Identification of a Novel Adélie Penguin Circovirus at Cape Crozier (Ross Island, Antarctica)


 ,
,  , ,
, , 

{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
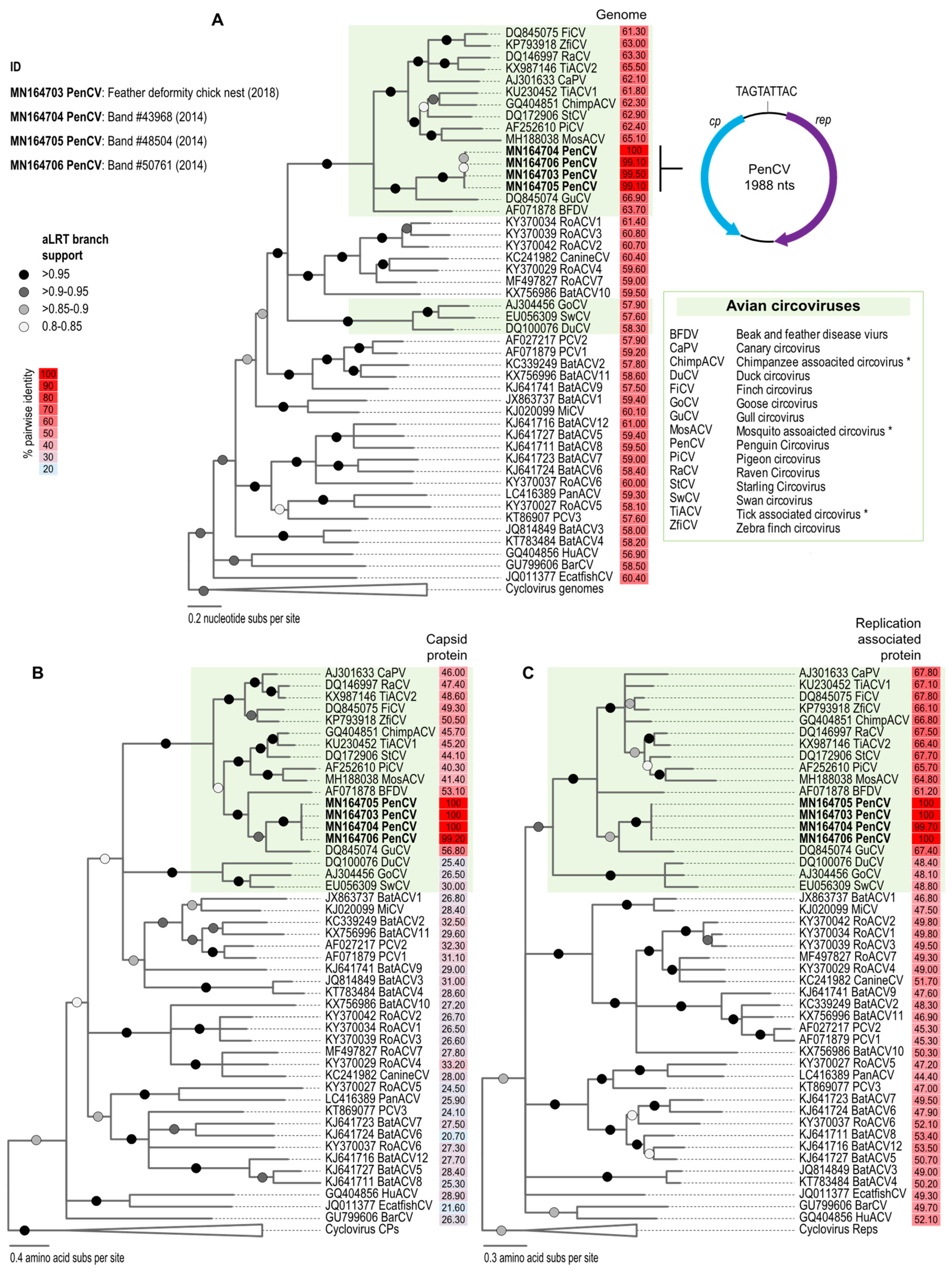
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kane, O.J.; Smith, J.R.; Boersma, P.D.; Parsons, N.J.; Strauss, V.; Garcia-Borboroglu, P.; Villanueva, C. Feather-Loss Disorder in African and Magellanic Penguins. Waterbirds 2010, 33, 415–421. [Google Scholar] [CrossRef]
- Barbosa, A.; Colominas-Ciuró, R.; Coria, N.; Centurión, M.; Sandler, R.; Negri, A.; Santos, M. First record of feather-loss disorder in Antarctic penguins. Antarct. Sci. 2014, 27, 69–70. [Google Scholar] [CrossRef]
- Lyver, P.O.; Barron, M.; Barton, K.J.; Ainley, D.G.; Pollard, A.; Gordon, S.; McNeill, S.; Ballard, G.; Wilson, P.R. Trends in the breeding population of Adelie penguins in the Ross Sea, 1981–2012: A coincidence of climate and resource extraction effects. PLoS ONE 2014, 9, e91188. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lynch, H.J.; LaRue, M.A. First global census of the Adélie Penguin. Auk 2014, 131, 457–466. [Google Scholar] [CrossRef]
- Grimaldi, W.W.; Hall, R.J.; White, D.D.; Wang, J.; Massaro, M.; Tompkins, D.M. First report of a feather loss condition in Adelie penguins (Pygoscelis adeliae) on Ross Island, Antarctica, and a preliminary investigation of its cause. Emu 2016, 115, 185–189. [Google Scholar] [CrossRef]
- Smeele, Z.E.; Ainley, D.G.; Varsani, A. Viruses associated with Antarctic wildlife: From serology based detection to identification of genomes using high throughput sequencing. Virus Res. 2018, 243, 91–105. [Google Scholar] [CrossRef]
- Phalen, D.N. Papillomaviruses and Polyomaviruses. In Infectious Diseases of Wild Birds; Thomas, N.J., Hunter, D.B., Atkinson, C.T., Eds.; Blackwell Publishing: Melbourne, VIC, Australia, 2007. [Google Scholar]
- Paré, J.A.; Robert, N. Circovirus. In Infectious Diseases of Wild Birds; Thomas, N.J., Hunter, D.B., Atkinson, C.T., Eds.; Blackwell Publishing: Melbourne, VIC, Australia, 2007. [Google Scholar]
- Varsani, A.; Porzig, E.L.; Jennings, S.; Kraberger, S.; Farkas, K.; Julian, L.; Massaro, M.; Ballard, G.; Ainley, D.G. Identification of an avian polyomavirus associated with Adelie penguins (Pygoscelis adeliae). J. Gen. Virol. 2015, 96, 851–857. [Google Scholar] [CrossRef]
- Rosario, K.; Breitbart, M.; Harrach, B.; Segales, J.; Delwart, E.; Biagini, P.; Varsani, A. Revisiting the taxonomy of the family Circoviridae: Establishment of the genus Cyclovirus and removal of the genus Gyrovirus. Arch. Virol. 2017, 162, 1447–1463. [Google Scholar] [CrossRef]
- Ritchie, B.W.; Niagro, F.D.; Lukert, P.D.; Steffens, W.L.; Latimer, K.S. Characterization of a New Virus from Cockatoos with Psittacine Beak and Feather Disease. Virology 1989, 171, 83–88. [Google Scholar] [CrossRef]
- Morozov, I.; Sirinarumitr, T.; Sorden, S.D.; Halbur, P.G.; Morgan, M.K.; Yoon, K.J.; Paul, P.S. Detection of a novel strain of porcine circovirus in pigs with postweaning multisystemic wasting syndrome. J. Clin. Microbiol. 1998, 36, 2535–2541. [Google Scholar]
- Hamel, A.L.; Lin, L.L.; Nayar, G.P.S. Nucleotide sequence of porcine circovirus associated with postweaning multisystemic wasting syndrome in pigs. J. Virol. 1998, 72, 5262–5267. [Google Scholar] [PubMed]
- Grau-Roma, L.; Fraile, L.; Segalés, J. Recent advances in the epidemiology, diagnosis and control of diseases caused by porcine circovirus type 2. Vet. J. 2011, 187, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Doneley, R.J. Acute beak and feather disease in juvenile African Grey parrots—An uncommon presentation of a common disease. Aust. Vet. J. 2003, 81, 206–207. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, B.W.; Niagro, F.D.; Lukert, P.D.; Latimer, K.S.; Steffens III, W.L.; Pritchard, N. A Review of Psittacine Beak and Feather Disease: Characteristics of the PBFD Virus. J. Assoc. Avian Vet. 1989, 3, 143–149. [Google Scholar] [CrossRef]
- McOrist, S.; Black, D.G.; Pass, D.A.; Scott, P.C.; Marshall, J. Beak and feather dystrophy in wild sulphur-crested cockatoos (Cacatua galerita). J. Wildl. Dis 1984, 20, 120–124. [Google Scholar] [CrossRef]
- Huff, D.G.; Schmidt, R.E.; Fudge, A.M. Psittacine beak and feather syndrome in a blue-fronted Amazon (Amazona aestiva). AAV Today 1988, 2, 84–86. [Google Scholar] [CrossRef]
- Ritchie, B.W.; Niagro, F.D.; Latimer, K.S.; Lukert, P.D.; Steffens, W.; Rakich, P.M.; Pritchard, N. Ultrastructural, protein composition, and antigenic comparison of psittacine beak and feather disease virus purified from four genera of psittacine birds. J. Wildl. Dis. 1990, 26, 196–203. [Google Scholar] [CrossRef]
- Pass, D.A.; Perry, R.A. The pathology of psittacine beak and feather disease. Aust. Vet. J. 1984, 61, 69–74. [Google Scholar] [CrossRef]
- Raidal, S.R. Viral skin diseases of birds. Semin. Avian Exot. Pet Med. 1995, 4, 72–82. [Google Scholar] [CrossRef]
- Ritchie, B.W.; Niagro, F.D.; Latimer, K.S.; Steffens, W.L.; Pesti, D.; Ancona, J.; Lukert, P.D. Routes and Prevalence of Shedding of Psittacine Beak and Feather Disease Virus. Am. J. Vet. Res. 1991, 52, 1804–1809. [Google Scholar]
- Rahaus, M.; Desloges, N.; Probst, S.; Loebbert, B.; Lantermann, W.; Wolff, M.H. Detection of beak and feather disease virus DNA in embryonated eggs of psittacine birds. Vet. Med. Czech. 2008, 53, 53–58. [Google Scholar] [CrossRef]
- Soike, D.; Kohler, B.; Albrecht, K. A circovirus-like infection in geese related to a runting syndrome. Avian Pathol. 1999, 28, 199–202. [Google Scholar] [CrossRef]
- Soike, D.; Albrecht, K.; Hattermann, K.; Schmitt, C.; Mankertz, A. Novel circovirus in mulard ducks with developmental and feathering disorders. Vet. Rec. 2004, 154, 792–793. [Google Scholar] [CrossRef] [PubMed]
- Todd, D.; Weston, J.; Ball, N.W.; Borghmans, B.J.; Smyth, J.A.; Gelmini, L.; Lavazza, A. Nucleotide sequence-based identification of a novel circovirus of canaries. Avian Pathol. 2001, 30, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Shivaprasad, H.L.; Hill, D.; Todd, D.; Smyth, J.A. Circovirus infection in a Gouldian finch (Chloebia gouldiae). Avian Pathol. 2004, 33, 525–529. [Google Scholar] [CrossRef]
- Mysore, J.; Read, D.; Daft, B. Circovirus-like particles in finches. In Proceedings of the Annual Conference of the American Association of Veterinary Laboratory Diagnosis, Histopathology Section. Reno, NV, USA, 18–23 October 2007. [Google Scholar]
- Circella, E.; Legretto, M.; Pugliese, N.; Caroli, A.; Bozzo, G.; Accogli, G.; Lavazza, A.; Camarda, A. Psittacine beak and feather disease-like illness in Gouldian finches (Chloebia gouldiae). Avian Dis. 2014, 58, 482–487. [Google Scholar] [CrossRef]
- Raidal, S.R.; Riddoch, P.A. A feather disease in Senegal doves (Streptopelia senegalensis) morphologically similar to psittacine beak and feather disease. Avian Pathol. 1997, 26, 829–836. [Google Scholar] [CrossRef]
- Pass, D.A.; Plant, S.L.; Sexton, N. Natural infection of wild doves (Streptopelia senegalensis) with the virus of psittacine beak and feather disease. Aust. Vet. J. 1994, 71, 307–308. [Google Scholar] [CrossRef]
- Woods, L.W.; Latimer, K.S. Circovirus infection of nonpsittacine birds. J. Avian Med. Surg. 2000, 14, 154–163. [Google Scholar] [CrossRef]
- Fahsbender, E.; Burns, J.M.; Kim, S.; Kraberger, S.; Frankfurter, G.; Eilers, A.A.; Shero, M.R.; Beltran, R.; Kirkham, A.; McCorkell, R.; et al. Diverse and highly recombinant anelloviruses associated with Weddell seals in Antarctica. Virus Evol. 2017, 3. [Google Scholar] [CrossRef]
- Varsani, A.; Kraberger, S.; Jennings, S.; Porzig, E.L.; Julian, L.; Massaro, M.; Pollard, A.; Ballard, G.; Ainley, D.G. A novel papillomavirus in Adelie penguin (Pygoscelis adeliae) faeces sampled at the Cape Crozier colony, Antarctica. J. Gen. Virol. 2014, 95, 1352–1365. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Posada, D. Selection of models of DNA evolution with jModelTest. Methods Mol. Biol. 2009, 537, 93–112. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast selection of best-fit models of protein evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef]
- Anisimova, M.; Gascuel, O. Approximate likelihood-ratio test for branches: A fast, accurate, and powerful alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef]
- Stover, B.C.; Muller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef] [PubMed]
- MacLachlan, N.J.; Dubovi, E.J. Circoviridae and Anelloviridae. In Fenner’s Veterinary Virology; MacLachlan, N.J., Dubovi, E.J., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 259–268. [Google Scholar]
- Fogell, D.J.; Martin, R.O.; Bunbury, N.; Lawson, B.; Sells, J.; McKeand, A.M.; Tatayah, V.; Trung, C.T.; Groombridge, J.J. Trade and conservation implications of new beak and feather disease virus detection in native and introduced parrots. Conserv. Biol. 2018, 32, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Ha, H.J.; Anderson, I.L.; Alley, M.R.; Springett, B.P.; Gartrell, B.D. The prevalence of beak and feather disease virus infection in wild populations of parrots and cockatoos in New Zealand. N. Z. Vet. J. 2007, 55, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Jackson, B.; Lorenzo, A.; Theuerkauf, J.; Barnaud, A.; Duval, T.; Guichard, P.; Bloc, H.; Baouma, A.; Stainton, D.; Kraberge, S.; et al. Preliminary surveillance for beak and feather disease virus in wild parrots of New Caledonia: Implications of a reservoir species for Ouvea Parakeets. Emu 2014, 114, 283–289. [Google Scholar] [CrossRef]
- Massaro, M.; Ortiz-Catedral, L.; Julian, L.; Galbraith, J.A.; Kurenbach, B.; Kearvell, J.; Kemp, J.; van Hal, J.; Elkington, S.; Taylor, G.; et al. Molecular characterisation of beak and feather disease virus (BFDV) in New Zealand and its implications for managing an infectious disease. Arch. Virol. 2012, 157, 1651–1663. [Google Scholar] [CrossRef]
- Ortiz-Catedral, L.; McInnes, K.; Hauber, M.E.; Brunton, D.H. First report of beak and feather disease virus (BFDV) in wild Red-fronted Parakeets (Cyanoramphus novaezelandiae) in New Zealand. Emu 2009, 109, 244–247. [Google Scholar] [CrossRef]
- Dugger, K.M.; Ainley, D.G.; Lyver, P.O.; Barton, K.; Ballard, G. Survival differences and the effect of environmental instability on breeding dispersal in an Adelie penguin meta-population. Proc. Natl. Acad. Sci. USA 2010, 107, 12375–12380. [Google Scholar] [CrossRef]
- Dugger, K.M.; Ballard, G.; Ainley, D.G.; Lyver, P.O.B.; Schine, C. Adelie penguins coping with environmental change: Results from a natural experiment at the edge of their breeding range. Front. Ecol. Evol. 2014, 2. [Google Scholar] [CrossRef]
- Ainley, D. The Adélie Penguin: Bellwether of Climate Change; Columbia University Press: New York, NY, USA, 2002. [Google Scholar]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morandini, V.; Dugger, K.M.; Ballard, G.; Elrod, M.; Schmidt, A.; Ruoppolo, V.; Lescroël, A.; Jongsomjit, D.; Massaro, M.; Pennycook, J.; et al. Identification of a Novel Adélie Penguin Circovirus at Cape Crozier (Ross Island, Antarctica). Viruses 2019, 11, 1088. https://doi.org/10.3390/v11121088
Morandini V, Dugger KM, Ballard G, Elrod M, Schmidt A, Ruoppolo V, Lescroël A, Jongsomjit D, Massaro M, Pennycook J, et al. Identification of a Novel Adélie Penguin Circovirus at Cape Crozier (Ross Island, Antarctica). Viruses. 2019; 11(12):1088. https://doi.org/10.3390/v11121088
Chicago/Turabian StyleMorandini, Virginia, Katie M. Dugger, Grant Ballard, Megan Elrod, Annie Schmidt, Valeria Ruoppolo, Amélie Lescroël, Dennis Jongsomjit, Melanie Massaro, Jean Pennycook, and et al. 2019. "Identification of a Novel Adélie Penguin Circovirus at Cape Crozier (Ross Island, Antarctica)" Viruses 11, no. 12: 1088. https://doi.org/10.3390/v11121088
APA StyleMorandini, V., Dugger, K. M., Ballard, G., Elrod, M., Schmidt, A., Ruoppolo, V., Lescroël, A., Jongsomjit, D., Massaro, M., Pennycook, J., Kooyman, G. L., Schmidlin, K., Kraberger, S., Ainley, D. G., & Varsani, A. (2019). Identification of a Novel Adélie Penguin Circovirus at Cape Crozier (Ross Island, Antarctica). Viruses, 11(12), 1088. https://doi.org/10.3390/v11121088