Virus-Induced Flowering by Apple Latent Spherical Virus Vector: Effective Use to Accelerate Breeding of Grapevine

Abstract
1. Introduction
2. Materials and Methods
2.1. Plants
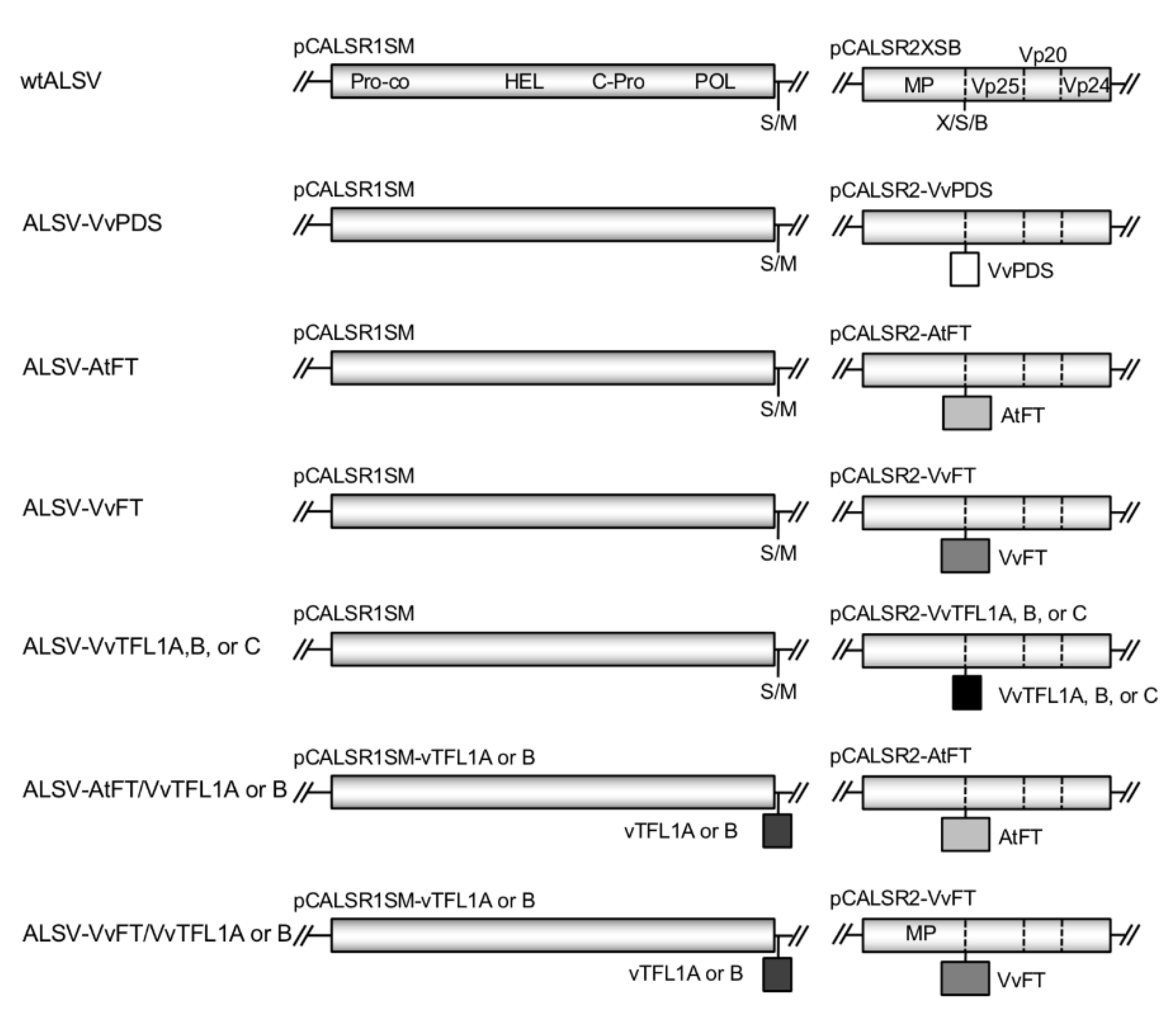
2.2. Construction of ALSV Vectors
2.3. Agro-Inoculation
2.4. Preparation of RNAs from Infected Leaves
2.5. ALSV Inoculation by Particle Bombardment
2.6. RT-PCR for Detection of Virus Infection
2.7. Quantitative RT-PCR
2.8. Tissue-Blot Hybridization
2.9. In Situ Hybridization
2.10. Analysis of Sugar, Acid, and Anthocyanin Content in Grape Berries
2.11. High-Temperature Treatment for Virus Elimination
3. Results
3.1. Construction of ALSV Vectors for Grapevine
3.2. ALSV Vector Inoculation Conditions for Grapevine Seedlings and In Vitro Cultures
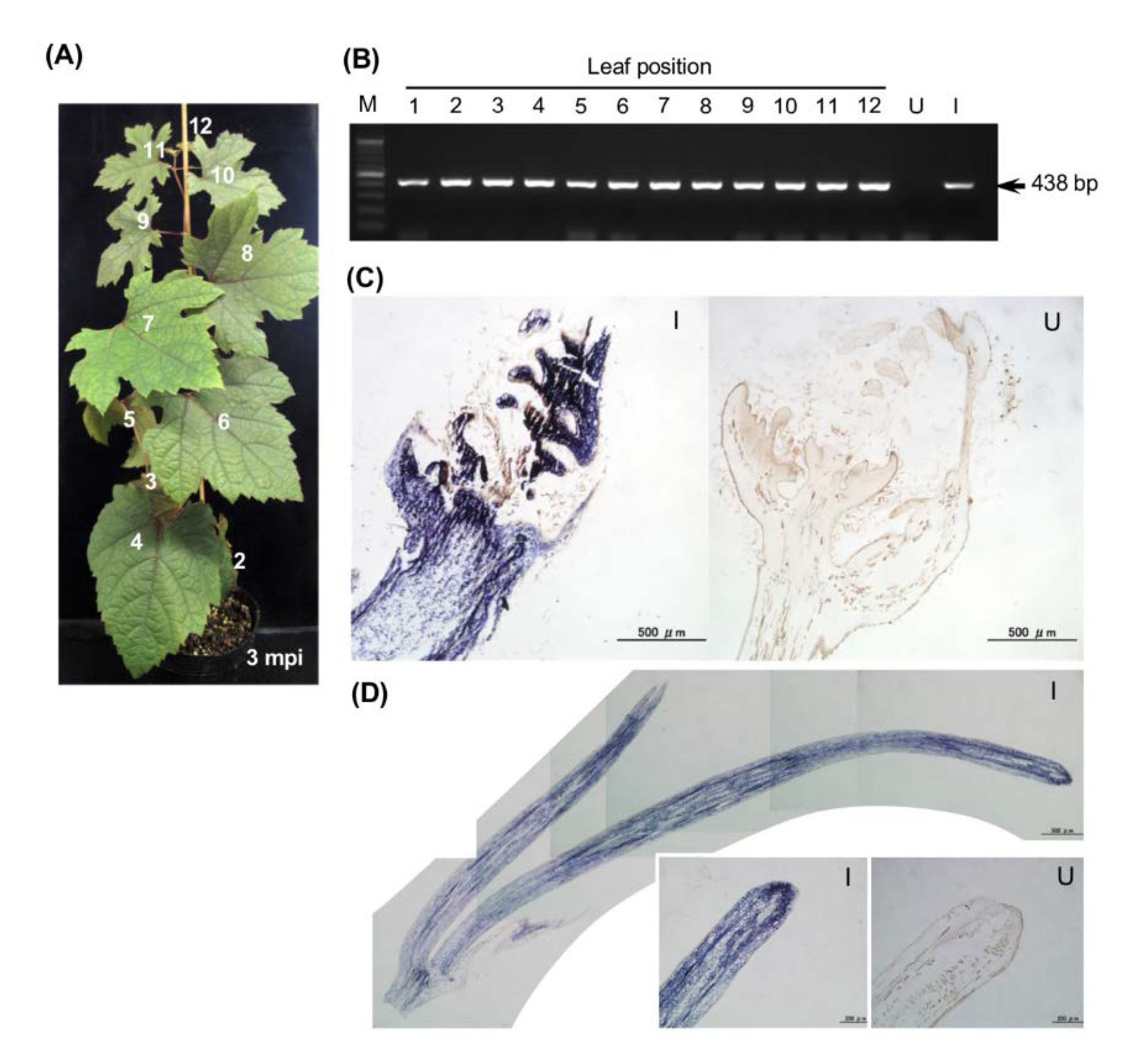
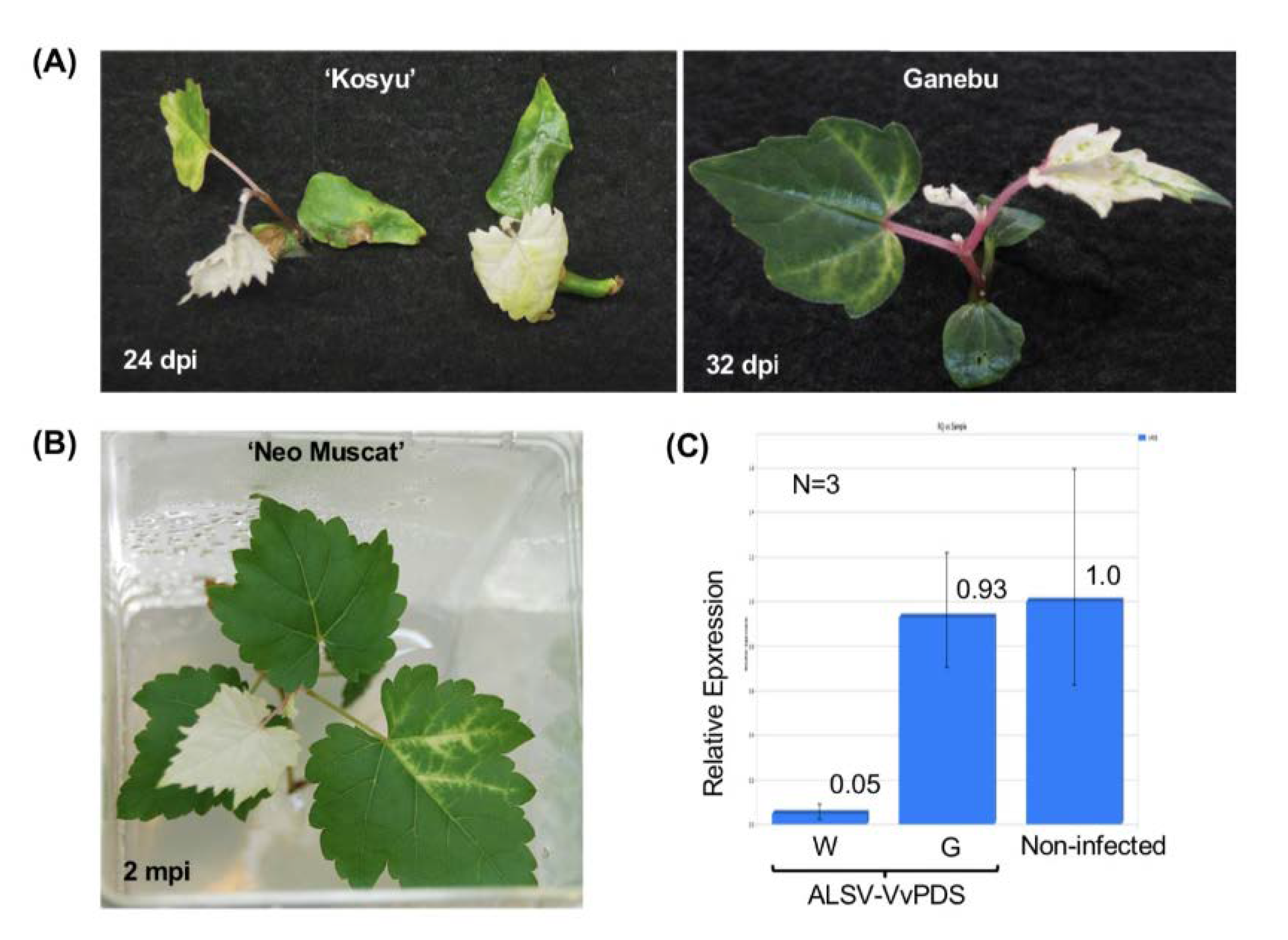
3.3. VIGS in Grapevine Using the ALSV Vector
3.4. VIF in Grapevine Using the ALSV Vector
3.5. Use of VIF for Evaluation of F1 Hybrids from Crossing between the Two Vitis Species
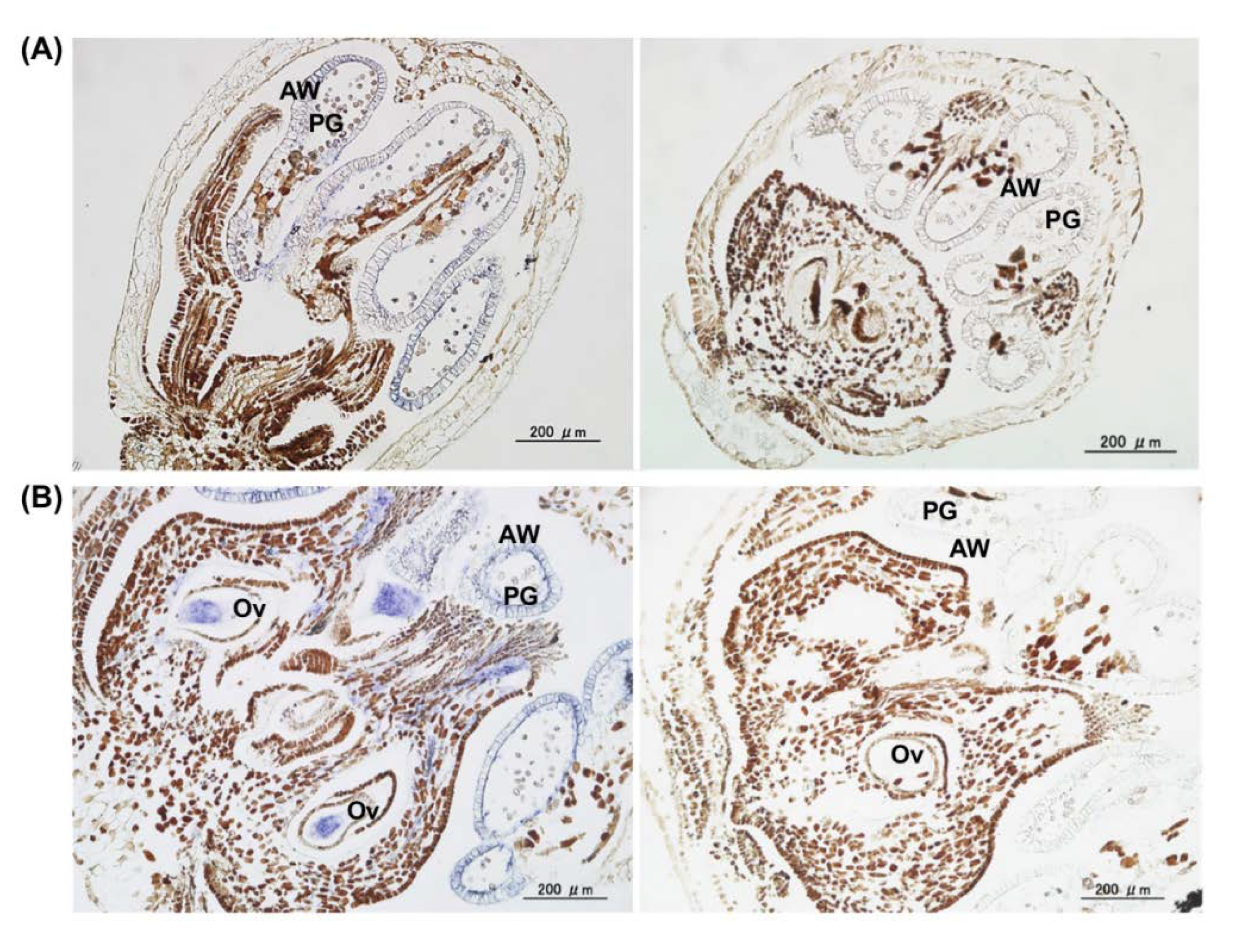
3.6. No Seed Transmission to Progeny Seedlings and Virus Elimination from Infected Grapevine Plants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carrington, J.C.; Kasschau, K.D.; Mahajan, S.K.; Schaad, M.C. Cell-to-cell and long-distance transport of viruses in plants. Plant Cell 1996, 8, 1669–1681. [Google Scholar] [CrossRef]
- Scholthof, H.B.; Scholthof, K.B.G.; Jackson, A.O. Plant virus gene vectors for transient expression of foreign proteins in plants. Annu. Rev. Phytopathol. 1996, 34, 299–323. [Google Scholar] [CrossRef] [PubMed]
- Burch-Smith, T.M.; Andersen, J.C.; Martin, G.B.; Dinesh-Kumar, S.P. Applications and advantages of virus-induced gene silencing for gene function studies in plants. Plant J. 2004, 39, 734–746. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, M.H.; Donson, J.; Della-Cioppa, G.; Harvey, D.; Hanley, K.; Grill, L.K. Cytoplasmic inhibition of carotenoid biosynthesis with virus-derived RNA. Proc. Natl. Acad. Sci. USA 1995, 92, 1679–1683. [Google Scholar] [CrossRef] [PubMed]
- Robertson, D. VIGS vectors for gene silencing: Many targets, many tools. Annu. Rev. Plant Biol. 2004, 55, 495–519. [Google Scholar] [CrossRef]
- Igarashi, A.; Yamagata, K.; Sugai, T.; Takahashi, Y.; Sugawara, E.; Tamura, A.; Yaegashi, H.; Yamagishi, N.; Takahashi, T.; Isogai, M.; et al. Apple latent spherical virus vectors for reliable and effective virus-induced gene silencing among a broad range of plants including tobacco, tomato, Arabidopsis thaliana, cucurbits, and legumes. Virology 2009, 386, 407–416. [Google Scholar] [CrossRef]
- Constantin, G.D.; Krath, B.N.; MacFarlane, S.A.; Nicolaisen, M.; Johansen, I.E.; Lund, O.S. Virus-induced gene silencing as a tool for functional genomics in a legume species. Plant J. 2004, 40, 622–631. [Google Scholar] [CrossRef]
- Dommes, A.B.; Gross, T.; Herbert, D.B.; Kivivirta, K.I.; Becker, A. Virus-induced gene silencing: Empowering genetics in non-model organism. J. Exp. Bot. 2019, 70, 757–779. [Google Scholar] [CrossRef]
- Grønlund, M.; Constantin, G.; Piendnoir, E.; Kovacev, J.; Johansen, I.E.; Lund, O.S. Virus-induced gene silencing in Medicago truncatula and Lathyrus odorata. Virus Res. 2008, 135, 345–349. [Google Scholar] [CrossRef]
- Holzberg, S.; Brosio, P.; Gross, C.; Pogue, G.P. Barley stripe mosaic virus-induced gene silencing in a monocot plant. Plant J. 2002, 30, 315–327. [Google Scholar] [CrossRef]
- Liu, Y.; Schiff, M.; Dinesh-Kumar, S.P. Virus-induced gene silencing in tomato. Plant J. 2002, 31, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Purkayastha, A.; Dasgupta, I. Virus-induced gene silencing: A versatile tool for discovery of gene functions in plants. Plant Physiol. Biochem. 2009, 47, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Scofield, S.R.; Nelson, R.S. Resources for virus-induced gene silencing in the grasses. Plant Physiol. 2009, 149, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Valentine, T.; Shaw, J.; Blok, V.C.; Phillips, M.S.; Oparka, K.J.; Lacomme, C. Efficient virus-induced gene silencing in roots using a modified Tobacco rattle virus vector. Plant Physiol. 2004, 136, 3999–4009. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Bradshaw, J.D.; Whitham, S.A.; Hill, J.H. The development of efficient multipurpose Bean pod mottle virus viral vector set for foreign gene expression and RNA silencing. Plant Physiol. 2010, 153, 52–65. [Google Scholar] [CrossRef]
- Zhao, F.; Lim, S.; Igori, D.; Too, R.H.; Kwon, S.-Y.; Kwon, S.-Y. Development of tobacco ringspot virus-based vectors for foreign gene expression and virus-induced gene silencing in a variety of plants. Virology 2016, 492, 166–178. [Google Scholar] [CrossRef]
- Folimonov, A.S.; Folimonova, S.Y.; Bar-Joseph, M.; Dawson, W.O. A stable RNA virus-based vector for citrus trees. Virology 2007, 368, 205–216. [Google Scholar] [CrossRef]
- Agüero, J.; Ruiz-Ruiz, S.; Vives, M.D.C.; Velázquez, K.; Navarro, L.; Peña, L.; Moreno, P.; Guerri, J. Development of viral vectors based on Citrus leaf blotch virus to express foreign proteins or analyze gene function in citrus plants. Mol. Plant Microbe Interact. 2012, 25, 1326–1337. [Google Scholar] [CrossRef]
- Agüero, J.; Vives, M.D.C.; Velázquez, K.; Pina, J.A.; Navarro, L.; Moreno, P.; Guerri, J. Effectiveness of gene silencing induced by viral vectors based on Citrus leaf blotch virus is different in Nicotiana benthamiana and citrus plants. Virology 2014, 460–461, 154–164. [Google Scholar] [CrossRef]
- Muruganantham, M.; Moskovitz, Y.; Haviv, S.; Horesh, T.; Fenigstein, A.; du Preez, J.; Stephan, D.; Burger, J.T.; Mawassi, M. Grapevine virus A-mediated gene silencing in Nicotiana benthamiana and Vitis vinifera. J. Virol. Methods 2009, 155, 167–174. [Google Scholar] [CrossRef]
- Kurth, E.G.; Peremyslov, V.V.; Prokhnevsky, A.I.; Kasschau, K.D.; Miller, M.; Carrington, J.C.; Dolja, V.V. Virus-derived gene expression and RNA interference vector for grapevine. J. Virol. 2012, 86, 6002–6009. [Google Scholar] [PubMed]
- Dawson, W.O.; Folimonova, S.Y. Virus-based transient expression vectors for woody crops: A new frontier for vector design and use. Annu. Rev. Phytopathol. 2013, 51, 321–337. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yamagishi, N.; Kasajima, I.; Yoshikawa, N. Virus-induced gene silencing and virus induced flowering in strawberry (Fragaria × ananassa) using apple latent spherical virus vector. Hortic. Res. 2019, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, S.; Yamagishi, N.; Yoshikawa, N. Efficient virus-induced gene silencing in apple, pear and Japanese pear using Apple latent spherical virus vectors. Plant Methods 2011, 7, 15. [Google Scholar] [CrossRef]
- Yamagishi, N.; Sasaki, S.; Yamagata, K.; Komori, S.; Nagase, M.; Wada, M.; Yamamoto, T.; Yoshikawa, N. Promotion of flowering and reduction of a generation time in apple seedlings by ectopical expression of the Arabidopsis thaliana FT gene using the Apple latent spherical virus vector. Plant Mol. Biol. 2011, 75, 193–204. [Google Scholar] [CrossRef]
- Yamagishi, N.; Kishigami, R.; Yoshikawa, N. Reduced generation time of apple seedlings to within a year by means of a plant virus vector: A new plant-breeding technique with no transmission of genetic modification to the next generation. Plant Biotechnol. J. 2014, 12, 60–68. [Google Scholar] [CrossRef]
- Thompson, J.R.; Dasgupta, I.; Fuchs, M.; Iwanami, T.; Karasev, A.V.; Petrzik, K.; Sanfacon, H.; Tzanetakis, I.; van der Vlugt, R.; Wetzel, T.; et al. ICTV virus taxonomy profile: Secoviridae. J. Gen. Virol. 2017, 98, 529–531. [Google Scholar] [CrossRef]
- Li, C.; Sasaki, N.; Isogai, M.; Yoshikawa, N. Stable expression of foreign proteins in herbaceous and apple plants using Apple latent spherical virus RNA2 vectors. Arch. Virol. 2004, 149, 1541–1558. [Google Scholar] [CrossRef]
- Li, C.; Yoshikawa, N.; Takahashi, T.; Ito, T.; Yoshida, K.; Koganezawa, H. Nucleotide sequence and genome organization of Apple latent spherical virus: A new virus classified into the family Comoviridae. J. Gen. Virol. 2000, 81, 541–547. [Google Scholar] [CrossRef]
- Fekih, R.; Yamagishi, N.; Yoshikawa, N. Apple latent spherical virus vector-induced flowering for shortening the juvenile phase in Japanese gentian and lisianthus plants. Planta 2016, 244, 203–214. [Google Scholar] [CrossRef]
- Fujita, N.; Kazama, Y.; Yamagishi, N.; Watanabe, K.; Ando, S.; Tsuji, H.; Kawano, S.; Yoshikawa, N.; Komatsu, K. Development of the VIGS system in the dioecious plant Silene latifolia. Int. J. Mol. Sci. 2019, 20, 1031. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, R.; Yamagishi, N.; Yoshikawa, N. A MYB transcription factor controls flower color in soybean. J. Hered. 2013, 104, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, N.; Yoshikawa, N. Virus-induced gene silencing in soybean seeds and emergence stage of soybean plants with Apple latent spherical virus vectors. Plant Mol. Biol. 2009, 71, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Tamura, A.; Kato, T.; Taki, A.; Sone, A.; Satoh, N.; Yamagishi, N.; Takahashi, T.; Ryo, B.-S.; Natsuaki, T.; Yoshikawa, N. Preventive and curative effects of Apple latent spherical virus vectors harboring part of the target virus genome against potyvirus and cucumovirus infections. Virology 2013, 446, 314–324. [Google Scholar] [CrossRef]
- Yamagishi, N.; Yoshikawa, N. Expression of FLOWERING LOCUS T from Arabidopsis thaliana induces precocious flowering in soybean irrespective of maturity group and stem growth habit. Planta 2011, 233, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, N.; Li, C.; Yoshikawa, N. Promotion of flowering by Apple latent spherical virus vector and virus elimination at high temperature allow accelerated breeding of apple and pear. Front. Plant Sci. 2016, 7, 171. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Yamagishi, N.; Isogai, M.; Komori, S.; Ito, T.; Yoshikawa, N. Seed and pollen transmission of Apple latent spherical virus in apple. J. Gen. Plant Pathol. 2011, 77, 48–53. [Google Scholar] [CrossRef]
- Where Are Grapes Grown? Worldatlas. Available online: https://www.worldatlas.com/articles/top-grape-growing-countries.html (accessed on 6 January 2020).
- Matsui, H. Physiological and Ecological Characteristics of Wild Grapes Originating from Japan. J. Brew. Soc. Jpn. 1989, 84, 687–693. (In Japanese) [Google Scholar] [CrossRef]
- Shiozaki, S.; Murakami, K. Flavonoid profiles of wild grapes native to Japan: Vitis coignetiae Pulliat and Vitis ficifolia Bunge var. ganebu Hatusima. Agric. Sci. 2017, 8, 239–252. [Google Scholar]
- Yamashita, H.; Mochioka, R. Wild grape germplasms in Japan. Adv. Hortic. Sci. 2014, 28, 214–224. [Google Scholar]
- Kon, T.; Yoshikawa, N. Induction and maintenance of DNA methylation in plant promoter sequences by apple latent spherical virus-induced transcriptional gene silencing. Front. Microbiol. 2014, 5, 595. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, N.; Sasaki, S.; Yoshikawa, N. Highly efficient inoculation method of apple viruses to apple seedlings. Julius-Kühn-Archiv 2010, 427, 226–229. [Google Scholar]
- Kamada, K.; Omata, S.; Yamagishi, N.; Kasajima, I.; Yoshikawa, N. Gentian (Gentiana triflora) prevents transmission of apple latent spherical virus (ALSV) vector to progeny seeds. Planta 2018, 248, 1431–1441. [Google Scholar] [CrossRef]
- Kasajima, I.; Ito, M.; Yamagishi, N.; Yoshikawa, N. Apple latent spherical virus (ALSV) vector as a tool for reverse genetic studies and non-transgenic breeding of a variety of crops. In Plant Epigenetics; Rajewsky, N., Jurga, S., Barciszewski, J., Eds.; RNA Technologies’ Series; Springer International Publishing AG: Cham, Switzerland, 2017; pp. 513–536. [Google Scholar]
- Namai, K.; Matsushima, Y.; Morishima, M.; Amagai, M.; Natusaki, T. Resistance to anthracnose is decreased by tissue culture but increased with longer acclimation in the resistant strawberry cultivar. J. Gen. Plant Pathol. 2013, 79, 402–411. [Google Scholar] [CrossRef]
- Carmona, M.J.; Calonje, M.; Martinez-Zapater, J.M. The FT/TFL1 gene family in grapevine. Plant Mol. Biol. 2007, 63, 637–650. [Google Scholar] [CrossRef]
- Yang, S.; Fresnedo-Ramirez, J.; Wang, M.; Cote, L.; Schweitzer, P.; Barba, P.; Takacs, E.M.; Clark, M.; Luby, J.; Manns, D.C.; et al. A next-generation marker genotyping platform (AmpSeq) in heterozygous crops: A case study for marker-assisted selection in grapevine. Hortic. Res. 2016, 3, 16002. [Google Scholar]
- Costa, L.D.; Malony, M.; Gribaudo, I. Breeding next generation tree fruit: Technical and legal challenges. Hortic. Res. 2017, 4, 17067. [Google Scholar]
- Malnoy, M.; Viola, R.; Jung, M.-H.; Koo, O.-J.; Kim, S.; Kim, J.-S.; Velasco, R.; Kanchiswarmy, C.N. DNA-free genetically edited grapevine and apple protoplast using CRISPER/Cas9 ribonucleoproteins. Front. Plant Sci. 2016, 7, 1904. [Google Scholar]
- Nakajima, I.; Ban, Y.; Azuma, A.; Onoue, N.; Moriguchi, T.; Yamamoto, T.; Toki, S.; Endo, M. CRISPER/Cas9-mediated targeted mutagenesis in grape. PLoS ONE 2017, 12, e0177966. [Google Scholar]
- Ren, C.; Liu, X.; Zhang, Z.; Wang, Y.; Duan, W.; Li, S.; Liang, Z. CRISPER/Cas9-mediated efficient targeted mutagenesis in Chardonnay (Vitis vinifera L.). Sci. Rep. 2016, 6, 32289. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material (Cultivar) | Growth Stage | System for Particle Bombardment * | ALSV Vector | No. of Infected/Inoculated Plants (%) | |
|---|---|---|---|---|---|
| Local ** | Systemic *** | ||||
| Seedlings (‘Koshu’) | Folded cotyledons | Helios Gene Gun | wtALSV | nt | 11/12 (92) |
| Expanded cotyledons | Helios Gene Gun | wtALSV | 10/10 (100) | 1/10 (10) | |
| Three true leaves | Helios Gene Gun | wtALSV | 10/10 (100) | 0/15 (0) | |
| Folded cotyledons | Helios Gene Gun | ALSV-VvPDS | nt | 10/11 (91) | |
| Expanded cotyledons | Helios Gene Gun | ALSV-VvPDS | 10/10 (100) | 1/12 (8) | |
| Three true leaves | Helios Gene Gun | ALSV-VvPDS | 15/15 (100) | 0/15 (0) | |
| Plants cultured in vitro (‘Neo Muscat’) | True leaf, non-rooted | PDS-1000/He™ | wtALSV | nt | 17/58 (29) |
| True leaf, rooted | GDS-80 | wtALSV | nt | 7/10 (70) | |
| True leaf, non-rooted | PDS-1000/He™ | ALSV-VvPDS | nt | 17/98 (17) | |
| Materials (Cultivar) | ALSV Vectors | No. of Flowered/Infected Plants (%) |
|---|---|---|
| Seedlings (‘Koshu’) | wtALSV | 0/10 (0) |
| ALSV-AtFT | 3/8 (38) | |
| ALSV-VvFT | 0/6 (0) | |
| ALSV-VvTFL1A | 0/9 (0) | |
| ALSV-VvTFL1B | 0/6 (0) | |
| ALSV-VvTFL1C | 0/5 (0) | |
| ALSV-AtFT/VvTFL1A | 8/9 (89) | |
| ALSV-AtFT/VvTFL1B | 5/9 (56) | |
| ALSV-VvFT/VvTFL1A | 0/6 (0) | |
| ALSV-VvFT/VvTFL1B | 0/3 (0) | |
| Seedlings (Ganebu) | wtALSV | 0/10 (0) |
| ALSV-AtFT | 6/6 (100) | |
| ALSV-VvFT | 0/6 (0) | |
| ALSV-VvTFL1A | 0/10 (0) | |
| ALSV-VvTFL1B | 0/10 (0) | |
| ALSV-VvTFL1C | 0/5 (0) | |
| ALSV-AtFT/VvTFL1A | 8/12 (67) | |
| ALSV-AtFT/VvTFL1B | 17/21 (81) | |
| ALSV-VvFT/VvTFL1A | 0/5 (0) | |
| ALSV-VvFT/VvTFL1B | 0/5 (0) | |
| In vitro cultures (‘Neo Muscut’) | wtALSV | 0/20 (0) |
| ALSV-AtFT/VvTFL1A | 11/16 (69) | |
| ALSV-AtFT/VvTFL1B | 5/7 (71) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maeda, K.; Kikuchi, T.; Kasajima, I.; Li, C.; Yamagishi, N.; Yamashita, H.; Yoshikawa, N. Virus-Induced Flowering by Apple Latent Spherical Virus Vector: Effective Use to Accelerate Breeding of Grapevine. Viruses 2020, 12, 70. https://doi.org/10.3390/v12010070
Maeda K, Kikuchi T, Kasajima I, Li C, Yamagishi N, Yamashita H, Yoshikawa N. Virus-Induced Flowering by Apple Latent Spherical Virus Vector: Effective Use to Accelerate Breeding of Grapevine. Viruses. 2020; 12(1):70. https://doi.org/10.3390/v12010070
Chicago/Turabian StyleMaeda, Kiyoaki, Teppei Kikuchi, Ichiro Kasajima, Chungjiang Li, Noriko Yamagishi, Hiroyuki Yamashita, and Nobuyuki Yoshikawa. 2020. "Virus-Induced Flowering by Apple Latent Spherical Virus Vector: Effective Use to Accelerate Breeding of Grapevine" Viruses 12, no. 1: 70. https://doi.org/10.3390/v12010070
APA StyleMaeda, K., Kikuchi, T., Kasajima, I., Li, C., Yamagishi, N., Yamashita, H., & Yoshikawa, N. (2020). Virus-Induced Flowering by Apple Latent Spherical Virus Vector: Effective Use to Accelerate Breeding of Grapevine. Viruses, 12(1), 70. https://doi.org/10.3390/v12010070