
Embryonic and Neonatal Mouse Cochleae Are Susceptible to Zika Virus Infection



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Collection of Embryos and Neonatal Pups
2.3. Cochlear Cultures
2.4. Histology
2.5. Microscopy and Evaluating Infection
2.6. Image Analysis of Cell Death
3. Results
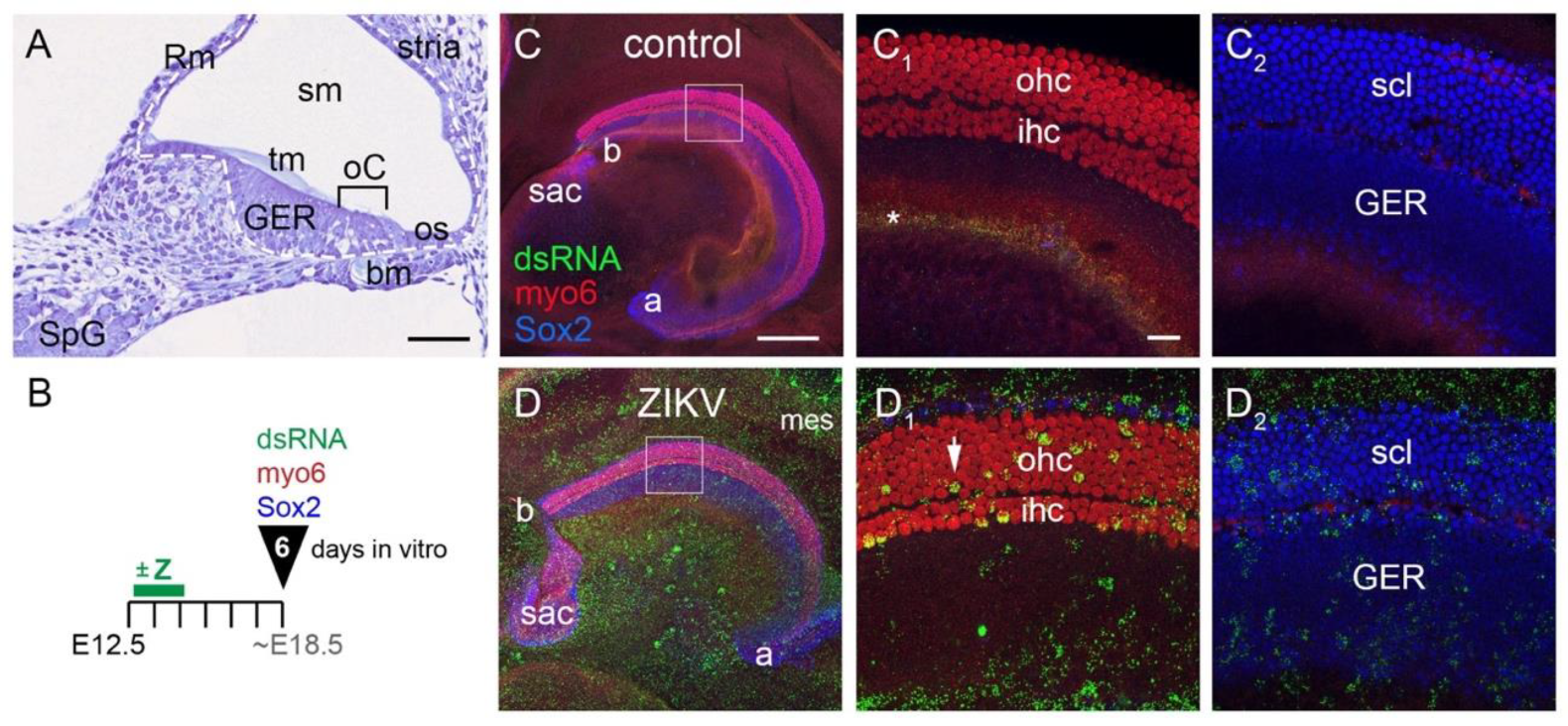
3.1. ZIKV Infects Both Prosensory (Mitotic) Cells and Post-Mitotic Sensory Cells in Cochlear Organ Cultures
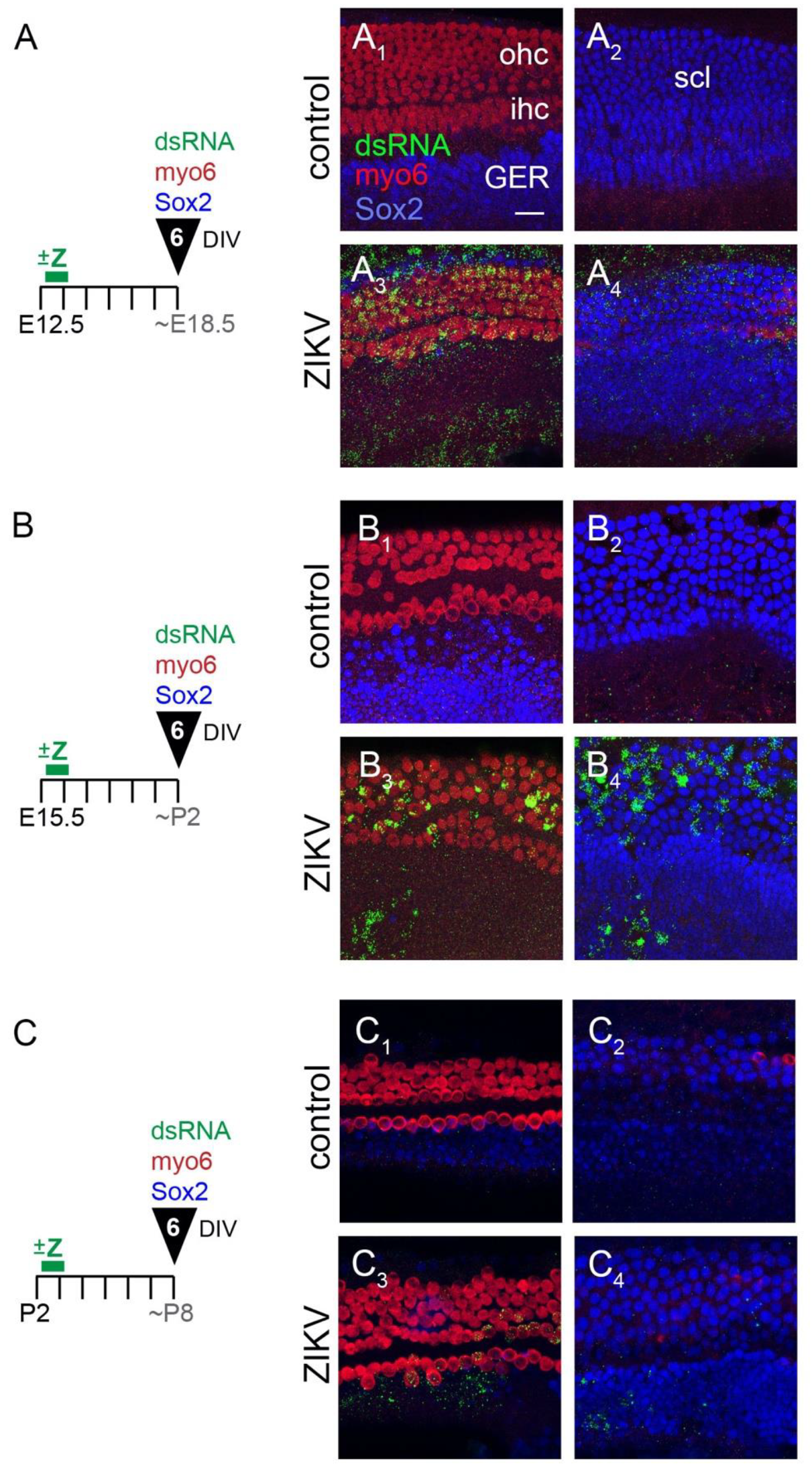
3.1.1. E12.5 Plus 6 Days In Vitro
3.1.2. E15.5 Plus 6 Days In Vitro
3.1.3. P2 Plus 6 Days In Vitro
3.2. ZIKV Infection Does Not Induce Extensive Cell Death in the Cochlear Epithelium
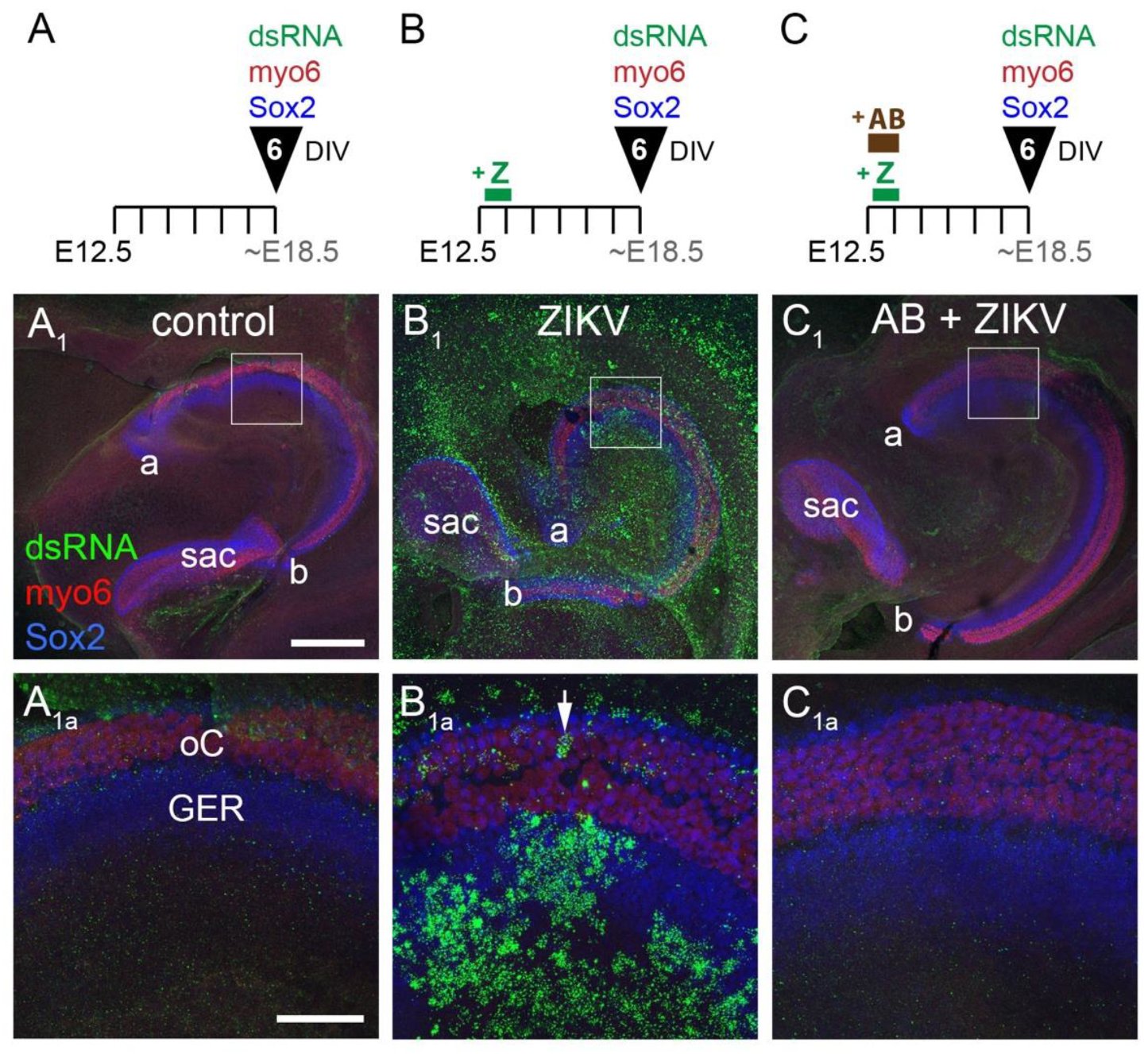
3.3. ZIKV Infection of the Cochlea Is Blocked by Preincubation with a Neutralizing Antibody
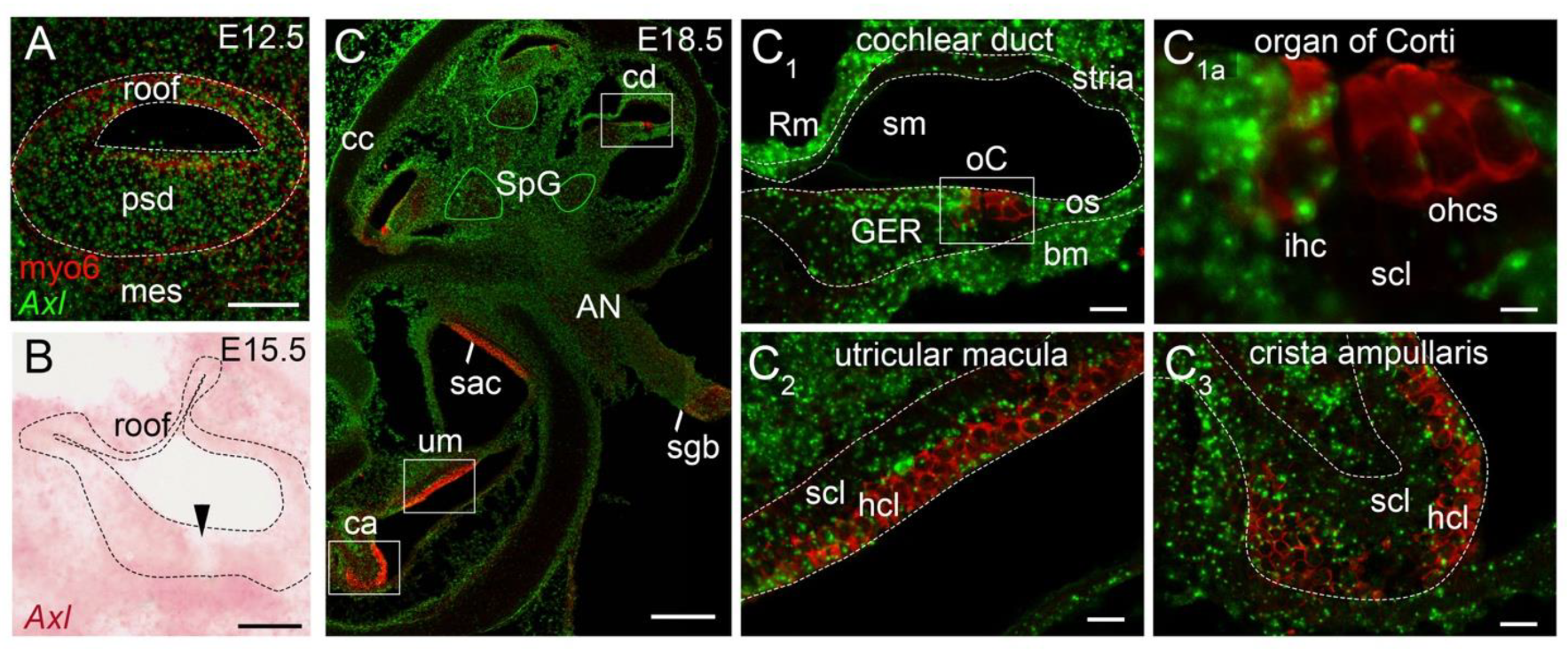
3.4. Axl Transcripts Are Expressed in Many Tissues of the Developing Inner Ear
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Brasil, P.; Pereira, J.P., Jr.; Moreira, M.E.; Ribeiro Nogueira, R.M.; Damasceno, L.; Wakimoto, M.; Rabello, R.S.; Valderramos, S.G.; Halai, U.A.; Salles, T.S.; et al. Zika Virus Infection in Pregnant Women in Rio de Janeiro. N. Engl. J. Med. 2016, 375, 2321–2334. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.A.; Staples, J.E.; Dobyns, W.B.; Pessoa, A.; Ventura, C.V.; Fonseca, E.B.; Ribeiro, E.M.; Ventura, L.O.; Neto, N.N.; Arena, J.F.; et al. Characterizing the Pattern of Anomalies in Congenital Zika Syndrome for Pediatric Clinicians. JAMA Pediatr. 2017, 171, 288–295. [Google Scholar] [CrossRef] [Green Version]
- Nielsen-Saines, K.; Brasil, P.; Kerin, T.; Vasconcelos, Z.; Gabaglia, C.R.; Damasceno, L.; Pone, M.; de Abreu Carvalho, L.M.; Pone, S.M.; Zin, A.A.; et al. Delayed childhood neurodevelopment and neurosensory alterations in the second year of life in a prospective cohort of ZIKV-exposed children. Nat. Med. 2019, 25, 1213–1217. [Google Scholar] [CrossRef]
- Barbosa, M.H.M.; Magalhaes-Barbosa, M.C.; Robaina, J.R.; Prata-Barbosa, A.; Lima, M.; Cunha, A. Auditory findings associated with Zika virus infection: An integrative review. Braz. J. Otorhinolaryngol. 2019, 85, 642–663. [Google Scholar] [CrossRef]
- Escobar-Ipuz, F.A.; Soria-Bretones, C.; Garcia-Jimenez, M.A.; Cueto, E.M.; Torres Aranda, A.M.; Sotos, J.M. Early detection of neonatal hearing loss by otoacoustic emissions and auditory brainstem response over 10 years of experience. Int. J. Pediatr. Otorhinolaryngol. 2019, 127, 109647. [Google Scholar] [CrossRef]
- Thompson, D.C.; McPhillips, H.; Davis, R.L.; Lieu, T.L.; Homer, C.J.; Helfand, M. Universal newborn hearing screening: Summary of evidence. JAMA 2001, 286, 2000–2010. [Google Scholar] [CrossRef] [Green Version]
- Bakhos, D.; Marx, M.; Villeneuve, A.; Lescanne, E.; Kim, S.; Robier, A. Electrophysiological exploration of hearing. Eur. Ann. Otorhinolaryngol. Head Neck Dis. 2017, 134, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Guinan, J.J., Jr.; Salt, A.; Cheatham, M.A. Progress in cochlear physiology after Bekesy. Hear Res. 2012, 293, 12–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thawani, A.; Sammudin, N.H.; Reygaerts, H.S.; Wozniak, A.N.; Munnamalai, V.; Kuhn, R.J.; Fekete, D.M. Zika virus can directly infect and damage the auditory and vestibular components of the embryonic chicken inner ear. Dev. Dyn. 2020, 249, 867–883. [Google Scholar] [CrossRef]
- Yee, K.T.; Neupane, B.; Bai, F.; Vetter, D.E. Zika virus infection causes widespread damage to the inner ear. Hear Res. 2020, 395, 108000. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Hammack, C.; Ogden, S.C.; Wen, Z.; Qian, X.; Li, Y.; Yao, B.; Shin, J.; Zhang, F.; Lee, E.M.; et al. Zika Virus Infects Human Cortical Neural Progenitors and Attenuates Their Growth. Cell Stem Cell 2016, 18, 587–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapparapu, G.; Fernandez, E.; Kose, N.; Bin, C.; Fox, J.M.; Bombardi, R.G.; Zhao, H.; Nelson, C.A.; Bryan, A.L.; Barnes, T.; et al. Neutralizing human antibodies prevent Zika virus replication and fetal disease in mice. Nature 2016, 540, 443–447. [Google Scholar] [CrossRef] [Green Version]
- Almansour, I.; Alfares, R.; Aljofi, H. Large-scale analysis of B-cell epitopes of envelope: Implications for Zika vaccine and immunotherapeutic development. F1000Research 2018, 7, 1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erasmus, J.H.; Archer, J.; Fuerte-Stone, J.; Khandhar, A.P.; Voigt, E.; Granger, B.; Bombardi, R.G.; Govero, J.; Tan, Q.; Durnell, L.A.; et al. Intramuscular Delivery of Replicon RNA Encoding ZIKV-117 Human Monoclonal Antibody Protects against Zika Virus Infection. Mol. Ther. Methods Clin. Dev. 2020, 18, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Robbiani, D.F.; Bozzacco, L.; Keeffe, J.R.; Khouri, R.; Olsen, P.C.; Gazumyan, A.; Schaefer-Babajew, D.; Avila-Rios, S.; Nogueira, L.; Patel, R.; et al. Recurrent Potent Human Neutralizing Antibodies to Zika Virus in Brazil and Mexico. Cell 2017, 169, 597–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, S.; Zhang, H.; Liang, Z.; Yang, X.; Cao, R. AXL, an Important Host Factor for DENV and ZIKV Replication. Front. Cell Infect. Microbiol. 2021, 11, 575346. [Google Scholar] [CrossRef]
- Munnamalai, V.; Fekete, D.M. Organotypic Culture of the Mouse Cochlea from Embryonic Day 12 to the Neonate. Methods Mol. Biol. 2016, 1427, 293–303. [Google Scholar]
- Munnamalai, V.; Fekete, D.M. Notch-Wnt-Bmp crosstalk regulates radial patterning in the mouse cochlea in a spatiotemporal manner. Development 2016, 143, 4003–4015. [Google Scholar] [CrossRef] [Green Version]
- Ruben, R.J. Development of the inner ear of the mouse: A radioautographic study of terminal mitosis. Acta Otolaryngol. Suppl. 1967, 220, 4–44. [Google Scholar]
- Chen, P.; Segil, N. p27(Kip1) links cell proliferation to morphogenesis in the developing organ of Corti. Development 1999, 126, 1581–1590. [Google Scholar] [CrossRef]
- Souza, B.S.; Sampaio, G.L.; Pereira, C.S.; Campos, G.S.; Sardi, S.I.; Freitas, L.A.; Figueira, C.P.; Paredes, B.D.; Nonaka, C.K.; Azevedo, C.M.; et al. Zika virus infection induces mitosis abnormalities and apoptotic cell death of human neural progenitor cells. Sci. Rep. 2016, 6, 39775. [Google Scholar] [CrossRef]
- Li, H.; Saucedo-Cuevas, L.; Regla-Nava, J.A.; Chai, G.; Sheets, N.; Tang, W.; Terskikh, A.V.; Shresta, S.; Gleeson, J.G. Zika Virus Infects Neural Progenitors in the Adult Mouse Brain and Alters Proliferation. Cell Stem Cell 2016, 19, 593–598. [Google Scholar] [CrossRef] [Green Version]
- Sevvana, M.; Rogers, T.F.; Miller, A.S.; Long, F.; Klose, T.; Beutler, N.; Lai, Y.C.; Parren, M.; Walker, L.M.; Buda, G.; et al. Structural Basis of Zika Virus Specific Neutralization in Subsequent Flavivirus Infections. Viruses 2020, 12, 1346. [Google Scholar] [CrossRef] [PubMed]
- Hasan, S.S.; Miller, A.; Sapparapu, G.; Fernandez, E.; Klose, T.; Long, F.; Fokine, A.; Porta, J.C.; Jiang, W.; Diamond, M.S.; et al. A human antibody against Zika virus crosslinks the E protein to prevent infection. Nat. Commun. 2017, 8, 14722. [Google Scholar] [CrossRef]
- Miner, J.J.; Diamond, M.S. Understanding How Zika Virus Enters and Infects Neural Target Cells. Cell Stem Cell 2016, 18, 559–560. [Google Scholar] [CrossRef] [PubMed]
- Nowakowski, T.J.; Pollen, A.A.; Di Lullo, E.; Sandoval-Espinosa, C.; Bershteyn, M.; Kriegstein, A.R. Expression Analysis Highlights AXL as a Candidate Zika Virus Entry Receptor in Neural Stem Cells. Cell Stem Cell 2016, 18, 591–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meertens, L.; Labeau, A.; Dejarnac, O.; Cipriani, S.; Sinigaglia, L.; Bonnet-Madin, L.; Le Charpentier, T.; Hafirassou, M.L.; Zamborlini, A.; Cao-Lormeau, V.M.; et al. Axl Mediates ZIKA Virus Entry in Human Glial Cells and Modulates Innate Immune Responses. Cell Rep. 2017, 18, 324–333. [Google Scholar] [CrossRef]
- Hamel, R.; Dejarnac, O.; Wichit, S.; Ekchariyawat, P.; Neyret, A.; Luplertlop, N.; Perera-Lecoin, M.; Surasombatpattana, P.; Talignani, L.; Thomas, F.; et al. Biology of Zika Virus Infection in Human Skin Cells. J. Virol. 2015, 89, 8880–8896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharyya, S.; Zagorska, A.; Lew, E.D.; Shrestha, B.; Rothlin, C.V.; Naughton, J.; Diamond, M.S.; Lemke, G.; Young, J.A. Enveloped viruses disable innate immune responses in dendritic cells by direct activation of TAM receptors. Cell Host Microbe 2013, 14, 136–147. [Google Scholar] [CrossRef] [Green Version]
- Lemke, G.; Rothlin, C.V. Immunobiology of the TAM receptors. Nat. Rev. Immunol. 2008, 8, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Orvis, J.; Gottfried, B.; Kancherla, J.; Adkins, R.S.; Song, Y.; Dror, A.A.; O’lley, D.; Rose, K.; Chrysostomou, E.; Kelly, M.C.; et al. gEAR: Gene Expression Analysis Resource portal for community-driven, multi-omic data exploration. Nat. Methods 2021, 18, 843–844. [Google Scholar] [CrossRef]
- Elkon, R.; Milon, B.; Morrison, L.; Shah, M.; Vijayakumar, S.; Racherla, M.; Leitch, C.C.; Silipino, L.; Hadi, S.; Weiss-Gayet, M.; et al. RFX transcription factors are essential for hearing in mice. Nat. Commun. 2015, 6, 8549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Chen, L.; Giffen, K.P.; Stringham, S.T.; Li, Y.; Judge, P.D.; Beisel, K.W.; He, D.Z.Z. Cell-Specific Transcriptome Analysis Shows That Adult Pillar and Deiters’ Cells Express Genes Encoding Machinery for Specializations of Cochlear Hair Cells. Front. Mol. Neurosci. 2018, 11, 356. [Google Scholar] [CrossRef] [PubMed]
- Kolla, L.; Kelly, M.C.; Mann, Z.F.; Anaya-Rocha, A.; Ellis, K.; Lemons, A.; Palermo, A.T.; So, K.S.; Mays, J.C.; Orvis, J.; et al. Characterization of the development of the mouse cochlear epithelium at the single cell level. Nat. Commun. 2020, 11, 2389. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Chen, J.; Yang, J. Autophagy precedes apoptosis during degeneration of the Kolliker’s organ in the development of rat cochlea. Eur. J. Histochem. 2019, 63, 3025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamiya, K.; Takahashi, K.; Kitamura, K.; Momoi, T.; Yoshikawa, Y. Mitosis and apoptosis in postnatal auditory system of the C3H/He strain. Brain Res. 2001, 901, 296–302. [Google Scholar] [CrossRef]
- Locher, H.; Frijns, J.H.; van Iperen, L.; de Groot, J.C.; Huisman, M.A.; de Chuva Sousa Lopes, S.M. Neurosensory development and cell fate determination in the human cochlea. Neural. Dev. 2013, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Pujol, R.; Lavigne-Rebillard, M. Early stages of innervation and sensory cell differentiation in the human fetal organ of Corti. Acta Otolaryngol. Suppl. 1985, 423, 43–50. [Google Scholar] [CrossRef]
- Leal, M.C.; Muniz, L.F.; Ferreira, T.S.; Santos, C.M.; Almeida, L.C.; Van Der Linden, V.; Ramos, R.C.; Rodrigues, L.C.; Neto, S.S. Hearing Loss in Infants with Microcephaly and Evidence of Congenital Zika Virus Infection-Brazil, November 2015–May 2016. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 917–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thawani, A.; Sirohi, D.; Kuhn, R.J.; Fekete, D.M. Zika Virus Can Strongly Infect and Disrupt Secondary Organizers in the Ventricular Zone of the Embryonic Chicken Brain. Cell Rep. 2018, 23, 692–700. [Google Scholar] [CrossRef] [Green Version]
- Meertens, L.; Carnec, X.; Lecoin, M.P.; Ramdasi, R.; Guivel-Benhassine, F.; Lew, E.; Lemke, G.; Schwartz, O.; Amara, A. The TIM and TAM families of phosphatidylserine receptors mediate dengue virus entry. Cell Host Microbe 2012, 12, 544–557. [Google Scholar] [CrossRef] [Green Version]
- Wells, M.F.; Salick, M.R.; Wiskow, O.; Ho, D.J.; Worringer, K.A.; Ihry, R.J.; Kommineni, S.; Bilican, B.; Klim, J.R.; Hill, E.J.; et al. Genetic Ablation of AXL Does Not Protect Human Neural Progenitor Cells and Cerebral Organoids from Zika Virus Infection. Cell Stem Cell 2016, 19, 703–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govero, J.; Esakky, P.; Scheaffer, S.M.; Fernandez, E.; Drury, A.; Platt, D.J.; Gorman, M.J.; Richner, J.M.; Caine, E.A.; Salazar, V.; et al. Zika virus infection damages the testes in mice. Nature 2016, 540, 438–442. [Google Scholar] [CrossRef] [Green Version]
- Hastings, A.K.; Yockey, L.J.; Jagger, B.W.; Hwang, J.; Uraki, R.; Gaitsch, H.F.; Parnell, L.A.; Cao, B.; Mysorekar, I.U.; Rothlin, C.V.; et al. TAM Receptors Are Not Required for Zika Virus Infection in Mice. Cell Rep. 2017, 19, 558–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.Y.; Wang, Z.; Zhen, Z.D.; Feng, K.H.; Guo, J.; Gao, N.; Fan, D.Y.; Han, D.S.; Wang, P.G.; An, J. Axl is not an indispensable factor for Zika virus infection in mice. J. Gen. Virol. 2017, 98, 2061–2068. [Google Scholar] [CrossRef]
- Pierson, T.C.; Diamond, M.S. The continued threat of emerging flaviviruses. Nat. Microbiol 2020, 5, 796–812. [Google Scholar] [CrossRef] [PubMed]
- Renauld, J.M.; Basch, M.L. Congenital Deafness and Recent Advances Towards Restoring Hearing Loss. Curr. Protoc. 2021, 1, e76. [Google Scholar] [CrossRef] [PubMed]
- Ficenec, S.C.; Schieffelin, J.S.; Emmett, S.D. A Review of Hearing Loss Associated with Zika, Ebola, and Lassa Fever. Am. J. Trop. Med. Hyg. 2019, 101, 484–490. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munnamalai, V.; Sammudin, N.H.; Young, C.A.; Thawani, A.; Kuhn, R.J.; Fekete, D.M. Embryonic and Neonatal Mouse Cochleae Are Susceptible to Zika Virus Infection. Viruses 2021, 13, 1823. https://doi.org/10.3390/v13091823
Munnamalai V, Sammudin NH, Young CA, Thawani A, Kuhn RJ, Fekete DM. Embryonic and Neonatal Mouse Cochleae Are Susceptible to Zika Virus Infection. Viruses. 2021; 13(9):1823. https://doi.org/10.3390/v13091823
Chicago/Turabian StyleMunnamalai, Vidhya, Nabilah H. Sammudin, Caryl A. Young, Ankita Thawani, Richard J. Kuhn, and Donna M. Fekete. 2021. "Embryonic and Neonatal Mouse Cochleae Are Susceptible to Zika Virus Infection" Viruses 13, no. 9: 1823. https://doi.org/10.3390/v13091823
APA StyleMunnamalai, V., Sammudin, N. H., Young, C. A., Thawani, A., Kuhn, R. J., & Fekete, D. M. (2021). Embryonic and Neonatal Mouse Cochleae Are Susceptible to Zika Virus Infection. Viruses, 13(9), 1823. https://doi.org/10.3390/v13091823