1. Introduction
Zika virus (
Flaviviridae;
Flavivirus; ZIKV) is a human pathogenic arthropod-borne virus (arbovirus) transmitted by mosquitoes. Originally discovered in 1947 in Uganda, the virus was introduced to Brazil in 2014 due to human activity [
1,
2,
3,
4]. In the following years, ZIKV caused major outbreaks among humans in South America, Central America, and in the Caribbean (reviewed in [
5]). Typical disease symptoms include febrile illness, rash, headache, and arthritis [
3]. Under certain circumstances, a fetus can develop severe neurological damage such as microcephaly when the mother has been infected with the virus [
6,
7,
8]. The rapid ZIKV outbreaks in the Western Hemisphere were fueled by the ubiquitous presence of the mosquito vectors,
Aedes aegypti and
Ae. albopictus, both transmitting strains of the virus in urban disease cycles, albeit with varying levels of efficiency [
9,
10,
11,
12].
An arthropod vector needs to become persistently infected with a virus in order to transmit the virus to a vertebrate host [
13]. A mosquito vector such as
Ae. aegypti acquires an arbovirus, i.e., ZIKV, along with a viremic bloodmeal from a vertebrate host (reviewed in [
14]). The bloodmeal enters the lumen of the mosquito midgut. Within a few hours (before the peritrophic matrix is formed) the virus has to enter midgut epithelial cells via receptor-mediated endocytosis. Flavivirus RNA replication and de novo synthesis of virions take place at the endoplasmic reticulum (ER) [
15,
16]. De novo synthesized virions, having a diameter of 40–50 nm [
17,
18], are then transported to the epithelial cells’ basal labyrinth in close proximity to the midgut basal lamina (BL). The BL is produced and secreted by the epithelial cells and constitutes a multi-stranded network of the extracellular matrix components lining the midgut epithelium [
19,
20]. Earlier studies indicated that flaviviruses, similar to alphaviruses [
21] exit the midgut by traversing the BL to disseminate to secondary tissues [
22,
23]. The secondary tissue tropism within the mosquito vector has been described in detail for the related flavivirus, dengue 2 virus (DENV2) [
24]. Following its exit from the mosquito midgut, DENV2 infects hemocytes, fat body, nerve tissue, and eventually the mosquito’s salivary glands. Once these are infected and virus gets released into the salivary ducts during salivation, the female mosquito is able to transmit the virus to vertebrate hosts for the remainder of her life.
Bloodmeal ingestion and digestion by the female mosquito strongly effect the structure and morphology of her midgut [
21,
25,
26,
27]. Most obvious is the distension of the posterior midgut, up to 20-fold. This distension causes a reduction in the density of microvilli and the stretching of the basal lamina, which appears now straightened. Within the initial 10–12 h post-bloodmeal (pbm), the basal labyrinth becomes enlarged. The peritrophic matrix is evident in the posterior region of the midgut. Around 20 h pbm, lipid inclusions and glycogen deposits become visible in the gut epithelium near the basal labyrinth. The time period from 36 h pbm onwards indicates the ending of the digestive cycle with lipid inclusions and glycogen deposits starting to disappear from the gut epithelium. By 72 h pbm, blood digestion has ended; however, the morphology of the posterior midgut does not entirely regain its original appearance. For example, cells that were joined before bloodmeal ingestion are now disconnected at their basal portion.
The basic components of the BL are type IV collagen, laminin, nidogen/entactin and proteoglycans of the perlecan type [
28]. As described in an ultrastructural analysis by Reinhardt and Hecker [
29], the midgut BL of female
Ae. aegypti forms a three-dimensional structure of hexagonal/cuboidal symmetry. Specifically, the BL consists of several stacked layers and each layer exhibits a grid-like superstructure. The width of the mesh formed by the grid-like structure of each BL layer was around 30 nm on average when analyzing the midguts of sugar-fed mosquitoes [
29]. The mesh width increased to around 48 nm directly after bloodmeal ingestion, and five days later, when the bloodmeal had been completely digested, the average mesh width became reduced (35 nm), however was still significantly larger than that of sugar-fed mosquitoes [
29]. The barrier character of the midgut BL became obvious for viruses when chikungunya virus (
Togaviridae;
Alphavirus; CHIKV) was intrathoracically injected into sugar-fed and blood-fed
Ae. aegypti (strain: Higgs’ White Eye, HWE) females [
25]. In seven-day old sugar-fed females (which had never received a bloodmeal before), injected virions, 60–70 nm in diameter [
30,
31], lined up at the basal side of the BL and seemed unable to traverse it [
25]. However, injection of CHIKV into females that had ingested a bloodmeal resulted in the presence and replication of the virus inside the midgut epithelium. Scanning electron microscopy (SEM) imaging showed that the outer surface of midguts obtained from artificially blood-fed females was severely ripped and distorted near the midgut-surrounding muscles, whereas the surface structure of midguts obtained from sugar-fed females looked intact [
21,
25]. In addition, we showed that substituting blood in the artificial meal for protein (based on bovine serum albumin [BSA]) or PBS (phosphate buffered saline, no nutrients) had no inhibitory effect on CHIKV dissemination [
25]. These combined observations led us to conclude that the presence of a (blood)meal in the midgut is temporally facilitating the dissemination of CHIKV from the organ as it causes the midgut BL to be overly distended thereby increasing its pore size exclusion limit.
Flaviviruses employ replication and virion assembly strategies that differ from those of alphaviruses (reviewed in [
32,
33,
34]). In a mosquito cell, flavivirus replication takes place within replication complexes residing within double-membrane vesicles, which are located at the membrane of the rough ER [
15]. Flaviviruses express their structural and non-structural proteins from the same RNA molecule [
32,
33]. De novo synthesized viral RNA forms a complex with the transmembrane capsid (C) protein. The capsid protein-RNA complex receives a lipid bilayer envelope originating from the ER containing the structural proteins envelope (E) and pre-membrane (prM). This results in the formation of an immature fusion-incompetent virion, which is budding into the ER lumen. When transported through the trans-Golgi network, E and prM undergo conformational changes forming a mature virion with prM molecules now covering the fusion loops of E. Via furin cleavage, prM is eventually cleaved into pr and M. Alphaviruses, by contrast, express their nonstructural and structural proteins from different viral RNA subunits and regulate their plus/minus strand RNA and protein synthesis in a temporal manner enabling them to quickly replicate to high titers [
32]. Virion assembly of alphaviruses within the infected cell involves a two-step assembly process [
34,
35]. Viral RNA replication is controlled by the non-structural proteins (nsP1-nsP4) and takes place within virus-induced replication factories consisting of spherules located at the plasma membrane and within large cytopathic vacuoles [
36]. Translation of the subgenomic RNA produces the structural polyprotein containing CP (capsid protein), E3, E2, 6K, and E1 [
34,
35]. CP is cleaved from the polyprotein and freely released into the cytoplasm to form the nucleocapsid together with de novo synthesized viral RNA released from the spherules. The glycoprotein precursors PE2 (including E2 and E3) and E1 are translocated across the ER and then processed and transported though the trans-Golgi network to the plasma membrane. There, the nucleocapsid, having bypassed the trans-Golgi network, associates with the glycoproteins, resulting in the assembly of mature virions, each now possessing 80 spikes consisting of trimers of E2 and E1 heterodimers [
32,
34,
36].
Generally, alphaviruses build up higher titers in a shorter time period than flaviviruses do in the same mosquito vector [
23,
37]. This prompted us to investigate whether the different infection dynamics between flaviviruses and alphaviruses are also leading to different midgut dissemination patterns for the two virus groups. Alternatively, the mechanics underlying distention and reconfiguration of the mosquito midgut/BL structure during bloodmeal digestion could perhaps dictate a common dissemination pattern for both virus groups. Here, we investigate the dissemination pattern of the flavivirus, ZIKV, from the mosquito midgut and discuss whether it differs from that previously described for the alphavirus, CHIKV [
21,
38]. In an in vivo growth curve, we revealed the time point when orally acquired ZIKV was detectable outside the mosquito midgut. In parallel, we observed infection patterns for the virus in situ using immunofluorescence assays and at the ultrastructural level via transmission electron microscopy (TEM). Furthermore, using a mixture of 5 nm and 50 nm gold nanoparticles, we show how meal ingestion into the midgut changed the size exclusion limit of the BL and discuss its relevance for the midgut dissemination of ZIKV. The effect of multiple bloodmeal ingestions by the mosquito on the intensity of infection was assessed for the flavivirus, dengue 4 virus (DENV4), at the ultrastructural level.
2. Materials and Methods
2.1. Mosquitoes and Viruses
The eye pigment deficient Higgs’ White Eye (HWE) strain of
Ae. aegypti [
39] was maintained in an insectary under a 12 h light/12 h dark cycle and a temperature and humidity regime of 28 °C and 80%, respectively. Larvae were reared at a moderate density (~100 larvae per shoe box size hatch pan and ~3 cm distilled water level) and fed with tropical fish food (Tetramin, Melle, Germany). One hundred emerged females and around 10 males were placed in a 64 oz. (~1 L) ice cream carton, which was covered with netting. Mosquitoes were supplied ad libitum with raisins and water. One-week post-emergence, females were used for experiments involving ZIKV, DENV4, or gold-nanoparticles.
ZIKV strain I-44 (GenBank #: KX856011), which was isolated in 2016 from mosquitoes captured in Chiapas State of Mexico was used in this study [
10,
40]. The virus was propagated for four days (until use in artificial blood-feeding experiments) in confluent Vero cells (ATCC #: CCL-81) at a multiplicity of infection (MOI) of 0.01. Vero cells were cultured in complete Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 7% fetal bovine serum (FBS). DENV4 strain H241 (GenBank #: KR011349) was isolated from a human patient in the Philippines in 1956 and represents the prototype for DENV serotype 4 [
41,
42].
Ae. albopictus C6/36 cells were cultivated at 28 °C without CO
2 supplement in Leibovitz L-15 medium supplemented with 7% FBS until 80%–90% confluency. Cells were then infected with DENV4 at a MOI of 0.001. Following virus inoculation, the cell culture medium was changed for fresh L-15 medium now containing 2% FBS and 1% non-essential amino acids (100x) (Corning Inc., Corning, NY, USA). Following 4–5 days of cultivation, the cell culture supernatant was harvested and used in artificial bloodmeals for oral infection of mosquitoes. All research work involving ZIKV and DENV4 was performed in the biosafety level 3 (BSL3) Virology Suite of the Laboratory for Infectious Disease Research (LIDR) at the University of Missouri.
2.2. ZIKV or DENV4 Infection of Mosquitoes via Artificial Bloodmeals
Twenty-four hours before blood-feeding, HWE females were deprived of their raisin food source and their water cups were removed 2–3 h prior to blood-feeding. ZIKV-containing Vero cell culture supernatant (at 96 h post-infection pi)) or DENV4-containing C6/36 cell culture supernatant (at 96–120 h pi) was mixed at a 1:1 ratio with defibrinated sheep blood (Colorado Serum Company, Denver, CO, USA) and 10 mM ATP. The artificial bloodmeal was pipetted into glass feeders (2 mL volume/feeder, one feeder/carton), which were covered with a hog gut membrane to be exposed to the mosquitoes in the carton for probing. The glass feeders were connected to a water jacket to maintain a temperature of 37 °C. Mosquitoes were allowed to feed for 1 h before fully engorged females were selected and maintained for further analysis in an environmental growth chamber at 28 °C and 80% relative humidity.
2.3. Plaque Assays for the Detection of ZIKV in Individual Mosquito Tissues
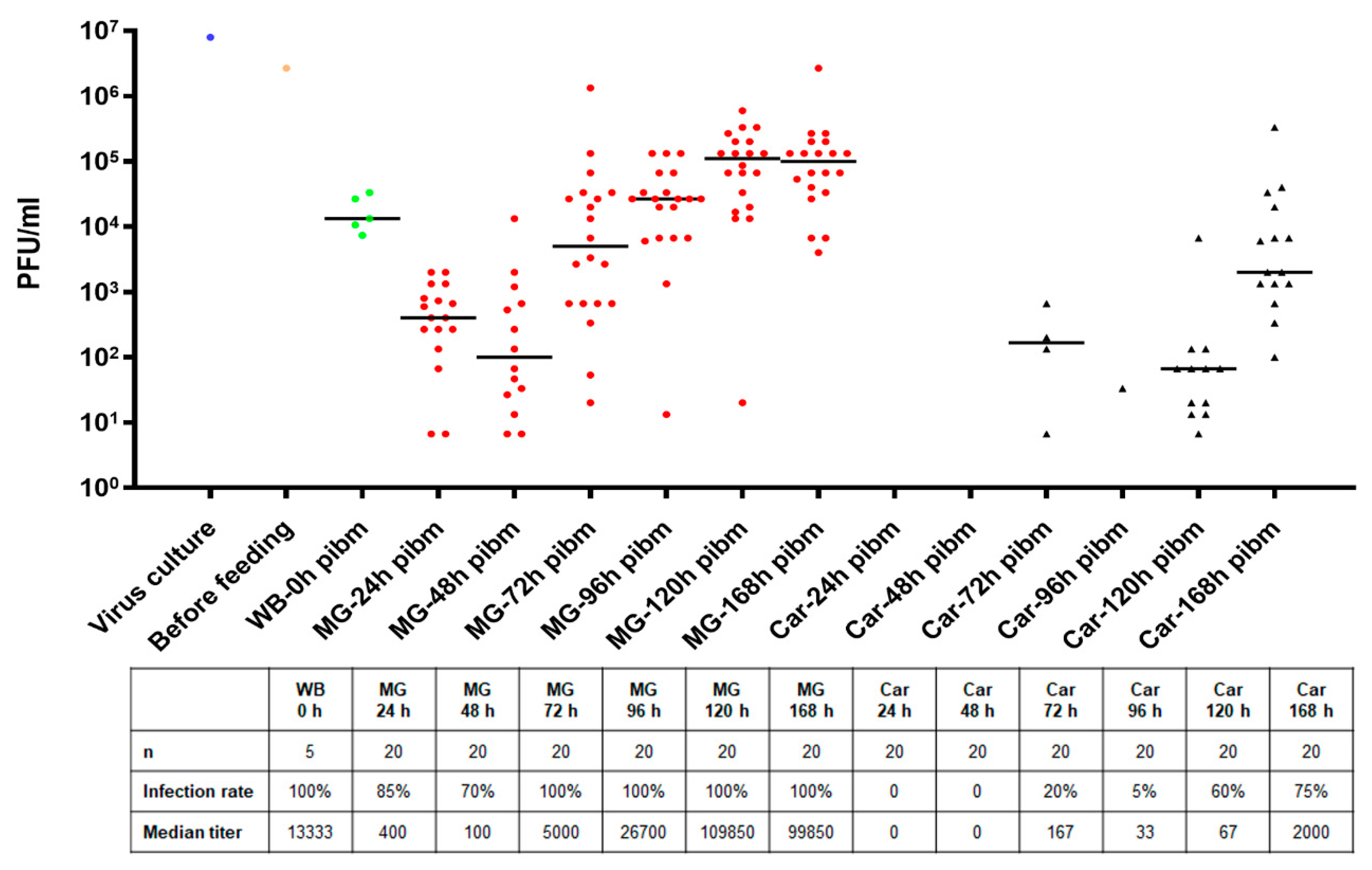
At 24, 48, 72, 96, 120, and 168 h post-infectious bloodmeal (pibm), midguts of blood-fed mosquitoes were dissected and separately maintained from the remaining body (carcass). To assess viral titers in individual tissues, midguts and carcasses were placed individually into 1.5 mL micro-centrifuge tubes containing 0.5–1.0 mL DMEM supplemented with 7% FBS, 5% HEPES and ground with a micro pestle before being filtrated using 0.22 µm Supor Membrane syringe filters (Pall Life Sciences, East Hills, NY, USA). Filtrated samples were 10-fold diluted and 150 µl of sample volume for each dilution step was used to inoculate ~90% confluent Vero cells seeded into 24-well plates. After a 1 h incubation period at 37 °C, inoculated Vero cells were overlaid with a 1% agarose-nutrient mixture containing 10% M199 (10x), 7% FBS, 0.5% MEM vitamin solution (100x), 0.5% MEM non-essential amino acid solution (100x), and 0.003% sodium bicarbonate solution (Gibco, ThermoFisher, Waltham, MA, USA). Following a four-day incubation period at 37 °C under 5% CO2 supplement, agarose containing plates were fixed with 10% formalin for 5 h. Thereafter, agarose ‘plugs’ were carefully removed without disturbing the Vero cell layer and cells were briefly stained with 0.2% crystal violet solution to visualize plaques. Plaques of each well were counted under a light microscope to calculate viral titers as plaque-forming units/mL (PFU/mL).
2.4. Detection of Flaviviral Antigen in Individual Mosquito Tissues by Immunofluorescence Assay (IFA)
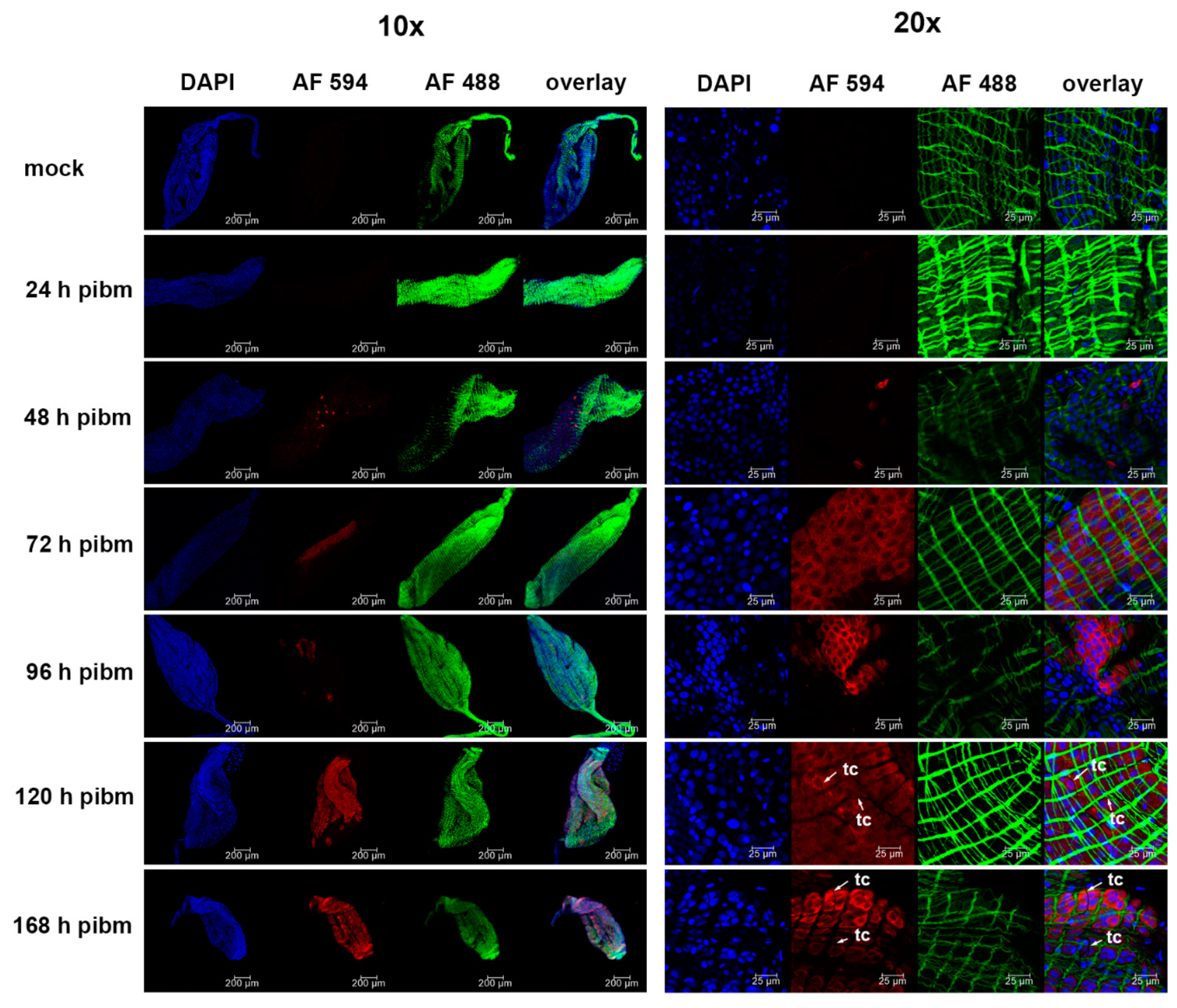
At 24, 48, 72, 96, 120, and 168 h pibm, midguts were dissected and fixed in 4% para-formaldehyde dissolved in 1x PBS. Fixed midguts were permeabilized using PBS-T (1x PBS containing 0.2% Triton X-100 and 1% BSA), before a 1 h incubation at room temperature (RT) with the primary monoclonal antibody Anti-Flavivirus Group Antigen D1-4G2-4-15 (ATCC #: VR-1852), diluted 1:500 in PBS-T. Following several washes with PBS-T, the secondary monoclonal antibody (goat anti-mouse IgG labeled with Alexa Fluor 594, Abcam # ab150120) was applied for 1 h at RT at a 1:500 dilution in PBS-T together with Alexa Fluor Phalloidin 488 (Invitrogen, Carlsbad, CA, USA) diluted 1:1000. Cell nuclei were stained with 1 µg/mL DAPI (Invitrogen) for 10 min. Midguts were then washed three times with PBS before being mounted on six-well printed slides, using Fluoromount G mounting medium (Electron Microcopy Sciences, Hatfield, PA, USA). Samples were viewed under an inverted spectral confocal microscope (TCP SP8 MP, Leica Microsystems, Wetzlar, Germany) at the Molecular Cytology Core of the University of Missouri.
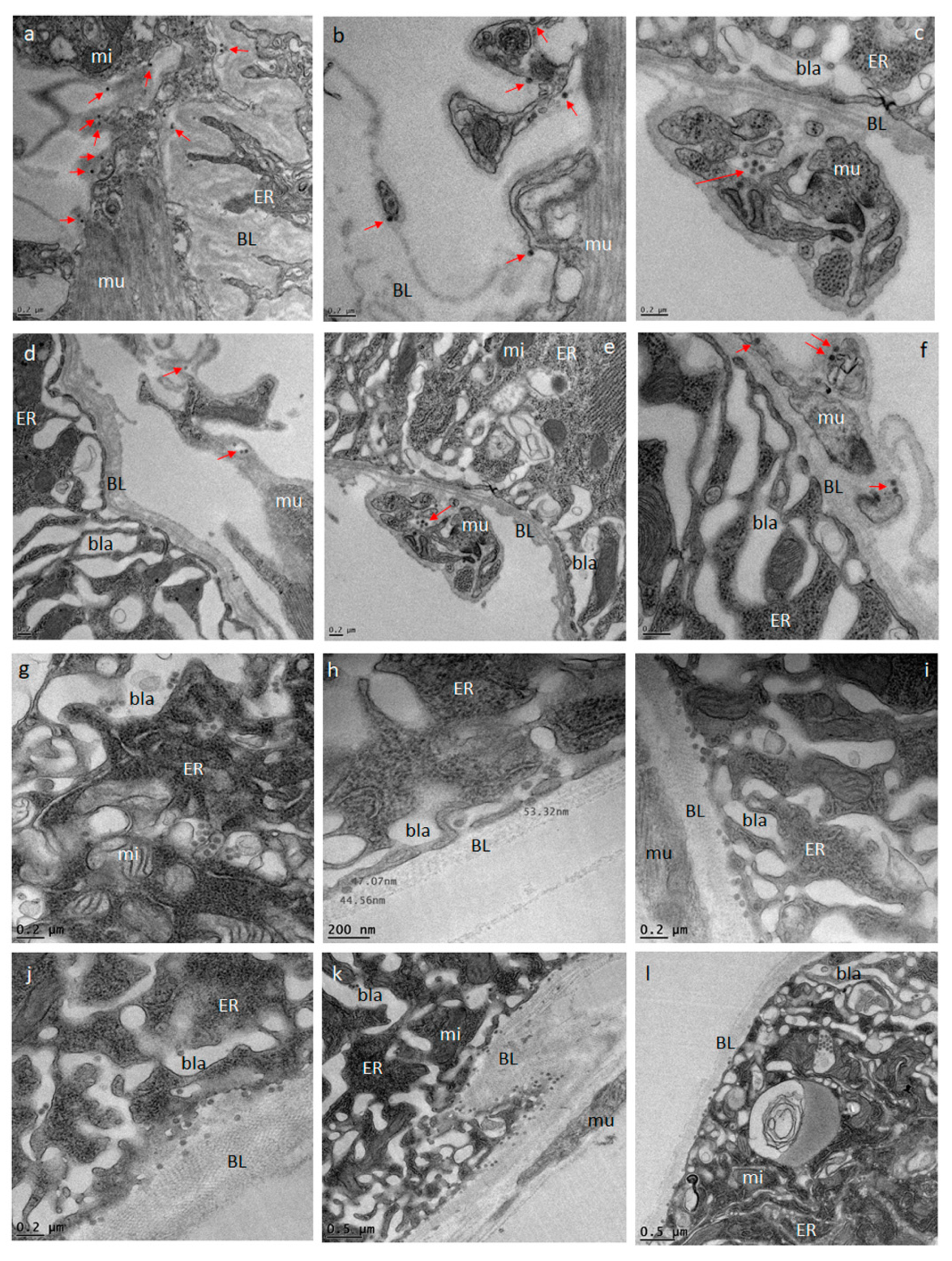
2.5. Detection of ZIKV and DENV4 in Midgut Tissue by TEM
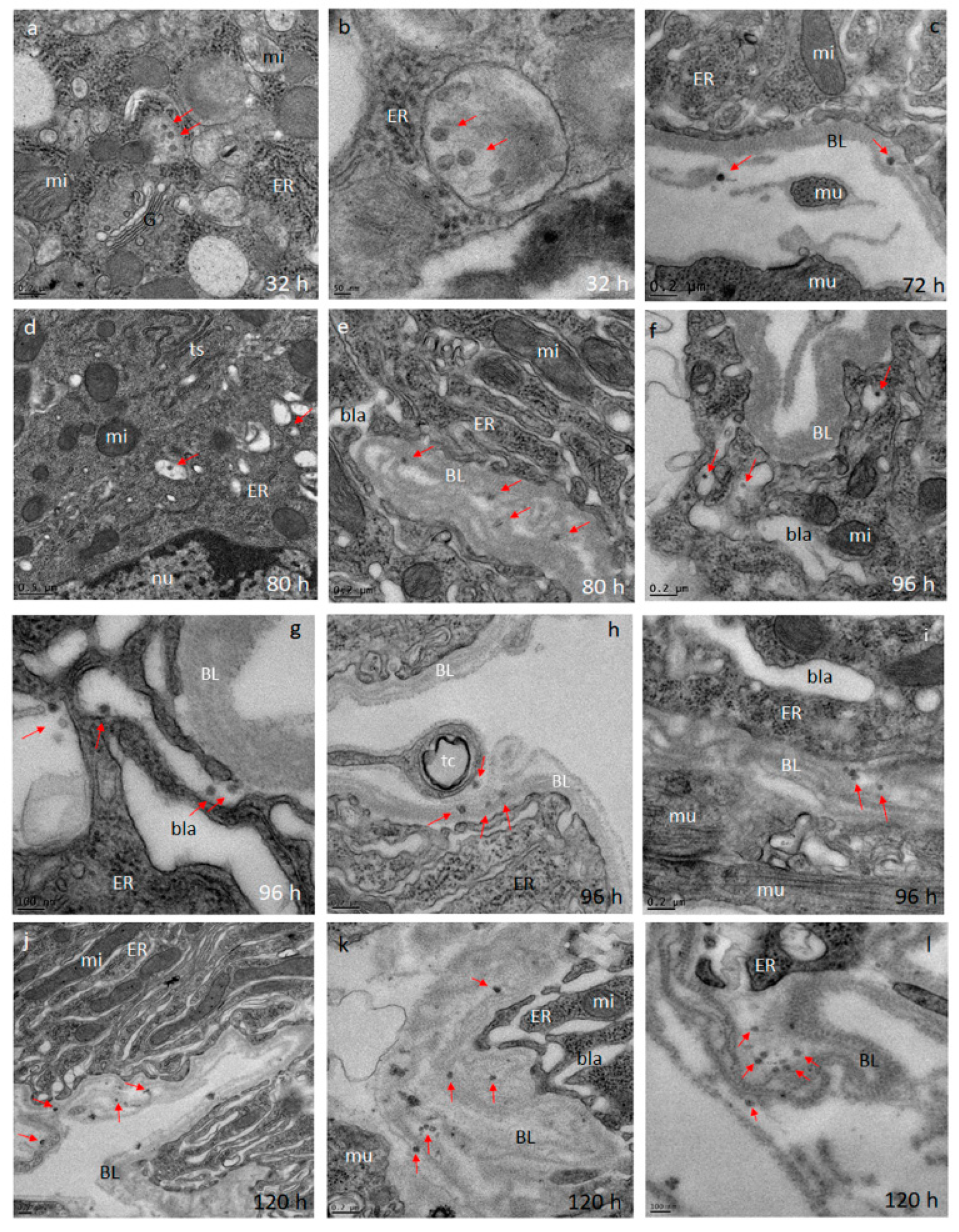
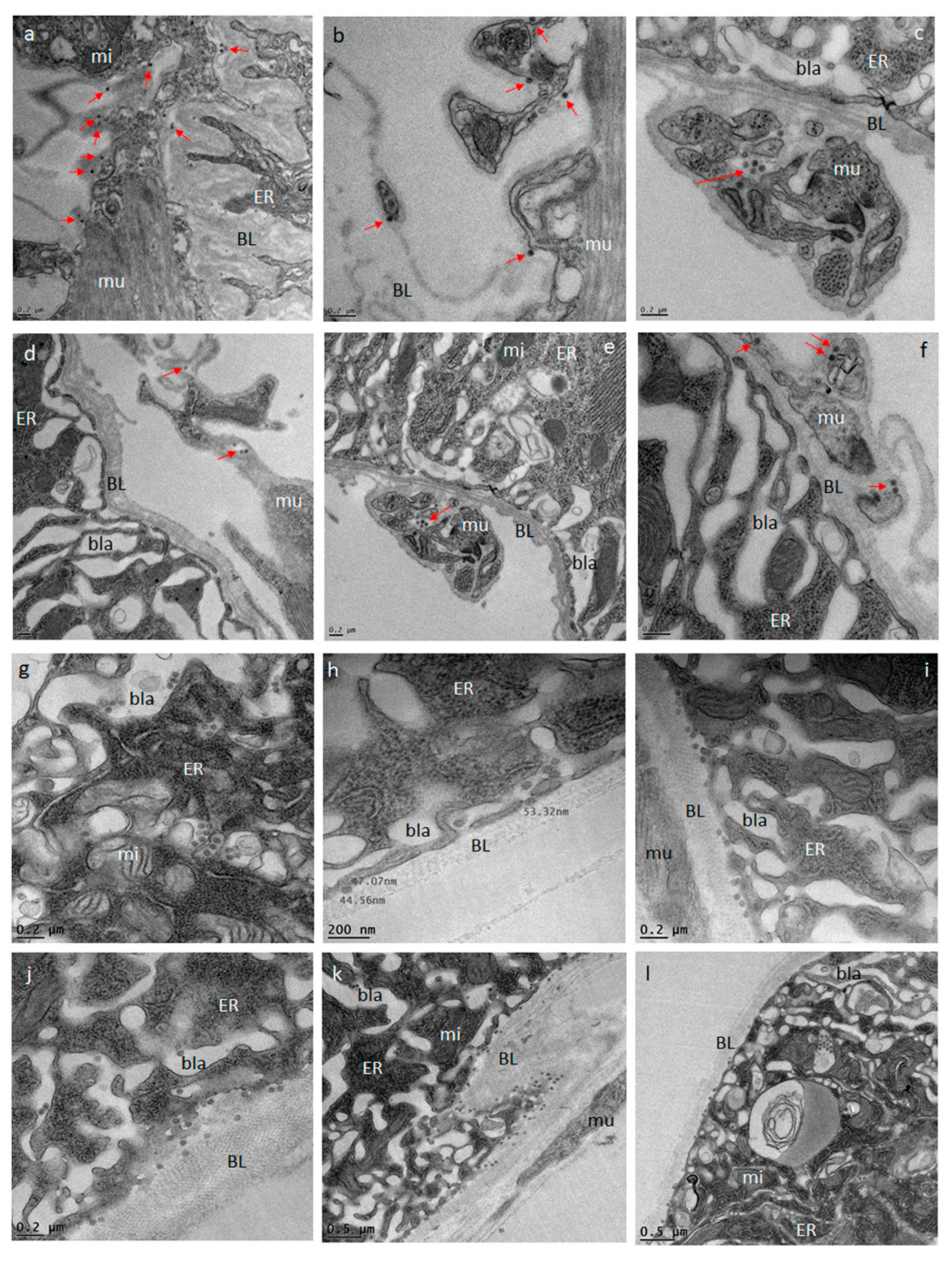
Midguts obtained from ZIKV infected (at 32, 72, 80, 96, and 120 h pibm) and DENV4 infected (at 144 h pibm) females were fixed in 100 mM sodium cacodylate buffer, pH 7.35 (Sigma Aldrich, St. Louis, MO, USA) supplemented with 2% paraformaldehyde and 2% glutaraldehyde. Sample embedding, ultrathin-sectioning, and TEM work were performed at the Electron Microscopy Core of the University of Missouri. Specimens were embedded in HistoGel (Thermo Scientific, Kalamazoo, MI, USA) before being rinsed with 100 mM sodium cacodylate buffer containing 130 mM sucrose. Secondary fixation was conducted in a Pelco Biowave (Ted Pella, Redding, CA, USA) using 100 mM sodium cacodylate buffer supplemented with 1% osmium tetroxide. Following a 1 h incubation at 4 °C, samples were en bloc stained over-night at 4 °C using 1% aqueous uranyl acetate. A graded dehydration series (100 Watts for 40 s per exchange) was performed in a Pelco Biowave, in which ethanol was initially chosen, followed by transition to acetone before dehydrated specimens were finally infiltrated with EPON resin (at 250 Watts for 3 min) and polymerized at 60 °C overnight. Embedded samples were ultrathin-sectioned (85 nm) using an ultra-microtome (Ultracut UCT, EM UC7, Leica Microsystems, Wetzlar, Germany) equipped with a diamond knife (Diatome, Hatfield, PA, USA). TEM images were acquired at 80 kV with a JEOL JEM 1400 transmission electron microscope (JEOL, Peabody, MA, USA), which was connected to a Gatan Ultrascan 1000 CCD camera (Gatan, Pleasanton, CA, USA).
2.6. Treatment of Dissected Midguts with Gold-Nanoparticles
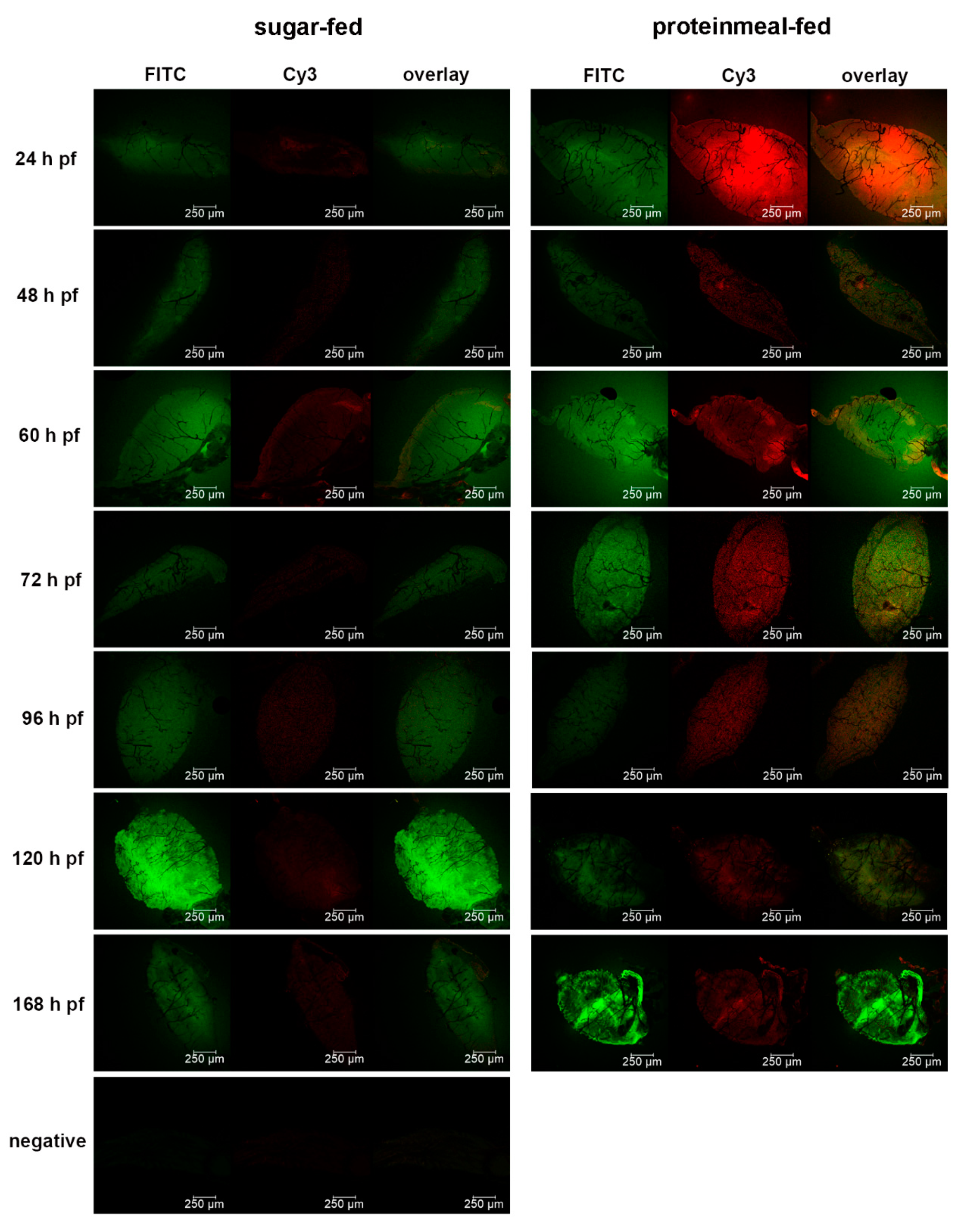
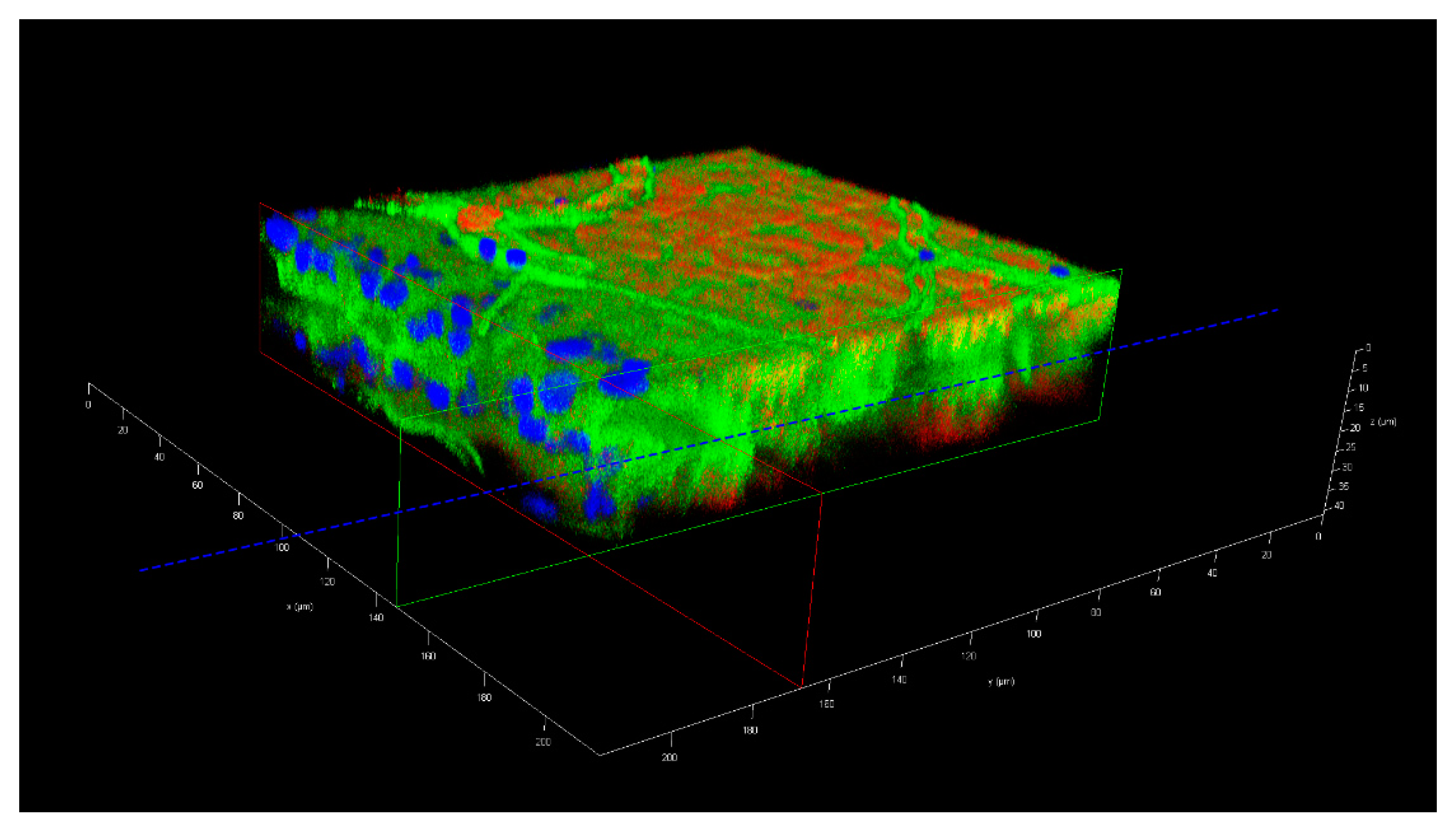
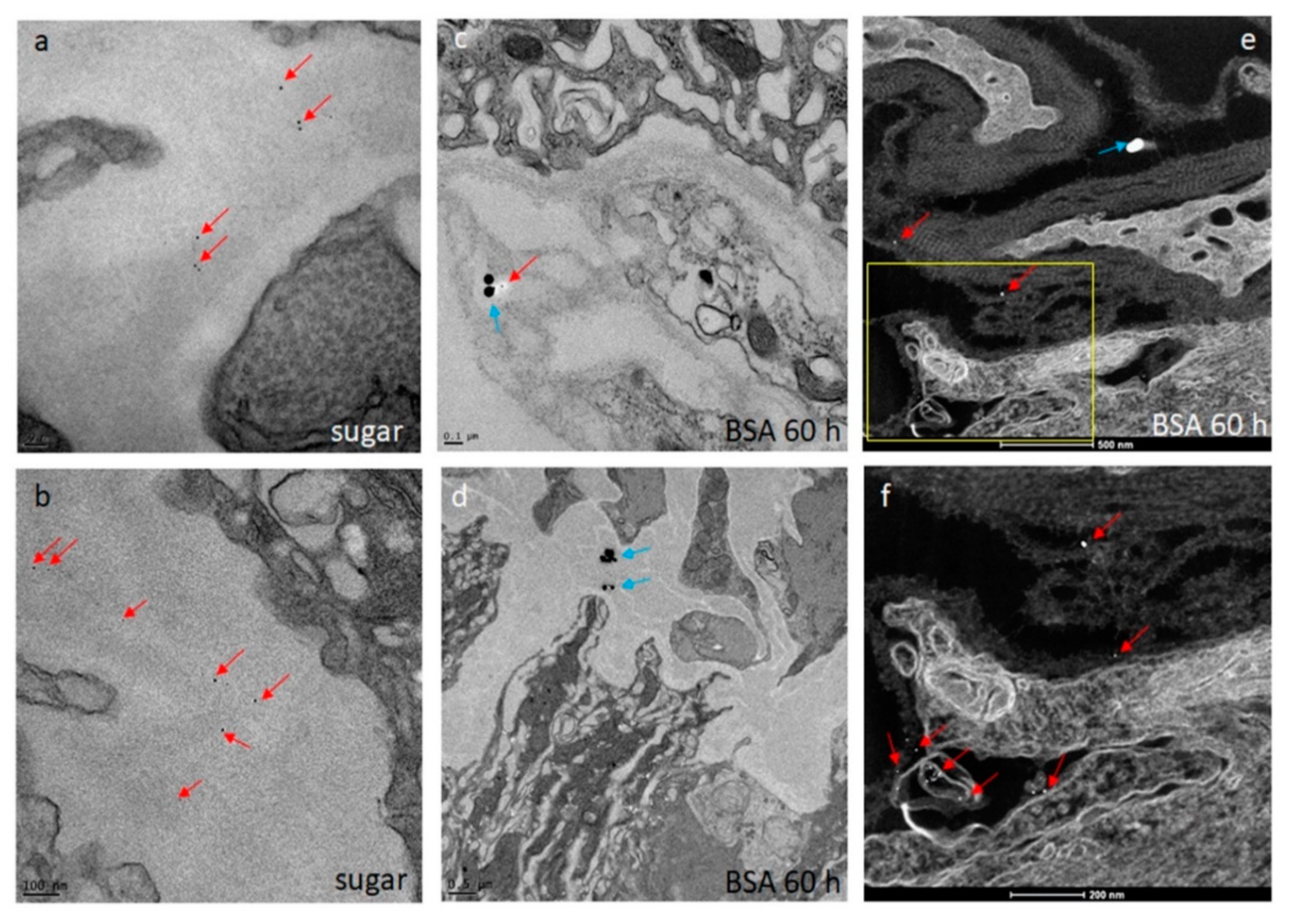
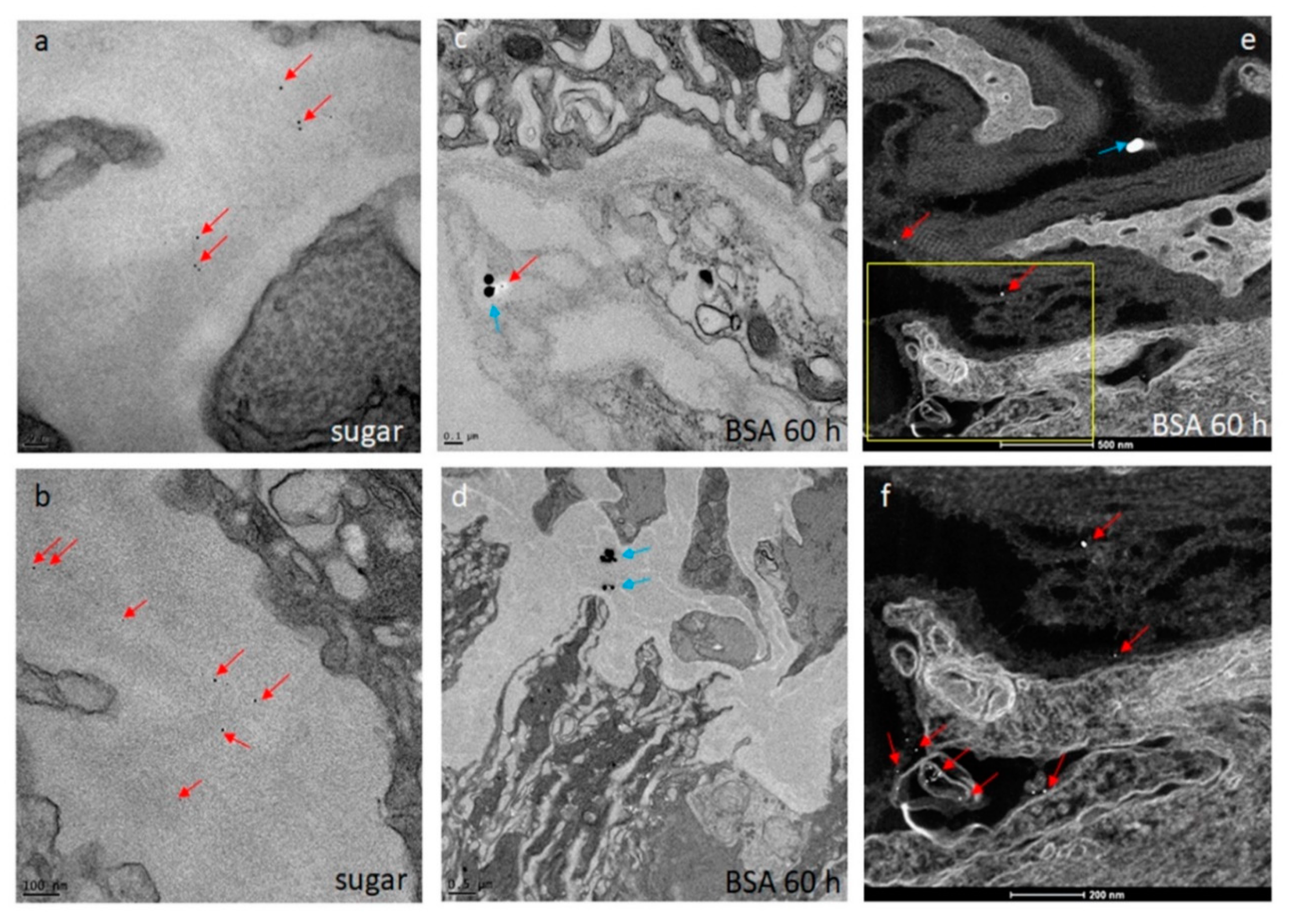
Seven days-old HWE mosquitoes were deprived of food for one day and water for 4 h before receiving a proteinmeal consisting of 20% BSA using artificial glass feeders as described above. Fully engorged mosquitoes were selected and their midguts dissected at 24, 48, 60, 72, 96, 120, and 168 h post-feeding (pf). Dissected midguts were soaked for 2 h in a suspension consisting of 5 nm FITC labeled gold-nanoparticles (1.99 × 109 nanoparticles/mL) and 50 nm Cy3 labeled gold-nanoparticles (1.99 × 109 nanoparticles/mL) (Nanopartz, Loveland, CO, USA). Midguts were then washed three times with 1x PBS before being stained with 1 µg/mL DAPI for 10 min. Following several washes with PBS, midguts were mounted on six-well printed slides and viewed under an inverted spectral confocal microscope (TCP SP8 MP, Leica Microsystems). A 3D-image was obtained from a midgut sample of a proteinmeal-fed female at 72 h pf. 3D-image reconstruction along the x, y, and z axes was performed using the LAS X 3D imaging software (Leica, Wetzlar, Germany).
2.7. Scanning Electron Microscopy (SEM) and Scanning Transmission Electron Microscopy (STEM) for the Detection of Gold-Nanoparticles in Midgut Samples
The initial SEM sample preparation steps followed the protocol for TEM sample preparation as described above. Following the graded dehydration series, samples were dried using the Tousimis Autosamdri 815 critical point dryer (Tousimis, Rockville, MD, USA), and then sputter-coated with 10 nm of platinum using the EMS 150T-ES Sputter Coater. Images were acquired with a FEI Quanta 600F environmental scanning electron microscope (FEI, Hillsboro, OR, USA). The STEM sample preparation procedure was similar to the TEM protocol as described above. Following the soaking of the midguts in the gold-nanoparticle suspension, samples were collected and processed for TEM as described. Scanning transmission electron microscopy of the TEM preparations (STEM) was performed in high angle annular dark field (HAADF) image mode on a ThermoFisher Tecnai F30 Twin 300 kV TEM operated at 200 kV. The intensity of STEM-HAADF imaging was proportional to the atomic number Z1.7, enabling gold-nanoparticles to be identified as bright particles in the sample.
4. Discussion
We investigated the midgut infection and dissemination pattern of ZIKV I-44 from the midgut of
Ae. aegypti (strain: HWE) by in vivo growth curve analysis, in situ detection of viral antigen over time, and ultrastructurally using TEM with the aim to explain the dissemination mechanism of the virus. In the in vivo growth curve analysis, dissemination of orally acquired ZIKV from the mosquito midgut was not observed before 72 h pibm since there was no infection of secondary tissues prior to that time point. At 72 h pibm, however, the midgut tissue was strongly infected with the virus (median virus titer: 5000 PFU/mL) and the bloodmeal completely digested. Furthermore, median titers in carcasses did not exceed ~170 PFU/mL before 168 h pibm. In our ultrastructural studies, ZIK virions were observed to associate with the midgut BL at 72 h pibm and did not appear in larger quantities between strands of the BL prior to 120–168 h pibm. In situ immunofluorescence assays confirmed our virus growth curve analysis as ZIKV antigen became apparent in the midgut at 48 h pibm, which coincided with the time point of observed viral de novo synthesis in this organ. Maximal presence of viral antigen was observed at 120–168 h pibm, when most parts of the organ looked infected. This matched the time period when the maximal virus titers were observed in the midgut. Furthermore, ZIKV did not infect the midgut associated tracheae or muscles as shown by IFA and TEM. All these observations lead to the conclusion that ZIKV dissemination from the midgut, similar to that of CHIKV, occurs by traversing the midgut BL [
21,
25,
38].
Other than that, the dissemination pattern of ZIKV differed substantially from that of CHIKV in the same strain of
Ae. aegypti. In a previous study, CHIK virions were already detected outside the mosquito midgut at 24 h pibm, before digestion of the bloodmeal [
38]. In the basal labyrinth of the midgut epithelial cells, a strong accumulation of mature virions in the process of traversing the BL, was observed between 24 and 32 h pibm [
21]. From 48 h pibm onwards, however, virion accumulation at the BL became less pronounced. We concluded that there was a “window of opportunity” during which large quantities of CHIK virions exited the midgut by traversing the BL during bloodmeal digestion when the midgut tissue was still overly distended. In contrast, as this study suggests, flaviviruses dissemination from the midgut primarily occurs after bloodmeal digestion. As described above, different replication and virion assembly strategies of alpha- and flaviviruses [
15,
16,
32,
33,
34,
35,
36] may be the underlying cause for their different infection and dissemination dynamics in
Ae. aegypti. Furthermore, mature virions of alphaviruses are substantially larger (60–70 nm in diameter) than those of flaviviruses (40–50 nm in diameter) [
17,
18,
30,
31]. Consequently, CHIKV may necessitate a more strongly distended midgut BL than ZIKV or DENV4 before being able to disseminate from the mosquito midgut. Bloodmeal ingestion leads to maximal BL distention during which the BL mesh size is significantly increased [
29]. Previously, we hypothesized that this bloodmeal induced “over”distention would increase the pore size exclusion limit of the BL sufficiently to enable CHIK virions to pass through [
21]. During subsequent bloodmeal digestion, the BL mesh size contracts again [
29]. According to our hypothesis, this would then steadily reduce the chance for CHIK virions to be able to traverse the BL. Thus, there would be a relatively narrow time span during which the mesh width of the BL would be just large enough for CHIK virions to pass through, which could explain the previously described “window of opportunity” dissemination pattern for CHIKV during bloodmeal digestion [
21]. However, after bloodmeal digestion, the mesh width of the midgut BL does not completely contract all the way to its original (pre-initial bloodmeal) width, which, in an earlier study, measured to be around 30 nm in
Ae. aegypti [
29]. This then could explain why flaviviruses, which are up to ~20 nm smaller in diameter than alphaviruses, would still be able to disseminate from the midgut after digestion of the bloodmeal. To further substantiate the idea, we decided to analyze the permissiveness of the midgut BL before, during, and following bloodmeal digestion using 5 nm and 50 nm size gold-nanoparticles. In agreement with Reinhardt and Hecker [
29], our experiments here suggest that following ingestion of an initial proteinmeal, the BL mesh width is enlarged due to midgut tissue distention and remains so for ~70 h after complete digestion of the meal. Whereas 5 nm gold-nanoparticles were consistently absorbed by the midgut BL of sugar-fed and proteinmeal-fed mosquitoes, 50 nm gold-nanoparticles predominantly associated with the BL of midguts obtained from proteinmeal-fed mosquitoes.
In another, earlier ultrastructural analysis, the permeability of the midgut BL of
Culex tarsalis for various particle sizes was analyzed [
43]. It was demonstrated that soaking dissected midguts of sugar-fed
Cx. tarsalis in suspensions containing lanthanum, horseradish peroxidase, cytochrome C, or colloidal thorium with sizes ranging from <2–8 nm resulted in particle absorption by the midgut BL, with particles occasionally found within the midgut epithelium. Particles of >8 nm in size accumulated at the basal site of the BL but did not traverse it, which is in accordance to our observations using gold-nanoparticles. Unfortunately, the authors did not provide any data showing whether or not particles of >8 nm in size were able to traverse the BL of midguts obtained from blood-fed mosquitoes. However, a statement was provided that there was no difference between sugar-fed and blood-fed mosquitoes, which contradicts previous work and our observations here [
29]. Two observations made during our study remain enigmatic: 1) The fact that gold-nanoparticles of either size generally did not completely penetrate or traverse the midgut BL although they were found located between strands of the BL and 2) that in some cases 50 nm nanoparticles were able to associate with the BL of sugar-fed mosquitoes. We speculate that their physical or electrostatic properties including their lack of elasticity/flexibility may have prevented those gold-nanoparticles from completely traversing the BL and entering the midgut epithelium. It may also be that the mesh width of the entire midgut BL is not completely uniform—neither in midguts of sugar-fed nor in those of (protein) meal-fed mosquitoes. Occasionally, even a midgut from a sugar-fed mosquito may contain zones were the BL mesh size is somewhat enlarged. In a few instances, SEM images showed slight tears in the BL surrounding the midgut of sugar-fed mosquitoes (see
Figure S1; [
21]. Our data also demonstrate that the number of midguts obtained from proteinmeal-fed mosquitoes showing association with 50 nm gold-nanoparticles strongly fluctuated between the various time points. In conjunction with our explanation above, we speculate that meal ingestion into the midgut may not uniformly expand the mesh width of the entire BL surrounding each midgut. Previously, we suggested that following bloodmeal digestion, the midgut BL structure would reconstitute to its pre-bloodmeal condition with the help of extracellular proteases such as matrix-metalloproteinases (MMPs) [
25,
44]. Our data here suggest that, similar to observations made by Reinhardt and Hecker [
29], the mesh width of the midgut BL remains distended for a prolonged period of time after bloodmeal digestion. We hypothesize that this is a critical phenomenon enabling ZIK virions to disseminate from the midgut after complete digestion of the bloodmeal. Our experiments did not confirm whether the midgut BL structure remains permanently altered after a single bloodmeal ingestion since our observation period did not exceed 168 h pf.
Previous reports have shown that DENV2 titers were significantly increased in mosquitoes, which had ingested a subsequent, non-infectious bloodmeal several days after ingestion of an initial, DENV2-containing bloodmeal [
45,
46]. Our ultrastructural data here show that DENV4 accumulation at the epithelial side of the midgut BL was strongly increased at 144 h pibm in those females that had acquired a second, non-infectious bloodmeal 120 h after the initial, DENV4 containing bloodmeal. Thus, the enhancement effect of subsequent bloodmeal ingestion was apparent before the virus exited the midgut. This leads us to conclude that ingestion of the second, non-infectious bloodmeal might provide a nutritional boost for the already virus-infected midgut epithelium thereby increasing virus production.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}