Influenza D Virus: Serological Evidence in the Italian Population from 2005 to 2017

 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Influenza Viruses
2.2. Serum Samples
2.3. Haemagglutination Inhibition Assay
2.4. Virus Neutralization Assay
2.5. Statistical Analysis
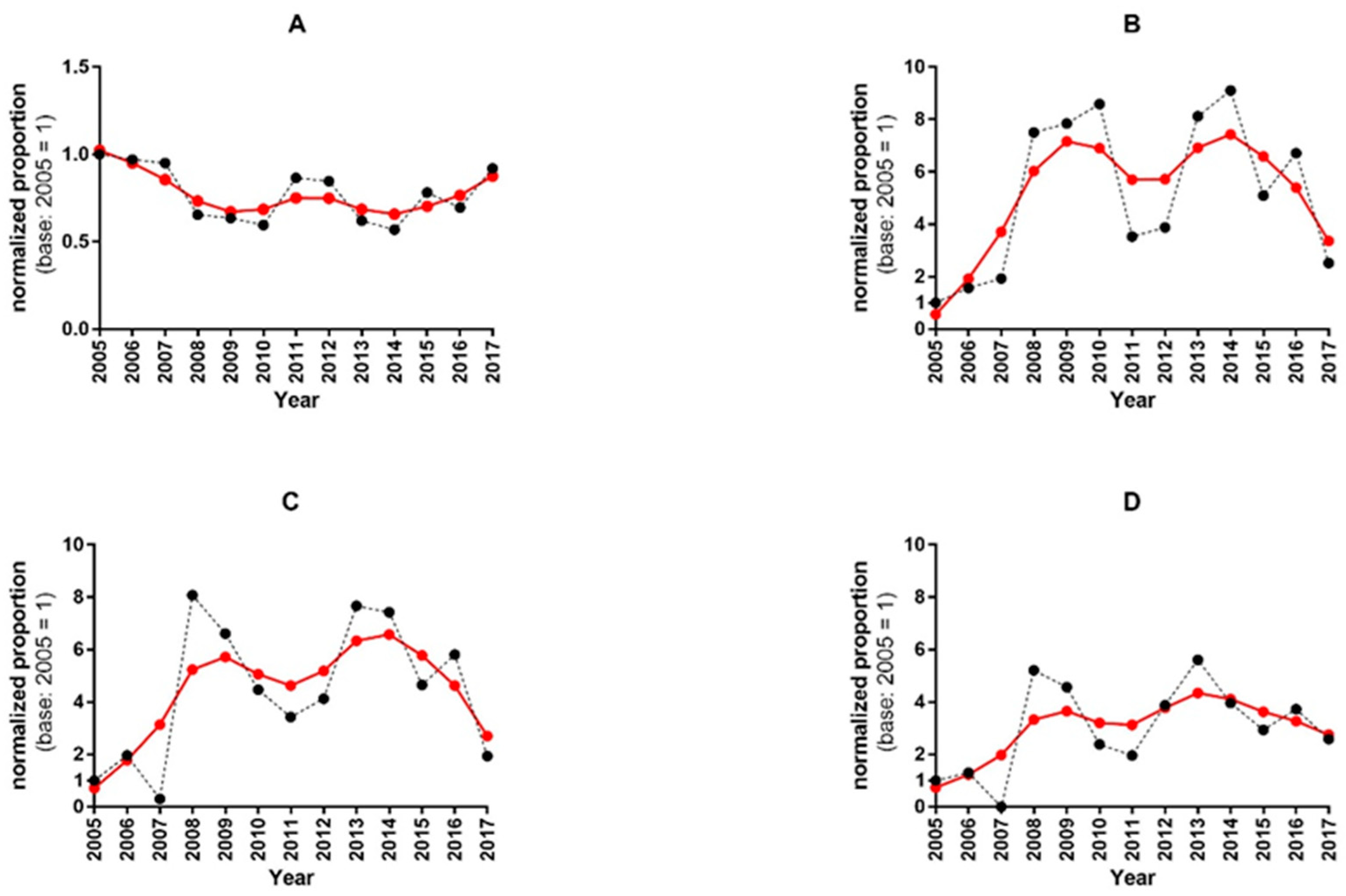
3. Results
3.1. Investigations on Potential Cross-Reactivity between IDV- and ICV-Positive Serum Samples
3.2. Analysis of Human Serum Samples for the Presence of IDV-Specific Antibodies
3.3. Pearson’s Chi-Squared Test for Multi-Proportions
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hause, B.M.; Ducatez, M.; Collin, E.A.; Ran, Z.; Liu, R.; Sheng, Z.; Armien, A.; Kaplan, B.; Chakravarty, S.; Hoppe, A.D.; et al. Isolation of a novel swine influenza virus from oklahoma in 2011 which is distantly related to human influenza c viruses. PLoS Pathog. 2013, 9, e1003176. [Google Scholar] [CrossRef] [PubMed]
- Centers for Excellence for Influenza Research and Surveillance. Introducing Influenza D: International Committee on Taxonomy of Viruses Names New Virus. Available online: http://www.niaidceirs.org/news/2017/02/introducing-influenza-d-international-committee-on-taxonomy-of-viruses-names-new-virus/ (accessed on 27 November 2019).
- Hause, B.M.; Collin, E.A.; Liu, R.; Huang, B.; Sheng, Z.; Lu, W.; Wang, D.; Nelson, E.A.; Li, F. Characterization of a novel influenza virus in cattle and swine: Proposal for a new genus in the orthomyxoviridae family. MBio 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Z.; Ran, Z.; Wang, D.; Hoppe, A.D.; Simonson, R.; Chakravarty, S.; Hause, B.M.; Li, F. Genomic and evolutionary characterization of a novel influenza-c-like virus from swine. Arch. Virol. 2014, 159, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Asha, K.; Kumar, B. Emerging influenza d virus threat: What we know so far! J. Clin. Med. 2019, 8, 192. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.M.; Wang, S.C.; Peng, C.; Yu, J.M.; Zhuang, Q.Y.; Hou, G.Y.; Liu, S.; Li, J.P.; Chen, J.M. Identification of a potential novel type of influenza virus in bovine in china. Virus Genes 2014, 49, 493–496. [Google Scholar] [CrossRef]
- Ducatez, M.F.; Pelletier, C.; Meyer, G. Influenza D virus in cattle, France, 2011–2014. Emerg. Infect. Dis. 2015, 21, 368–371. [Google Scholar] [CrossRef]
- Collin, E.A.; Sheng, Z.; Lang, Y.; Ma, W.; Hause, B.M.; Li, F. Cocirculation of two distinct genetic and antigenic lineages of proposed influenza d virus in cattle. J. Virol. 2015, 89, 1036–1042. [Google Scholar] [CrossRef]
- Ferguson, L.; Eckard, L.; Epperson, W.B.; Long, L.P.; Smith, D.; Huston, C.; Genova, S.; Webby, R.; Wan, X.F. Influenza d virus infection in mississippi beef cattle. Virology 2015, 486, 28–34. [Google Scholar] [CrossRef]
- Ferguson, L.; Olivier, A.K.; Genova, S.; Epperson, W.B.; Smith, D.R.; Schneider, L.; Barton, K.; McCuan, K.; Webby, R.J.; Wan, X.F. Pathogenesis of influenza d virus in cattle. J. Virol. 2016, 90, 5636–5642. [Google Scholar] [CrossRef]
- Murakami, S.; Endoh, M.; Kobayashi, T.; Takenaka-Uema, A.; Chambers, J.K.; Uchida, K.; Nishihara, M.; Hause, B.; Horimoto, T. Influenza d virus infection in herd of cattle, Japan. Emerg. Infect. Dis. 2016, 22, 1517–1519. [Google Scholar] [CrossRef]
- Horimoto, T.; Hiono, T.; Mekata, H.; Odagiri, T.; Lei, Z.; Kobayashi, T.; Norimine, J.; Inoshima, Y.; Hikono, H.; Murakami, K.; et al. Nationwide distribution of bovine influenza d virus infection in japan. PLoS ONE 2016, 11, e0163828. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Ferguson, L.; Smith, D.R.; Woolums, A.R.; Epperson, W.B.; Wan, X.F. Serological evidence for high prevalence of influenza d viruses in cattle, nebraska, united states, 2003–2004. Virology 2017, 501, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Hill, J.E.; Godson, D.L.; Ngeleka, M.; Fernando, C.; Huang, Y. The pulmonary virome, bacteriological and histopathological findings in bovine respiratory disease from western Canada. Transbound. Emerg. Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Dane, H.; Duffy, C.; Guelbenzu, M.; Hause, B.; Fee, S.; Forster, F.; McMenamy, M.J.; Lemon, K. Detection of influenza d virus in bovine respiratory disease samples, UK. Transbound. Emerg. Dis. 2019, 66, 2184–2187. [Google Scholar] [CrossRef]
- Flynn, O.; Gallagher, C.; Mooney, J.; Irvine, C.; Ducatez, M.; Hause, B.; McGrath, G.; Ryan, E. Influenza d virus in cattle, Ireland. Emerg. Infect. Dis. 2018, 24, 389–391. [Google Scholar] [CrossRef]
- Mitra, N.; Cernicchiaro, N.; Torres, S.; Li, F.; Hause, B.M. Metagenomic characterization of the virome associated with bovine respiratory disease in feedlot cattle identified novel viruses and suggests an etiologic role for influenza d virus. J. Gen. Virol. 2016, 97, 1771–1784. [Google Scholar] [CrossRef]
- Snoeck, C.J.; Oliva, J.; Pauly, M.; Losch, S.; Wildschutz, F.; Muller, C.P.; Hubschen, J.M.; Ducatez, M.F. Influenza d virus circulation in cattle and swine, Luxembourg, 2012–2016. Emerg. Infect. Dis. 2018, 24, 1388–1389. [Google Scholar] [CrossRef]
- Quast, M.; Sreenivasan, C.; Sexton, G.; Nedland, H.; Singrey, A.; Fawcett, L.; Miller, G.; Lauer, D.; Voss, S.; Pollock, S.; et al. Serological evidence for the presence of influenza d virus in small ruminants. Vet. Microbiol. 2015, 180, 281–285. [Google Scholar] [CrossRef]
- Salem, E.; Cook, E.A.J.; Lbacha, H.A.; Oliva, J.; Awoume, F.; Aplogan, G.L.; Hymann, E.C.; Muloi, D.; Deem, S.L.; Alali, S.; et al. Serologic evidence for influenza c and d virus among ruminants and camelids, Africa, 1991–2015. Emerg. Infect. Dis. 2017, 23, 1556–1559. [Google Scholar] [CrossRef]
- Zhai, S.L.; Zhang, H.; Chen, S.N.; Zhou, X.; Lin, T.; Liu, R.; Lv, D.H.; Wen, X.H.; Wei, W.K.; Wang, D.; et al. Influenza d virus in animal species in Guangdong province, Southern China. Emerg. Infect. Dis. 2017, 23, 1392–1396. [Google Scholar] [CrossRef]
- Nedland, H.; Wollman, J.; Sreenivasan, C.; Quast, M.; Singrey, A.; Fawcett, L.; Christopher-Hennings, J.; Nelson, E.; Kaushik, R.S.; Wang, D.; et al. Serological evidence for the co-circulation of two lineages of influenza d viruses in equine populations of the Midwest United States. Zoonoses Public Health 2018, 65, e148–e154. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Odagiri, T.; Melaku, S.K.; Bazartseren, B.; Ishida, H.; Takenaka-Uema, A.; Muraki, Y.; Sentsui, H.; Horimoto, T. Influenza d virus infection in dromedary camels, Ethiopia. Emerg. Infect. Dis. 2019, 25, 1224–1226. [Google Scholar] [CrossRef] [PubMed]
- O’Donovan, T.; Donohoe, L.; Ducatez, M.F.; Meyer, G.; Ryan, E. Seroprevalence of influenza d virus in selected sample groups of irish cattle, sheep and pigs. Ir. Vet. J. 2019, 72, 11. [Google Scholar] [CrossRef] [PubMed]
- Foni, E.; Chiapponi, C.; Baioni, L.; Zanni, I.; Merenda, M.; Rosignoli, C.; Kyriakis, C.S.; Luini, M.V.; Mandola, M.L.; Bolzoni, L.; et al. Influenza d in Italy: Towards a better understanding of an emerging viral infection in swine. Sci. Rep. 2017, 7, 11660. [Google Scholar] [CrossRef]
- Su, S.; Fu, X.; Li, G.; Kerlin, F.; Veit, M. Novel influenza d virus: Epidemiology, pathology, evolution and biological characteristics. Virulence 2017, 8, 1580–1591. [Google Scholar] [CrossRef]
- Ferguson, L.; Luo, K.; Olivier, A.K.; Cunningham, F.L.; Blackmon, S.; Hanson-Dorr, K.; Sun, H.; Baroch, J.; Lutman, M.W.; Quade, B.; et al. Influenza d virus infection in feral swine populations, United States. Emerg. Infect. Dis. 2018, 24, 1020–1028. [Google Scholar] [CrossRef]
- Chiapponi, C.; Faccini, S.; De Mattia, A.; Baioni, L.; Barbieri, I.; Rosignoli, C.; Nigrelli, A.; Foni, E. Detection of influenza d virus among swine and cattle, Italy. Emerg. Infect. Dis. 2016, 22, 352–354. [Google Scholar] [CrossRef]
- Smith, D.B.; Gaunt, E.R.; Digard, P.; Templeton, K.; Simmonds, P. Detection of influenza c virus but not influenza d virus in Scottish respiratory samples. J. Clin. Virol. 2016, 74, 50–53. [Google Scholar] [CrossRef]
- White, S.K.; Ma, W.; McDaniel, C.J.; Gray, G.C.; Lednicky, J.A. Serologic evidence of exposure to influenza d virus among persons with occupational contact with cattle. J. Clin. Virol. 2016, 81, 31–33. [Google Scholar] [CrossRef]
- Trombetta, C.M.; Perini, D.; Mather, S.; Temperton, N.; Montomoli, E. Overview of serological techniques for influenza vaccine evaluation: Past, present and future. Vaccines 2014, 2, 707–734. [Google Scholar] [CrossRef]
- Clopper, C.J.; Pearson, E.S. The use of confidence or fiducial limits illustrated in the case of the binomial. Biometrika 1934, 26, 404–413. [Google Scholar] [CrossRef]
- Hodrick, R.J.; Prescott, E.C. Postwar U.S. Business cycles: An empirical investigation. J. Money Credit Bank. 1997, 29, 1–16. [Google Scholar] [CrossRef]
- Borkenhagen, L.K.; Mallinson, K.A.; Tsao, R.W.; Ha, S.J.; Lim, W.H.; Toh, T.H.; Anderson, B.D.; Fieldhouse, J.K.; Philo, S.E.; Chong, K.S.; et al. Surveillance for respiratory and diarrheal pathogens at the human-pig interface in Sarawak, Malaysia. PLoS ONE 2018, 13, e0201295. [Google Scholar] [CrossRef] [PubMed]
- Bailey, E.S.; Choi, J.Y.; Zemke, J.; Yondon, M.; Gray, G.C. Molecular surveillance of respiratory viruses with bioaerosol sampling in an airport. Trop. Dis. Travel Med. Vaccines 2018, 4, 11. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasan, C.; Thomas, M.; Sheng, Z.; Hause, B.M.; Collin, E.A.; Knudsen, D.E.; Pillatzki, A.; Nelson, E.; Wang, D.; Kaushik, R.S.; et al. Replication and transmission of the novel bovine influenza d virus in a guinea pig model. J. Virol. 2015, 89, 11990–12001. [Google Scholar] [CrossRef]
- Song, H.; Qi, J.; Khedri, Z.; Diaz, S.; Yu, H.; Chen, X.; Varki, A.; Shi, Y.; Gao, G.F. Correction: An open receptor-binding cavity of hemagglutinin-esterase-fusion glycoprotein from newly-identified influenza d virus: Basis for its broad cell tropism. PLoS Pathog. 2016, 12, e1005505. [Google Scholar] [CrossRef]
- Holwerda, M.; Kelly, J.; Laloli, L.; Sturmer, I.; Portmann, J.; Stalder, H.; Dijkman, R. Determining the replication kinetics and cellular tropism of influenza d virus on primary well-differentiated human airway epithelial cells. Viruses 2019, 11, 337. [Google Scholar] [CrossRef]
- Chiapponi, C.; Faccini, S.; Fusaro, A.; Moreno, A.; Prosperi, A.; Merenda, M.; Baioni, L.; Gabbi, V.; Rosignoli, C.; Alborali, G.L.; et al. Detection of a new genetic cluster of influenza d virus in Italian cattle. Viruses 2019, 11, 1110. [Google Scholar] [CrossRef]
- Odagiri, T.; Ishida, H.; Li, J.Y.; Endo, M.; Kobayashi, T.; Kamiki, H.; Matsugo, H.; Takenaka-Uema, A.; Murakami, S.; Horimoto, T. Antigenic heterogeneity among phylogenetic clusters of influenza d viruses. J. Vet. Med Sci. 2018, 80, 1241–1244. [Google Scholar] [CrossRef]


{kind=link}
| Influenza Viruses | IDV Antiserum HI Titre | ICV Antiserum HI Titre |
|---|---|---|
| D/bovine/Oklahoma/660/2013 | 1:10,240 | 5 |
| C/Victoria/2/2012 | 5 | 1:640 |
| HI Assay | |||||||||||||||
| Year | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | Total | |
| Samples | 99 | 101 | 82 | 95 | 101 | 83 | 101 | 102 | 100 | 100 | 101 | 115 | 101 | 1281 | |
| Titre | 5 | 94 | 93 | 74 | 59 | 61 | 47 | 83 | 82 | 59 | 54 | 75 | 76 | 89 | 946 |
| ≥1:10 | 5 | 8 | 8 | 36 | 40 | 36 | 18 | 20 | 41 | 46 | 26 | 39 | 12 | 335 | |
| ≥1:20 | 4 | 8 | 1 | 31 | 27 | 15 | 14 | 17 | 31 | 30 | 19 | 27 | 12 | 236 | |
| ≥1:40 | 3 | 4 | 0 | 15 | 14 | 6 | 6 | 12 | 17 | 12 | 9 | 13 | 8 | 119 | |
| ≥1:80 | 2 | 1 | 0 | 7 | 9 | 3 | 2 | 8 | 7 | 2 | 2 | 4 | 5 | 52 | |
| ≥1:160 | 1 | 0 | 0 | 5 | 2 | 0 | 2 | 0 | 2 | 0 | 0 | 3 | 3 | 18 | |
| N≥1:10 | 5 | 8 | 8 | 36 | 40 | 36 | 18 | 20 | 41 | 46 | 26 | 39 | 12 | 335 | |
| %≥1:10 | 5.1% | 7.9% | 9.8% | 37.9% | 39.6% | 43.4% | 17.8% | 19.6% | 41.0% | 46.0% | 25.7% | 33.9% | 11.9% | 26.2% | |
| CI 95% lower | 1.7% | 3.5% | 4.3% | 28.1% | 30.0% | 32.5% | 10.9% | 12.4% | 31.3% | 36.0% | 17.6% | 25.3% | 6.3% | 23.8% | |
| CI 95% upper | 11.4% | 15.0% | 18.3% | 48.4% | 49.8% | 54.7% | 26.7% | 28.6% | 51.3% | 56.3% | 35.4% | 43.3% | 19.8% | 28.6% | |
| (A) | |||||||||||||||
| VN Assay | |||||||||||||||
| Year | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | Total | |
| Samples | 5 | 8 | 8 | 36 | 40 | 36 | 18 | 20 | 41 | 46 | 26 | 39 | 12 | 335 | |
| Titre | 5 | 0 | 1 | 0 | 2 | 7 | 16 | 1 | 5 | 2 | 4 | 4 | 9 | 4 | 55 |
| ≥1:10 | 5 | 7 | 8 | 34 | 33 | 20 | 17 | 15 | 39 | 42 | 22 | 30 | 8 | 280 | |
| ≥1:20 | 5 | 5 | 8 | 33 | 18 | 17 | 15 | 13 | 29 | 35 | 35 | 20 | 7 | 240 | |
| ≥1:40 | 4 | 4 | 2 | 10 | 2 | 5 | 9 | 9 | 12 | 14 | 14 | 7 | 4 | 96 | |
| ≥1:80 | 1 | 1 | 0 | 1 | 0 | 1 | 2 | 0 | 4 | 0 | 0 | 0 | 1 | 11 | |
| ≥1:160 | 0 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 1 | 0 | 0 | 0 | 0 | 4 | |
| N≥1:10 | 5 | 7 | 8 | 34 | 33 | 20 | 17 | 15 | 39 | 42 | 22 | 30 | 8 | 280 | |
| %≥1:10 | 5.1% | 6.9% | 9.8% | 35.8% | 32.7% | 24.1% | 16.8% | 14.7% | 39.0% | 42.0% | 21.8% | 26.1% | 7.9% | 21.9% | |
| CI 95% lower | 1.7% | 2.8% | 4.3% | 26.2% | 23.7% | 15.4% | 10.1% | 8.5% | 29.4% | 32.2% | 14.2% | 18.3% | 3.5% | 19.6% | |
| CI 95% upper | 11.4% | 13.8% | 18.3% | 46.3% | 42.7% | 34.7% | 25.6% | 23.1% | 49.3% | 52.3% | 31.1% | 35.1% | 15.0% | 24.2% | |
| (B) | |||||||||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trombetta, C.M.; Marchi, S.; Manini, I.; Kistner, O.; Li, F.; Piu, P.; Manenti, A.; Biuso, F.; Sreenivasan, C.; Druce, J.; et al. Influenza D Virus: Serological Evidence in the Italian Population from 2005 to 2017. Viruses 2020, 12, 30. https://doi.org/10.3390/v12010030
Trombetta CM, Marchi S, Manini I, Kistner O, Li F, Piu P, Manenti A, Biuso F, Sreenivasan C, Druce J, et al. Influenza D Virus: Serological Evidence in the Italian Population from 2005 to 2017. Viruses. 2020; 12(1):30. https://doi.org/10.3390/v12010030
Chicago/Turabian StyleTrombetta, Claudia M., Serena Marchi, Ilaria Manini, Otfried Kistner, Feng Li, Pietro Piu, Alessandro Manenti, Fabrizio Biuso, Chithra Sreenivasan, Julian Druce, and et al. 2020. "Influenza D Virus: Serological Evidence in the Italian Population from 2005 to 2017" Viruses 12, no. 1: 30. https://doi.org/10.3390/v12010030
APA StyleTrombetta, C. M., Marchi, S., Manini, I., Kistner, O., Li, F., Piu, P., Manenti, A., Biuso, F., Sreenivasan, C., Druce, J., & Montomoli, E. (2020). Influenza D Virus: Serological Evidence in the Italian Population from 2005 to 2017. Viruses, 12(1), 30. https://doi.org/10.3390/v12010030