Pseudomonas aeruginosa PA5oct Jumbo Phage Impacts Planktonic and Biofilm Population and Reduces Its Host Virulence


 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacteriophages Propagation and Purification
2.2. Phage Host Range and Phage Receptor Analysis
2.3. Airway Surface Liquid Infection Model
2.4. Biofilm Eradication Analysis on Nephrophane Membrane
2.5. Isolation of Phage Resistant Clones from Treated Biofilm
2.6. Pyocyanin and Pyoverdine Production
2.7. Twitching Motility Assay
2.8. Lipopolysaccharide Structure Patterns Analysis
2.9. Growth Rate Measurement
2.10. TLR Stimulation Profile in THP1-XBlue™ Cell Line, NF-κB/AP-1-Reporter Monocytes
2.11. Galleria Mellonella Larvae Infection Model
2.12. PA5oct Phage DNA Detection in Bacterial Clones
2.13. Real-Time PCR Quantification of Bacterial versus Phage DNA Copies Increment over Time
3. Results
3.1. The Host Range of Phage PA5oct Suggests an Increased Activity against Clinical CF Isolates
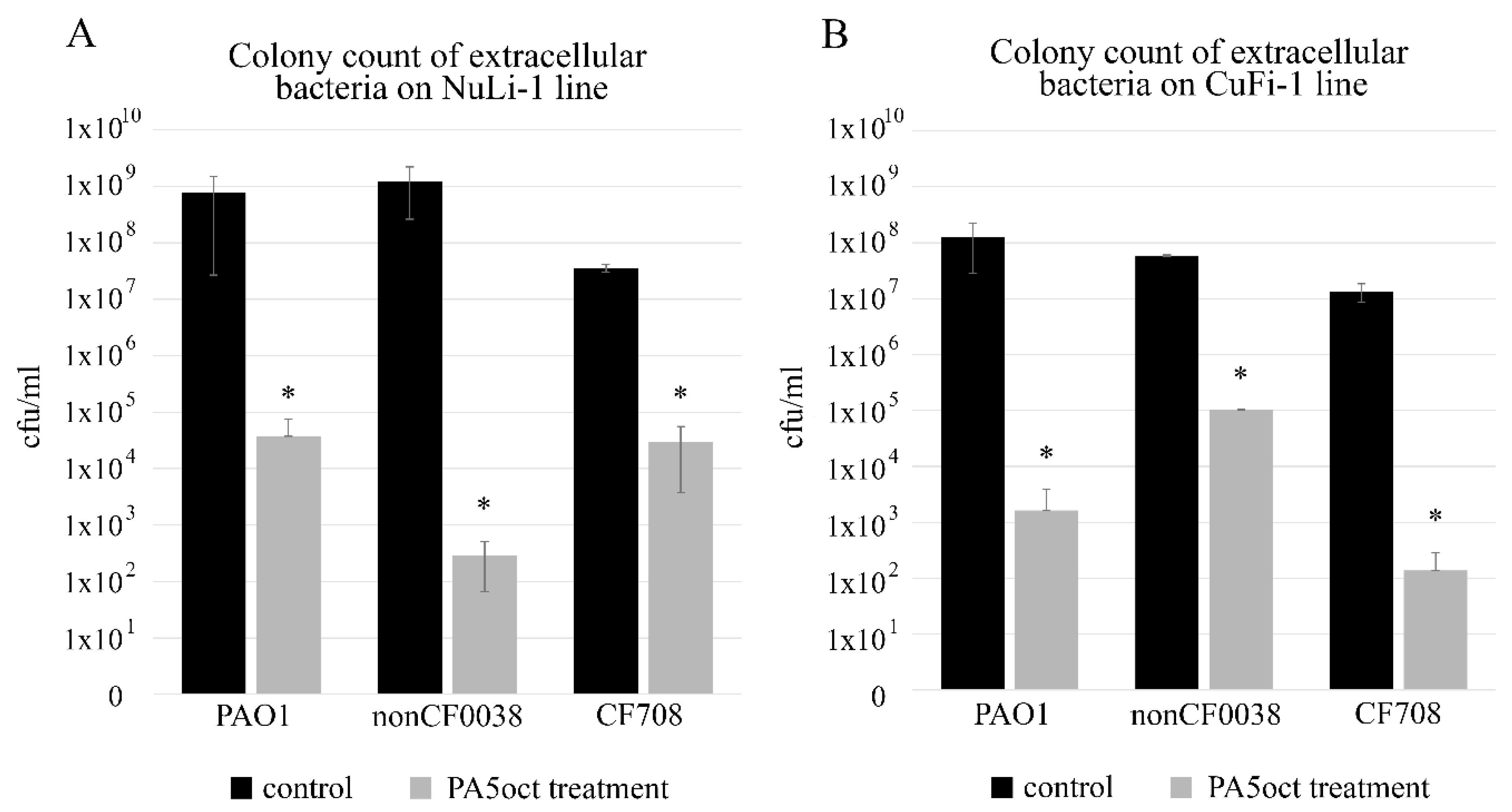
3.2. Phage PA5oct Infection Causes Significant Reduction of Planktonic/Cell Line-Associated Bacteria in an Airway Surface Liquid Infection Model
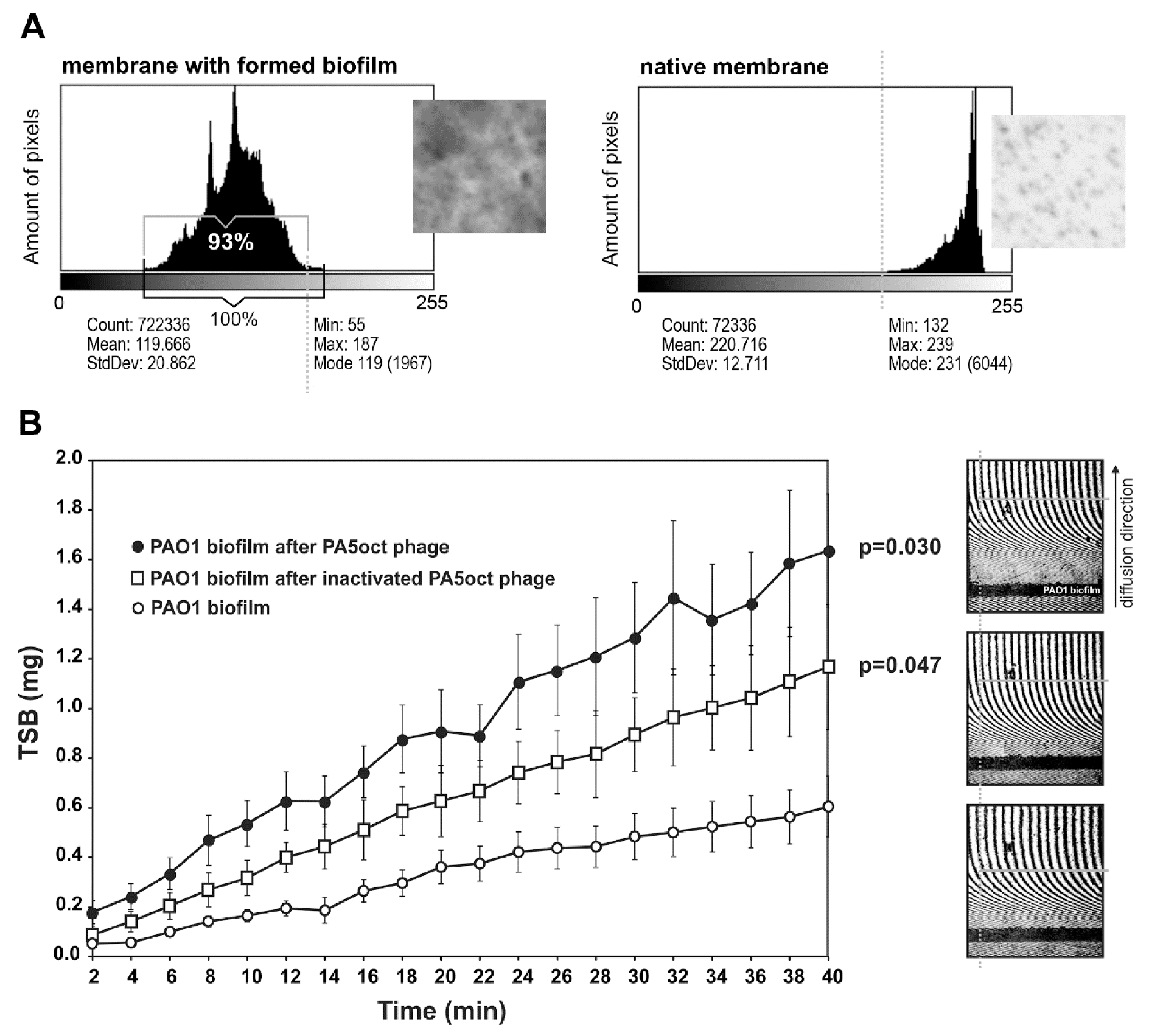
3.3. Real Time Measurement of P. aeruginosa Biofilm Diffusion Properties
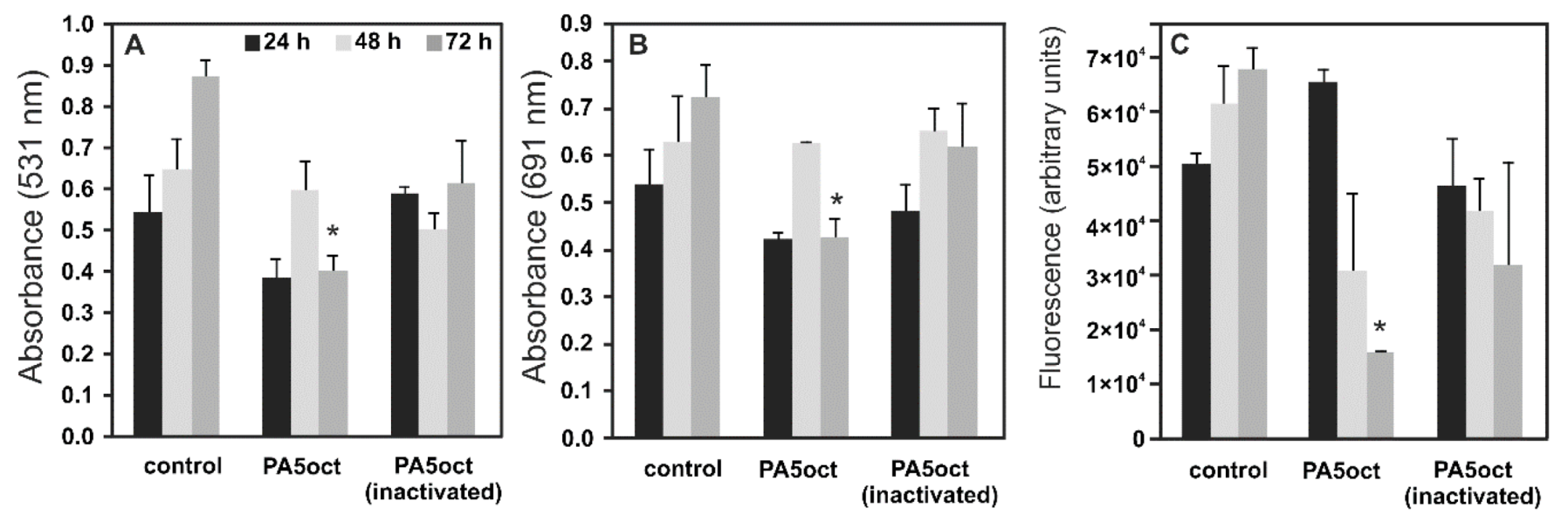
3.4. Phage PA5oct Impact on PAO1 Biofilm-Living Population after Infection
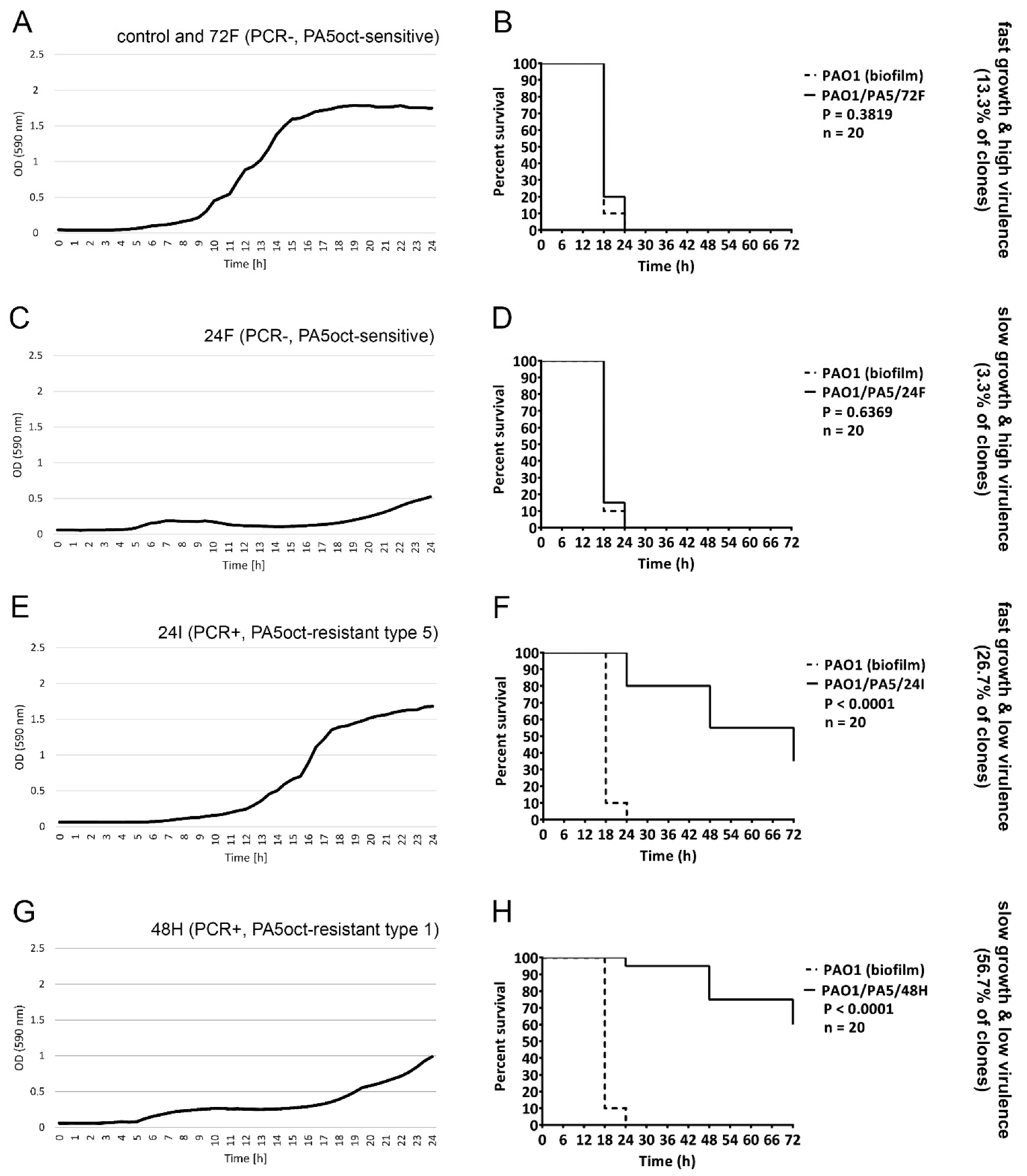
3.5. Emerging Phage PA5oct-Resistant Clones Showed a Reduced Virulence
3.6. The Reduced Virulence in PA5oct-Resistant Clones Is Correlated with the Persistence of Phage in Bacterial Population
3.7. The Presence of PA5oct in PAO1 Population Induce Pro-Inflammatory Response in Monocytes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Azam, A.H.; Tanji, Y. Bacteriophage-host arm race: An update on the mechanism of phage resistance in bacteria and revenge of the phage with the perspective for phage therapy. Appl. Microbiol. Biotechnol. 2019, 103, 2121–2131. [Google Scholar] [CrossRef] [PubMed]
- Latka, A.; Maciejewska, B.; Majkowska-Skrobek, G.; Briers, Y.; Drulis-Kawa, Z. Bacteriophage-encoded virion-associated enzymes to overcome the carbohydrate barriers during the infection process. Appl. Microbiol. Biotechnol. 2017, 101, 3103–3119. [Google Scholar] [CrossRef] [PubMed]
- Wittebole, X.; De Roock, S.; Opal, S.M. A historical overview of bacteriophage therapy as an alternative to antibiotics for the treatment of bacterial pathogens. Virulence 2014, 5, 226–235. [Google Scholar] [CrossRef]
- Forti, F.; Roach, D.R.; Cafora, M.; Pasini, M.E.; Horner, D.S.; Fiscarelli, E.V.; Rossitto, M.; Cariani, L.; Briani, F.; Debarbieux, L.; et al. Design of a broad-range bacteriophage cocktail that reduces Pseudomonas aeruginosa biofilms and treats acute infections in two animal models. Antimicrob. Agents Chemother. 2018, 62, e02573-17. [Google Scholar] [CrossRef]
- Galtier, M.; De Sordi, L.; Sivignon, A.; de Vallée, A.; Maura, D.; Neut, C.; Rahmouni, O.; Wannerberger, K.; Darfeuille-Michaud, A.; Desreumaux, P.; et al. Bacteriophages targeting adherent invasive Escherichia coli strains as a promising new treatment for Crohn’s disease. J. Crohn’s Colitis 2017, 11, 840–847. [Google Scholar] [CrossRef]
- Roach, D.R.; Leung, C.Y.; Henry, M.; Morello, E.; Singh, D.; Di Santo, J.P.; Weitz, J.S.; Debarbieux, L. Synergy between the host immune system and bacteriophage is essential for successful phage therapy against an acute respiratory pathogen. Cell Host Microbe 2017, 22, 38–47.e4. [Google Scholar] [CrossRef]
- Pirnay, J.-P.; Verbeken, G.; Ceyssens, P.-J.; Huys, I.; De Vos, D.; Ameloot, C.; Fauconnier, A. The Magistral phage. Viruses 2018, 10, 64. [Google Scholar] [CrossRef]
- Hendrix, R.W. Jumbo bacteriophages. Curr. Top. Microbiol. Immunol. 2009, 328, 229–240. [Google Scholar]
- Krylov, V.; Shaburova, O.; Krylov, S.; Pleteneva, E. A genetic approach to the development of new therapeutic phages to fight Pseudomonas aeruginosa in wound infections. Viruses 2012, 5, 15–53. [Google Scholar] [CrossRef]
- Yuan, Y.; Gao, M. Jumbo bacteriophages: An overview. Front. Microbiol. 2017, 8, 403. [Google Scholar] [CrossRef] [PubMed]
- Monson, R.; Foulds, I.; Foweraker, J.; Welch, M.; Salmond, G.P.C. The Pseudomonas aeruginosa generalized transducing phage phiPA3 is a new member of the phiKZ-like group of “jumbo” phages, and infects model laboratory strains and clinical isolates from cystic fibrosis patients. Microbiology 2011, 157, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Serwer, P. The conformation of DNA packaged in bacteriophage G. Biophys. J. 1997, 72, 958–963. [Google Scholar] [CrossRef]
- Lecoutere, E.; Ceyssens, P.J.; Miroshnikov, K.A.; Mesyanzhinov, V.V.; Krylov, V.N.; Noben, J.P.; Robben, J.; Hertveldt, K.; Volckaert, G.; Lavigne, R. Identification and comparative analysis of the structural proteomes of φKZ and EL, two giant Pseudomonas aeruginosa bacteriophages. Proteomics 2009, 9, 3215–3219. [Google Scholar] [CrossRef]
- Mesyanzhinov, V.V.; Robben, J.; Grymonprez, B.; Kostyuchenko, V.A.; Bourkaltseva, M.V.; Sykilinda, N.N.; Krylov, V.N.; Volckaert, G. The genome of bacteriophage φKZ of Pseudomonas aeruginosa. J. Mol. Biol. 2002, 317, 1–19. [Google Scholar] [CrossRef]
- Fokine, A.; Battisti, A.J.; Bowman, V.D.; Efimov, A.V.; Kurochkina, L.P.; Chipman, P.R.; Mesyanzhinov, V.V.; Rossmann, M.G. Cryo-EM Study of the Pseudomonas bacteriophage φKZ. Structure 2007, 15, 1099–1104. [Google Scholar] [CrossRef]
- Ceyssens, P.-J.; Minakhin, L.; Van den Bossche, A.; Yakunina, M.; Klimuk, E.; Blasdel, B.; De Smet, J.; Noben, J.-P.; Bläsi, U.; Severinov, K.; et al. Development of giant bacteriophage phiKZ is independent of the host transcription apparatus. J. Virol. 2014, 88, 10501–10510. [Google Scholar] [CrossRef]
- Chaikeeratisak, V.; Nguyen, K.; Khanna, K.; Brilot, A.F.; Erb, M.L.; Coker, J.K.C.; Vavilina, A.; Newton, G.L.; Buschauer, R.; Pogliano, K.; et al. Assembly of a nucleus-like structure during viral replication in bacteria. Science (80-.) 2017, 355, 194–197. [Google Scholar] [CrossRef]
- Krylov, V.N.; Dela Cruz, D.M.; Hertveldt, K.; Ackermann, H.-W. “φKZ-like viruses”, a proposed new genus of myovirus bacteriophages. Arch. Virol. 2007, 152, 1955–1959. [Google Scholar] [CrossRef]
- Danis-Wlodarczyk, K.; Vandenheuvel, D.; Jang, H.B.; Briers, Y.; Olszak, T.; Arabski, M.; Wasik, S.; Drabik, M.; Higgins, G.; Tyrrell, J.; et al. A proposed integrated approach for the preclinical evaluation of phage therapy in Pseudomonas infections. Sci. Rep. 2016, 6, 28115. [Google Scholar] [CrossRef]
- Drulis-Kawa, Z.; Olszak, T.; Danis, K.; Majkowska-Skrobek, G.; Ackermann, H.-W. A giant Pseudomonas phage from Poland. Arch. Virol. 2014, 159, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Lood, C.; Danis-Wlodarczyk, K.; Blasdel, B.; Jang, H.B.; Vandenheuvel, D.; Briers, Y.; Noben, J.-P.; van Noort, V.; Drulis-Kawa, Z.; Lavigne, R. Integrative omics analysis of Pseudomonas aeruginosa virus PA5oct highlights the molecular complexity of jumbo phages. bioRxiv 2019, 679506. [Google Scholar] [CrossRef]
- Danis-Wlodarczyk, K.; Olszak, T.; Arabski, M.; Wasik, S.; Majkowska-Skrobek, G.; Augustyniak, D.; Gula, G.; Briers, Y.; Jang, H.B.; Vandenheuvel, D.; et al. Characterization of the newly isolated lytic bacteriophages KTN6 and KT28 and their efficacy against Pseudomonas aeruginosa biofilm. PLoS ONE 2015, 10, e0127603. [Google Scholar]
- Ceyssens, P.-J.; Lavigne, R.; Mattheus, W.; Chibeu, A.; Hertveldt, K.; Mast, J.; Robben, J.; Volckaert, G. Genomic analysis of Pseudomonas aeruginosa phages LKD16 and LKA1: Establishment of the KMV subgroup within the T7 supergroup. J. Bacteriol. 2006, 188, 6924–6931. [Google Scholar] [CrossRef]
- Ceyssens, P.-J.; Mesyanzhinov, V.; Sykilinda, N.; Briers, Y.; Roucourt, B.; Lavigne, R.; Robben, J.; Domashin, A.; Miroshnikov, K.; Volckaert, G.; et al. The genome and structural proteome of YuA, a new Pseudomonas aeruginosa phage resembling M6. J. Bacteriol. 2008, 190, 1429–1435. [Google Scholar] [CrossRef]
- Adams, M.H. Enumeration of phage particles. In Bacteriophages; Adams, M.H., Ed.; Interscience Publishers: New York, NY, USA, 1959; pp. 27–30. [Google Scholar]
- Kutter, E. Phage host range and efficiency of plating. Methods Mol. Biol. 2009, 501, 141–149. [Google Scholar]
- Zabner, J.; Karp, P.; Seiler, M.; Phillips, S.L.; Mitchell, C.J.; Saavedra, M.; Welsh, M.; Klingelhutz, A.J. Development of cystic fibrosis and noncystic fibrosis airway cell lines. Am. J. Physiol. Cell. Mol. Physiol. 2003, 284, L844–L854. [Google Scholar] [CrossRef]
- Turnbull, L.; Whitchurch, C.B. Motility assay: twitching motility. In Pseudomonas Methods and Protocols; Filloux, A., Ramos, J.-L., Eds.; Humana Press Inc.: New York, NY, USA, 2014; pp. 73–86. [Google Scholar]
- Marolda, C.L.; Welsh, J.; Dafoe, L.; Valvano, M.A. Genetic analysis of the O7-polysaccharide biosynthesis region from the Escherichia coli O7:K1 strain VW187. J. Bacteriol. 1990, 172, 3590–3599. [Google Scholar] [CrossRef]
- Latino, L.; Caroff, M.; Pourcel, C. Fine structure analysis of lipopolysaccharides in bacteriophage-resistant Pseudomonas aeruginosa PAO1 mutants. Microbiology (United Kingdom) 2017, 163, 848–855. [Google Scholar] [CrossRef]
- Lee, C.H.; Tsai, C.M. Quantification of bacterial lipopolysaccharides by the purpald assay: Measuring formaldehyde generated from 2-keto-3-deoxyoctonate and heptose at the inner core by periodate oxidation. Anal. Biochem. 1999, 267, 161–168. [Google Scholar] [CrossRef]
- Tsai, C.M.; Frasch, C.E. A sensitive silver stain for detecting lipopolysaccharides in polyacrylamide gels. Anal. Biochem. 1982, 119, 115–119. [Google Scholar] [CrossRef]
- Cullen, L.; Weiser, R.; Olszak, T.; Maldonado, R.; Moreira, A.; Slachmuylders, L.; Brackman, G.; Paunova-Krasteva, T.; Zarnowiec, P.; Czerwonka, G.; et al. Phenotypic characterization of an international Pseudomonas aeruginosa reference panel: Strains of cystic fibrosis (CF) origin show less in vivo virulence than non-CF strains. Microbiology 2015, 161, 1961–1977. [Google Scholar] [CrossRef] [PubMed]
- Olszak, T.; Zarnowiec, P.; Kaca, W.; Danis-Wlodarczyk, K.; Augustyniak, D.; Drevinek, P.; de Soyza, A.; McClean, S.; Drulis-Kawa, Z. In vitro and in vivo antibacterial activity of environmental bacteriophages against Pseudomonas aeruginosa strains from cystic fibrosis patients. Appl. Microbiol. Biotechnol. 2015, 99, 6021–6033. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Khlebodarova, T.M.; Likhoshvai, V.A. Persister cells—A plausible outcome of neutral coevolutionary drift. Sci. Rep. 2018, 8, 14309. [Google Scholar] [CrossRef] [PubMed]
- De Soyza, A.; Hall, A.J.; Mahenthiralingam, E.; Drevinek, P.; Kaca, W.; Drulis-Kawa, Z.; Stoitsova, S.R.; Toth, V.; Coenye, T.; Zlosnik, J.E.A.; et al. Developing an international Pseudomonas aeruginosa reference panel. Microbiologyopen 2013, 2, 1010–1023. [Google Scholar] [CrossRef]
- Lorè, N.I.; Cigana, C.; De Fino, I.; Riva, C.; Juhas, M.; Schwager, S.; Eberl, L.; Bragonzi, A. Cystic fibrosis-niche adaptation of Pseudomonas aeruginosa reduces virulence in multiple infection hosts. PLoS ONE 2012, 7, e35648. [Google Scholar] [CrossRef]
- Bradbury, R.S.; Reid, D.W.E.C.; Inglis, T.J.J.; Champion, A.C. Decreased virulence of cystic fibrosis Pseudomonas aeruginosa in Dictyostelium discoideum. Microbiol. Immunol. 2011, 55, 224–230. [Google Scholar] [CrossRef]
- Cigana, C.; Curcurù, L.; Leone, M.R.; Ieranò, T.; Lorè, N.I.; Bianconi, I.; Silipo, A.; Cozzolino, F.; Lanzetta, R.; Molinaro, A.; et al. Pseudomonas aeruginosa exploits lipid A and muropeptides modification as a strategy to lower innate immunity during cystic fibrosis lung infection. PLoS ONE 2009, 4, e8439. [Google Scholar] [CrossRef]
- Worlitzsch, D.; Tarran, R.; Ulrich, M.; Schwab, U.; Cekici, A.; Meyer, K.C.; Birrer, P.; Bellon, G.; Berger, J.; Weiss, T.; et al. Effects of reduced mucus oxygen concentration in airway Pseudomonas infections of cystic fibrosis patients. J. Clin. Invest. 2002, 109, 317–325. [Google Scholar] [CrossRef]
- Prince, A. Adhesins and receptors of Pseudomonas aeruginosa associated with infection of the respiratory tract. Microb. Pathog. 1992, 13, 251–260. [Google Scholar] [CrossRef]
- Saiman, L.; Tabibi, S.; Starner, T.D.; San Gabriel, P.; Winokur, P.L.; Jia, H.P.; McCray, J.; Tack, B.F. Cathelicidin peptides inhibit multiply antibiotic-resistant pathogens from patients with cystic fibrosis. Antimicrob. Agents Chemother. 2001, 45, 2838–2844. [Google Scholar] [CrossRef] [PubMed]
- Dean, S.N.; Bishop, B.M.; van Hoek, M.L. Susceptibility of Pseudomonas aeruginosa biofilm to alpha-helical peptides: D-enantiomer of LL-37. Front. Microbiol. 2011, 2, 128. [Google Scholar] [CrossRef] [PubMed]
- Dalcin, D.; Ulanova, M. The role of human beta-defensin-2 in Pseudomonas aeruginosa pulmonary infection in cystic pibrosis patients. Infect. Dis. Ther. 2013, 2, 159–166. [Google Scholar] [CrossRef][Green Version]
- Dechecchi, M.C.; Nicolis, E.; Bezzerri, V.; Vella, A.; Colombatti, M.; Assael, B.M.; Mettey, Y.; Borgatti, M.; Mancini, I.; Gambari, R.; et al. MPB-07 reduces the inflammatory response to Pseudomonas aeruginosa in cystic fibrosis bronchial cells. Am. J. Respir. Cell Mol. Biol. 2007, 36, 615–624. [Google Scholar] [CrossRef]
- Dechecchi, M.C.; Nicolis, E.; Norez, C.; Bezzerri, V.; Borgatti, M.; Mancini, I.; Rizzotti, P.; Ribeiro, C.M.P.; Gambari, R.; Becq, F.; et al. Anti-inflammatory effect of miglustat in bronchial epithelial cells. J. Cyst. Fibros. 2008, 7, 555–565. [Google Scholar] [CrossRef]
- Abedon, S.T. Phage “delay” towards enhancing bacterial escape from biofilms: A more comprehensive way of viewing resistance to bacteriophages. AIMS Microbiol. 2017, 3, 186–226. [Google Scholar] [CrossRef]
- Abedon, S.T. Bacteriophage exploitation of bacterial biofilms: Phage preference for less mature targets? FEMS Microbiol. Lett. 2016, 363, fnv246. [Google Scholar] [CrossRef]
- Azeredo, J.; Sutherland, I. The use of phages for the removal of infectious biofilms. Curr. Pharm. Biotechnol. 2008, 9, 261–266. [Google Scholar] [CrossRef]
- Pires, D.P.; Oliveira, H.; Melo, L.D.R.; Sillankorva, S.; Azeredo, J. Bacteriophage-encoded depolymerases: Their diversity and biotechnological applications. Appl. Microbiol. Biotechnol. 2016, 100, 2141–2151. [Google Scholar] [CrossRef]
- Olszak, T.; Shneider, M.M.; Latka, A.; Maciejewska, B.; Browning, C.; Sycheva, L.V.; Cornelissen, A.; Danis-Wlodarczyk, K.; Senchenkova, S.N.; Shashkov, A.S.; et al. The O-specific polysaccharide lyase from the phage LKA1 tailspike reduces Pseudomonas virulence. Sci. Rep. 2017, 7, 16302. [Google Scholar] [CrossRef] [PubMed]
- Serwer, P.; Hayes, S.J.; Thomas, J.A.; Hardies, S.C. Propagating the missing bacteriophages: A large bacteriophage in a new class. Virol. J. 2007, 4, 21. [Google Scholar] [CrossRef] [PubMed]
- Le, S.; Yao, X.; Lu, S.; Tan, Y.; Rao, X.; Li, M.; Jin, X.; Wang, J.; Zhao, Y.; Wu, N.C.; et al. Chromosomal DNA deletion confers phage resistance to Pseudomonas aeruginosa. Sci. Rep. 2014, 4, 4738. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Zhang, H.; Shen, W.; Zou, Z.; Lu, S.; Li, G.; He, X.; Agnello, M.; Shi, W.; Hu, F.; et al. Pseudomonas aeruginosa MutL promotes large chromosomal deletions through non-homologous end joining to prevent bacteriophage predation. Nucleic Acids Res. 2018, 46, 4505–4514. [Google Scholar] [CrossRef]
- Latino, L.; Midoux, C.; Vergnaud, G.; Pourcel, C. Investigation of Pseudomonas aeruginosa strain PcyII-10 variants resisting infection by N4-like phage Ab09 in search for genes involved in phage adsorption. PLoS ONE 2019, 14, e0215456. [Google Scholar] [CrossRef]
- Loc-Carrillo, C.; Abedon, S.T. Pros and cons of phage therapy. Bacteriophage 2011, 1, 111–114. [Google Scholar] [CrossRef]
- Oechslin, F. Resistance development to bacteriophages occurring during bacteriophage therapy. Viruses 2018, 10, 351. [Google Scholar] [CrossRef]
- Cenens, W.; Makumi, A.; Govers, S.K.; Lavigne, R.; Aertsen, A. Viral transmission dynamics at single-cell resolution reveal transiently immune subpopulations caused by a carrier state association. PLoS Genet. 2015, 11, e1005770. [Google Scholar] [CrossRef]
- Latino, L.; Midoux, C.; Hauck, Y.; Vergnaud, G.; Pourcel, C. Pseudolysogeny and sequential mutations build multiresistance to virulent bacteriophages in Pseudomonas aeruginosa. Microbiology (UK) 2016, 162, 748–763. [Google Scholar] [CrossRef]
- Argov, T.; Azulay, G.; Pasechnek, A.; Stadnyuk, O.; Ran-Sapir, S.; Borovok, I.; Sigal, N.; Herskovits, A.A. Temperate bacteriophages as regulators of host behavior. Curr. Opin. Microbiol. 2017, 38, 81–87. [Google Scholar] [CrossRef]
- Cenens, W.; Mebrhatu, M.T.; Makumi, A.; Ceyssens, P.-J.; Lavigne, R.; Van Houdt, R.; Taddei, F.; Aertsen, A. Expression of a novel P22 ORFan gene reveals the phage carrier state in Salmonella Typhimurium. PLoS Genet. 2013, 9, e1003269. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowska, K. Phage therapy: What factors shape phage pharmacokinetics and bioavailability? Systematic and critical review. Med. Res. Rev. 2019, 39, 2000–2025. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PAO1 Isolates | Phenotype | PA5oct Activity |
|---|---|---|
| ATCC 15692 (PAO1) | Wild type | + |
| Δrmd (A−, B+) * | Deficiency in D-rhamnose biosynthesis; lack of A-band LPS | + |
| ΔwaaL (A−, B−) * | Lack of WaaL ligating O-polymer to core-lipid A; LPS is devoid of A-band and B-band, semirough (SR-LPS, or core-plus-one O-antigen) | −/+ |
| ΔwbpL (A−, B−) * | Lack of glucosyltransferase WbpL essential for initiation of both A-band and B-band synthesis | − |
| ΔfliC ΔalgC ΔpilA ** | Lack of flagella; lack of AlgC required for A-band, core oligosaccharide, and alginate biosynthesis; lack of Type IV pili | − |
| ΔfliC ΔpilA ** | Lack of flagella; lack of Type IV pili | − |
| ΔfliC WTpilA ** | Lack of flagella | −/+ |
| Bacterial Clones | Susceptibility to Phage Infection | Number of Isolates | |||||||
|---|---|---|---|---|---|---|---|---|---|
| LPS/Pili | LPS-Dependent | Pili-Dependent | |||||||
| PA5oct | LBL3 | KT28 | KTN6 | LUZ7 | KTN4 | phiKZ | LUZ19 | ||
| Control planktonic PAO1 | + | + | + | + | + | + | + | + | 1 |
| Control biofilm PAO1 | + | + | + | + | + | + | + | + | 30 |
| PA5oct sensitive | + | + | + | + | + | + | + | + | 10 |
| PA5oct resistant type 1 | − | − | + | + | + | − | + | + | 1 |
| PA5oct resistant type 2 | − | − | + | + | + | − | − | + | 2 |
| PA5oct resistant type 3 | − | + | − | − | + | − | + | + | 2 |
| PA5oct resistant type 4 | − | + | − | − | + | + | + | + | 5 |
| PA5oct resistant type 5 | − | − | − | − | + | + | + | + | 10 |
| Bacterial Clones | Name | LPS Pattern | Growth Rate [OD590/24 h] | Larvae Survival Rate [%] (18/24/48/72 h) | Twitching Motility [mm] | Phage DNA Presence (PCR) | ΔCT PAO1/ΔCT PA5oct Ratio |
|---|---|---|---|---|---|---|---|
| control planktonic | PAO1 | S | >1.5 | 10/0/0/0 | 24.2 ± 1.3 | − | − |
| control biofilm | K72A | S | >1.5 | 30/0/0/0 | 23 ± 2.0 | − | − |
| control biofilm | K72B | S | >1.5 | 30/0/0/0 | 22.9 ± 1.9 | − | − |
| control biofilm | K72C | S | >1.5 | 10/0/0/0 | 22.8 ± 2.3 | − | − |
| control biofilm | K72D | S | >1.5 | 5/0/0/0 | 22.7 ±1.5 | − | − |
| control biofilm | K72E | S | >1.5 | 15/0/0/0 | 23.5 ± 1.3 | − | − |
| control biofilm | K72F | S | >1.5 | 0/0/0/0 | 23.8 ± 1.2 | − | − |
| control biofilm | K72G | S | >1.5 | 5/0/0/0 | 23.7 ± 1.6 | − | − |
| control biofilm | K72H | S | >1.5 | 35/0/0/0 | 23.3 ± 2.1 | − | − |
| control biofilm | K72I | S | >1.5 | 15/0/0/0 | 23.8 ± 1.1 | − | − |
| control biofilm | K72J | S | >1.5 | 25/0/0/0 | 23.2 ± 1.5 | − | − |
| PA5oct sensitive | 24F | S | <1.0 | 15/0/0/0 | 18.2 ± 0.9 * | − | − |
| PA5oct sensitive | 48B | S | >1.5 | 20/10/0/0 | 18.5 ± 1.0 * | − | − |
| PA5oct sensitive | 48C | S | >1.5 | 80/35/0/0 ** | 19.3 ± 1.3 * | − | − |
| PA5oct sensitive | 48F | S | >1.5 | 30/0/0/0 | 15.7 ± 0.8 * | − | − |
| PA5oct sensitive | 48G | S | >1.5 | 80/40/15/0 ** | 19 ± 0.8 * | − | − |
| PA5oct sensitive | 72B | S | >1.5 | 30/25/10/0 ** | 18.4 ± 1.3 * | − | − |
| PA5oct sensitive | 72C | S | <1.0 | 80/55/10/0 ** | 15.5 ± 1.3 * | + | 1.7 |
| PA5oct sensitive | 72E | S | >1.5 | 55/25/0/0 ** | 14.6 ± 0.7 * | − | − |
| PA5oct sensitive | 72F | S | >1.5 | 20/0/0/0 | 18.3 ± 0.8 * | − | − |
| PA5oct sensitive | 72G | S | >1.5 | 30/15/0/0 | 18.5 ± 1.1 * | − | − |
| PA5oct resistant type 1 | 48H | S | <1.0 | 100/95/75/60 ** | 5.5 ± 1.3 * | + | 0.8 |
| PA5oct resistant type 2 | 24C | S | <1.0 | 75/60/55/35 ** | 15.4 ± 1.0 * | + | 0.8 |
| PA5oct resistant type 2 | 48A | S | <1.0 | 70/65/30/0 ** | 17.5 ± 1.8 * | + | 1.0 |
| PA5oct resistant type 3 | 48E | S | <1.0 | 100/45/0/0 ** | 19.1 ± 1.3 * | + | 0.9 |
| PA5oct resistant type 3 | 72H | S | <1.0 | 95/55/10/0 ** | 13.6 ± 1.6 * | + | 0.8 |
| PA5oct resistant type 4 | 24A | S | <1.0 | 50/15/15/15 ** | 16.7 ± 2.0 * | + | 2.0 |
| PA5oct resistant type 4 | 24D | S | >1.5 | 75/60/40/20 ** | 11 ± 2.3 * | + | 1.0 |
| PA5oct resistant type 4 | 48I | S | <1.0 | 85/60/20/0 ** | 13.2 ± 0.6 * | + | 0.9 |
| PA5oct resistant type 4 | 48J | S | <1.0 | 80/60/15/15 ** | 15.4 ± 1.2 * | + | 1.1 |
| PA5oct resistant type 4 | 72I | S | <1.0 | 55/50/5/0 ** | 14 ± 1.1 * | + | 1.2 |
| PA5oct resistant type 5 | 24B | S | <1.0 | 85/80/40/40 ** | 16.5 ± 1.6 * | + | 1.0 |
| PA5oct resistant type 5 | 24E | S | <1.0 | 75/70/55/30 ** | 15.4 ± 1.6 * | + | 1.0 |
| PA5oct resistant type 5 | 24G | S | <1.0 | 75/70/65/35 ** | 12.5 ± 2.1 * | + | 1.0 |
| PA5oct resistant type 5 | 24H | S | <1.0 | 95/75/45/35 ** | 12.2 ± 1.3 * | + | 0.8 |
| PA5oct resistant type 5 | 24I | S | >1.5 | 100/80/55/35 ** | 13.3 ± 0.8 * | + | 1.0 |
| PA5oct resistant type 5 | 24J | S | >1.5 | 85/70/50/30 ** | 14.5 ± 1.7 * | + | 1.3 |
| PA5oct resistant type 5 | 48D | S | <1.0 | 85/45/15/0 ** | 14.4 ± 0.7 * | + | 0.8 |
| PA5oct resistant type 5 | 72A | S | <1.0 | 95/70/0/0 ** | 12.5 ± 1.7 * | + | 1.0 |
| PA5oct resistant type 5 | 72D | S | >1.5 | 100/75/15/0 ** | 13.2 ± 0.9 * | + | 0.9 |
| PA5oct resistant type 5 | 72J | S | <1.0 | 95/80/10/0 ** | 22.8 ± 1.3 * | + | 1.0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olszak, T.; Danis-Wlodarczyk, K.; Arabski, M.; Gula, G.; Maciejewska, B.; Wasik, S.; Lood, C.; Higgins, G.; Harvey, B.J.; Lavigne, R.; et al. Pseudomonas aeruginosa PA5oct Jumbo Phage Impacts Planktonic and Biofilm Population and Reduces Its Host Virulence. Viruses 2019, 11, 1089. https://doi.org/10.3390/v11121089
Olszak T, Danis-Wlodarczyk K, Arabski M, Gula G, Maciejewska B, Wasik S, Lood C, Higgins G, Harvey BJ, Lavigne R, et al. Pseudomonas aeruginosa PA5oct Jumbo Phage Impacts Planktonic and Biofilm Population and Reduces Its Host Virulence. Viruses. 2019; 11(12):1089. https://doi.org/10.3390/v11121089
Chicago/Turabian StyleOlszak, Tomasz, Katarzyna Danis-Wlodarczyk, Michal Arabski, Grzegorz Gula, Barbara Maciejewska, Slawomir Wasik, Cédric Lood, Gerard Higgins, Brian J. Harvey, Rob Lavigne, and et al. 2019. "Pseudomonas aeruginosa PA5oct Jumbo Phage Impacts Planktonic and Biofilm Population and Reduces Its Host Virulence" Viruses 11, no. 12: 1089. https://doi.org/10.3390/v11121089
APA StyleOlszak, T., Danis-Wlodarczyk, K., Arabski, M., Gula, G., Maciejewska, B., Wasik, S., Lood, C., Higgins, G., Harvey, B. J., Lavigne, R., & Drulis-Kawa, Z. (2019). Pseudomonas aeruginosa PA5oct Jumbo Phage Impacts Planktonic and Biofilm Population and Reduces Its Host Virulence. Viruses, 11(12), 1089. https://doi.org/10.3390/v11121089