
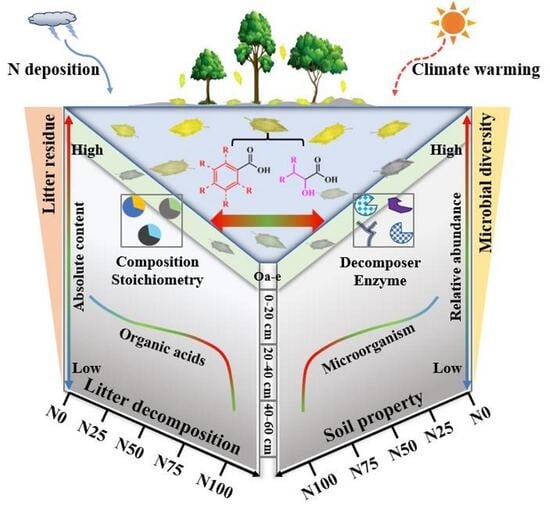
Nitrogen Addition-Induced Variations in Stoichiometric Ratio of Organic Acids from Litter Decomposition in a Temperate Forest


 ,
,
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Study Sites and Climatic Conditions
2.2. Experimental Design and Sampling
2.3. Sample Determination
2.4. Statistical Analysis
3. Results
3.1. Variation in the Characteristics of Litter Decomposition Components Across N Additions and Soil Layers
3.2. Stoichiometric Characteristics of Individual Organic Acids Across N Additions and Soil Layers
3.3. Relationships Between Stoichiometric Ratios of Organic Acids and Individual Soil Parameters
4. Discussion
4.1. Effect of Nitrogen Addition on Organic Acid Composition and Source Soil Conditions in Litter Decomposition
4.2. Application Potential of Organic Acid Stoichiometric Indicator in Litter Decomposition Adaption to N Resource Utilization and Climate Environment Change
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.X.; Guo, X.W.; Chen, L.X.; Kuzyakov, Y.; Wang, R.Z.; Zhang, H.Y.; Han, X.G.; Jiang, Y.; Sun, O.J.X. Global pattern of organic carbon pools in forest soils. Glob. Chang. Biol. 2024, 30, e17386. [Google Scholar] [CrossRef]
- Feng, X.J.; Dai, G.H.; Liu, T.; Jia, J.; Zhu, E.X.; Liu, C.Z.; Zhao, Y.P.; Wang, Y.; Kang, E.Z.; Xiao, J.; et al. Understanding the mechanisms and potential pathways of soil carbon sequestration from the biogeochemistry perspective. Sci. China Earth Sci. 2024, 67, 3386–3396. [Google Scholar] [CrossRef]
- Pessoa, D.V.; da Cunha, M.V.; de Mello, A.C.L.; dos Santos, M.V.F.; Soares, G.S.C.; Camelo, D.; Apolinário, V.X.O.; Junior, J.C.B.D.; Coelho, J.J. Litter deposition and decomposition in a tropical grass-legume silvopastoral system. J. Soil Sci. Plant Nutr. 2024, 24, 3504–3518. [Google Scholar] [CrossRef]
- Beidler, K.V.; Phillips, R.P.; Andrews, E.; Maillard, F.; Mushinski, R.M.; Kennedy, P.G. Substrate quality drives fungal necro mass decay and decomposer community structure under contrasting vegetation types. J. Ecol. 2020, 108, 1845–1859. [Google Scholar] [CrossRef]
- Abubakar, A.; Mayer, M.; Neumann, M.; Gao, Q.; Wang, D. Nitrogen addition reduces litter decomposition but does not affect litter production and chemistry in an alpine shrub land. Plant Soil 2025, 509, 795–807. [Google Scholar] [CrossRef]
- Chen, H.; Xu, Q.H.; van Groenigen, K.J.; Hungate, B.A.; Smith, P.; Li, D.J.; Moorhead, D.L.; Osborne, B.B.; Ma, Z.L.; Olesen, J.E.; et al. Linking soil extracellular enzymes with soil respiration under altered litter inputs. Agric. For. Meteorol. 2025, 367, 110503. [Google Scholar] [CrossRef]
- López, J.; Vancampenhout, K.; Muys, B.; Ponette, Q. Tree community composition modulates early-stage decomposition of standard litter through chemical and physical engineering. For. Ecosyst. 2025, 15, 100387. [Google Scholar] [CrossRef]
- Castillo-Figueroa, D. The effect of forest microenvironment on litter decomposition in the Andean tropical mountains. J. For. Res. 2025, 36, 102. [Google Scholar] [CrossRef]
- Zhao, Y.D.; Lu, N.; Shi, H.; Huang, J.B.; Fu, B.J. Patterns and driving factors of litter decomposition rates in global dryland ecosystem. Glob. Chang. Biol. 2025, 31, e70025. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, Y.N.; Wang, M.R.; Wang, K.; Meng, F.L.; Liu, L.; Zhao, Y.H.; Ma, L.; Zhu, Q.C.; Xu, W.; et al. Atmospheric nitrogen deposition: A review of quantification methods and its spatial pattern derived from the global monitoring networks. Ecotoxicol. Environ. Saf. 2021, 216, 112180. [Google Scholar] [CrossRef] [PubMed]
- Cook-Patton, S.C.; Leavitt, S.M.; Gibbs, D.; Harris, N.L.; Lister, K.; Ander-son-Teixeira, K.J.; Briggs, R.D.; Chazdon, R.L.; Crowther, T.W.; Ellis, P.W.; et al. Mapping carbon accumulation potential from global natural forest regrowth. Nature 2020, 585, 545–550. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Q.; Yang, W.; Wang, N.; Fan, P.; You, C.; Yu, L.; Gao, Q.; Wang, H.; Zheng, P. Response mechanisms of 3 typical plants nitrogen and phosphorus nutrient cycling to nitrogen deposition in temperate meadow grasslands. Front. Plant Sci. 2023, 14, 1140080. [Google Scholar] [CrossRef]
- Yu, G.R.; Jia, Y.L.; He, N.P.; Zhu, J.X.; Chen, Z.; Wang, Q.F.; Piao, S.L.; Liu, X.J.; He, H.L.; Guo, X.B.; et al. Stabilization of atmospheric nitrogen deposition in China over the past decade. Nat. Geosci. 2019, 12, 424–429. [Google Scholar] [CrossRef]
- Vogels, J.J.; Van de Waal, D.B.; WallisDeVries, M.F.; Van den Burg, A.B.; Nijssen, M.; Bobbink, R.; Berg, M.P.; Olde Venterink, H.; Siepel, H. Towards a mechanistic understanding of the impacts of nitrogen deposition on producer–consumer interactions. Biol. Rev. 2023, 98, 1712–1731. [Google Scholar] [CrossRef]
- Zhang, G.L.; Wu, Y.J.; Ouyang, S.N.; Duan, H.L.; Wang, J.; Tie, L.H. The impact of nitrogen and phosphorus enrichment on litter decomposition: Soil biota roles and biochemical pathways. Plant Soil 2025, 517, 677–698. [Google Scholar] [CrossRef]
- Kirui, A.; Zhao, W.C.; Deligey, F.; Yang, H.; Kang, X.; Mentink-Vigier, F.; Wang, T. Carbohydrate-aromatic interface and molecular architecture of lignocellulose. Nat. Commun. 2022, 13, 538. [Google Scholar] [CrossRef]
- He, J.H.; Nie, Y.X.; Tan, X.P.; Hu, A.; Li, Z.Q.; Dai, S.P.; Ye, Q.; Zhang, G.X.; Shen, W.J. Latitudinal patterns and drivers of plant lignin and microbial necromass accumulation in forest soils: Disentangling microbial and abiotic controls. Soil Biol. Biochem. 2024, 194, 109438. [Google Scholar] [CrossRef]
- Chen, J.; Luo, Y.Q.; Van Groenigen, K.J.; Huang, B.A.; Cao, J.J.; Zhou, X.H.; Wang, R.W.; Wang, Y. A keystone microbial enzyme for nitrogen control of soil carbon storage. Sci. Adv. 2018, 4, eaaq1689. [Google Scholar] [CrossRef] [PubMed]
- Li, C.H.; Feng, J.G.; Liu, L.; Sun, X.; Li, J.S.; Qian, Z.Z.; Fu, R.R.; Yu, Q.S.; Zhu, B.; Tao, X. Nitrogen addition weakens home-field advantage of litter decomposition by altering soil pH and bacterial communities in asubtropical forest. J. Plant Ecol. 2025, 18, rtaf089. [Google Scholar] [CrossRef]
- Pichon, N.A.; Cappelli, S.L.; Soliveres, S.; Hölzel, N.; Klaus, V.H.; Kleinebecker, T.; Allan, E. Decomposition disentangled: A test of the multiple mechanisms by which nitrogen enrichment alters litter decomposition. Funct. Ecol. 2020, 34, 1485–1496. [Google Scholar] [CrossRef]
- Benito-Carnero, G.; Gartzia-Bengoetxea, N.; Arias-González, A.; Rousk, J. Low-quality carbon and lack of nutrients result in a stronger fungal than bacterial home-field advantage during the decomposition of leaf litter. Funct. Ecol. 2021, 35, 1783–1796. [Google Scholar] [CrossRef]
- Shigyo, N.; Umeki, K.; Hirao, T. Soil microbial identity explains home-field advantage for litter decomposition. New Phytol. 2024, 243, 2146–2156. [Google Scholar] [CrossRef]
- Yang, S.; Jia, Z.; Chang, P.F.; Wu, Y.T.; Huang, J.S.; Wang, J.; Deng, M.F.; Su, J.; Hong, S.B.; He, Y.; et al. Significant impact of UV exposure on litter decomposition across diverse climate zones. Glob. Chang. Biol. 2025, 31, e70456. [Google Scholar] [CrossRef]
- Kong, B.S.; Zhou, J.L.; Qi, L.G.; Jiao, S.Y.; Ma, L.J.; Geng, W.W.; Zhao, Y.H.; Gao, T.; Gong, J.; Li, K.; et al. Effects of nitrogen deposition on leaf litter decomposition and soil organic carbon density in arid and barren rocky mountainous regions: A case study of Yimeng Mountain. Forests 2023, 14, 1351. [Google Scholar] [CrossRef]
- Chen, J.C.; Bai, E.; Liang, Y.T.; Liu, Z.P.; Ji, Y.X.; Sun, T.T.; Guo, Z.X.; Huo, Y.D.; Liu, S.S.; Berg, B. The origin and succession of the microbial community in decomposing litter. ISME Commun. 2025, 5, ycaf155. [Google Scholar] [CrossRef]
- Fanin, N.; Lin, D.; Freschet, G.T.; Keiser, A.D.; Augusto, L.; Wardle, D.A.; Veen, G.G. Home-field advantage of litter decomposition: From the phyllosphere to the soil. New Phytol. 2021, 231, 1353–1358. [Google Scholar] [CrossRef]
- Wang, Y.J.; Zhang, W.T.; Wang, Z.Y.; Lyu, S.X. A polylactic acid degrading lipase from Bacillus safensis: Characterization and structural analysis. Int. J. Biol. Macromol. 2024, 268, 131916. [Google Scholar] [CrossRef]
- Dao, T.T.; Mikutta, R.; Sauheitl, L.; Gentsch, N.; Shibistova, O.; Wild, B.; Schnecker, J.; Bárta, J.; Čapek, P.; Gittel, A.; et al. Lignin preservation and microbial carbohydrate metabolism in permafrost soils. J. Geophys. Res.-Biogeosci. 2022, 127, e2020JG006181. [Google Scholar] [CrossRef]
- Zhang, Z.L.; Bodenheimer, J.; Scott, G.; Dukes, J.S.; Suseela, V. Climatic stress-induced changes in plant chemistry alter the compound-specific degradation of litter during decomposition. New Phytol. 2025, 248, 92–106. [Google Scholar] [CrossRef]
- Anhäuser, T.; Greule, M.; Zech, M.; Kalbitz, K.; McRoberts, C.; Keppler, F. Stable hydrogen and carbon isotope ratios of methoxyl groups during plant litter degradation. Isot. Environ. Health Stud. 2015, 51, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.Q.; Jia, L.L.; Awasthi, M.K.; Jing, G.H.; Wang, Y.B.; He, L.Y.; Zhao, N.; Chen, Z.K.; Zhang, Z.; Shi, X.W. Variations in lignin monomer contents and stable hydrogen isotope ratios in methoxy groups during the biodegradation of garden biomass. Sci. Rep. 2022, 12, 8734. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, S.Z.; Wang, H.Y.; Cao, Y.N.; Hu, J.; Liu, W.G. Apparent fractionation of hydrogen isotope from precipitation to leaf wax n-alkanes from natural environments and manipulation experiments. Sci. Total Environ. 2023, 877, 162970. [Google Scholar] [CrossRef]
- Bhatnagar, J.M.; Peay, K.G.; Treseder, K.K. Litter chemistry influences decomposition through activity of specific microbial functional guilds. Ecol. Monogr. 2018, 88, 429–444. [Google Scholar] [CrossRef]
- Yao, B.; Kong, X.S.; Tian, K.; Zeng, X.Y.; Lu, W.S.; Pang, L.; Sun, S.C.; Tian, X.J. Initial litter chemistry and UV radiation drive chemical divergence in litter during decomposition. Microorganisms 2024, 12, 1535. [Google Scholar] [CrossRef]
- Guo, J.Q.; Liu, X.H.; Ge, W.S.; Zhao, L.J.; Fan, W.J.; Zhang, X.Y.; Lu, Q.Q.; Xing, X.Y.; Zhou, Z.H. Tracking photosynthetic phenology using spectral indices at the leaf and canopy scales in temperate evergreen and deciduous trees. Agric. For. Meteorol. 2024, 344, 109809. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Kang, H.H.; Liu, X.H.; Zhou, J.; Liu, M.D.; Wang, L.X.; Xing, X.Y.; Lu, Q.Q.; Zeng, X.M.; Wei, N.; et al. Comparative foliar atmospheric mercury accumulation across functional types in temperate trees. Environ. Sci. Technol. 2025, 59, 2082–2094. [Google Scholar] [CrossRef]
- Jiang, J.L.; Liu, Y.M.; Mao, J.Y.; Wu, G.X. Extreme heatwave over Eastern China in summer 2022: The role of three oceans and local soil moisture feedback. Environ. Res. Lett. 2023, 18, 044025. [Google Scholar] [CrossRef]
- Zhou, S.; Li, Y.; Wang, J.Y.; He, L.E.; Wang, J.; Guo, Y.X.; Zhao, F.Z. Contrasting soil microbial functional potential for phosphorus cycling in subtropical and temperate forests. Forests 2022, 13, 2002. [Google Scholar] [CrossRef]
- Bastin, J.F.; Finegold, Y.; Garcia, C.; Mollicone, D.; Rezende, M.; Routh, D.; Zohner, C.M.; Crowther, T.W. The global tree restoration potential. Science 2019, 365, 76–79. [Google Scholar] [CrossRef]
- Yang, Z.C.; Guerrieri, R.; Ye, N.; Shao, Y.L.; Yu, F.; Jiao, T.Z.; Yang, H.C.; Chen, J.; Zheng, M.H.; Wang, A.; et al. Underappreciated role of canopy nitrogen deposition for forest productivity. Commun. Earth Environ. 2026, 7, 316. [Google Scholar] [CrossRef]
- Wei, L.; Razavi, B.S.; Wang, W.Q.; Zhu, Z.K.; Liu, S.L.; Wu, J.S.; Kuzyakov, Y.; Ge, T.D. Labile carbon matters more than temperature for enzyme activity in paddy soil. Soil Biol. Biochem. 2019, 135, 134–143. [Google Scholar] [CrossRef]
- Olson, J.S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Wang, X.P.; Ma, Y.S.; Zhang, S.T. Litter quality dependent dynamics of bacteria communities in litter and soil layers during litter decomposition. Plant Soil 2025, 508, 663–675. [Google Scholar] [CrossRef]
- Prescott, C.E.; Vesterdal, L. Decomposition and transformations along the continuum from litter to soil organic matter in forest soils. For. Ecol. Manag. 2021, 498, 119522. [Google Scholar] [CrossRef]
- Wang, A.; Fang, Y.T.; Chen, D.X.; Phillips, O.; Koba, K.; Zhu, W.X.; Zhu, J.J. High nitrogen isotope fractionation of nitrate during denitrification in four forest soils and its implications for denitrification rate estimates. Sci. Total Environ. 2018, 633, 1078–1088. [Google Scholar] [CrossRef]
- Kaiser, C.; Koranda, M.; Kitzler, B.; Fuchslueger, L.; Schnecker, J.; Schweiger, P.; Rasche, F.; Zechmeister-Boltenstern, S.; Sessitsch, A.; Richter, A. Belowground carbon allocation by trees drives seasonal patterns of extracellular enzyme activities by altering microbial community composition in a beech forest soil. New Phytol. 2010, 187, 843–858. [Google Scholar] [CrossRef] [PubMed]
- DeForest, J.L. The influence of time, storage temperature, and substrate age on potential soil enzyme activity in acidic forest soils using MUB-linked substrates and L-DOPA. Soil Biol. Biochem. 2009, 41, 1180–1186. [Google Scholar] [CrossRef]
- Logue, J.B.; Stedmon, C.A.; Kellerman, A.M.; Nielsen, N.J.; Andersson, A.F.; Laudon, H.; Lindström, E.S.; Kritzberg, E.S. Experimental insights into the importance of aquatic bacterial community composition to the degradation of dissolved organic matter. ISME J. 2016, 10, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.D.; Feng, J.G.; Ao, G.K.L.; Qin, W.K.; Han, M.G.; Shen, Y.W.; Liu, M.L.; Chen, Y.; Zhu, B. Globally nitrogen addition alters soil microbial community structure, but has minor effects on soil microbial diversity and richness. Soil Biol. Biochem. 2023, 179, 108982. [Google Scholar] [CrossRef]
- Heidke, I.; Scholz, D.; Hoffmann, T. Quantification of lignin oxidation products as vegetation biomarkers in speleothems and cave drip water. Biogeosciences 2018, 15, 5831–5845. [Google Scholar] [CrossRef]
- Bentler, P.M.; Chou, C.P. Practical issues in structural modeling. Sociol. Methods Res. 1987, 16, 78–117. [Google Scholar] [CrossRef]
- Hong, S.B.; Cong, N.; Ding, J.Z.; Piao, S.L.; Liu, L.L.; Peñuelas, J.; Chen, A.P.; Quine, T.A.; Zeng, H.; Houlton, B.Z. Effects of afforestation on soil carbon and nitrogen accumulation depend on initial soil nitrogen status. Glob. Biogeochem. Cycles 2023, 37, e2022GB007490. [Google Scholar] [CrossRef]
- Xi, J.Z.; Wang, J.Y.; Zhu, Y.F.; Xu, M.P. Nitrogen deposition reduces the rate of leaf litter decomposition: A Global Study. Forests 2024, 15, 1492. [Google Scholar] [CrossRef]
- Vivanco, L.; Austin, A.T. Nitrogen addition stimulates forest litter decomposition and disrupts species interactions in Patagonia, Argentina. Glob. Chang. Biol. 2011, 17, 1963–1974. [Google Scholar] [CrossRef]
- DeForest, J.L. Chronic phosphorus enrichment and elevated pH suppresses Quercus spp. leaf litter decomposition in a temperate forest. Soil Biol. Biochem. 2019, 135, 206–212. [Google Scholar] [CrossRef]
- Rothwell, J.J.; Futter, M.N.; Dise, N.B. A classification and regression tree model of controls on dissolved inorganic nitrogen leaching from European forests. Environ. Pollut. 2008, 156, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Chomel, M.; Guittonny-Larchevque, M.; Baldy, V. Effect of mixing herbaceous litter with tree litters on decomposition and N release in boreal plantations. Plant Soil 2016, 398, 229–241. [Google Scholar] [CrossRef]
- Chen, X.; Cao, J.J.; Sinsabaugh, R.L.; Moorhead, D.L.; Bardgett, R.D.; Fanin, K.; Nottingham, A.T.; Zheng, X.H.; Chen, J. Soil extracellular enzymes as drivers of soil carbon storage under nitrogen addition. Biol. Rev. 2025, 100, 1234–1250. [Google Scholar] [CrossRef]
- Marshall, J.D.; Peichl, M.; Tarvainen, L.; Lim, H.; Lundmark, T.; Nasholm, T.; Oquist, M.; Linder, S. A carbon-budget approach shows that reduced decomposition causes the nitrogen-induced increase in soil carbon in a boreal forest. For. Ecol. Manag. 2021, 502, 119750. [Google Scholar] [CrossRef]
- Zhao, J.N.; Feng, X.H.; Hu, J.; He, M.; Wang, S.Y.; Yang, Y.H.; Chen, L.Y. Mineral and microbial properties drive the formation of mineral-associated organic matter and its response to increased temperature. Glob. Chang. Biol. 2024, 30, e70004. [Google Scholar] [CrossRef]
- Mao, Z.X.; Yue, M.; Wang, Y.C.; Li, L.J.; Li, Y. Soil microorganisms mediated the responses of the plant–soil systems of Neotrinia splendens to nitrogen addition and warming in a desert ecosystem. Agronomy 2024, 14, 132. [Google Scholar] [CrossRef]
- Erdenebileg, E.; Wang, C.W.; Yu, W.Y.; Ye, X.H.; Pan, X.; Huang, Z.Y.; Liu, G.F.; Cornelissen, J.H.C. Carbon versus nitrogen release from root and leaf litter is modulated by litter position and plant functional type. J. Ecol. 2023, 111, 198–213. [Google Scholar] [CrossRef]
- Yin, H.Y.; Xu, L.B.; Porter, N.A. Free radical lipid peroxidation: Mechanisms and analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.X.; Zhang, L.X.; Wang, J.C.; Li, G.; Qiu, Y.Z.; Ren, L.H.; Wang, P. Microbial enzymes for the biodegradation of polylactic acid (PLA): Molecular docking simulation and biodegradation pathway proposal. Environ. Rev. 2024, 33, 1–18. [Google Scholar] [CrossRef]
- Qin, N.; Li, L.Y.; Wang, Z.; Shi, S.B. Microbial production of odd-chain fatty acids. Biotechnol. Bioeng. 2023, 120, 917–931. [Google Scholar] [CrossRef]
- Thomas, C.L.; Jansen, B.; van Loon, E.E.; Wiesenberg, G.L. Transformation of n- alkanes from plant to soil: A review. Soil 2021, 7, 785–809. [Google Scholar] [CrossRef]
- Cook, S.D. An historical review of phenylacetic acid. Plant Cell Physiol. 2019, 60, 243–254. [Google Scholar] [CrossRef]
- Nike, P.I.; Pérez Pantoja, D.; de Lorenzo, V. Pyridine nucleotide transhydrogenases enable redox balance of Pseudomonas putida during biodegradation of aromatic compounds: Transhydrogenases, Redox Balance and Biodegradation. Environ. Microbiol. 2016, 18, 3565–3582. [Google Scholar] [CrossRef]
- Heng, Y.C.; Wong, G.W.J.; Kittelmann, S. Expanding the biosynthesis spectrum of hydroxy fatty acids: Unleashing the potential of novel bacterial fatty acid hydratases. Biotechnol. Biofuels 2024, 17, 131. [Google Scholar] [CrossRef]
- Evangelista, S.; Cooper, D.G.; Yargeau, V. The effect of structure and a secondary carbon source on the microbial degradation of chlorophenoxy acids. Chemosphere 2010, 79, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Córdova, S.C.; Olk, D.C.; Dietzel, R.N.; Mueller, K.E.; Archontouilis, S.V.; Castellano, M.J. Plant litter quality affects the accumulation rate, composition, and stability of mineral-associated soil organic matter. Soil Biol. Biochem. 2018, 125, 115–124. [Google Scholar] [CrossRef]
- Chen, F.S.; Wang, G.G.; Fang, X.M.; Wan, S.Z.; Zhang, Y.; Liang, C. Nitrogen deposition effect on forest litter decomposition is interactively regulated by endogenous litter quality and exogenous resource supply. Plant Soil 2019, 437, 413–426. [Google Scholar] [CrossRef]
- Craig, M.E.; Geyer, K.M.; Beidler, K.V.; Brzostek, E.R.; Frey, S.D.; Grandy, A.S.; Liang, C.; Phillips, R.P. Fast-decaying plant litter enhances soil carbon in temperate forests but not through microbial physiological traits. Nat. Commun. 2022, 13, 1229. [Google Scholar] [CrossRef]
- Zhou, J.C.; Luo, Y.Q.; Chen, J. Dilemmas in linking microbial carbon use efficiency with soil organic carbon dynamics. Glob. Chang. Biol. 2025, 31, e70047. [Google Scholar] [CrossRef]
- Chen, H.; Li, D.J.; Zhao, J.; Zhang, W.; Xiao, K.C.; Wang, K.L. Nitrogen addition aggravates microbial carbon limitation: Evidence from ecoenzymatic stoichiometry. Geoderma 2018, 329, 61–64. [Google Scholar] [CrossRef]
- Song, Y.Y.; Song, C.C.; Ren, J.S.; Tan, W.W.; Jin, S.F.; Jiang, L. Influence of nitrogen additions on litter decomposition, nutrient dynamics, and enzymatic activity of two plant species in a peatland in Northeast China. Sci. Total Environ. 2018, 625, 640–646. [Google Scholar] [CrossRef]
- Wu, Q.X.; Ni, X.Y.; Sun, X.Y.; Chen, Z.H.; Hong, S.B.; Berg, B.; Zheng, M.H.; Chen, J.; Zhu, J.J.; Ai, L.; et al. Substrate and climate determine terrestrial litter decomposition. Proc. Natl. Acad. Sci. USA 2025, 122, e2420664122. [Google Scholar] [CrossRef]
- Tian, D.S.; Niu, S.L. A global analysis of soil acidification caused by nitrogen addition. Environ. Res. Lett. 2015, 10, 024019. [Google Scholar] [CrossRef]
- Gao, M.X.; Lin, G.G.; Zhu, F.F.; Wu, Z.; Gundersen, P.; Zeng, D.H.; Hobbie, E.A.; Zhu, W.X.; Fang, Y.T. Higher resistance of larch-broadleaf mixed forests than larch forests against soil acidification under experimental nitrogen addition. Plant Soil 2024, 505, 335–349. [Google Scholar] [CrossRef]
- Wu, J.P.; Liu, W.F.; Zhang, W.X.; Shao, Y.H.; Duan, H.L.; Chen, B.D.; Wei, X.H.; Fan, H.B. Long-term nitrogen addition changes soil microbial community and litter decomposition rate in a subtropical forest. Appl. Soil Ecol. 2019, 142, 43–51. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, X.L.; Liu, L.X.; Li, T.; Dou, Y.X.; Qiao, J.B.; Wang, Y.Q.; An, S.S.; Chang, S.X. Nitrogen fertilization weakens the linkage between soil carbon and microbial diversity: A global meta-analysis. Glob. Chang. Biol. 2022, 28, 6446–6461. [Google Scholar] [CrossRef]
- Sauvadet, M.; Fanin, N.; Chauvat, M.; Bertrand, I. Can the comparison of above-and below-ground litter decomposition improve our understanding of bacterial and fungal successions? Soil Biol. Biochem. 2019, 132, 24–27. [Google Scholar] [CrossRef]
- Kroeger, M.E.; DeVan, M.R.; Thompson, J.; Johansen, R.; Galle-gos-Graves, L.V.; Lopez, D.; Runde, A.; Yoshida, T.; Munsky, B.; Sevanto, S.; et al. Microbial community composition controls carbon flux across litter types in early phase of litter decomposition. Environ. Microbiol. 2021, 23, 6676–6693. [Google Scholar] [CrossRef]
- Lin, H.; Li, Y.N.; Bruelheide, H.; Zhang, S.R.; Ren, H.B.; Zhang, N.L.; Ma, K.P. What drives leaf litter decomposition and the decomposer community in subtropical forests–The richness of the above-ground tree community or that of the leaf litter? Soil Biol. Biochem. 2021, 160, 108314. [Google Scholar] [CrossRef]
- Hobbie, E.A.; Diepen, L.T.A.V.; Lilleskov, E.A.; Ouimette, A.P.; Finzi, A.C.; Hofmockel, K.S. Fungal functioning in a pine forest: Evidence from a 15N-labeled global change experiment. New Phytol. 2014, 201, 1431–1439. [Google Scholar] [CrossRef] [PubMed]
- Brewer, T.E.; Handley, K.M.; Carini, P.; Gilbert, J.A.; Fierer, N. Genome reduction in an abundant and ubiquitous soil bacterium ‘Candidatus Udaeobacter copiosus’. Nat. Microbiol. 2016, 2, 16198. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, F.L.P.; Giger, W.; Guenther, K.; Kohler, H.P.E. Differential Degradation of Nonylphenol Isomers by Sphingomonas xenophaga Bayram. Appl. Environ. Microbiol. 2005, 71, 1123–1129. [Google Scholar] [CrossRef] [PubMed]
- Canessa, R.; van den Brink, L.; Saldaña, A.; Rios, R.S.; Hättenschwiler, S.; Mueller, C.W.; Prater, I.; Tielbörger, K.; Bader, M.Y. Relative effects of climate and litter traits on decomposition change with time, climate and trait variability. J. Ecol. 2021, 109, 447–458. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Lu, Q.; Cai, X.; Zeng, X.; Chen, J.; Chang, F.; Jing, G.; Guo, J.; Zhou, S.; Chen, Z.; Jia, L.; et al. Nitrogen Addition-Induced Variations in Stoichiometric Ratio of Organic Acids from Litter Decomposition in a Temperate Forest. Forests 2026, 17, 622. https://doi.org/10.3390/f17050622
Lu Q, Cai X, Zeng X, Chen J, Chang F, Jing G, Guo J, Zhou S, Chen Z, Jia L, et al. Nitrogen Addition-Induced Variations in Stoichiometric Ratio of Organic Acids from Litter Decomposition in a Temperate Forest. Forests. 2026; 17(5):622. https://doi.org/10.3390/f17050622
Chicago/Turabian StyleLu, Qiangqiang, Xinping Cai, Xiaomin Zeng, Ji Chen, Fan Chang, Guanghua Jing, Jiaqi Guo, Sha Zhou, Zhikun Chen, Lili Jia, and et al. 2026. "Nitrogen Addition-Induced Variations in Stoichiometric Ratio of Organic Acids from Litter Decomposition in a Temperate Forest" Forests 17, no. 5: 622. https://doi.org/10.3390/f17050622
APA StyleLu, Q., Cai, X., Zeng, X., Chen, J., Chang, F., Jing, G., Guo, J., Zhou, S., Chen, Z., Jia, L., Liu, J., & Liu, T. (2026). Nitrogen Addition-Induced Variations in Stoichiometric Ratio of Organic Acids from Litter Decomposition in a Temperate Forest. Forests, 17(5), 622. https://doi.org/10.3390/f17050622