Wildlife Management Practices Associated with Pathogen Exposure in Non-Native Wild Pigs in Florida, U.S.

 ,
, 
Abstract
1. Introduction
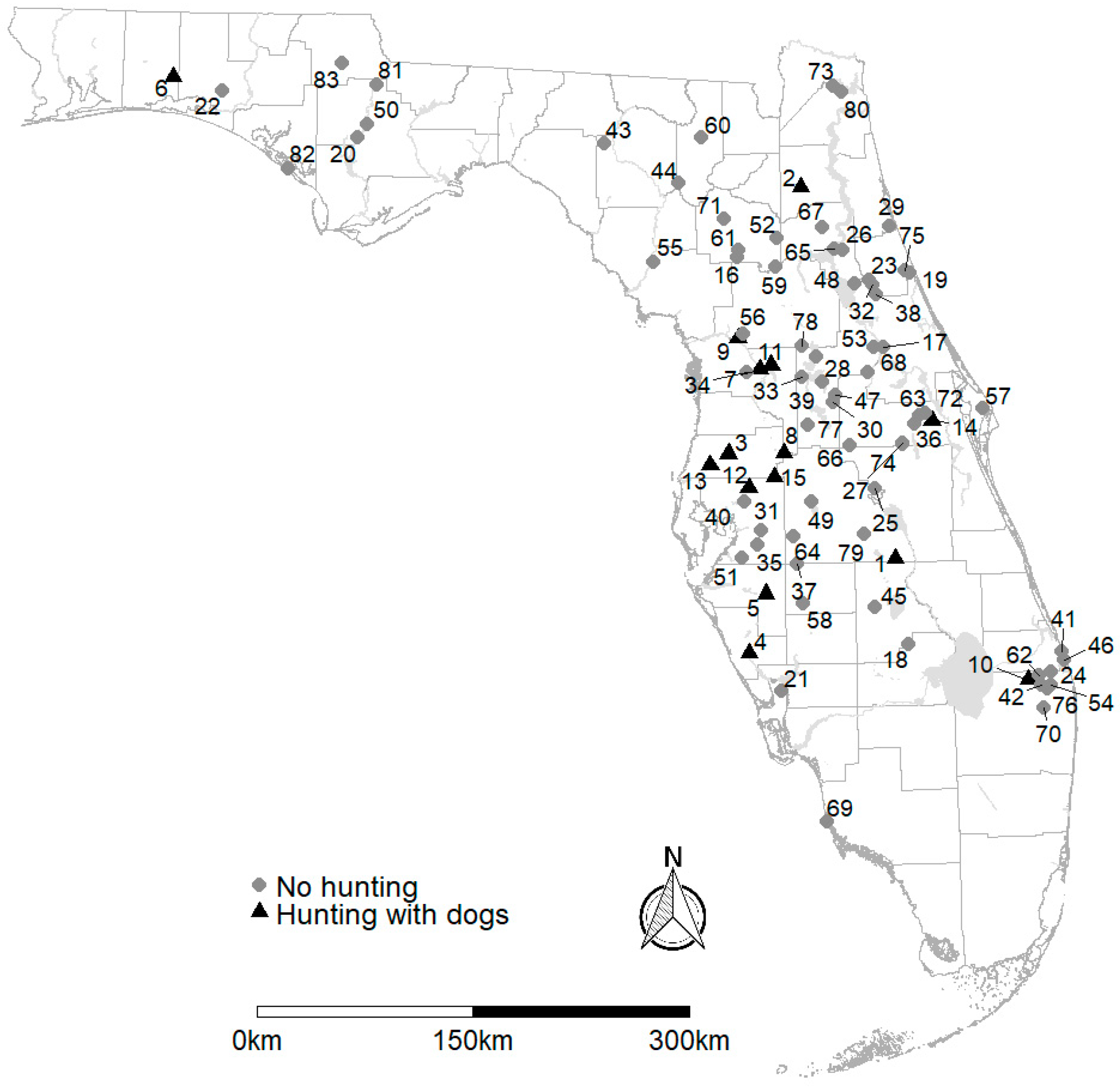
2. Materials and Methods
3. Results
3.1. Probability of Disease Detection
3.2. Effect of Dog Hunting on the Prevalence of Disease
3.3. Effect of Hunting on the Odds of Pathogen Exposure
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- King, L.J.; Anderson, L.R.; Blackmore, C.G.; Blackwell, M.J.; Lautner, E.A.; Marcus, L.C.; Meyer, T.E.; Monath, T.P.; Nave, J.E.; Ohle, J.; et al. Executive summary of the AVMA One Health Initiative Task Force report. J. Am. Vet. Med. Assoc. 2008, 233, 259–261. [Google Scholar] [CrossRef] [PubMed]
- Patz, J.A.; Graczyk, T.K.; Geller, N.; Vittor, A.Y. Effects of environmental change on emerging parasitic diseases. Int. J. Parasitol. 2000, 30, 1395–1405. [Google Scholar] [CrossRef]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging infectious diseases of wildlife—Threats to biodiversity and human health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Patz, J.A.; Daszak, P.; Tabor, G.M.; Aguirre, A.A.; Pearl, M.; Epstein, J.; Wolfe, N.D.; Kilpatrick, A.M.; Foufopoulos, J.; Molyneux, D.; et al. Unhealthy landscapes: Policy recommendations on land use change and infectious disease emergence. Environ. Health Perspect. 2004, 112, 1092–1098. [Google Scholar] [CrossRef] [PubMed]
- Farnsworth, M.L.; Wolfe, L.L.; Hobbs, N.T.; Burnham, K.P.; Williams, E.S.; Theobald, D.M.; Conner, M.M.; Miller, M.W. Human land use influences chronic wasting disease prevalence in mule deer. Ecol. Appl. 2005, 15, 119–126. [Google Scholar] [CrossRef]
- Chua, K.B.; Goh, K.J.; Wong, K.T.; Kamarulzaman, A.; Tan, P.S.K.; Ksiazek, T.G.; Zaki, S.R.; Paul, G.; Lam, S.K.; Tan, C.T. Fatal encephalitis due to Nipah virus among pig-farmers in Malaysia. Lancet 1999, 354, 1257–1259. [Google Scholar] [CrossRef]
- Rose, J.B.; Epstein, P.R.; Lipp, E.K.; Sherman, B.H.; Bernard, S.M.; Patz, J.A. Climate variability and change in the United States: Potential impacts on water- and foodborne diseases caused by microbiologic agents. Environ. Health Perspect. 2001, 109, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Van Houtan, K.S.; Hargrove, S.K.; Balazs, G.H. Land use, macroalgae, and a tumor-forming disease in marine turtles. PLoS ONE 2010, 5, e12900. [Google Scholar] [CrossRef] [PubMed]
- Ballantyne, E.E.; O’Donoghue, J.G. Rabies control in Alberta. J. Am. Vet. Med. Assoc. 1954, 125, 316–326. [Google Scholar] [PubMed]
- Macdonald, D.W. Rabies and Wildlife: A Biologist’s Perspective; Earth Resources Research Limited: Oxford, UK, 1980.
- Woodroffe, R. Managing disease threats to wild mammals. Anim. Conserv. 1999, 2, 185–193. [Google Scholar] [CrossRef]
- Woodroffe, R.; Cleaveland, S.; Courtenay, O.; Laurenson, M.K.; Artois, M. Infectious disease in the management and conservation of wild canids. In The Biology and Conservation of Wild Canids; Macdonald, D.W., Sillero-Zubiri, C., Eds.; Oxford University Press: Oxford, UK, 2004; pp. 123–142. [Google Scholar]
- Carter, S.P.; Roy, S.S.; Cowan, D.P.; Massei, G.; Ji, W.; Rossi, S.; Woodroffe, R.; Wilson, G.J. Options for the control of disease 2: Targeting hosts. In Management of Disease in Wild Mammals; Delahay, R.J., Smith, G.C., Hutchings, M.R., Eds.; Springer: Tokyo, Japan, 2009; pp. 129–146. [Google Scholar]
- Potapov, A.; Merrill, E.; Lewis, M.A. Wildlife disease elimination and density dependence. Proc. R. Soc. Lond. 2012, 279, 3139–3145. [Google Scholar] [CrossRef]
- Bulai, I.M.; Cavoretto, R.; Chialva, B.; Duma, D.; Venturino, E. Comparing disease-control policies for interacting wild populations. Nonlinear Dyn. 2015, 79, 1881–1900. [Google Scholar] [CrossRef]
- Guberti, V.; Rutili, D.; Ferrari, G.; Patta, C.; Oggiano, A. Estimate the threshold abundance for the persistence of classical swine fever in the wild boar population of the eastern Sardinia. In Proceedings of the Meeting Measures to Control Classical Swine Fever in European Wild Boar, Perugia, Italy, 6–7 April 1998; pp. 54–61. [Google Scholar]
- Swinton, J.M.E.; Woolhouse, M.E.G.; Begon, M.E.; Dobson, A.P.; Ferroglio, E.; Grenfell, B.T.; Guberti, V.; Hails, R.S.; Heesterbeek, J.A.P.; Lavazza, A.; et al. Microparasite transmission and persistence. In The Ecology of Wildlife Diseases, 1st ed.; Hudson, P.J., Rizzoli, A., Grenfell, B.T., Heesterbeek, H., Dobson, A.P., Eds.; Oxford University Press: New York, NY, USA, 2002; pp. 83–101. [Google Scholar]
- Rossi, S.; Fromot, E.; Pontier, D.; Crucière, C.; Hars, J.; Barrat, J.; Pacholek, X.; Artois, M. Incidence and persistence of classical swine fever in free-ranging wild boar (Sus scrofa). Epidemiol. Infect. 2005, 133, 559–568. [Google Scholar] [CrossRef]
- Pauli, J.N.; Buskirk, S.W. Risk-disturbance overrides density dependence in a hunted colonial rodent, the black- tailed prairie dog Cynomys ludovicianus. J. Appl. Ecol. 2007, 44, 1219–1230. [Google Scholar] [CrossRef]
- Bateson, P.; Bradshaw, E.L. Physiological effects of hunting red deer (Cervus elaphus). Proc. R. Soc. Lond. 1997, 264, 1707–1714. [Google Scholar] [CrossRef]
- Mason, G. The physiology of the hunted deer. Nature 1998, 391, 22. [Google Scholar] [CrossRef] [PubMed]
- Padgett, D.A.; Glaser, R. How stress influences the immune response. Trends Immunol. 2003, 24, 444–448. [Google Scholar] [CrossRef]
- Choisy, M.; Rohani, P. Harvesting can increase severity of wildlife disease epidemics. Proc. R. Soc. Lond. 2006, 273, 2025–2034. [Google Scholar] [CrossRef] [PubMed]
- Crowl, T.A.; Crist, T.O.; Parmenter, R.R.; Belovsky, G.; Lugo, A.E. The spread of invasive species and infectious disease as drivers of ecosystem change. Front. Ecol. Environ. 2008, 6, 238–246. [Google Scholar] [CrossRef]
- Leiser, O.P.; Corn, J.L.; Schmit, B.S.; Keim, P.S.; Foster, J.T. Feral swine brucellosis in the United States and prospective genomic techniques for disease epidemiology. Vet. Microbiol. 2013, 166, 1–10. [Google Scholar] [CrossRef]
- Pedersen, K.; Bevins, S.N.; Baroch, J.A.; Cumbee, J.C., Jr.; Chandler, S.C.; Woodruff, B.S.; Bigelow, T.T.; DeLiberto, T.J. Pseudorabies in feral swine in the United States, 2009–2012. J. Wildl. Dis. 2013, 49, 709–713. [Google Scholar] [CrossRef] [PubMed]
- Pellet, P.E.; Roizman, B. Herpesviridae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2013; pp. 1802–1822. [Google Scholar]
- Kluge, J.P.; Beran, G.W.; Hill, H.T.; Platt, K.B. Pseudorabies (Aujeszky’s disease). In Diseases of Swine, 8th ed.; Straw, B.E., Dállaire, S., Mengeling, W.L., Taylor, D.J., Eds.; Blackwell Science: Oxford, UK, 1999; pp. 233–246. [Google Scholar]
- Young, E.J. An overview of human brucellosis. Clin. Infect. Dis. 1995, 21, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Hahn, E.; Page, G.; Hahn, P.; Gillis, K.; Romero, C. Mechanisms of transmission of Aujeszky’s disease virus originating from feral swine in the USA. Vet. Microbiol. 1997, 55, 123–130. [Google Scholar] [CrossRef]
- Müller, T.F.; Teuffert, J.; Zellmer, R.; Conraths, F.J. Experimental infection of European wild boars and domestic pigs with pseudorabies viruses with differing virulence. Am. J. Vet. Res. 2001, 62, 252–258. [Google Scholar] [CrossRef]
- USDA (United States Department of Agriculture). Pseudorabies (Aujeszky’s Disease) and its Eradication: A Review of the U.S. Experience; Technical Bulletin No. 1923; USDA: Washington, DC, USA, 2008.
- Florida Department of Agriculture and Consumer Services. Florida Feral Swine Trappers: Feral Swine in Florida; FDACS-P-1510; Division of Animal: Husbandry, FL, USA, 2016.
- Mayer, J.; Brisbin, I.L. Wild Pigs: Biology, Damage, Control Techniques and Management; No. SRNL-RP-2009-00869; Savannah River Site: Aiken, SC, USA, 2009.
- Hernández, F.A.; Parker, B.M.; Pylant, C.L.; Smyser, T.J.; Piaggio, A.J.; Lance, S.L.; Milleson, M.P.; Austin, J.D.; Wisely, S.M. Invasion ecology of wild pigs (Sus scrofa) in Florida, USA: The role of humans in the expansion and colonization of an invasive wild ungulate. Biol. Invasions 2018, 20, 1865–1880. [Google Scholar] [CrossRef]
- Giuliano, W.M. Wild Hogs in Florida: Ecology and Management; Document WEC277; University of Florida Institute of Food and Agricultural Sciences: Gainesville, FL, USA, 2016. [Google Scholar]
- Matschke, G.H. Aging European wild hogs by dentition. J. Wildl. Manag. 1967, 31, 109–113. [Google Scholar] [CrossRef]
- University of Florida GeoPlan Center. 2015. Available online: https://www.fgdl.org (accessed on 30 March 2018).
- Florida Department of Environmental Protection. 1989. Available online: https://www.fgdl.org (accessed on 30 March 2018).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: http://www.R-project.org/ (accessed on 14 November 2018).
- Pannwitz, G.; Freuling, C.; Denzin, N.; Schaarschmidt, U.; Nieper, H.; Hlinak, A.; Burkhardt, S.; Klopries, M.; Dedek, J.; Hoffman, L.; et al. A long-term serological survey on Aujeszky’s disease virus infections in wild boar in East Germany. Epidemiol. Infect. 2012, 140, 348–358. [Google Scholar] [CrossRef]
- Cross, P.C.; Edwards, W.H.; Scurlock, B.M.; Maichak, E.J.; Rogerson, J.D. Effects of management and climate on elk brucellosis in the Greater Yellowstone Ecosystem. Ecol. Appl. 2007, 17, 957–964. [Google Scholar] [CrossRef]
- Gilbert, A.T.; Fooks, A.R.; Hayman, D.T.; Horton, D.L.; Müller, T.; Plowright, R.; Peel, A.J.; Bowen, R.; Wood, J.L.; Mills, J.; et al. Deciphering serology to understand the ecology of infectious diseases in wildlife. EcoHealth 2013, 10, 298–313. [Google Scholar] [CrossRef]
- McClintock, B.T.; Nichols, J.D.; Bailey, L.L.; MacKenzie, D.I.; Kendal, W.L.; Franklin, A.B. Seeking a second opinion: Uncertainty in disease ecology. Ecol. Lett. 2010, 13, 659–674. [Google Scholar] [CrossRef]
- Corn, J.L.; Stallknecht, D.E.; Mechlin, N.M.; Luttrell, M.P.; Fischer, J.R. Persistence of pseudorabies virus in feral swine populations. J. Wildl. Dis. 2004, 40, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Hines, J.E. PRESENCE2—Software to Estimate Patch Occupancy and Related Parameters. 2006. Available online: http://www.mbr-pwrc.gov/software/presence.html (accessed on 1 December 2016).
- Chinsakchai, S.; Molitor, T.W. Replication and immunosuppressive effects of Pseudorabies virus on swine peripheral blood mononuclear cells. Vet. Immunol. Immunopath. 1992, 30, 247–260. [Google Scholar] [CrossRef]
- Dhabar, F.S.; McEwen, B.S. Enhancing versus suppressive effects of stress hormones on skin immune function. Proc. Natl. Acad. Sci. USA 1999, 96, 1059–1064. [Google Scholar] [CrossRef]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar] [CrossRef] [PubMed]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Van der Leek, M.L.; Becker, H.N.; Humphrey, P.; Adams, C.L.; Belden, R.C.; Frankenberger, W.B.; Nicoletti, P.L. Prevalence of brucellosis sp. antibodies in feral swine in Florida. J. Wildl. Dis. 1993, 29, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Van der Leek, M.L.; Becker, H.N.; Pirtle, E.C.; Humphrey, P.; Adams, C.L.; All, B.P.; Erickson, G.A.; Belden, R.C.; Frankenberger, W.B.; Gibbs, E.P.J. Prevalence of pseudorabies (Aujeszky’s disease) virus antibodies in feral swine in Florida. J. Wildl. Dis. 1993, 29, 403–409. [Google Scholar] [CrossRef]
- Pedersen, K.; Bevins, S.N.; Schmit, B.S.; Lutman, M.W.; Milleson, M.P. Apparent prevalence of swine brucellosis in feral swine in the United States. Hum.-Wildl. Interact. 2012, 6, 38–47. [Google Scholar]
- Boelaert, F.; Deluyker, H.; Maes, D.; Godfroid, J.; Raskin, A.; Varewijck, H.; Pensaert, M.B.; Nauwynck, H.S.; Castryck, F.; Miry, C.; et al. Prevalence of herds with young sows seropositive to pseudorabies (Aujeszky’s disease) in northern Belgium. Prev. Vet. Med. 1999, 41, 239–255. [Google Scholar] [CrossRef]
- Risco, D.; Garcia, A.; Serrano, E.; Fernandez-Llario, P.; Benitez, J.M.; Martinez, R.; Garcia, W.L.; Hermoso de Mendoza, J. High-density dependence but low impact on selected reproduction parameters of Brucella suis biovar 2 in wild boar hunting estates from south-western Spain. Transbound. Emerg. Dis. 2014, 61, 555–562. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Bartón, K. R package “MuMIn”: Multi-Model Inference (Version 1.40.0). 2017. Available online: http://CRAN.R-project.org/package=MuMIn (accessed on 14 November 2018).
- Schoenbaum, M.A.; Beran, G.W.; Murphy, D.P. A study comparing the immunologic responses of swine to pseudorabies viral antigens based on the ELISA, serum neutralization, and latex agglutination tests. J. Vet. Diagn. Investig. 1990, 2, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Oren, S.L.; Swenson, S.L.; Kinker, D.L. Evaluation of serological pseudorabies tests for the detection of antibodies during early infection. J. Vet. Diagn. Invest. 1993, 5, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, S.J.; Karges, S.L.; Saliki, J.T. Evaluation of a semiautomated latex agglutination test for the detection of pseudorabies antibody in swine sera. J. Vet. Diagn. Investig. 1996, 8, 168–171. [Google Scholar] [CrossRef]
- Mikolon, A.B.; Gardner, L.A.; Hietala, S.K.; de Anda, J.H.; Pestaña, E.C.; Hennager, S.G.; Edmondson, A.J. Evaluation of North American antibody detection tests for diagnosis of brucellosis in goats. J. Clin. Microbiol. 1998, 36, 1716–1722. [Google Scholar] [PubMed]
- Nielsen, K. Diagnosis of brucellosis by serology. Vet. Microbiol. 2002, 90, 447–459. [Google Scholar] [CrossRef]
- Gaston, W.; Armstrong, J.B.; Arjo, W.; Stribling, H.L. Home range and habitat use of feral hogs (Sus scrofa) on Lowndes County WMA, Alabama. In Proceedings of the National Conference on Feral Hogs, St. Louis, MO, USA, 13–15 April 2008. [Google Scholar]
- Pepin, K.M.; Davis, A.J.; Beasley, J.; Boughton, R.; Campbell, T.; Cooper, S.M.; Gaston, W.; Hartley, S.; Kilgo, J.C.; Wisely, S.M.; et al. Contact heterogeneities in feral swine: Implications for disease management and future research. Ecosphere 2016, 7, e01230. [Google Scholar] [CrossRef]
- Bieber, C.; Ruf, T. Population dynamics in wild boar Sus scrofa: Ecology, elasticity of growth rate and implications for the management of pulsed resource consumers. J. Appl. Ecol. 2005, 42, 1203–1213. [Google Scholar] [CrossRef]
- Thiry, E.; Saliki, J.; Bublot, M.; Pastoret, P.P. Reactivation of infectious bovine rhinotracheitis virus by transport. Comp. Immunol. Microbiol. Infect. Dis. 1987, 10, 59–63. [Google Scholar] [CrossRef]
- Padgett, D.A.; Sheridan, J.F.; Dorne, J.; Berntson, G.G.; Candelora, J.; Glaser, R. Social stress and the reactivation of latent herpes simplex virus type 1. Proc. Nat. Acad. Sci. USA 1998, 95, 7231–7235. [Google Scholar] [CrossRef]
- Weigler, B.J.; Hird, D.W.; Hilliard, J.K.; Lerche, N.W.; Roberts, J.A.; Scott, L.M. Epidemiology of cercopithecine herpesvirus 1 (B virus) infection and shedding in a large breeding cohort of rhesus macaques. J. Infect. Dis. 1993, 167, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Tozzini, F.; Poli, A.; Croce, G.D. Experimental infection of European wild swine (Sus scrofa L.) with pseudorabies virus. J. Wildl. Dis. 1982, 18, 425–428. [Google Scholar] [CrossRef] [PubMed]
- Capua, I.; Fico, R.; Banks, M.; Tamba, M.; Calzetta, G. Isolation and characterisation of an Aujeszky’s disease virus naturally infecting a wild boar (Sus scrofa). Vet. Microbiol. 1997, 55, 141–146. [Google Scholar] [CrossRef]
- Pirtle, E.C.; Sacks, J.M.; Nettles, V.F.; Rollor, E.A., III. Prevalence and transmission of pseudorabies virus in an isolated population of feral swine. J. Wildl. Dis. 1989, 25, 605–607. [Google Scholar] [CrossRef] [PubMed]
- Romero, C.; Meade, P.; Santagata, J.; Gillis, K. Genital infection and transmission of pseudorabies virus in feral swine in Florida, USA. Vet. Microbiol. 1997, 55, 131–139. [Google Scholar] [CrossRef]
- Romero, C.H.; Meade, P.N.; Schultz, J.E.; Chung, H.Y.; Gibbs, E.P.; Hahn, E.C.; Lollis, G. Venereal transmission of pseudorabies virus indigenous to feral swine. J. Wildl. Dis. 2001, 37, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.; Hahn, E.C.; Tottewitz, F.; Kramer, M.; Klupp, B.G.; Mettenleiter, T.C.; Freuling, C. Pseudorabies virus in wild swine: A global perspective. Arch. Virol. 2011, 156, 1691–1705. [Google Scholar] [CrossRef]
- McIlroy, J.C.; Saillard, R.J. The effect of hunting with dogs on the numbers and movements of feral pigs, Sus-scrofa, and the subsequent success of poisoning exercises in Namadgi-National-Park, A.C.T. Aust. Wildl. Res. 1989, 16, 353–363. [Google Scholar] [CrossRef]
- Caley, P. Population dynamics of feral pigs (Sus scrofa) in a tropical riverine habitat complex. Wildl. Res. 1993, 20, 625–636. [Google Scholar] [CrossRef]
- Kay, S.L.; Fisher, J.W.; Monaghan, A.J.; Beasley, J.C.; Boughton, R.; Campbell, T.A.; Cooper, S.M.; Ditchkoff, S.S.; Hartley, S.B.; Kilgo, J.C.; et al. Quantifying drivers of wild pig movements across multiple spatial and temporal scales. Mov. Ecol. 2017, 5, 14. [Google Scholar] [CrossRef]
- Franckowiak, G.A.; Poché, R.M. Short-term home range and habitat selection by feral hogs in northern Texas. Am. Midl. Nat. 2018, 179, 28–37. [Google Scholar] [CrossRef]
- Hernández, F.A.; Carr, A.N.; Milleson, M.P.; Merrill, H.R.; Avery, M.L.; Parker, B.M.; Pylant, C.L.; Austin, J.D.; Wisely, S.M. Dispersal and land cover as contributing factors of pseudorabies virus (PrV) exposure in invasive wild pigs (Sus scrofa). Basic Appl. Ecol. 2018. Submitted. [Google Scholar]
- Glass, C.M.; McLean, R.G.; Katz, J.B.; Maehr, D.S.; Cropp, C.B.; Kirk, L.J.; McKeirnan, A.J.; Evermann, J.F. Isolation of pseudorabies (Aujeszky’s disease) virus from a Florida panther. J. Wildl. Dis. 1994, 30, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Franco, M.P.; Mulder, M.; Gilman, R.H.; Smits, H.L. Human brucellosis. Lancet 2007, 7, 775–786. [Google Scholar] [CrossRef]
- Wildlife Services. Table G. Animals Taken by Wildlife Services—FY 2012; USDA APHIS Wildlife Damage Program Data Reports; Wildlife Services: Washington, DC, USA, 2012.
- Kokko, H.; Lindström, J. Seasonal density dependence, timing of mortality, and sustainable harvesting. Ecol. Model. 1998, 110, 293–304. [Google Scholar] [CrossRef]
- Boyce, M.S.; Sinclair, A.R.E.; White, G.C. Seasonal compensation of predation and harvesting. Oikos 1999, 87, 419–426. [Google Scholar] [CrossRef]
- Jonzén, N.; Lundberg, P. Temporally structured density-dependence and population management. Ann. Zool. Fenn. 1999, 36, 39–44. Available online: https://www.jstor.org/stable/23735640 (accessed on 10 December 2018).
- Xu, C.; Boyce, M.S.; Daley, D.J. Harvesting in seasonal environments. J. Math. Biol. 2005, 50, 663–682. [Google Scholar] [CrossRef]
- Hone, J. Predator-prey theory and feral pig control, with emphasis on evaluation of shooting from a helicopter. Aust. Wildl. Res. 1999, 17, 123–130. [Google Scholar] [CrossRef]
- Saunders, G. Observation on the effectiveness of shooting feral pigs from helicopters. Eur. J. Wildl. Res. 1993, 20, 771–776. [Google Scholar] [CrossRef]
- Campbell, T.A.; Long, D.B.; Leland, B.R. Feral swine behavior relative to aerial gunning in southern Texas. J. Wildl. Manag. 2010, 74, 337–342. [Google Scholar] [CrossRef]


{kind=link}
| Pathogen | Level | p | Reference |
|---|---|---|---|
| PrV | State | 1.00 | This study |
| Site | 0.81 | This study | |
| Assay | |||
| LAT | 0.99 | [59,60,61] | |
| ELISA | 1.00 | [59,60,61] | |
| Compounded | |||
| LAT | 0.80 | This study | |
| ELISA | 0.81 | This study | |
| Brucella spp. | State | 1.00 | This study |
| Site | 0.63 | This study | |
| Assay | |||
| Card | 0.73 | [62,63] | |
| RIV | 0.60 | [62,63] | |
| FPA | 0.99 | [63] | |
| Compounded | |||
| Card | 0.46 | This study | |
| RIV | 0.38 | This study | |
| FPA | 0.62 | This study |
| Treatment | No Disease | PrV | Brucella spp. | PrV-Brucella spp. |
|---|---|---|---|---|
| Dog-hunted | 426 ↓*** | 326 ↑** | 42 ns | 101 ↑*** |
| Not dog-hunted | 694 ↑*** | 291 ↓** | 53 ns | 58 ↓** |
| Pathogen | Model | K | ΔAIC | wi | R2m | R2c |
|---|---|---|---|---|---|---|
| PrV | Harvest + Age + Harvest x Age | 6 | 0.00 | 0.65 | 0.20 | 0.50 |
| Harvest + Age | 5 | 1.24 | 0.35 | 0.18 | 0.48 | |
| Age | 4 | 12.55 | 0.00 | 0.06 | 0.46 | |
| Brucella spp. | Harvest + Age | 5 | 0.00 | 0.77 | 0.13 | 0.45 |
| Harvest + Age + Harvest x Age | 6 | 3.61 | 0.13 | 0.14 | 0.45 | |
| Age | 4 | 4.03 | 0.10 | 0.08 | 0.44 |
| Pathogen | Variable | Coefficient | OR | SEOR | RI | p-value |
|---|---|---|---|---|---|---|
| PrV | (Intercept) | −3.62 | 0.03 | 1.93 | < 0.001 | |
| Harvest | 2.47 | 11.81 | 2.45 | 1.00 | 0.006 | |
| Age | 1.00 | |||||
| Adult | 2.30 | 10.01 | 1.89 | < 0.001 | ||
| Sub-adult | 0.74 | 2.10 | 1.87 | 0.234 | ||
| Harvest x Age | 0.65 | |||||
| Harvest x Adult | −0.83 | 0.44 | 2.22 | 0.301 | ||
| Harvest x Sub-adult | −0.55 | 0.57 | 2.06 | 0.445 | ||
| Brucella spp. | (Intercept) | −5.06 | 0.01 | 2.31 | < 0.001 | |
| Harvest | 1.13 | 3.09 | 2.26 | 0.90 | 0.167 | |
| Age | 1.00 | |||||
| Adult | 2.47 | 11.80 | 2.22 | 0.002 | ||
| Sub-adult | 1.52 | 4.56 | 2.34 | 0.075 | ||
| Harvest x Age | 0.13 | |||||
| Harvest x Adult | −0.08 | 0.92 | 1.77 | 0.884 | ||
| Harvest x Sub-adult | −0.04 | 0.96 | 1.78 | 0.942 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carr, A.N.; Milleson, M.P.; Hernández, F.A.; Merrill, H.R.; Avery, M.L.; Wisely, S.M. Wildlife Management Practices Associated with Pathogen Exposure in Non-Native Wild Pigs in Florida, U.S. Viruses 2019, 11, 14. https://doi.org/10.3390/v11010014
Carr AN, Milleson MP, Hernández FA, Merrill HR, Avery ML, Wisely SM. Wildlife Management Practices Associated with Pathogen Exposure in Non-Native Wild Pigs in Florida, U.S. Viruses. 2019; 11(1):14. https://doi.org/10.3390/v11010014
Chicago/Turabian StyleCarr, Amanda N., Michael P. Milleson, Felipe A. Hernández, Hunter R. Merrill, Michael L. Avery, and Samantha M. Wisely. 2019. "Wildlife Management Practices Associated with Pathogen Exposure in Non-Native Wild Pigs in Florida, U.S." Viruses 11, no. 1: 14. https://doi.org/10.3390/v11010014
APA StyleCarr, A. N., Milleson, M. P., Hernández, F. A., Merrill, H. R., Avery, M. L., & Wisely, S. M. (2019). Wildlife Management Practices Associated with Pathogen Exposure in Non-Native Wild Pigs in Florida, U.S. Viruses, 11(1), 14. https://doi.org/10.3390/v11010014