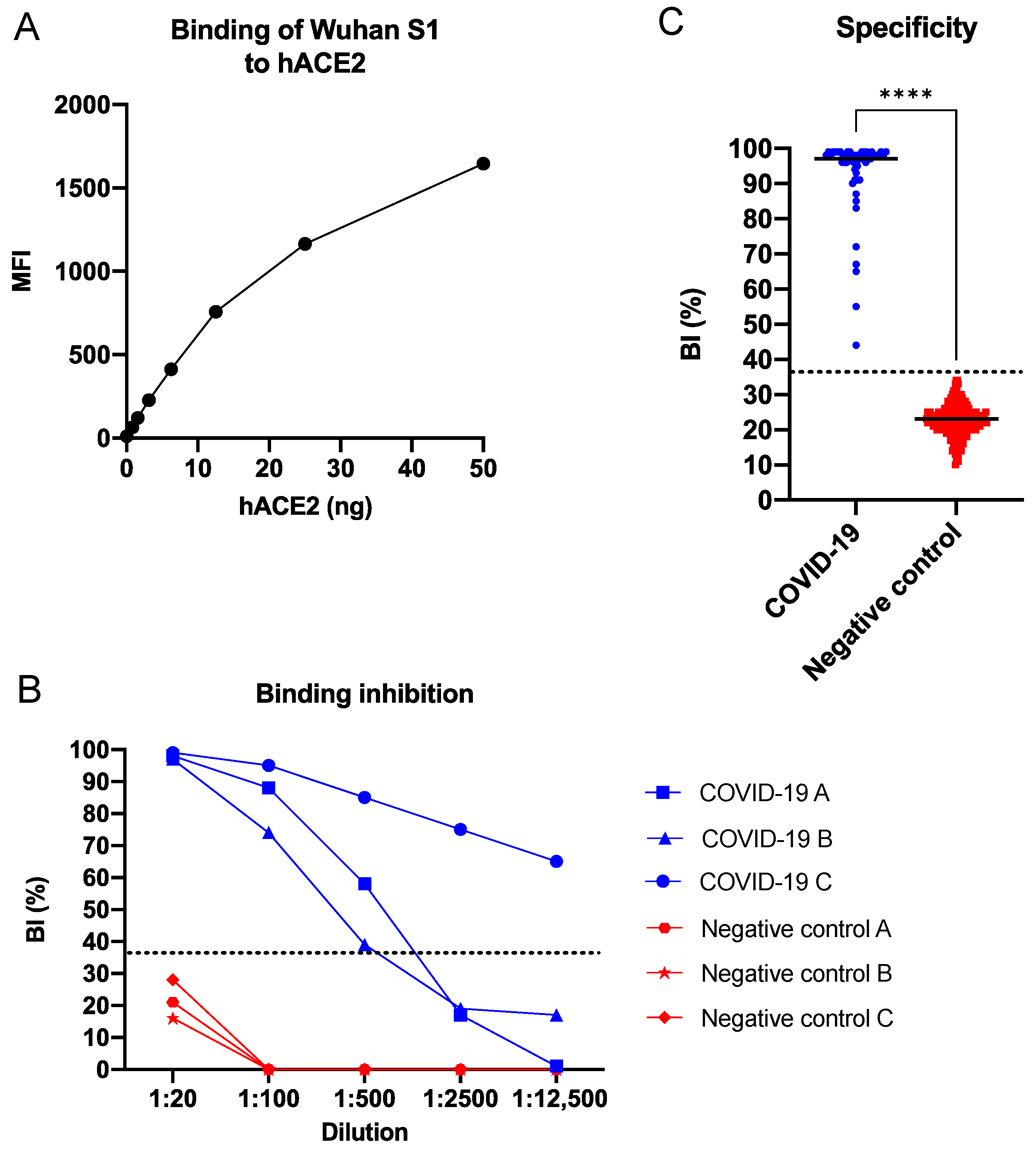
3.1.1. Cut-Off Determination
3.1.2. Binding Evaluation
3.1.3. Inter and Intra Assay Variability
3.2. SARS-CoV-2 sVNT in Multiplex Format
3.3. Correlation between Wuhan S1 SARS-CoV-2 sVNT (Singleplex Format) and cVNT
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, C.; Horby, P.W.; Hayden, F.G.; Gao, G.F. A novel coronavirus outbreak of global health concern. Lancet 2020, 395, 470–473. [Google Scholar] [CrossRef] [PubMed]
- Worldometer. COVID-19 Coronavirus Pandemic. Available online: https://www.worldometers.info/coronavirus/ (accessed on 24 April 2023).
- Satarker, S.; Nampoothiri, M. Structural proteins in severe acute respiratory syndrome coronavirus-2. Arch. Med. Res. 2020, 51, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Li, F. Structure, function, and evolution of coronavirus spike proteins. Annu. Rev. Virol. 2016, 3, 237–261. [Google Scholar] [CrossRef] [PubMed]
- Stamatatos, L.; Czartoski, J.; Wan, Y.H.; Homad, L.J.; Rubin, V.; Glantz, H.; Neradilek, M.; Seydoux, E.; Jennewein, M.F.; MacCamy, A.J.; et al. mRNA vaccination boosts cross-variant neutralizing antibodies elicited by SARS-CoV-2 infection. Science 2021, 372, 1413–1418. [Google Scholar] [CrossRef] [PubMed]
- CDC. SARS-CoV-2 Variant Classifications and Definitions. Available online: https://www.cdc.gov/coronavirus/2019-ncov/variants/variant-classifications.html#anchor_1632154493691 (accessed on 9 August 2022).
- WHO. Enhancing Response to Omicron SARS-CoV-2 Variant: Technical Brief and Priority Actions for Member States; Update #5: 7 January 2022; World Health Organization: Geneva, Switzerland, 2022; Available online: https://www.who.int/docs/default-source/coronaviruse/2022-01-07-global-technical-brief-and-priority-action-on-omicron---corr2.pdf?sfvrsn=918b09d_20 (accessed on 9 August 2022).
- Wu, L.; Zhou, L.; Mo, M.; Liu, T.; Wu, C.; Gong, C.; Lu, K.; Gong, L.; Zhu, W.; Xu, Z. SARS-CoV-2 Omicron RBD shows weaker binding affinity than the currently dominant Delta variant to human ACE2. Signal Transduct. Target. Ther. 2022, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Mögling, R.; Colavita, F.; Reimerink, J.; Melidou, A.; Leitmeyer, K.; Keramarou, M.; Lapa, D.; Francalancia, M.; Murk, J.-L.; Vossen, A.; et al. External quality assessment of SARS-CoV-2 serology in European expert laboratories, April 2021. Eurosurveillanve 2022, 27, 2101057. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, T.; Kolstad, L.; Lindahl, J.F.; Albinsson, B.; Bergqvist, A.; Rönnberg, B.; Lundkvist, Å. Diagnostic potential of a Luminex-based coronavirus disease 2019 suspension immunoassay (COVID-19 SIA) for the detection of antibodies against SARS-CoV-2. Viruses 2021, 13, 993. [Google Scholar] [CrossRef] [PubMed]
- Akaberi, D.; Krambrich, J.; Ling, J.; Luni, C.; Hedenstierna, G.; Järhult, J.D.; Lennerstrand, J.; Lundkvist, Å. Mitigation of the replication of SARS-CoV-2 by nitric oxide in vitro. Redox. Biol. 2020, 37, 101734. [Google Scholar] [CrossRef] [PubMed]
- Klasse, P.J. Neutralization of virus infectivity by antibodies: Old problems in new perspectives. Adv. Biol. 2014, 2014, 157895. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.; Bertoglio, F.; Steinke, S.; Heine, P.A.; Ynga-Durand, M.A.; Maass, H.; Sammartino, J.C.; Cassaniti, I.; Zuo, F.; Du, L.; et al. Human serum from SARS-CoV-2-vaccinated and COVID-19 patients shows reduced binding to the RBD of SARS-CoV-2 Omicron variant. BMC Med. 2022, 20, 102. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Liu, P.; Zhang, Y.; Du, T.; Zhou, Y.; Lu, S.; Peng, X. Characteristic analysis of Omicron-included SARS-CoV-2 variants of concern. MedComm 2022, 3, e129. [Google Scholar] [CrossRef] [PubMed]
- Glocker, M.O.; Opuni, K.F.M.; Thiesen, H.J. From free binding energy calculations of SARS-CoV-2-receptor interactions to cellular immune responses. Medicina 2022, 58, 226. [Google Scholar] [CrossRef] [PubMed]
- Lupala, C.S.; Ye, Y.; Chen, H.; Su, X.D.; Liu, H. Mutations on RBD of SARS-CoV-2 Omicron variant result in stronger binding to human ACE2 receptor. Biochem. Biophys. Res. Commun. 2022, 590, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Cameroni, E.; Bowen, J.E.; Rosen, L.E.; Saliba, C.; Zepeda, S.K.; Culap, K.; Pinto, D.; VanBlargan, L.A.; De Marco, A.; di Iulio, J.; et al. Broadly neutralizing antibodies overcome SARS-CoV-2 Omicron antigenic shift. Nature 2022, 602, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Meng, B.; Abdullahi, A.; Ferreira, I.; Goonawardane, N.; Saito, A.; Kimura, I.; Yamasoba, D.; Gerber, P.P.; Fatihi, S.; Rathore, S.; et al. Altered TMPRSS2 usage by SARS-CoV-2 Omicron impacts infectivity and fusogenicity. Nature 2022, 603, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Murugan, N.A.; Srivastava, V. Improved binding affinity of Omicron’s spike protein for the human angiotensin-converting enzyme 2 receptor is the key behind its increased virulence. Int. J. Mol. Sci. 2022, 23, 3409. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variant (Lineage) | Company | Product No. | Expression System | Purity a | Amino Acids (aa) | No. of aa Changes b | No. of aa Changes in RBD c |
|---|---|---|---|---|---|---|---|
| Wuhan S1 (Wuhan-Hu-1) | SinoBiological | 40591-V08H | HEK293 | ≥92% | 16–685 | ||
| Delta S1 (B.1.617.2) | SinoBiological | 40591-V08H23 | HEK293 | 92.7% | 1–685 | 8 | 2 e |
| Omicron S1 (B.1.1.529) | SinoBiological | 40591-V08H41 | HEK293 | 97.0% | 1–685 | 28 | 15 f |
| Omicron S1 (B.1.1.529) | Abcam | ab290828 | HEK293 | >95% | 16–671 | 29 | 15 f |
| Omicron S1 (B.1.1.529) | ACROBiosystems | S1N-C52Ha | HEK293 | >95% | 16–685 | 28 | 15 f |
| Omicron S1 (B.1.1.529) | Native Antigen | REC32006 | HEK293 | >95% | 16–671 | 29 | 15 f |
| Omicron S1 + S2 (B.1.1.529) d | SinoBiological | 40589-V08H26 | HEK293 | 94% | 16–1234 | 40 | 15 f |
| Omicron RBD (B.1.1.529) | SinoBiological | 40592-V08H121 | HEK293 | 97.8% | 319–541 | 15 f | |
| Wuhan N (Wuhan-Hu-1) | SinoBiological | 40588-V08B | BVI | 96% | 1–419 |
| cVNT (Cut-Off: 50%) | cVNT (Cut-Off: 80%) | ||||
|---|---|---|---|---|---|
| COVID-19 patients (n = 54) | Pos | Neg | Pos | Neg | |
| sVNT (Cut-off: 36%) | Pos | 54 | 0 | 54 | 0 |
| Neg | 0 | 0 | 0 | 0 | |
| Blood donors (n = 18) | Pos | Neg | Pos | Neg | |
| sVNT (Cut-off: 36%) | Pos | 0 | 0 | 0 | 0 |
| Neg | 0 | 18 | 0 | 18 | |
| cVNT | sVNT | ||
|---|---|---|---|
| ID | Control Serum/Plasma | Inhibition (%) | Inhibition (%) |
| (1:20) | (1:20) | ||
| EQA A | SARS-CoV-2 Abs (Hospitalized patient) | 100 * | 91 |
| EQA B | SARS-CoV-2 Abs (Mild infection) | 100 | 94 |
| EQA C | Negative pre-pandemic sera (Other human CoV) | 0 | 0 |
| EQA D | SARS-CoV-2 Abs (Hospitalized patient) | 100 | 97 |
| EQA E | Acute EBV/CMV sera (Other human CoV) | 0 | 0 |
| EQA F | Negative pre-pandemic sera (Other human CoV) | 0 | 0 |
| EQA G | SARS-CoV-2 Abs (Mild infection) | 100 | 80 |
| EQA H | SARS-CoV-2 Abs (Hospitalized patient) | 100 | 89 |
| JCR 017 | SARS-CoV-2 Abs (Standard) | 98 | 80 |
| JCR 018 | SARS-CoV-2 Abs (Standard) | 92 | 87 |
| 20/136 | SARS-CoV-2 Abs (Standard) | 100 | 96 |
| 20/140 (20/268) | SARS-CoV-2 Abs + NAbs (Lowest Ab levels) | 53 | 54 |
| 20/142 (20/268) | Negative human sera | 6 | 34 |
| 20/144 (20/268) | SARS-CoV-2 Abs + NAbs (Low Ab levels) | 92 | 50 |
| 20/148 (20/268) | SARS-CoV-2 Abs + NAbs (Mid Ab levels) | 97 | 86 |
| 20/150 (20/268) | SARS-CoV-2 Abs + NAbs (High Ab levels) | 100 | 96 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoffman, T.; Kolstad, L.; Akaberi, D.; Järhult, J.D.; Rönnberg, B.; Lundkvist, Å. Reduced Binding between Omicron B.1.1.529 and the Human ACE2 Receptor in a Surrogate Virus Neutralization Test for SARS-CoV-2. Viruses 2023, 15, 1280. https://doi.org/10.3390/v15061280
Hoffman T, Kolstad L, Akaberi D, Järhult JD, Rönnberg B, Lundkvist Å. Reduced Binding between Omicron B.1.1.529 and the Human ACE2 Receptor in a Surrogate Virus Neutralization Test for SARS-CoV-2. Viruses. 2023; 15(6):1280. https://doi.org/10.3390/v15061280
Chicago/Turabian StyleHoffman, Tove, Linda Kolstad, Dario Akaberi, Josef D. Järhult, Bengt Rönnberg, and Åke Lundkvist. 2023. "Reduced Binding between Omicron B.1.1.529 and the Human ACE2 Receptor in a Surrogate Virus Neutralization Test for SARS-CoV-2" Viruses 15, no. 6: 1280. https://doi.org/10.3390/v15061280
APA StyleHoffman, T., Kolstad, L., Akaberi, D., Järhult, J. D., Rönnberg, B., & Lundkvist, Å. (2023). Reduced Binding between Omicron B.1.1.529 and the Human ACE2 Receptor in a Surrogate Virus Neutralization Test for SARS-CoV-2. Viruses, 15(6), 1280. https://doi.org/10.3390/v15061280