Pathogenic Characteristics of a Porcine Astrovirus Strain Isolated in China

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Viruses and Cells
2.2. Animals
2.3. Clinical and Necropsy Assessment
2.4. Histopathology and Morphometry
2.5. qRT-PCR
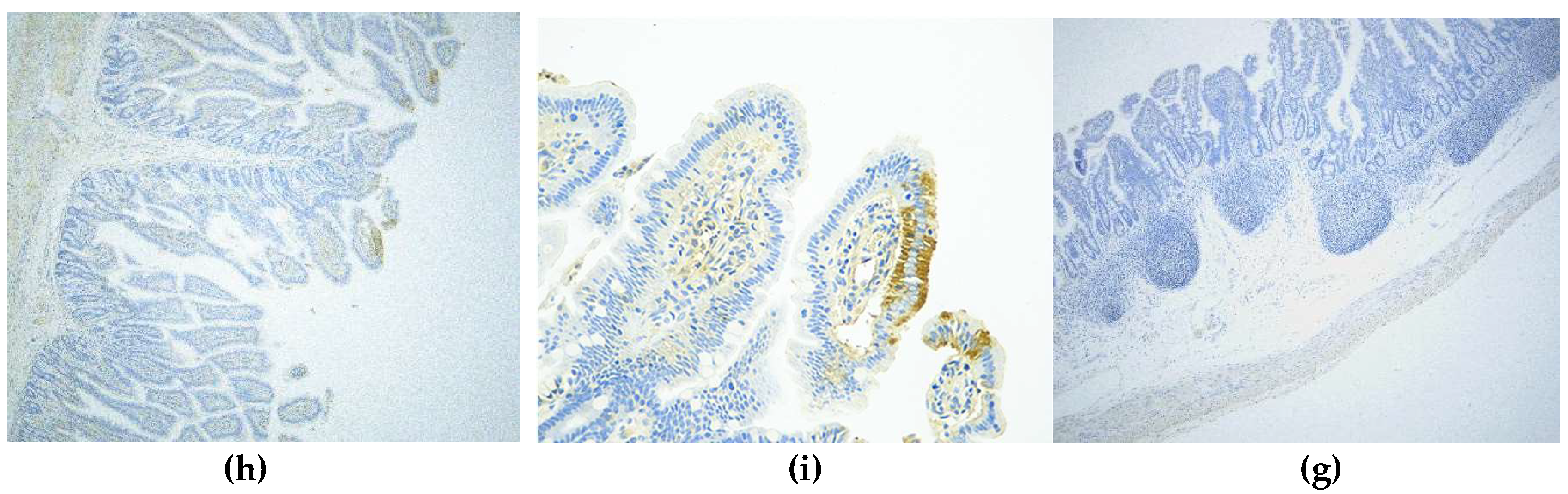
2.6. Immunohistochemistry (IHC)
2.7. Statistical Analysis
3. Results
3.1. Infection of Pigs with PAstV1-GX1 Results in Mild Diarrhea
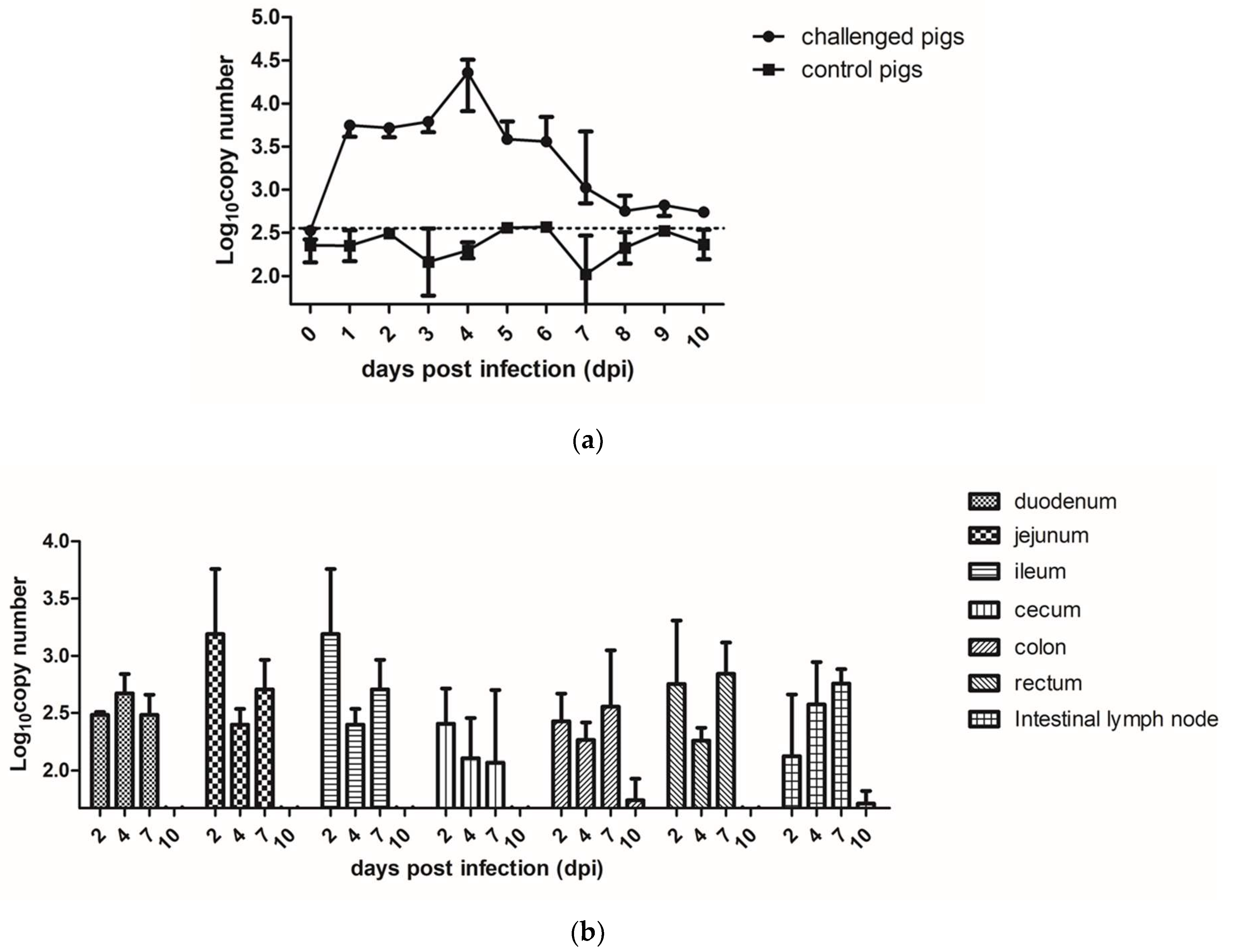
3.2. Fecal Shedding, Viral Load, and Tissue Distribution in PAstV-Inoculated Pigs
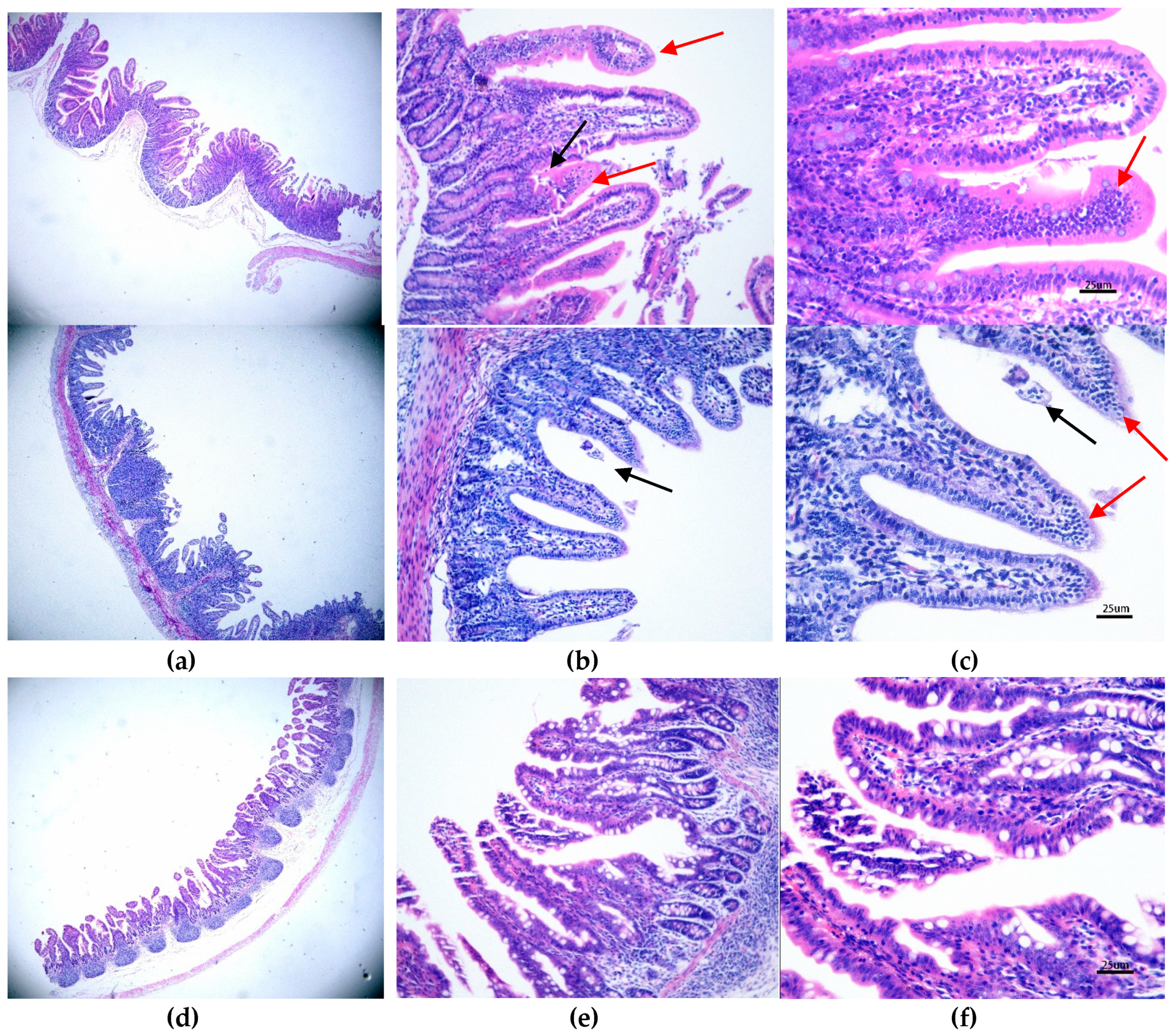
3.3. Histopathological and Morphometric Changes Induced by PAstV1-GX1 Infection
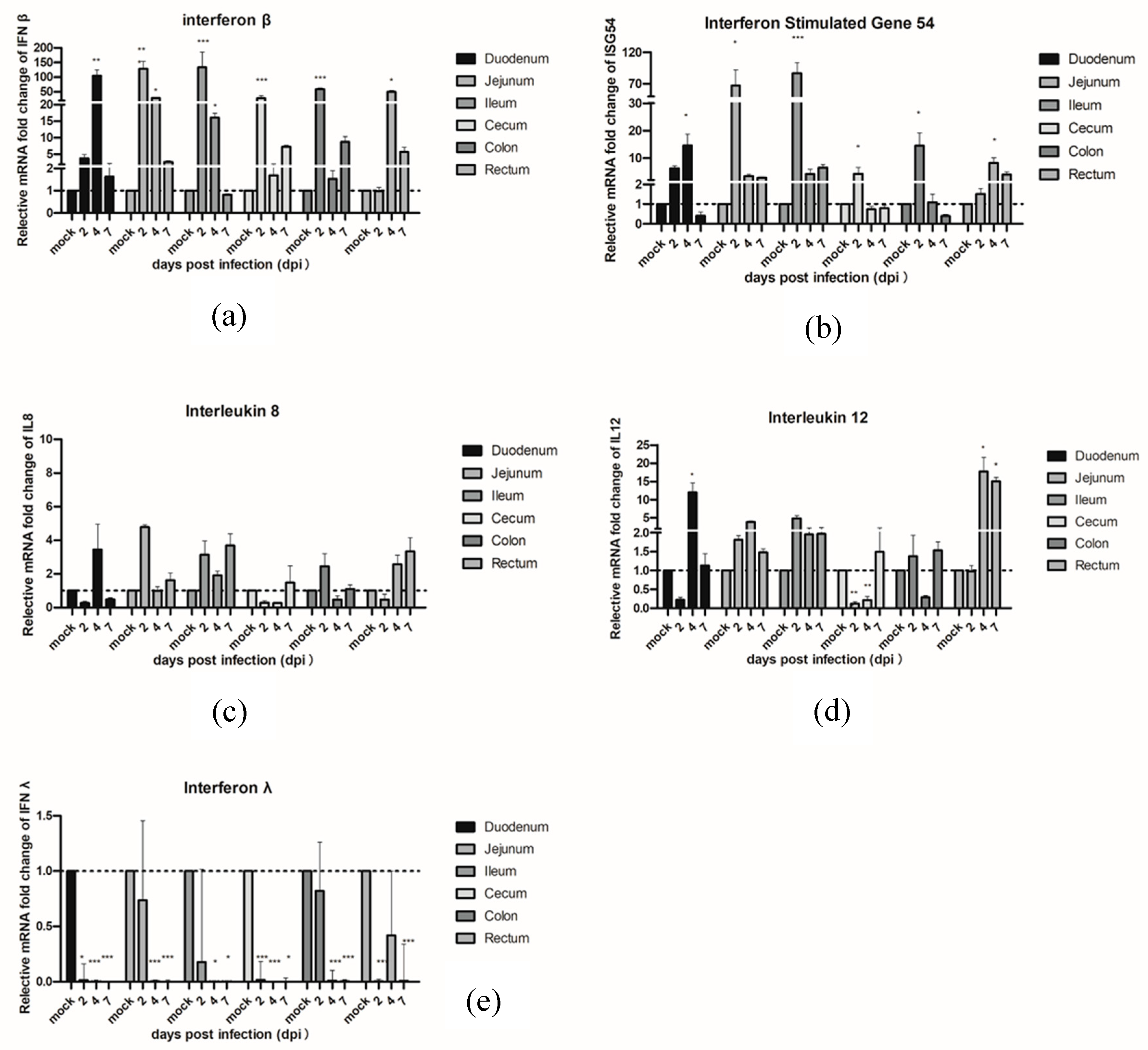
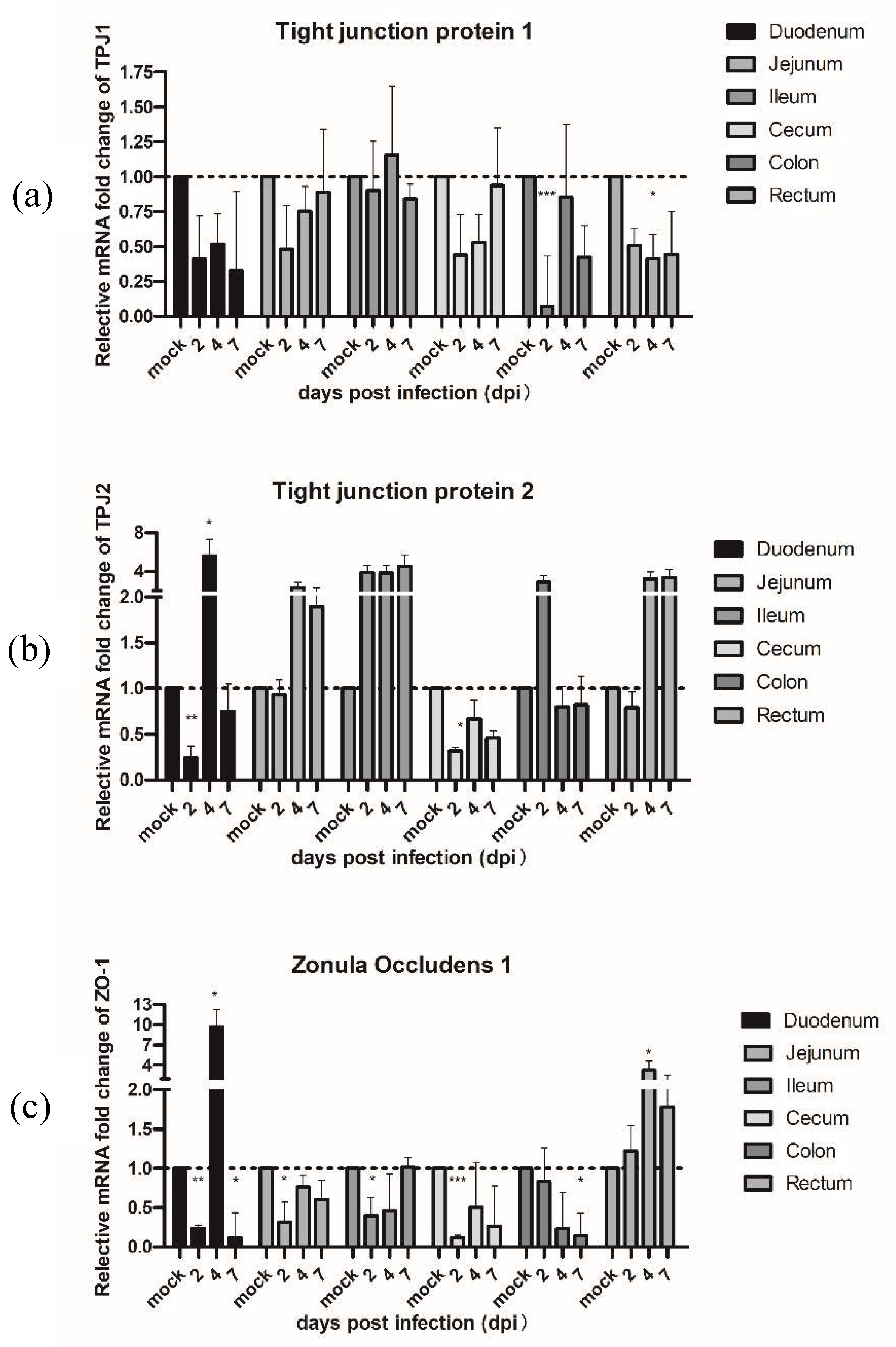
3.4. The Effects of PAstV on Transcriptional Activation of Inflammatory Cytokines and Permeable Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Benedictis, P.D.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals–Molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G.; Pankovics, P.; Delwart, E.; Boros, Á. Identification of a novel astrovirus in domestic sheep in Hungary. Arch. Virol. 2011, 156, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Bridger, J.C. Detection by electron microscopy of caliciviruses, astroviruses and rotavirus-like particles in the faeces of piglets with diarrhoea. Vet. Rec. 1980, 107, 532–533. [Google Scholar] [PubMed]
- Brnić, D.; Jemeršić, L.; Keros, T.; Prpić, J. High prevalence and genetic heterogeneity of porcine astroviruses in domestic pigs. Vet. J. 2014, 202, 390–392. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Yin, W.; Zhou, Y.; Li, B.; Ai, L.; Pan, M.; Guo, W. Molecular detection of Porcine astrovirus in Sichuan Province, China. Virol. J. 2016, 13, 6. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.T.; Giménez-Lirola, L.G.; Gerber, P.F.; Jiang, Y.H.; Halbur, P.G.; Opriessnig, T. Identification and characterization of novel porcine astroviruses (PAstVs) with high prevalence and frequent co-infection of individual pigs with multiple PAstV types. J. Gen. Virol. 2013, 94, 570. [Google Scholar] [CrossRef]
- Zhou, W.; Ullman, K.; Chowdry, V.; Reining, M.; Benyeda, Z.; Baule, C.; Juremalm, M.; Wallgren, P.; Schwarz, L.; Zhou, E. Molecular investigations on the prevalence and viral load of enteric viruses in pigs from five European countries. Vet. Microbiol. 2016, 182, 75–81. [Google Scholar] [CrossRef]
- Lan, D.; Ji, W.; Shan, T.; Cui, L.; Yang, Z.; Yuan, C.; Hua, X. Molecular characterization of a porcine astrovirus strain in China. Arch. Virol. 2011, 156, 1869–1875. [Google Scholar] [CrossRef]
- Ashley, C.R.; Caul, E.O.; Paver, W.K. Astrovirus-associated gastroenteritis in children. Lancet 1975, 306, 559. [Google Scholar] [CrossRef]
- Cortez, V.; Meliopoulos, V.A.; Karlsson, E.A.; Hargest, V.; Johnson, C.; Schultz-Cherry, S. Astrovirus biology and pathogenesis. Annu. Rev. Virol. 2017, 4, 327–348. [Google Scholar] [CrossRef]
- Bosch, A.; Pinto, R.M.; Guix, S. Human astroviruses. Clin. Microbiol. Rev. 2014, 27, 1048–1074. [Google Scholar] [CrossRef] [PubMed]
- Mendenhall, I.H.; Smith, G.J.; Vijaykrishna, D. Ecological drivers of virus evolution: Astrovirus as a case study. J. Virol. 2015, 89, 6978–6981. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, E.A.; Small, C.T.; Freiden, P.; Feeroz, M.M.; Matsen, T.F.; San, S.; Hasan, M.K.; Wang, D.; Jonesengel, L.; Schultzcherry, S. Non-human primates harbor diverse mammalian and avian astroviruses including those associated with human infections. PLoS Pathog. 2015, 11, e1005225. [Google Scholar] [CrossRef] [PubMed]
- Pankovics, P.; Boros, A.; Kiss, T.; Delwart, E.; Reuter, G. Detection of a mammalian-like astrovirus in bird, European roller (Coracias garrulus). Infect. Genet. Evol. 2015, 34, 114–121. [Google Scholar] [CrossRef]
- Finkbeiner, S.R.; Le, B.M.; Holtz, L.R.; Storch, G.A.; Wang, D. Detection of newly described astrovirus MLB1 in stool samples from children. Emerg. Infect. Dis. 2009, 15, 441–444. [Google Scholar] [CrossRef]
- Rivera, R.; Nollens, H.H.; Venn-Watson, S.; Gulland, F.M.; Wellehan, F.X., Jr. Characterization of phylogenetically diverse astroviruses of marine mammals. J. Gen. Virol. 2010, 91, 166. [Google Scholar] [CrossRef]
- Johnson, C.; Hargest, V.; Cortez, V.; Meliopoulos, V.A.; Stacey, S.-C. Astrovirus pathogenesis. Viruses 2017, 9, 22. [Google Scholar] [CrossRef]
- Indik, S.; Valíček, L.; Šmíd, B.; Dvořáková, H.; Rodák, L. Isolation and partial characterization of a novel porcine astrovirus. Vet. Microbiol. 2006, 117, 276–283. [Google Scholar] [CrossRef]
- Mor, S.K.; Chander, Y.; Marthaler, D.; Patnayak, D.P.; Goyal, S.M. Detection and molecular characterization of Porcine astrovirus strains associated with swine diarrhea. J. Vet. Diagn. Investig. 2012, 24, 1064–1067. [Google Scholar] [CrossRef]
- Shan, T.; Li, L.; Simmonds, P.; Wang, C.; Moeser, A.; Delwart, E. The fecal virome of pigs on a high-density farm. J. Virol. 2011, 85, 11697. [Google Scholar] [CrossRef]
- Brnić, D.; Prpić, J.; Keros, T.; Roić, B.; Starešina, V.; Jemeršić, L. Porcine astrovirus viremia and high genetic variability in pigs on large holdings in Croatia. Infect. Genet. Evol. 2013, 14, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Arruda, B.; Arruda, P.; Hensch, M.; Chen, Q.; Zheng, Y.; Yang, C.; Irh, G.; Ferreyra, F.M.; Gauger, P.; Schwartz, K. Porcine astrovirus type 3 in central nervous system of swine with polioencephalomyelitis. Emerg. Infect. Dis. 2017, 23, 2097–2100. [Google Scholar] [CrossRef] [PubMed]
- Boros, Á.; Albert, M.; Pankovics, P.; Bíró, H.; Pesavento, P.A.; Phan, T.G.; Delwart, E.; Reuter, G. Outbreaks of neuroinvasive astrovirus associated with encephalomyelitis, weakness, and paralysis among weaned pigs, Hungary. Emerg. Infect. Dis. 2017, 23, 1982–1993. [Google Scholar] [CrossRef] [PubMed]
- Behling-kelly, E.; Schultz-cherry, S.; Koci, M.; Kelley, L.; Larsen, D.; Brown, C. Localization of astrovirus in experimentally infected turkeys as determined by in situ hybridization. Vet. Pathol. 2002, 39, 595–598. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Wang, F.; Shi, J.; Zheng, L.; Wang, X.; Zhang, D. Molecular characterization of a duck hepatitis virus 3-like astrovirus. Vet. Microbiol. 2014, 170, 39–47. [Google Scholar] [CrossRef]
- Cordey, S.; Vu, D.L.; Schibler, M.; L’Huillier, A.G.; Brito, F.; Docquier, M.; Posfay-Barbe, K.M.; Petty, T.J.; Turin, L.; Zdobnov, E.M. Astrovirus mlb2, a new gastroenteric virus associated with meningitis and disseminated infection. Emerg. Infect. Dis. 2016, 22, 846–853. [Google Scholar] [CrossRef]
- Brown, J.R.; Morfopoulou, S.; Hubb, J.; Emmett, W.A.; Ip, W.; Shah, D.; Brooks, T.; Paine, S.M.L.; Anderson, G.; Virasami, A. Astrovirus va1/hmo-c: An increasingly recognized neurotropic pathogen in immunocompromised patients. Clin. Infect. Dis. 2015, 60, 881–888. [Google Scholar] [CrossRef]
- Selimovic-Hamza, S.; Boujon, C.L.; Hilbe, M.; Oevermann, A.; Seuberlich, T. Frequency and pathological phenotype of bovine astrovirus ch13/neuros1 infection in neurologically-diseased cattle: Towards assessment of causality. Viruses 2017, 9, 12. [Google Scholar] [CrossRef]
- Blomström, A.L.; Widén, F.; Hammer, A.S.; Belák, S.; Berg, M. Detection of a novel astrovirus in brain tissue of mink suffering from shaking mink syndrome by use of viral metagenomics. J. Clin. Microbiol. 2010, 48, 4392–4396. [Google Scholar] [CrossRef]
- Qin, Y.; Fang, Q.; Liu, H.; Ji, C.; Chen, Y.; Ouyang, K.; Wei, Z.; Huang, W. Construction of a reverse genetic system for porcine astrovirus. Arch. Virol. 2018, 163, 1511–1518. [Google Scholar] [CrossRef]
- McCarty, K.S., Jr.; Miller, L.S.; Cox, E.B.; Konrath, J.; McCarty, K.S., Sr. Estrogen receptor analyses. Correlation of biochemical and immunohistochemical methods using monoclonal antireceptor antibodies. Arch. Pathol. Lab. Med. 1985, 109, 716–721. [Google Scholar] [PubMed]
- Luo, Z.; Roi, S.; Dastor, M.; Gallice, E.; Laurin, M.A.; L’Homme, Y. Multiple novel and prevalent astroviruses in pigs. Vet. Microbiol. 2011, 149, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Monini, M.; Bartolo, I.D.; Ianiro, G.; Angeloni, G.; Magistrali, C.F.; Ostanello, F.; Ruggeri, F.M. Detection and molecular characterization of zoonotic viruses in swine fecal samples in Italian pig herds. Arch. Virol. 2015, 160, 2547–2556. [Google Scholar] [CrossRef] [PubMed]
- Machnowska, P.; Ellerbroek, L.; Johne, R. Detection and characterization of potentially zoonotic viruses in faeces of pigs at slaughter in Germany. Vet. Microbiol. 2014, 168, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Kumthip, K.; Khamrin, P.; Saikruang, W.; Kongkaew, A.; Vachirachewin, R.; Ushijima, H.; Maneekarn, N. Detection and genetic characterization of porcine astroviruses in piglets with and without diarrhea in Thailand. Arch. Virol. 2018, 163, 1823–1829. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Jeoung, H.Y.; Park, H.R.; Lim, J.A.; Song, J.Y.; An, D.J. Phylogenetic analysis of porcine astrovirus in domestic pigs and wild boars in South Korea. Virus Genes 2013, 46, 175–181. [Google Scholar] [CrossRef]
- Xiao, C.T.; Luo, Z.; Lv, S.L.; Opriessnig, T.; Li, R.C.; Yu, X.L. Identification and characterization of multiple porcine astrovirus genotypes in Hunan province, China. Arch. Virol. 2016, 162, 1–10. [Google Scholar] [CrossRef]
- Snodgrass, D.R.; Gray, E.W. Detection and transmission of 30 nm virus particles (astroviruses) in faeces of lambs with diarrhoea. Arch. Virol. 1977, 55, 287–291. [Google Scholar] [CrossRef]
- Tzipori, S.; Menzies, J.D.; Gray, E.W. Detection of astrovirus in the faeces of red deer. Vet. Rec. 1981, 108, 286. [Google Scholar] [CrossRef]
- Woode, G.N.; Bridger, J.C. Isolation of small viruses resembling astroviruses and caliciviruses from acute enteritis of calves. J. Med Microbiol. 1978, 11, 441–452. [Google Scholar] [CrossRef]
- Bosch, A.; Guix, S.; Pintó, R.M. Epidemiology of Human Astroviruses; Springer: New York, NY, USA, 2012. [Google Scholar] [CrossRef]
- Méndez-Toss, M.; Griffin, D.D.; Calva, J.; Contreras, J.F.; Puerto, F.I.; Mota, F.; Guiscafré, H.; Cedillo, R.; Muñoz, O.; Herrera, I. Prevalence and genetic diversity of human astroviruses in Mexican children with symptomatic and asymptomatic infections. J. Clin. Microbiol. 2004, 42, 151. [Google Scholar] [CrossRef] [PubMed]
- Koci, M.D.; Moser, L.A.; Kelley, L.A.; Diane, L.; Brown, C.C.; Stacey, S.C. Astrovirus induces diarrhea in the absence of inflammation and cell death. J. Virol. 2003, 77, 11798–11808. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, C.C.; Loh, J.T.; Zhao, G.; Duan, E.; Freiden, P.; Heath, R.J.; Schultz-Cherry, S.; Stappenbeck, T.S.; Wang, D.; Huang, H.V. Adaptive immunity restricts replication of novel murine astroviruses. J. Virol. 2012, 86, 12262–12270. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.L.; Cordey, S.; Brito, F.; Kaiser, L. Novel human astroviruses: Novel human diseases? J. Clin. Virol. 2016, 82, 56–63. [Google Scholar] [CrossRef]
- Quan, P.L.; Wagner, T.A.; Briese, T.; Torgerson, T.R.; Hornig, M.; Tashmukhamedova, A.; Firth, C.; Palacios, G.; Baisre-De-Leon, A.; Paddock, C.D. Astrovirus encephalitis in boy with X-linked agammaglobulinemia. Emerg. Infect. Dis. 2010, 16, 918–925. [Google Scholar] [CrossRef]
- Bouzalas, I.G.; Daniel, W.; Julia, W.; Cord, D.G.; Andreas, Z.; Marc, V.; Anna, O.; Rémy, B.; Torsten, S. Neurotropic astrovirus in cattle with nonsuppurative encephalitis in Europe. J. Clin. Microbiol. 2014, 52, 3318–3324. [Google Scholar] [CrossRef]
- Schlottau, K.; Schulze, C.; Bilk, S.; Hanke, D.; Höper, D.; Beer, M.; Hoffmann, B. Detection of a Novel Bovine Astrovirus in a Cow with Encephalitis. Transbound. Emerg. Dis. 2016, 63, 253–259. [Google Scholar] [CrossRef]
- Pfaff, F.; Schlottau, K.; Scholes, S.; Courtenay, A.; Hoffmann, B.; Höper, D.; Beer, M. A novel astrovirus associated with encephalitis and ganglionitis in domestic sheep. Transbound. Emerg. Dis. 2017, 64, 677–682. [Google Scholar] [CrossRef]
- Marvin, S.A.; Huerta, C.T.; Sharp, B.; Freiden, P.; Cline, T.D.; Schultz-Cherry, S. Type I interferon response limits astrovirus replication and protects against increased barrier permeability in vitro and in vivo. J. Virol. 2016, 90, 1988–1996. [Google Scholar] [CrossRef]
- Guix, S.; Perez-Bosque, A.; Miro, L.; Moreto, M.; Bosch, A.; Pinto, R.M. Type I interferon response is delayed in human astrovirus infections. PLoS ONE 2015, 10, e0123087. [Google Scholar] [CrossRef]
- Wang, D.; Fang, L.; Liu, L.; Zhong, H.; Chen, Q.; Luo, R.; Liu, X.; Zhang, Z.; Chen, H.; Xiao, S. Foot-and-mouth disease virus (FMDV) leader proteinase negatively regulates the porcine interferon-lambda1 pathway. Mol. Immunol. 2011, 49, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Meliopoulos, V.A.; Marvin, S.A.; Freiden, P.; Moser, L.A.; Nighot, P.; Ali, R.; Blikslager, A.; Reddivari, M.; Heath, R.J.; Koci, M.D. Oral administration of astrovirus capsid protein is sufficient to induce acute diarrhea in vivo. Mbio 2016, 7, e01494-16. [Google Scholar] [CrossRef] [PubMed]
- Moser, L.A.; Michael, C.; Stacey, S.C. Astrovirus increases epithelial barrier permeability independently of viral replication. J. Virol. 2007, 81, 11937–11945. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| dpi | PAstV-Challenged | Control |
|---|---|---|
| 2 d | Stomach bloat, gastric capillary congestion, small intestine bloat containing yellow foam liquid, mesenteric congestion | Mildly flatulence in individual pigs of the small intestine |
| 4 d | Stomach bloat, gastric capillary congestion, small intestine bloat containing yellow foam liquid, mesenteric congestion, cecum slightly flatulent | No lesions |
| 7 d | Yellow liquid contained in individual pigs of the small intestine | No lesions |
| 10 d | No lesions | No lesions |
| PAstV-Challenged | Control | |
|---|---|---|
| Duodenum | 0.42 ± 0.19 | 0 |
| Jejunum | 1.17 ± 0.24 | 0.73 ± 0.20 |
| Ileum | 0.83 ± 0.21 | 0.36 ± 0.15 |
| cecum | 0.17 ± 0.11 | 0 |
| colon | 0.08 ± 0.08 | 0.09 ± 0.09 |
| rectum | 0 | 0 |
| mesenteric lymph | 0.5 ± 0.15 | 0 |
| spleen | 0 | 0 |
| ung | 0 | 0 |
| kidney | 0 | 0 |
| stomach | 0.5 ± 0.15 | 0.27 ± 0.14 |
| dpi | Duodenum | Jejunum | Ileum | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | Challenged | p | Control | Challenged | p | Control | Challenged | p | ||
| 2 | Villous height (μm) | 544.9 ± 15.88 | 447.7 ± 25.82 | 0.038 (*) | 461.6 ± 27.97 | 268.1 ± 27.97 | 0.0001 (***) | 344.9 ± 47.67 | 278.6 ± 47.67 | 0.2328 (NS) |
| Crypt depth (μm) | 147.1 ± 8.644 | 180.7 ± 13.21 | 0.0328 (*) | 147.0 ± 4.974 | 171.5 ± 9.240 | 0.0161 (*) | 121.6 ± 6.506 | 179.5 ± 22.36 | 0.002 (*) | |
| Ratio (μm/μm) | 3.704 ± 1.828 | 2.478 ± 1.955 | 3.140 ± 5.623 | 1.563 ± 0.0003 | 2.836 ± 7.327 | 1.558 ± 1.21 | ||||
| 4 | Villous height (μm) | 513.9 ± 68.77 | 274.1 ± 16.22 | 0.0003 (***) | 445.4 ± 47.62 | 260.9 ± 14.73 | 0.0003 (***) | 333.7 ± 11.59 | 264 ± 14.76 | 0.0001 (***) |
| Crypt depth (μm) | 168.3 ± 5.602 | 190.4 ± 11.12 | 0.1936 (NS) | 162.6 ± 7.897 | 161.3 ± 6.637 | 0.917 (NS) | 122.5 ± 6.575 | 145.6 ± 11.06 | 0.0649 (NS) | |
| Ratio (μm/μm) | 3.053 ± 12.276 | 1.440 ± 1.459 | 2.739 ± 6.03 | 1.617 ± 2.219 | 2.734 ± 1.763 | 1.813 ± 1.335 | ||||
| 7 | Villous height (μm) | 456.5 ± 32.36 | 420.1 ± 24.66 | 0.4361 (NS) | 442.3 ± 29.53 | 451.6 ± 40.22 | 0.8539 (NS) | 380 ± 22.29 | 310.2 ± 22.29 | 0.0728 (NS) |
| Crypt depth (μm) | 176.7 ± 5.904 | 182.0 ± 14.98 | 0.8042 (NS) | 162.6 ± 15.35 | 177.6 ± 11.22 | 0.4573 (NS) | 157.3 ± 6.014 | 138.8 ± 11.52 | 0.1380 (NS) | |
| Ratio (μm/μm) | 2.583 ± 5.481 | 2.308 ± 1.646 | 2.720 ± 1.924 | 2.543 ± 3.585 | 2.416 ± 3.706 | 2.235 ± 1.935 | ||||
| 10 | Villous height (μm) | 510.1 ± 9.831 | 485.4 ± 10.76 | 0.1181 (NS) | 312.6 ± 16.88 | 336 ± 17.5 | 0.3513 (NS) | 394.9 ± 8.937 | 367.8 ± 21.71 | 0.3243 (NS) |
| Crypt depth (μm) | 188.4 ± 14.14 | 211.7 ± 14.12 | 0.5219 (NS) | 194.3 ± 7.024 | 192.4 ± 8.213 | 0.8751 (NS) | 158 ± 16.27 | 162.6 ± 8.119 | 0.7906 (NS) | |
| Ratio (μm/μm) | 2.571 ± 0.695 | 2.293 ± 0.762 | 1.609 ± 2.403 | 1.746 ± 2.131 | 2.429 ± 0.549 | 2.262 ± 2.674 | ||||
| dpi | Number of Sections | Duodenum | Jejunum | Ileum | Cecum | Colon | Rectum | Mesenteric Lymph |
|---|---|---|---|---|---|---|---|---|
| 2 | 3 | 6.67 ± 0.67 | 8.33 ± 2.33 | 9.00 ± 1.73 | 1.67 ± 0.33 | 2.00 ± 1.00 | 2.00 ± 1.00 | 7.33 ± 1.67 |
| 4 | 4 | 8.50 ± 1.66 | 6.25 ± 1.32 | 3.25 ± 0.75 | 1.25 ± 0.25 | 2.25 ± 1.32 | 1.13 ± 0.48 | 2.00 ± 1.35 |
| 7 | 3 | 5.00 ± 2.08 | 4.67 ± 1.33 | 2.33 ± 0.67 | 2.33 ± 0.88 | 1.67 ± 0.88 | 1.33 ± 0.67 | 1.33 ± 0.33 |
| 10 | 3 | 4.00 ± 2.65 | 2.00 ± 1.16 | 1.67 ± 1.20 | 1.00 ± 0.58 | 0.67 ± 0.33 | 0.67 ± 0.33 | 0.67 ± 0.33 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, Q.; Wang, C.; Liu, H.; Wu, Q.; Liang, S.; Cen, M.; Dong, Q.; Wei, Y.; Chen, Y.; Ouyang, K.; et al. Pathogenic Characteristics of a Porcine Astrovirus Strain Isolated in China. Viruses 2019, 11, 1156. https://doi.org/10.3390/v11121156
Fang Q, Wang C, Liu H, Wu Q, Liang S, Cen M, Dong Q, Wei Y, Chen Y, Ouyang K, et al. Pathogenic Characteristics of a Porcine Astrovirus Strain Isolated in China. Viruses. 2019; 11(12):1156. https://doi.org/10.3390/v11121156
Chicago/Turabian StyleFang, Qingli, Cui Wang, Huan Liu, Qingping Wu, Siying Liang, Minli Cen, Qinting Dong, Yingyi Wei, Ying Chen, Kang Ouyang, and et al. 2019. "Pathogenic Characteristics of a Porcine Astrovirus Strain Isolated in China" Viruses 11, no. 12: 1156. https://doi.org/10.3390/v11121156
APA StyleFang, Q., Wang, C., Liu, H., Wu, Q., Liang, S., Cen, M., Dong, Q., Wei, Y., Chen, Y., Ouyang, K., Wei, Z., & Huang, W. (2019). Pathogenic Characteristics of a Porcine Astrovirus Strain Isolated in China. Viruses, 11(12), 1156. https://doi.org/10.3390/v11121156