
Influences of the China–Russia Crude Oil Pipelines on the Characteristics of Soil Bacterial and Fungal Communities in Permafrost Regions of the Da Xing’anling Mountains, Northeast China

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design and Sampling
2.3. Laboratory Analysis and Determination
2.4. Soil Microbial DNA Extraction and High-Throughput Sequencing
2.5. Data Analysis
3. Results
3.1. Effects of Pipeline Engineering Disturbances on Soil Physicochemical Properties
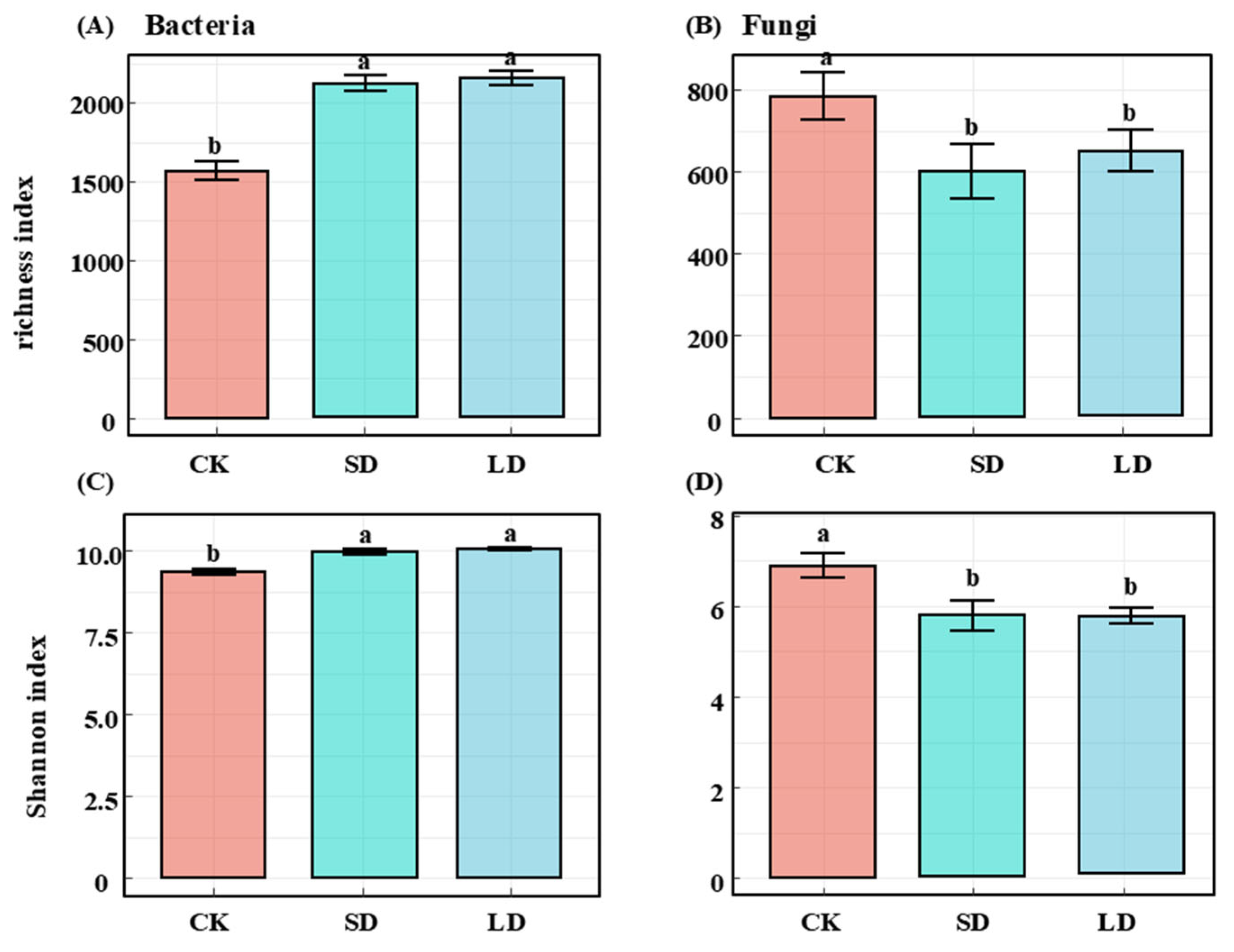
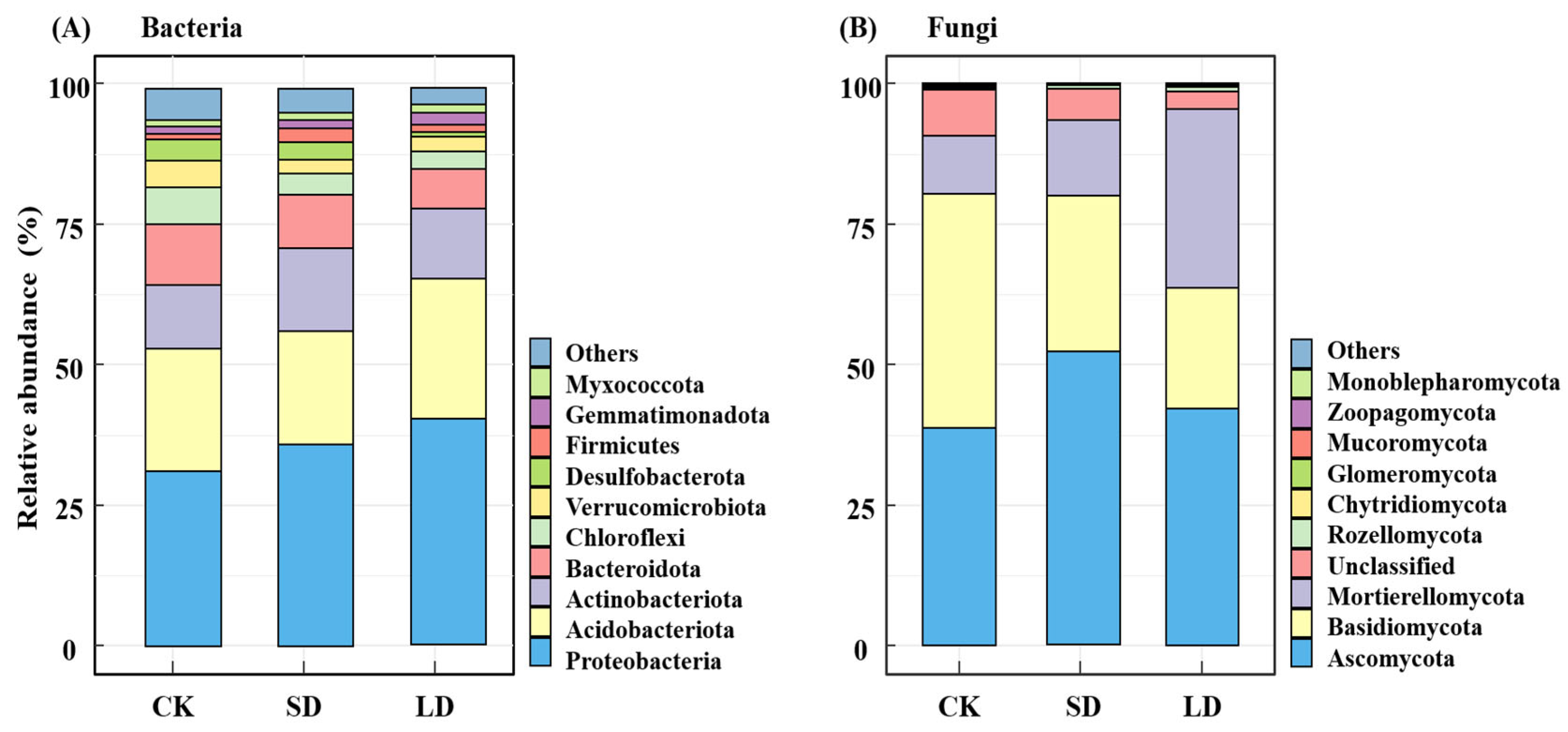
3.2. Effects of Pipeline Engineering Disturbances on Diversity and Composition of Bacterial and Fungal Communities
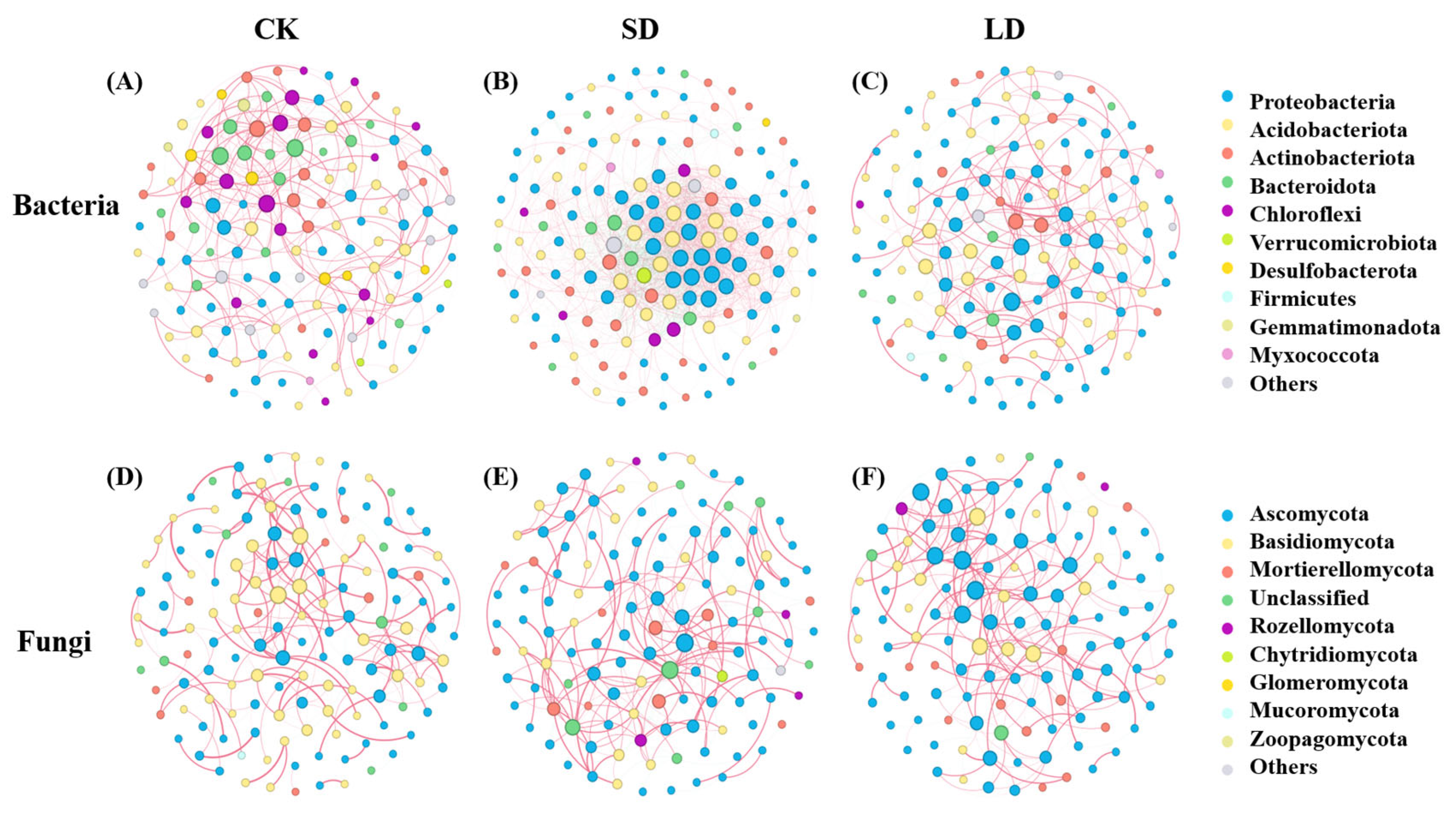
3.3. Co-Occurrence Networks of Soil Bacterial and Fungal Communities Under Pipeline Engineering Disturbances
3.4. Relationship Between Soil Microbial Communities and Soil Physicochemical Properties
4. Discussion
4.1. Changes in Soil Physicochemical Properties Due to Pipeline Engineering Disturbance
4.2. Microbial Community Responses and Underlying Mechanisms
4.3. Implications for Ecological Restoration and Management in Permafrost Regions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fan, X.W.; Lin, Z.J.; Gao, Z.Y.; Meng, X.L.; Niu, F.J.; Luo, J.; Yin, G.A.; Zhou, F.J.; Lan, A.Y. Cryostructures and ground ice content in ice-rich permafrost area of the Qinghai-Tibet Plateau with Computed Tomography Scanning. J. Mt. Sci. 2021, 18, 1208–1221. [Google Scholar] [CrossRef]
- Luo, D.; Jin, H.J.; Marchenko, S.S.; Romanovsky, V.E. Difference between near-surface air, land surface and ground surface temperatures and their influences on the frozen ground on the Qinghai-Tibet Plateau. Geoderma 2018, 312, 74–85. [Google Scholar] [CrossRef]
- Li, X.; Cui, Y.; Ma, D.; Song, D.; Liu, L. Vertical distribution of bacterial community diversity in the Greater Khingan Mountain permafrost region. Ecol. Evol. 2022, 12, e9106. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Li, G.Y.; Cao, Y.P.; Chen, D.; Wu, G.; Du, Q.S.; Wang, F.; Alexander, F.; Che, F.Q.; Zhang, Z.R.; et al. Permafrost thawing caused by the China-Russia Crude oil pipeline based on multi-type data and its impacts on geomorphological reshaping and water erosion. Catena 2024, 242, 108134. [Google Scholar] [CrossRef]
- Li, G.Y.; Cao, Y.P.; Ma, W.; Jin, X.Y.; Chen, P.C.; Yu, Q.H.; Zhang, Z.Q.; Mu, Y.H.; Jin, H.J. Permafrost engineering problem along China-Russia Crude Oil Pipeline and mitigative measure. Bull. Chin. Acad. Sci. 2021, 36, 150–159. [Google Scholar] [CrossRef]
- Wang, F.; Li, G.; Ma, W.; Wu, Q.; Serban, M.; Vera, S.; Alexandr, F.; Jiang, N.; Wang, B. Pipeline–permafrost interaction monitoring system along the China–Russia crude oil pipeline. Eng. Geol. 2019, 254, 113–125. [Google Scholar] [CrossRef]
- Jin, H.J.; Hao, J.; Chang, X.; Zhang, J.; Yu, Q.H.; Qi, J.; Lü, L.; Wang, S. Zonation and assessment of frozen-ground conditions for engineering geology along the China–Russia crude oil pipeline route from Mo’he to Daqing, Northeastern China. Cold Reg. Sci. Technol. 2010, 64, 213–225. [Google Scholar] [CrossRef]
- Chai, M.T.; Li, G.Y.; Ma, W.; Cao, Y.P.; Wu, G.; Mu, Y.H.; Chen, D.; Zhang, J.; Zhou, Z.W.; Zhou, Y.; et al. Assessment of freeze–thaw hazards and water features along the China–Russia Crude Oil Pipeline in permafrost regions. Remote Sens. 2020, 12, 3576. [Google Scholar] [CrossRef]
- Li, X.Y.; Jin, X.Y.; Wang, X.B.; Jin, H.J.; Tang, L.; Li, X.Y.; He, R.X.; Li, Y.; Huang, C.J.; Zhang, S.F. Investigation of permafrost engineering geological environment with electrical resistivity tomography: A case study along the China-Russia crude oil pipelines. Eng. Geol. 2021, 291, 106237. [Google Scholar] [CrossRef]
- Wang, F.; Li, G.Y.; Ma, W.; Mu, Y.H.; Zhou, Z.W.; Mao, Y.C. Permafrost thawing along the China-Russia Crude Oil Pipeline and countermeasures: A case study in Jiagedaqi, Northeast China. Cold Reg. Sci. Technol. 2018, 155, 308–313. [Google Scholar] [CrossRef]
- Olson, E.R.; Doherty, J.M. The legacy of pipeline installation on the soil and vegetation of southeast Wisconsin wetlands. Ecol. Eng. 2012, 39, 53–62. [Google Scholar] [CrossRef]
- Wang, J.H.; Cai, T.J.; Ge, S.S.; Liu, J.X.; Qu, C.Y.; Sun, X.X. Effects of Mohe-Daqing oil pipeline project construction on typical forest ecosystems in Daxing’an. J. Beijing For. Univ. 2015, 37, 58–66. [Google Scholar] [CrossRef]
- Wu, M.H.; Chen, S.Y.; Chen, J.W.; Xue, K.; Chen, S.L.; Wang, X.M.; Chen, T.; Kang, S.C.; Rui, J.P.; Thies, J.E.; et al. Reduced microbial stability in the active layer is associated with carbon loss under alpine permafrost degradation. Proc. Natl. Acad. Sci. USA 2021, 118, e2025321118. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Li, F.; Sun, Q.Y.; Xie, Y.H.; Zhang, J.; Wang, Z. Review on the Study of Soil Microorganisms in Wetland Ecosystems. Chin. J. Appl. Environ. Biol. 2013, 19, 547–552. [Google Scholar] [CrossRef]
- Philippot, L.; Griffiths, B.S.; Langenheder, S. Microbial community resilience across ecosystems and multiple disturbances. Microbiol. Mol. Biol. Rev. 2021, 85, e00026-20. [Google Scholar] [CrossRef]
- Yu, X.F.; Wang, G.P.; Zou, Y.C.; Wang, Q.; Zhao, H.M.; Lu, X.G. Effects of Pipeline Construction on Wetland Ecosystems: Russia–China Oil Pipeline Project (Mohe-Daqing Section). Ambio 2010, 39, 447–450. [Google Scholar] [CrossRef]
- Rui, W.H.; Chen, J.X.; Wang, Y.G.; Zhang, T.; Liu, H.; Li, M. Effects of Natural Gas Pipeline Construction on Soil Nutrients and Bacteria Diversity in the Fragile Loess Region. Acta Sci. Circumst. 2018, 38, 3278–3285. [Google Scholar] [CrossRef]
- Jin, X.Y.; Jin, H.J.; Yang, X.; Wang, W.H.; Huang, S.; Zhang, S.R.; Yang, S.Q.; Li, X.Y.; Wang, H.W.; He, R.X.; et al. Shrubification along pipeline corridors in permafrost regions. Forests 2022, 13, 1093. [Google Scholar] [CrossRef]
- Xiao, J.; Wang, Y.F.; Shi, P.; Yang, L.; Chen, L.D. Potential effects of large linear pipeline construction on soil and vegetation in ecologically fragile regions. Environ. Monit. Assess. 2014, 186, 8037–8048. [Google Scholar] [CrossRef]
- Su, H.; Mi, S.F.; Peng, X.W.; Han, Y.J. The mutual influence between corrosion and the surrounding soil microbial communities of buried petroleum pipelines. RSC Adv. 2019, 9, 18930–18940. [Google Scholar] [CrossRef]
- Ji, M.K.; Kong, W.D.; Liang, C.; Zhang, T.Q.; Jia, H.Z.; Dong, X.B. Permafrost thawing exhibits a greater influence on bacterial richness and community structure than permafrost age in Arctic permafrost soils. Cryosphere 2020, 16, 1–22. [Google Scholar] [CrossRef]
- Barbato, R.A.; Jones, R.M.; Douglas, T.A.; Doherty, S.J.; Messan, X.K.; Foley, K.L.; Perkins, E.J.; Thurston, A.K.; Garcia-Reyero, N. Not all permafrost microbiomes are created equal: Influence of permafrost thaw on the soil microbiome in a laboratory incubation study. Soil Biol. Biochem. 2022, 167, 108605. [Google Scholar] [CrossRef]
- Perez-Mon, C.; Stierli, B.; Plötze, M.; Frey, B. Fast and persistent responses of alpine permafrost microbial communities to in situ warming. Sci. Total Environ. 2022, 807, 150720. [Google Scholar] [CrossRef] [PubMed]
- Messan, K.S.; Jones, R.M.; Doherty, S.J.; Foley, K.; Douglas, T.A.; Barbato, R.A. The role of changing temperature in microbial metabolic processes during permafrost thaw. PLoS ONE 2020, 15, e0232169. [Google Scholar] [CrossRef]
- Pei, G.T.; Sun, J.F.; He, T.X.; Hu, B.Q. Effects of long-term human disturbances on soil microbial diversity and community structure in a karst grassland ecosystem of northwestern Guangxi, China. Chin. J. Plant Ecol. 2021, 45, 74–84. [Google Scholar] [CrossRef]
- Bonomo, M.G.; Scrano, L.; Mang, S.M.; Scalese, B.E.; Bufo, S.A.; Modley, L.A.; Buongarzone, E.; Salzano, G. Changes in the Bacterial Community Composition of Cultivated Soil after Digging up Operations for Laying a Pipeline. Agriculture 2023, 13, 1189. [Google Scholar] [CrossRef]
- Tang, M.; Chen, Q.; Xiao, X.Z.; Lyu, Y.; Sun, W.L. Differential impacts of water diversion and environmental factors on bacterial, archaeal, and fungal communities in the eastern route of the South-to-North water diversion project. Environ. Int. 2025, 195, 109280. [Google Scholar] [CrossRef]
- Cameron, E.A.; Lantz, T.C. Persistent Changes to Ecosystems following Winter Road Construction and Abandonment in an Area of Discontinuous Permafrost, Nahanni National Park Reserve, Northwest Territories, Canada. Arct. Antarct. Alp. Res. 2017, 49, 259–276. [Google Scholar] [CrossRef]
- Yang, S.Z.; Wen, X.; Jin, H.J.; Wu, Q.B. Pyrosequencing Investigation into the Bacterial Community in Permafrost Soils along the China-Russia Crude Oil Pipeline (CRCOPs). PLoS ONE 2012, 7, e52730. [Google Scholar] [CrossRef]
- Sun, S.; Li, S.; Avera, B.N.; Strahm, B.D.; Badgley, B.D. Soil Bacterial and Fungal Communities Show Distinct Recovery Patterns during Forest Ecosystem Restoration. Appl. Environ. Microbiol. 2017, 83, e00966-17. [Google Scholar] [CrossRef]
- Dong, X.; Liu, C.; Ma, D.L.; Wu, Y.F.; Man, H.R.; Wu, X.W.; Li, M.; Zang, S.Y. Organic Carbon Mineralization and Bacterial Community of Active Layer Soils Response to Short-Term Warming in the Great Hing’an Mountains of Northeast China. Front. Microbiol. 2021, 12, 802213. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.Y.; Li, M.S.; Liu, X.L.; Yin, W.P.; Li, G.F.; Mu, L.Q.; Cui, X.Y.; Cheng, Z.C. Soil Bacterial Community Composition and Diversity of Typical Permafrost in Greater Khingan Mountains. Microbiol. China. 2020, 47, 2759–2770. [Google Scholar] [CrossRef]
- Wahdan, S.F.M.; Ji, L.; Schädler, M.; Wu, Y.T.; Sansupa, C.; Tanunchai, B.; Buscot, F.; Purahong, W. Future climate conditions accelerate wheat straw decomposition alongside altered microbial community composition, assembly patterns, and interaction networks. ISME J. 2023, 17, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.H.; Zheng, Y.T.; Li, P.F.; Cui, J.X.; Sui, P.; Chen, Y.Q.; Gao, W.S. Organic management increases beneficial microorganisms and promotes the stability of microecological networks in tea plantation soil. Front. Microbiol. 2023, 14, 1237842. [Google Scholar] [CrossRef]
- Cui, J.; Cai, T.J.; Yang, Y.; Yang, S. Effects of Mohe-Daqing Oil Pipeline Poroject on Soil Nutrientin the Areas Along the Line. J. Soil Water Conserv. 2013, 27, 143–149. [Google Scholar] [CrossRef]
- Shao, M.; Zhang, S.Y.; Niu, B.; Pei, Y.; Song, S.; Lei, T.Z.; Yun, H.B. Soil texture influences soil bacterial biomass in the permafrost-affected alpine desert of the Tibetan plateau. Front. Microbiol. 2022, 13, 1007194. [Google Scholar] [CrossRef]
- Yang, K.; Zhu, J.J.; Zhang, W.W.; Zhang, Q.; Lu, D.L.; Zhang, Y.K.; Zheng, X.; Xu, S.; Wang, G.G. Litter decomposition and nutrient release from monospecific and mixed litters: Comparisons of litter quality, fauna and decomposition site effects. J. Ecol. 2022, 110, 1673–1686. [Google Scholar] [CrossRef]
- Liang, Q.Y.; Fu, D.X.; Zhao, J.M.; Wang, Z.X.; Wang, X.Y.; Li, X.G. Effects of Terrain on Soil Hydrological Characteristics of Alpine Grasslands. J. For. Environ. 2024, 44, 367–375. [Google Scholar] [CrossRef]
- Brehm, T.; Culman, S.W. Pipeline installation effects on soils and plants: A review and quantitative synthesis. Agrosyst. Geosci. Environ. 2022, 5, e20312. [Google Scholar] [CrossRef]
- Richman, D.; Tucker, C.L.; Koehler, P.G. Influence of Portland cement amendment on soil pH and residual soil termiticide performance. Pest Manag. Sci. 2006, 62, 1216–1223. [Google Scholar] [CrossRef]
- Soon, Y.K.; Rice, W.A.; Arshad, M.A.; Millis, P.F. Effect of pipeline installation on crop yield and some biological properties of boreal soils. Can. J. Soil Sci. 2000, 80, 483–488. [Google Scholar] [CrossRef]
- Shi, P.; Xiao, J.; Wang, Y.F.; Chen, L.D. Assessment of Ecological and Human Health Risks of Heavy Metal Contamination in Agriculture Soils Disturbed by Pipeline Construction. Int. J. Environ. Res. Public Health 2014, 11, 2504–2520. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Xiao, J.; Wang, Y.F.; Chen, L.D. The effects of pipeline construction disturbance on soil properties and restoration cycle. Environ. Monit. Assess. 2014, 186, 1825–1835. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dong, S.K.; Gao, Q.Z.; Liu, S.L.; Ganjurjav, H.; Wang, X.X.; Su, X.K.; Wu, X.Y. Soil bacterial and fungal diversity differently correlated with soil biochemistry in alpine grassland ecosystems in response to environmental changes. Sci Rep. 2017, 7, 43077. [Google Scholar] [CrossRef]
- Shi, Y.X.; Zhou, J.Q.; Sun, Y.H.; Yang, L.; Chen, T.; Li, R.; Wang, X. The variation trends of plant and soil fungi communities in alpine grassland along the Sichuan-Tibet Railway. Acta Agrestia Sin. 2024, 32, 2707–2717. [Google Scholar] [CrossRef]
- Terrat, S.; Horrigue, W.; Dequietd, S.; Saby, N.; Lelièvre, M.; Nowak, V.; Tripied, J.; Régnier, T.; Jolivet, C.; Arrouays, D.; et al. Mapping and predictive variations of soil bacterial richness across France. PLoS ONE 2017, 12, e0186766. [Google Scholar] [CrossRef]
- Seaton, F.M.; George, P.B.L.; Lebron, I.; Jones, D.L.; Creer, S.; Robinson, D.A. Soil textural heterogeneity impacts bacterial but not fungal diversity. Soil Biol. Biochem. 2020, 144, 107766. [Google Scholar] [CrossRef]
- Wang, C.; Yu, Q.Y.; Ji, N.N.; Zheng, Y.; Taylo, J.W.; Guo, L.D.; Gao, C. Bacterial genome size and gene functional diversity negatively correlate with taxonomic diversity along a pH gradient. Nat. Commun. 2023, 14, 7437. [Google Scholar] [CrossRef]
- Ren, B.H.; Hu, Y.M.; Bu, R.C. Vertical distribution patterns and drivers of soil bacterial communities across the continuous permafrost region of northeastern China. Ecol. Process. 2022, 11, 1–13. [Google Scholar] [CrossRef]
- Liu, Z.H.; Huang, F.Y.; Li, J.L.; Zhang, P.; Yang, B.P.; Ding, R.X.; Nie, J.F.; Jia, Z.K. Effects of farmland mulching patterns on soil microbial diversity and community structure in dryland. Acta Ecol. Sin. 2021, 41, 2750–2760. [Google Scholar] [CrossRef]
- Zhao, Y.T.; Li, W.Y.; Huo, R.; Wu, C.B.; Di, Y.L.; Shi, K.; Zhou, S.L. Response mechanism of bacterial community and physicochemical factors evolution to ecological restoration in sediment and water dual medium of Baiyangdian Lake. J. Water Process Eng. 2025, 70, 106949. [Google Scholar] [CrossRef]
- Yu, H.; Liu, Y.; Deng, Y.; Lu, G.X.; Yan, H.L.; Wang, Y.C. Effects of the Transformation from Natural Alpine Grassland to Mixed Artificial Grassland on the Characteristics of Soil Microbial Community. Environ. Sci. 2023, 44, 2928–2935. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.M.; Qu, M.J.; Shao, S.; Li, J.W. Soil fungal guilds as important integrators linking plant richness and carbon, nitrogen and phosphorus stocks in oasis–desert ecosystems. Soil Biol. Biochem. 2023, 177, 108930. [Google Scholar] [CrossRef]
- Xia, Q.; Rufty, T.; Shi, W. Soil microbial diversity and composition: Links to soil texture and associated properties. Soil Biol. Biochem. 2020, 149, 107953. [Google Scholar] [CrossRef]
- Zhao, P.P.; Huang, Y.T.; Liu, B.Y.; Chen, J.Y.; Lei, Z.Y.; Zhang, Y.H.; Cheng, B.H.; Zhou, T.; Peng, S.L. Effects of daytime and nighttime warming on soil microbial diversity. Geoderma 2024, 447, 116909. [Google Scholar] [CrossRef]
- Yang, H.L.; Cheng, L.; Che, L.G.; Su, Y.Z.; Li, Y.L. Nutrients addition decreases soil fungal diversity and alters fungal guilds and co-occurrence networks in a semi - arid grassland in northern China. Sci. Total Environ. 2024, 926, 172100. [Google Scholar] [CrossRef]
- Shen, C.C.; Xiong, J.B.; Zhang, H.Y.; Feng, Y.Z.; Li, X.J.; Li, X.Y.; Liang, W.J.; Chu, H.Y. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Jones, R.T.; Robeson, M.S.; Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses. ISME J. 2009, 3, 442–453. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Alvarez, A.; Saez, J.M.; Costa, J.S.D.; Colin, V.L.; Fuentes, M.S.; Cuozzo, S.A.; Benimeli, C.S.; Polti, M.A.; Amoroso, M.J. Actinobacteria: Current research and perspectives for bioremediation of pesticides and heavy metals. Chemosphere 2017, 166, 41–62. [Google Scholar] [CrossRef]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing - based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.H.; Lu, D.X.; Jin, H.; Yang, A.L.; Qin, P.; Wei, J.Q.; Guo, R.; Zhang, W.Q. Relationship between the bacterial community and environmental factors in the rhizosphere soil of wild morels in Gansu. Microbiol. China. 2022, 49, 514–528. [Google Scholar]
- Naumoff, D.G.; Dedysh, S.N. Lateral gene transfer between the Bacteroidetes and Acidobacteria: The case of α-l-rhamnosidases. FEBS Lett. 2012, 586, 3843–3851. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.S.; Wang, S.S.; Fang, W.; Zheng, M.Q.; Jiang, B.H.; Shao, S.; Ma, X.M.; Xu, Q.F. Bamboo invasion surrounding forest increased soil pH, changed soil chemical nutrient and microbial community: A meta-analysis. Acta Pedol. Sin. 2024, 61, 862–877. [Google Scholar] [CrossRef]
- Wang, M.; Lin, M.; Liu, Q.H.; Li, C.; Pang, X.Y. Fungal, but not bacterial, diversity and network complexity promote network stability during roadside slope restoration. Sci. Total Environ. 2024, 922, 171007. [Google Scholar] [CrossRef]
- Wu, H.K.; Ji, S.; Qiu, X.X.; Du, S.B.; Xie, H.C. Structural characteristics of soil fungal communities at different altitudes on the southern slope of the Qilian Mountains. J. For. Environ. 2024, 44, 571–580. [Google Scholar] [CrossRef]
- Li, K.; DiLegge, M.J.; Minas, I.S.; Hamm, A.; Manter, D.; Vivanco, J.M. Soil sterilization leads to re-colonization of a healthier rhizosphere microbiome. Rhizosphere 2019, 12, 100176. [Google Scholar] [CrossRef]
- Shade, A.; Peter, H.; Allison, S.D.; Baho, D.L.; Berga, M.; Bürgmann, H.; Huber, D.H.; Langenheder, S.; Lennon, J.T.; Martiny, J.B.H.; et al. Fundamentals of Microbial Community Resistance and Resilience. Front. Microbiol. 2012, 3, 417. [Google Scholar] [CrossRef]
- Allison, S.D.; Martiny, J.B.H. Resistance, resilience, and redundancy in microbial communities. Proc. Natl. Acad. Sci. USA 2008, 105 (Suppl. 1), 11512–11519. [Google Scholar] [CrossRef]
- Santillan, E.; Neshat, S.A.; Wuertz, S. Disturbance and stability dynamics in microbial communities for environmental biotechnology applications. Curr. Opin. Biotechnol. 2025, 93, 103304. [Google Scholar] [CrossRef]
- Chen, L.; Yan, H.; Di, S.S.; Guo, C.; Zhang, H.; Zhang, S.Q.; Gold, A.; Wang, Y.; Hu, M.; Wu, D.Y.; et al. Mapping Pesticide-Induced Metabolic Alterations in Human Gut Bacteria. bioRxiv. 2024, 16, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Inglese, C.N.; Christiansen, C.T.; Lamhonwah, D.; Moniz, K.; Montross, S.N.; Lamoureux, S.; Lafrenière, M.; Grogan, P.; Walker, V.K. Examination of soil microbial communities after permafrost thaw subsequent to an active layer detachment in the high Arctic. Arct. Antarct. Alpine Res. 2017, 49, 455–472. [Google Scholar] [CrossRef]
- Medriano, C.A.; Chan, A.; De Sotto, R.; Bae, S. Different types of land use influence soil physicochemical properties, the abundance of nitrifying bacteria, and microbial interactions in tropical urban soil. Sci. Total Environ. 2023, 869, 161722. [Google Scholar] [CrossRef] [PubMed]
- Tardy, V.; Mathieu, O.; Lévêque, J.; Terrat, S.; Chabbi, A.; Lemanceau, P.; Ranjard, L.; Maron, P.-A. Stability of soil microbial structure and activity depends on microbial diversity. Environ. Microbiol. Rep. 2014, 6, 173–183. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Norton, D.A. Towards a conceptual framework for restoration ecology. Restor. Ecol. 1996, 4, 93–110. [Google Scholar] [CrossRef]
- Wagg, C.; Bender, S.F.; Widmer, F.; Van Der Heijden, M.G.A. Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proc. Natl. Acad. Sci. USA 2014, 111, 5266–5270. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Property | CK | SD | LD |
|---|---|---|---|
| clay (%) | 3.39 ± 0.45 b | 7.44 ± 0.41 a | 7.65 ± 0.39 a |
| silt (%) | 59.26 ± 1.99 a | 57.99 ± 1.03 a | 55.65 ± 1.56 a |
| sand (%) | 37.34 ± 2.38 a | 34.56 ± 1.11 a | 36.71 ± 1.57 a |
| SWC (%) | 405.65 ± 26.9 a | 60.61 ± 16.33 b | 85 ± 21.16 b |
| pH | 4.16 ± 0. 04 b | 4.71 ± 0.03 a | 4.62 ± 0.03 a |
| SOC (g kg−1) | 356.74 ± 18.67 a | 131.57 ± 18.15 b | 105.59 ± 14.32 b |
| TN (g kg−1) | 19.47 ± 1.01 a | 7.63 ± 0.97 b | 6.33 ± 0.82 b |
| TP (g kg−1) | 1.49 ± 0.06 a | 0.95 ± 0.05 b | 0.92 ± 0.04 b |
| Topological Properties | Bacteria | Fungi | ||||
|---|---|---|---|---|---|---|
| CK | SD | LD | CK | SD | LD | |
| Positive ratio | 88.76% | 78.46% | 68.27% | 92.67% | 77.08% | 89.17% |
| Negative ratio | 11.24% | 21.54% | 31.13% | 7.33% | 22.92% | 10.83% |
| Nodes | 141 | 159 | 140 | 142 | 127 | 125 |
| Edges | 356 | 1639 | 312 | 232 | 253 | 240 |
| Average degree | 5.05 | 20.62 | 4.46 | 3.27 | 3.99 | 3.84 |
| Graph density | 0.036 | 0.13 | 0.032 | 0.023 | 0.032 | 0.031 |
| Average path length | 4.35 | 3.07 | 4.09 | 5.43 | 4.27 | 4.69 |
| Soil Property | Bacteria | Fungi | ||
|---|---|---|---|---|
| Richness | Shannon Index | Richness | Shannon Index | |
| clay | 0.58 ** | 0.51 ** | −0.44 ** | −0.61 ** |
| silt | −0.11 | −0.11 | 0.14 | 0.12 |
| sand | −0.09 | 0.06 | 0.01 | 0.04 |
| SWC | −0.66 ** | −0.59 ** | 0.49 ** | 0.67 ** |
| pH | 0.70 ** | 0.68 ** | −0.49 ** | −0.49 ** |
| SOC | −0.69 ** | −0.62 ** | 0.46 ** | 0.65 ** |
| TN | −0. 66 ** | −0.58 ** | 0.46 ** | 0.65 ** |
| TP | −0.54 ** | −0.42 ** | 0.54 ** | 0.70 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Shi, Y.; Jin, X.; Li, Z.; Wang, W.; Huang, S.; Jin, H. Influences of the China–Russia Crude Oil Pipelines on the Characteristics of Soil Bacterial and Fungal Communities in Permafrost Regions of the Da Xing’anling Mountains, Northeast China. Forests 2025, 16, 1038. https://doi.org/10.3390/f16071038
Yang X, Shi Y, Jin X, Li Z, Wang W, Huang S, Jin H. Influences of the China–Russia Crude Oil Pipelines on the Characteristics of Soil Bacterial and Fungal Communities in Permafrost Regions of the Da Xing’anling Mountains, Northeast China. Forests. 2025; 16(7):1038. https://doi.org/10.3390/f16071038
Chicago/Turabian StyleYang, Xue, Yanling Shi, Xiaoying Jin, Zuwang Li, Wenhui Wang, Shuai Huang, and Huijun Jin. 2025. "Influences of the China–Russia Crude Oil Pipelines on the Characteristics of Soil Bacterial and Fungal Communities in Permafrost Regions of the Da Xing’anling Mountains, Northeast China" Forests 16, no. 7: 1038. https://doi.org/10.3390/f16071038
APA StyleYang, X., Shi, Y., Jin, X., Li, Z., Wang, W., Huang, S., & Jin, H. (2025). Influences of the China–Russia Crude Oil Pipelines on the Characteristics of Soil Bacterial and Fungal Communities in Permafrost Regions of the Da Xing’anling Mountains, Northeast China. Forests, 16(7), 1038. https://doi.org/10.3390/f16071038