
Growth Responses of European Beech (Fagus sylvatica L.) and Oriental Beech (Fagus orientalis Lipsky) Along an Elevation Gradient Under Global Climate Change

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Material and Methods
2.1. Study Site
2.2. Data Collection
2.3. Data Analyses
3. Results
3.1. Production and Structure
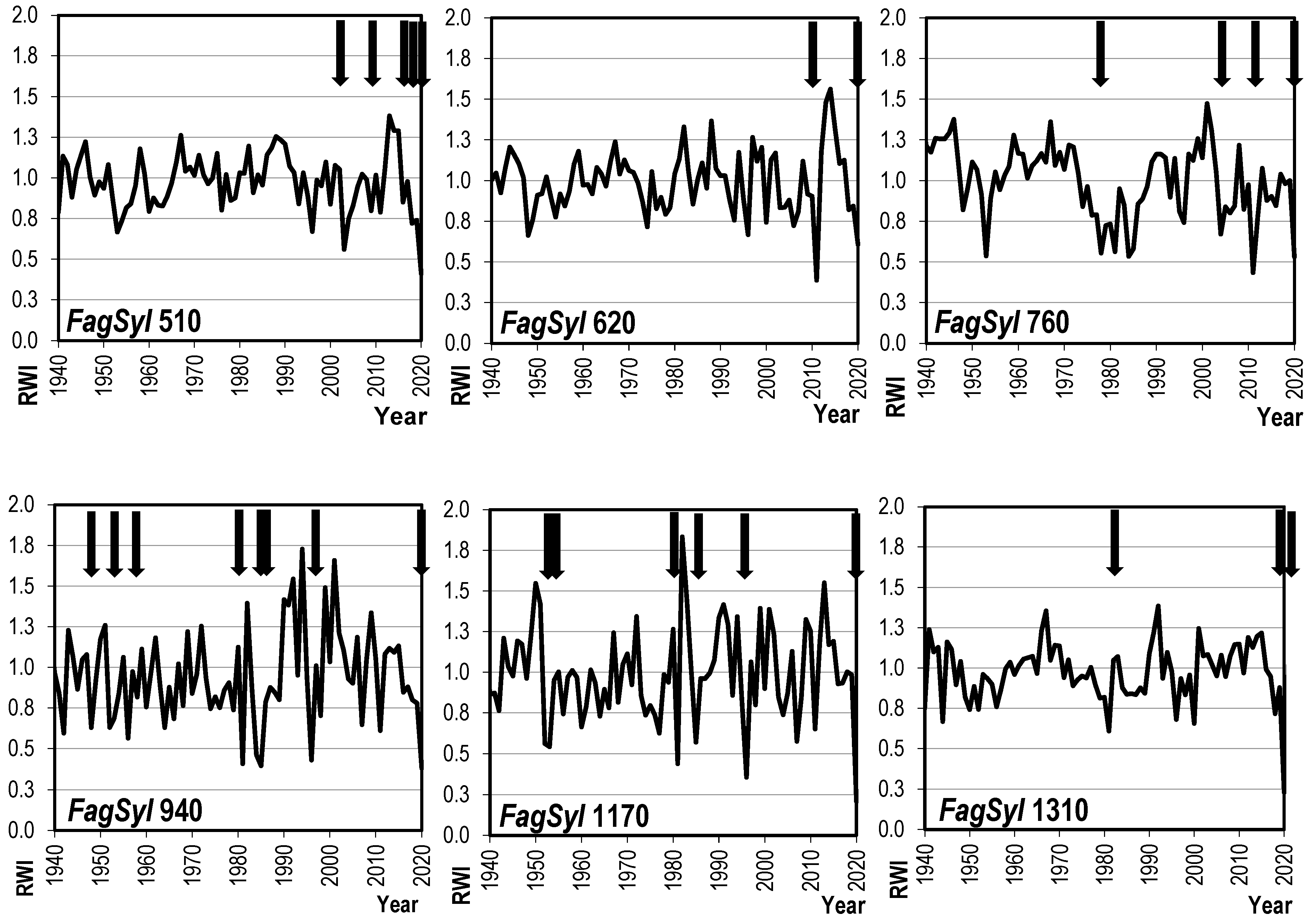
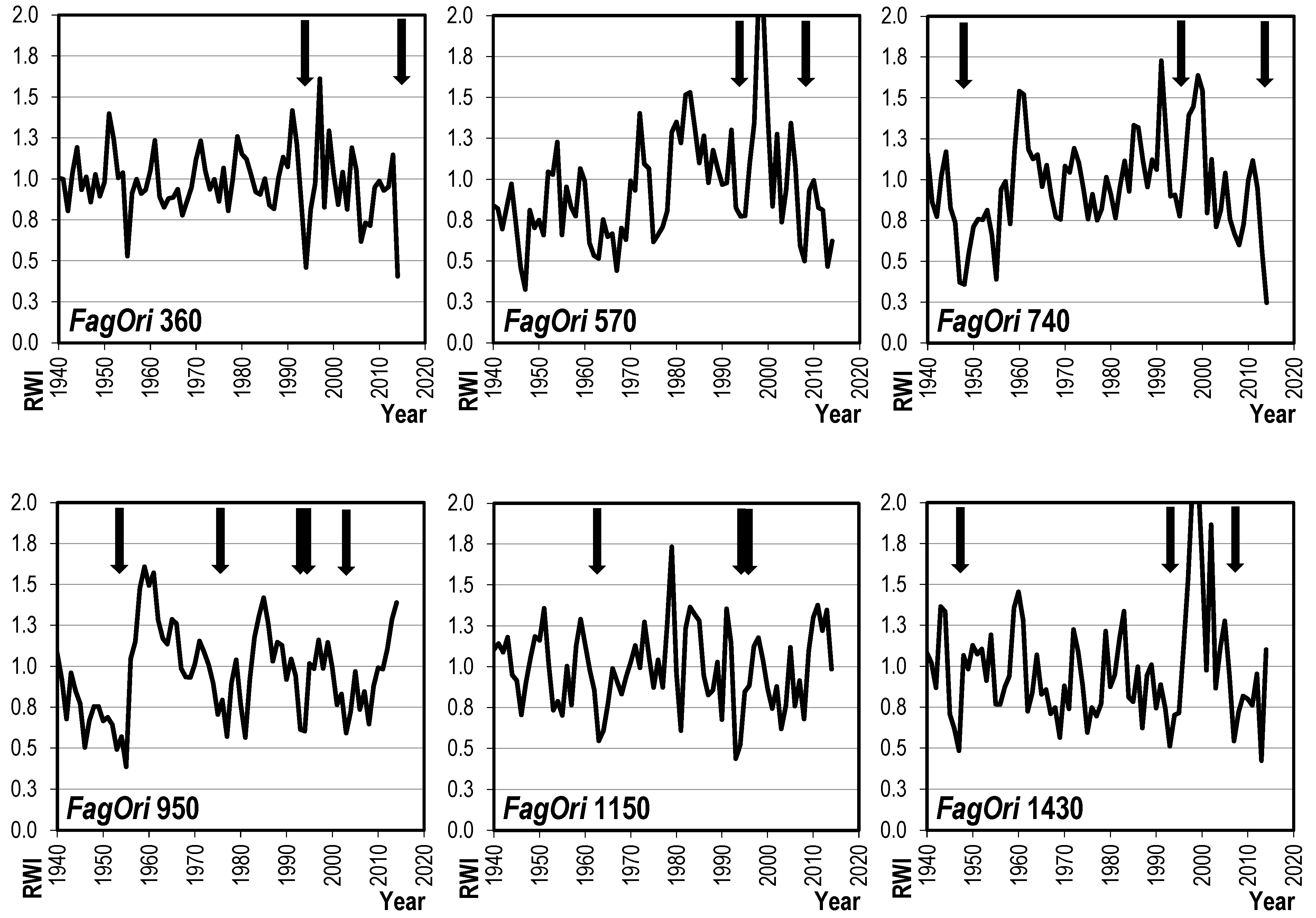
3.2. Dynamics of Radial Growth
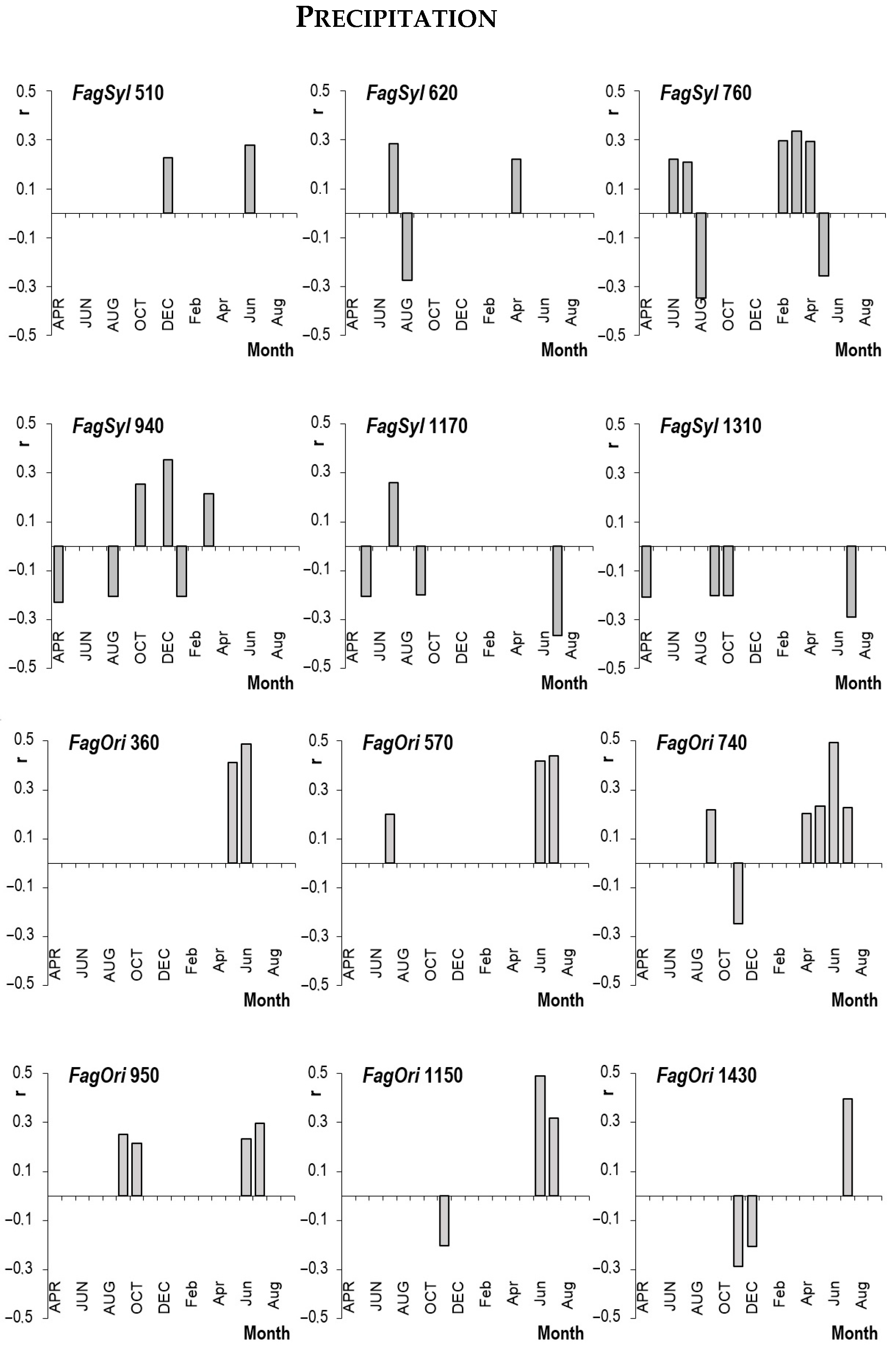
3.3. Effect of Climate Factors
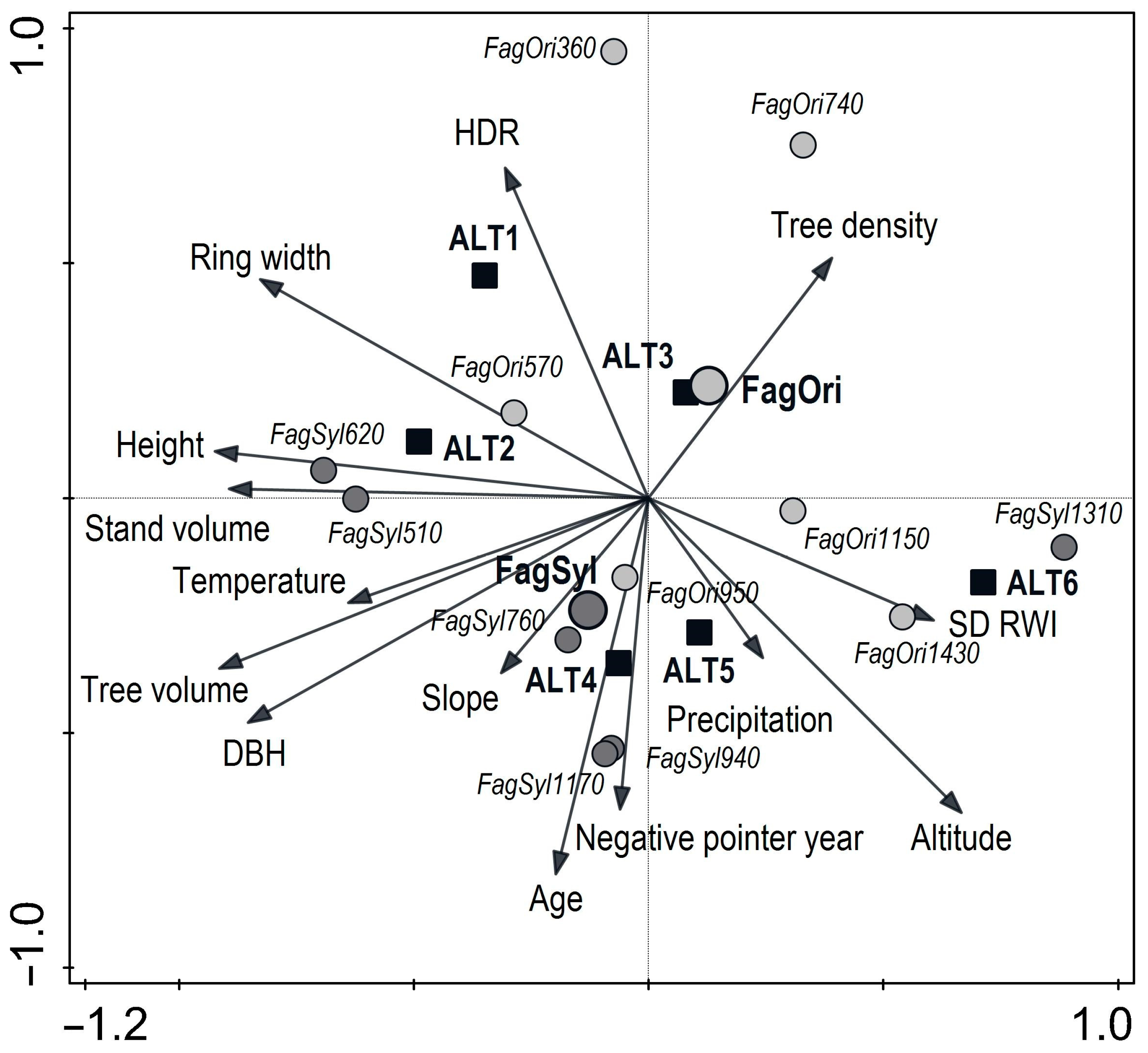
3.4. Relationships Between Radial Growth, Climate, Site, and Stand Parameters
4. Discussion
4.1. Productivity Potential and Stand Structure
4.2. Radial Growth Trends and Variability
4.3. Impact of Climatic Factors on Growth
4.4. Study Limitations and Recommendations
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berndes, G.; Abt, B.; Asikainen, A.; Cowie, A.; Dale, V.; Egnell, G.; Lindner, M.; Marelli, L.; Paré, D.; Pingoud, K. Forest Biomass, Carbon Neutrality and Climate Change Mitigation. Sci. Policy 2016, 3, 1–27. [Google Scholar]
- Harris, N.L.; Gibbs, D.A.; Baccini, A.; Birdsey, R.A.; de Bruin, S.; Farina, M.; Fatoyinbo, L.; Hansen, M.C.; Herold, M.; Houghton, R.A.; et al. Global Maps of Twenty-First Century Forest Carbon Fluxes. Nat. Clim. Change 2021, 11, 234–240. [Google Scholar] [CrossRef]
- Lindner, M.; Fitzgerald, J.B.; Zimmermann, N.E.; Reyer, C.; Delzon, S.; van der Maaten, E.; Schelhaas, M.J.; Lasch, P.; Eggers, J.; van der Maaten-Theunissen, M.; et al. Climate Change and European Forests: What Do We Know, What Are the Uncertainties, and What Are the Implications for Forest Management? J. Environ. Manag. 2014, 146, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest Disturbances under Climate Change. Nat. Clim. Change 2017, 7, 395–402. [Google Scholar] [CrossRef]
- Pureswaran, D.S.; Roques, A.; Battisti, A. Forest Insects and Climate Change. Curr. For. Rep. 2018, 4, 35–50. [Google Scholar] [CrossRef]
- Vacek, Z.; Vacek, S.; Cukor, J. European Forests under Global Climate Change: Review of Tree Growth Processes, Crises and Management Strategies. J. Environ. Manag. 2023, 332, 117353. [Google Scholar] [CrossRef]
- Kramer, K.; Degen, B.; Buschbom, J.; Hickler, T.; Thuiller, W.; Sykes, M.T.; de Winter, W. Modelling Exploration of the Future of European Beech (Fagus sylvatica L.) under Climate Change-Range, Abundance, Genetic Diversity and Adaptive Response. Ecol. Manag. 2010, 259, 2213–2222. [Google Scholar] [CrossRef]
- Mellert, K.H.; Šeho, M. Suitability of Fagus orientalis Lipsky at Marginal Fagus sylvatica L. Forest Sites in Southern Germany. IForest 2022, 15, 417–423. [Google Scholar] [CrossRef]
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas mit den Alpen, 6th ed.; Eugen Ulmer Verlag: Stuttgart, Germany, 2010; 1357p. [Google Scholar]
- Serin, H.; Akay, A.E.; Pak, M. Estimating the Effects of Optimum Bucking on the Economic Value of Brutian Pine (Pinus brutia) Logs Extracted in Mediterranean Region of Turkey. Afr. J. Agric. Res. 2010, 5, 916–921. [Google Scholar]
- Shahverdi, M.; Dashti, H.; Taghiyari, H.R.; Heshmati, S.; Gholamiyan, H.; Hossein, M. The Impact of Red Heartwood on Drying Characteristics and Mass Transfer Coefficients in Beech Wood. Austrian J. For. Sci. 2012, 130, 85–101. [Google Scholar]
- Pak, M.; Gulci, N. A Comparative Economic Evaluation of Bucking Deciduous Trees: A Case Study of Oriental Beech (Fagus orientalis) Stands in Northeastern Turkey. J. Fac. For.-Istanb. Univ. 2017, 67, 72–79. [Google Scholar]
- Drobyshev, I.; Niklasson, M.; Mazerolle, M.J.; Bergeron, Y. Reconstruction of a 253-Year Long Mast Record of European Beech Reveals Its Association with Large Scale Temperature Variability and No Long-Term Trend in Mast Frequencies. Agric. Meteorol. 2014, 192, 9–17. [Google Scholar] [CrossRef]
- Hlásny, T.; Mátyás, C.; Seidl, R.; Kulla, L.; Merganičová, K.; Trombik, J.; Dobor, L.; Barcza, Z.; Konôpka, B. Climate Change Increases the Drought Risk in Central European Forests: What Are the Options for Adaptation? For. J. 2014, 60, 5–18. [Google Scholar] [CrossRef]
- Ertekin, M.; Kırdar, E.; Ayan, S. The Effects of Exposure, Elevation and Tree Age on Seed Characteristics of Fagus orientalis Lipsky. South-East Eur. For. 2015, 6, 15–23. [Google Scholar] [CrossRef]
- Yildiz, O.; Eşen, D. Effects of Different Rhododendron Control Methods in Eastern Beech (Fagus orientalis Lipsky) Ecosystems in the Western Black Sea Region of Turkey. Ann. Appl. Biol. 2006, 149, 235–242. [Google Scholar] [CrossRef]
- Peterken, G.F. Natural Woodland: Ecology and Conservation in Northern Temperate Regions; Cambridge University Press: Cambridge, UK, 1996; ISBN 0521367921. [Google Scholar]
- Barna, M.; Bosela, M. Tree Species Diversity Change in Natural Regeneration of a Beech Forest under Different Management. Ecol. Manag. 2015, 342, 93–102. [Google Scholar] [CrossRef]
- Fuchs, Z.; Vacek, Z.; Vacek, S.; Cukor, J.; Šimůnek, V.; Štefančík, I.; Brabec, P.; Králíček, I. European Beech (Fagus sylvatica L.): A Promising Candidate for Future Forest Ecosystems in Central Europe amid Climate Change. Cent. Eur. For. J. 2024, 70, 62–76. [Google Scholar] [CrossRef]
- Nabuurs, G.J.; Pussinen, A.; van Brusselen, J.; Schelhaas, M.J. Future Harvesting Pressure on European Forests. Eur. J. Res. 2007, 126, 391–400. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.J.; Nabuurs, G.J.; Zimmermann, N.E. Climate Change May Cause Severe Loss in the Economic Value of European Forest Land. Nat. Clim. Change 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Sefidi, K.; Esfandiary Darabad, F.; Azaryan, M. Effect of Topography on Tree Species Composition and Volume of Coarse Woody Debris in an Oriental Beech (Fagus orientalis Lipsky) Old Growth Forests, Northern Iran. IForest 2016, 9, 658–665. [Google Scholar] [CrossRef]
- Ammer, C. Diversity and Forest Productivity in a Changing Climate. New Phytol. 2019, 221, 50–66. [Google Scholar] [CrossRef] [PubMed]
- Mäkinen, H.; Nöjd, P.; Kahle, H.P.; Neumann, U.; Tveite, B.; Mielikäinen, K.; Röhle, H.; Spiecker, H. Radial Growth Variation of Norway Spruce (Picea abies (L.) Karst.) across Latitudinal and Altitudinal Gradients in Central and Northern Europe. Ecol. Manag. 2002, 171, 243–259. [Google Scholar] [CrossRef]
- Lyu, L.; Suvanto, S.; Nöjd, P.; Henttonen, H.M.; Mäkinen, H.; Zhang, Q.-B. Tree Growth and Its Climate Signal along Latitudinal and Altitudinal Gradients: Comparison of Tree Rings between Finland and the Tibetan Plateau. Biogeosciences 2017, 14, 3083–3095. [Google Scholar] [CrossRef]
- Vacek, Z.; Vacek, S.; Cukor, J.; Bulušek, D.; Slávik, M.; Lukáčik, I.; Štefančík, I.; Sitková, Z.; Eşen, D.; Ripullone, F.; et al. Dendrochronological Data from Twelve Countries Proved Definite Growth Response of Black Alder (Alnus glutinosa [L.] Gaertn.) to Climate Courses across Its Distribution Range. Cent. Eur. For. J. 2022, 68, 139–153. [Google Scholar] [CrossRef]
- Mátyás, C.; Berki, I.; Czúcz, B.; Gálos, B.; Móricz, N.; Rasztovits, E. Future of Beech in Southeast Europe from the Perspective of Evolutionary Ecology. Acta Silv. Lignaria Hung. 2010, 6, 91–110. [Google Scholar] [CrossRef]
- Dulamsuren, C.; Hauck, M.; Kopp, G.; Ruff, M.; Leuschner, C. European Beech Responds to Climate Change with Growth Decline at Lower, and Growth Increase at Higher Elevations in the Center of Its Distribution Range (SW Germany). Trees—Struct. Funct. 2017, 31, 673–686. [Google Scholar] [CrossRef]
- Ningre, F.; Colin, F. Frost Damage on the Terminal Shoot as a Risk Factor of Fork Incidence on Common Beech (Fagus sylvatica L.). Ann. Sci. 2007, 64, 79–86. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Colangelo, M.; de Luis, M.; Martínez del Castillo, E.; Serra-Maluquer, X. Summer Drought and Spring Frost, but Not Their Interaction, Constrain European Beech and Silver Fir Growth in Their Southern Distribution Limits. Agric. Meteorol. 2019, 278, 107695. [Google Scholar] [CrossRef]
- Von Oheimb, G.; Westphal, C.; Tempel, H.; Härdtle, W. Structural Pattern of a Near-Natural Beech Forest (Fagus sylvatica) (Serrahn, North-East Germany). Ecol. Manag. 2005, 212, 253–263. [Google Scholar] [CrossRef]
- Janík, D.; Král, K.; Adam, D.; Hort, L.; Samonil, P.; Unar, P.; Vrska, T.; McMahon, S. Tree Spatial Patterns of Fagus sylvatica Expansion over 37 Years. Ecol. Manag. 2016, 375, 134–145. [Google Scholar] [CrossRef]
- Bilela, S.; Dounavi, A.; Fussi, B.; Konnert, M.; Holst, J.; Mayer, H.; Rennenberg, H.; Simon, J. Natural Regeneration of Fagus sylvatica L. Adapts with Maturation to Warmer and Drier Microclimatic Conditions. Ecol. Manag. 2012, 275, 60–67. [Google Scholar] [CrossRef]
- Rennenberg, H.; Seiler, W.; Matyssek, R.; Gessler, A.; Kreuzwieser, J. European Beech (Fagus sylvatica L.)—A Forest Tree Without Future in the South of Central Europe? Allg. Forst Jagdztg. 2004, 175, 210–224. [Google Scholar]
- Baumbach, L.; Niamir, A.; Hickler, T.; Yousefpour, R. Regional Adaptation of European Beech (Fagus sylvatica) to Drought in Central European Conditions Considering Environmental Suitability and Economic Implications. Reg. Environ. Change 2019, 19, 1159–1174. [Google Scholar] [CrossRef]
- Langer, G.J.; Bußkamp, J. Vitality Loss of Beech: A Serious Threat to Fagus sylvatica in Germany in the Context of Global Warming. J. Plant Dis. Prot. 2023, 130, 1101–1115. [Google Scholar] [CrossRef]
- Geßler, A.; Keitel, C.; Kreuzwieser, J.; Matyssek, R.; Seiler, W.; Rennenberg, H. Potential Risks for European Beech (Fagus sylvatica L.) in a Changing Climate. Trees—Struct. Funct. 2007, 21, 1–11. [Google Scholar] [CrossRef]
- Vacek, S.; Hejcman, M. Natural Layering, Foliation, Fertility and Plant Species Composition of a Fagus sylvatica Stand above the Alpine Timberline in the Giant (Krkonoše) Mts., Czech Republic. Eur. J. Res. 2012, 131, 799–810. [Google Scholar] [CrossRef]
- Dittmar, C.; Zech, W.; Elling, W. Growth Variations of Common Beech (Fagus sylvatica L.) under Different Climatic and Environmental Conditions in Europe—A Dendroecological Study. Ecol. Manag. 2003, 173, 63–78. [Google Scholar] [CrossRef]
- Allevato, E.; Saulino, L.; Cesarano, G.; Chirico, G.B.; D’Urso, G.; Bolognesi, S.F.; Rita, A.; Rossi, S.; Saracino, A.; Bonanomi, G. Canopy Damage by Spring Frost in European Beech Along the Apennines: Effect of Latitude, Altitude and Aspect. Remote Sens. Environ. 2019, 225, 431–440. [Google Scholar] [CrossRef]
- Dorren, L.K.A.; Berger, F.; Le Hir, C.; Mermin, E.; Tardif, P. Mechanisms, Effects and Management Implications of Rockfall in Forests. Ecol. Manag. 2005, 215, 183–195. [Google Scholar] [CrossRef]
- Johnson, O. Collins Tree Guide; HarperCollins Publishers: New York, NY, USA, 2004; ISBN 0007139543. [Google Scholar]
- Augusto, L.; Ranger, J.; Binkley, D.; Rothe, A. Impact of Several Common Tree Species of European Temperate Forests on Soil Fertility. Ann. Sci. 2002, 59, 233–253. [Google Scholar] [CrossRef]
- Houston Durrant, T.; de Rigo, D.; Caudullo, G. Fagus sylvatica and Other Beeches in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; European Commission: Luxembourg, 2016; pp. 94–95. [Google Scholar]
- Vacek, Z.; Vacek, S.; Slanař, J.; Bílek, L.; Bulušek, D.; Štefančík, I.; Králíček, I.; Vančura, K. Adaption of Norway Spruce and European Beech Forests under Climate Change: From Resistance to Close-to-Nature Silviculture. Cent. Eur. For. J. 2019, 65, 129–144. [Google Scholar] [CrossRef]
- Aertsen, W.; Janssen, E.; Kint, V.; Bontemps, J.-D.; Van Orshoven, J.; Muys, B. Long-Term Growth Changes of Common Beech (Fagus sylvatica L.) Are Less Pronounced on Highly Productive Sites. Ecol. Manag. 2014, 312, 252–259. [Google Scholar] [CrossRef]
- Králíček, I.; Vacek, Z.; Vacek, S.; Remeš, J.; Bulušek, D.; Král, J.; Štefančík, I.; Putalová, T. Dynamics and Structure of Mountain Autochthonous Spruce-Beech Forests: Impact of Hilltop Phenomenon, Air Pollutants and Climate. Dendrobiology 2017, 77, 119–137. [Google Scholar] [CrossRef]
- Šimůnek, V.; Vacek, Z.; Vacek, S.; Králíček, I.; Vančura, K. Growth Variability of European Beech (Fagus sylvatica L.) Natural Forests: Dendroclimatic Study from Krkonoše National Park. Cent. Eur. For. J. 2019, 65, 92–102. [Google Scholar] [CrossRef]
- Di Filippo, A.; Biondi, F.; Čufar, K.; De Luis, M.; Grabner, M.; Maugeri, M.; Presutti Saba, E.; Schirone, B.; Piovesan, G. Bioclimatology of Beech (Fagus sylvatica L.) in the Eastern Alps: Spatial and Altitudinal Climatic Signals Identified through a Tree-ring Network. J. Biogeogr. 2007, 34, 1873–1892. [Google Scholar] [CrossRef]
- Di Filippo, A.; Biondi, F.; Piovesan, G.; Ziaco, E. Tree Ring-based Metrics for Assessing Old-growth Forest Naturalness. J. Appl. Ecol. 2017, 54, 737–749. [Google Scholar] [CrossRef]
- Tegel, W.; Seim, A.; Hakelberg, D.; Hoffmann, S.; Panev, M.; Westphal, T.; Büntgen, U. A Recent Growth Increase of European Beech (Fagus sylvatica L.) at Its Mediterranean Distribution Limit Contradicts Drought Stress. Eur. J. Res. 2014, 133, 61–71. [Google Scholar] [CrossRef]
- Tognetti, R.; Lasserre, B.; Di Febbraro, M.; Marchetti, M. Modeling Regional Drought-Stress Indices for Beech Forests in Mediterranean Mountains Based on Tree-Ring Data. Agric. Meteorol. 2019, 265, 110–120. [Google Scholar] [CrossRef]
- Leuschner, C. Perspectives in Plant Ecology, Evolution and Systematics Drought Response of European Beech (Fagus sylvatica L.)—A Review. Perspect. Plant Ecol. Evol. Syst. 2020, 47, 125576. [Google Scholar] [CrossRef]
- Stjepanović, S.; Matović, B.; Stojanović, D.; Lalić, B.; Levanič, T.; Orlović, S.; Gutalj, M. The Impact of Adverse Weather and Climate on the Width of European Beech (Fagus sylvatica L.) Tree Rings in Southeastern Europe. Atmosphere 2018, 9, 451. [Google Scholar] [CrossRef]
- Haghshenas, M.; Mohadjer, M.R.M.; Attarod, P.; Pourtahmasi, K.; Feldhaus, J.; Sadeghi, S.M.M. Climate Effect on Tree-Ring Widths of Fagus Orientalis in the Caspian Forests, Northern Iran. For. Sci. Technol. 2016, 12, 176–182. [Google Scholar] [CrossRef]
- Alavi, S.J.; Ahmadi, K.; Dormann, C.F.; Serra-Diaz, J.M.; Nouri, Z. Assessing the Dominant Height of Oriental Beech (Fagus orientalis L.) in Relation to Edaphic and Physiographic Variables in the Hyrcanian Forests of Iran. BASE 2020, 24, 262–273. [Google Scholar] [CrossRef]
- Komitov, B. The European Beech Annual Tree Ring Widths Time Series, Solar–Climatic Relationships and Solar Dynamo Regime Changes. Atmosphere 2021, 12, 829. [Google Scholar] [CrossRef]
- Šimůnek, V.; Sharma, R.P.; Vacek, Z.; Vacek, S.; Hůnová, I. Sunspot Area as Unexplored Trend inside Radial Growth of European Beech in Krkonoše Mountains: A Forest Science from Different Perspective. Eur. J. Res. 2020, 139, 999–1013. [Google Scholar] [CrossRef]
- EEA. Impacts of Europe’s Changing Climate; Report No. 2/2004; European Environment Agency: Copenhagen, Denmark, 2004; ISBN 9789291673728. [Google Scholar]
- Atalay, I.; Efe, R.; Öztürk, M. Ecology and Classification of Forests in Turkey. Procedia-Soc. Behav. Sci. 2014, 120, 788–805. [Google Scholar] [CrossRef]
- Machar, I.; Vlckova, V.; Bucek, A.; Vozenilek, V.; Salek, L.; Jerabkova, L. Modelling of Climate Conditions in Forest Vegetation Zones as a Support Tool for Forest Management Strategy in European Beech Dominated Forests. Forests 2017, 8, 82. [Google Scholar] [CrossRef]
- Strengbom, J.; Dahlberg, A.; Larsson, A.; Lindelöw, Å.; Sandström, J.; Widenfalk, O.; Gustafsson, L. Introducing Intensively Managed Spruce Plantations in Swedish: Forest Landscapes Will Impair Biodiversity Decline. Forests 2011, 2, 610–630. [Google Scholar] [CrossRef]
- Seidl, R.; Rammer, W.; Lexer, M.J. Climate Change Vulnerability of Sustainable Forest Management in the Eastern Alps. Clim. Change 2011, 106, 225–254. [Google Scholar] [CrossRef]
- Di Filippo, A.; Biondi, F.; Maugeri, M.; Schirone, B.; Piovesan, G. Bioclimate and Growth History Affect Beech Lifespan in the Italian Alps and Apennines. Glob. Change Biol. 2012, 18, 960–972. [Google Scholar] [CrossRef]
- Temperli, C.; Bugmann, H.; Elkin, C. Adaptive Management for Competing Forest Goods and Services under Climate Change. Ecol. Appl. 2012, 22, 2065–2077. [Google Scholar] [CrossRef]
- Pötzelsberger, E.; Wolfslehner, B.; Hasenauer, H. Climate Change Impacts on Key Forest Functions of the Vienna Woods. Eur. J. Res. 2015, 134, 481–496. [Google Scholar] [CrossRef]
- Bulušek, D.; Vacek, Z.; Vacek, S.; Král, J.; Bílek, L.; Králíček, I. Spatial Pattern of Relict Beech (Fagus sylvatica L.) Forests in the Sudetes of the Czech Republic and Poland. J. For. Sci. 2016, 62, 293–305. [Google Scholar] [CrossRef]
- Vacek, Z.; Prokůpková, A.; Vacek, S.; Bulušek, D.; Šimůnek, V.; Hájek, V.; Králíček, I. Mixed vs. Monospecific Mountain Forests in Response to Climate Change: Structural and Growth Perspectives of Norway Spruce and European Beech. Ecol. Manag. 2021, 488, 119019. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated World Map of the Köppen-Geiger Climate Classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Vacek, Z.; Vacek, S.; Esen, D.; Yildiz, O.; Král, J.; Gallo, J. Effect of Invasive Rhododendron ponticum L. on Natural Regeneration and Structure of Fagus orientalis Lipsky Forests in the Black Sea Region. Forests 2020, 11, 603. [Google Scholar] [CrossRef]
- Kantarcı, M.D. Toprak İlmi [Soil Science]; Istanbul Universitesi Yayin: Istanbul, Turkey, 2000. [Google Scholar]
- Matějka, K.; Vacek, S.; Podrázský, V. Development of Forest Soils in the Krkonoše Mts. in the Period 1980–2009. J. Sci. 2010, 56, 485–504. [Google Scholar] [CrossRef]
- Kraft, G. Beiträge zur Lehre von Den Durchforstungen, Schlagstellungen und Lichtungshieben; Klindworth: Hannover, Germany, 1884. [Google Scholar]
- Haneca, K.; Čufar, K.; Beeckman, H. Oaks, Tree-Rings and Wooden Cultural Heritage: A Review of the Main Characteristics and Applications of Oak Dendrochronology in Europe. J. Archaeol. Sci. 2009, 36, 1–11. [Google Scholar] [CrossRef]
- Fabrika, M.; Ďurský, J. Algorithms and Software Solution of Thinning Models for SIBYLA Growth Simulator. J. Sci. 2005, 51, 431–445. [Google Scholar] [CrossRef]
- Petráš, R.; Pajtík, J. Sustava Cesko-Slovenskych Objemovych Tabuliek Drevin. Lesn. Cas. 1991, 37, 49–56. [Google Scholar]
- Carus, S. Increment and Growth in Even Aged Beech (Fagus orientalis Lipsky.) Forests. Postgraduate Thesis, The Graduate School of Natural and Applied Sciences, University of Istanbul, Istanbul, Turkey, 1998. [Google Scholar]
- Crookston, N.L. Percent Canopy Cover and Stand Structure Statistics from the Forest Vegetation Simulator; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 1999. [Google Scholar]
- Reineke, L.H. Perfecting a Stand-Density Index for Even-Aged Forest. J. Agric. Res. 1933, 46, 627–638. [Google Scholar]
- Halaj, J.; Grék, J.; Pánek, F.; Petráš, R.; Řehák, J. Rastové Tabuľky Hlavných Drevín ČSSR; Príroda Bratislava: Bratislava, Slovakia, 1987; 361p. [Google Scholar]
- Sharma, R.P.; Vacek, Z.; Vacek, S. Modeling Individual Tree Height to Diameter Ratio for Norway Spruce and European Beech in Czech Republic. Trees—Struct. Funct. 2016, 30, 1969–1982. [Google Scholar] [CrossRef]
- Petráš, R.; Kosut, M.; Oszlányi, J. Listová Biomasa Stromov Smreka, Borovice a Buka. Lesn. Cas. 1985, 31, 121–135. [Google Scholar]
- Ledermann, T.; Neumann, M. Biomass Equations from Data of Old Long-Term Experimental Plots. Austrian J. For. Sci. 2006, 123, 47–64. [Google Scholar]
- Seifert, T.; Schuck, J.; Block, J.; Pretzsch, H. Simulation von Biomasse-Und Nährstoffgehalt von Waldbäumen. Beiträge Jahrestag. Vom 2006, 29, 31. [Google Scholar]
- Speight, J.G. Properties of Fuels from Wood Sources; Royal Society of Chemistry: Cambridge, UK, 2011. [Google Scholar] [CrossRef]
- Drexhage, M.; Colin, F. Estimating Root System Biomass from Breast-height Diameters. Forestry 2001, 74, 491–497. [Google Scholar] [CrossRef]
- Bublinec, E. Koncentrácia, Akumulácia a Kolobeh Prvkov v Bukovom a Smrekovom Ekosystéme; Veda: Bratislava, Slovakia, 1994; ISBN 8022401277. [Google Scholar]
- Bunn, A.G. Statistical and Visual Crossdating in R Using the DplR Library. Dendrochronologia 2010, 28, 251–258. [Google Scholar] [CrossRef]
- Cook, E.R.; Shiyatov, S.G.; Mazepa, V.S.; Ecology, A.; Branch, U. Methods of Dendrochronology Applications. In Tree-Ring Laboratory, Lamont-Ooherty Geological Observatory; Cook, E.R., Kairiukstis, L.A., Eds.; Columbia University: New York, NY, USA, 1990; Volume 10, pp. 978–994. [Google Scholar]
- Bunn, A.; Korpela, M.; Biondi, F.; Campelo, F.; Mérian, P.; Qeadan, F.; Zang, C. DplR: Dendrochronology Program Library in R; R Package Version 2020; R Foundation: Vienna, Austria, 2020; Volume 1. [Google Scholar]
- Desplanque, C.; Rolland, C.; Schweingruber, F.H. Influence of Species and Abiotic Factors on Extreme Tree Ring Modulation: Picea abies and Abies alba in Tarentaise and Maurienne (French Alps). Trees—Struct. Funct. 1999, 13, 218–227. [Google Scholar] [CrossRef]
- Biondi, F.; Waikul, K. DENDROCLIM2002: A C++ Program for Statistical Calibration of Climate Signals in Tree-Ring Chronologies. Comput. Geosci. 2004, 30, 303–311. [Google Scholar] [CrossRef]
- Špulák, O.; Souček, J. The Sibyla Model and Development of Beech Forests Affected by Air Pollution. Cent. Eur. J. Biol. 2010, 5, 371–383. [Google Scholar] [CrossRef]
- Slanař, J.; Vacek, Z.; Vacek, S.; Bulušek, D.; Cukor, J.; Štefančík, I.; Bílek, L.; Krl, J. Long-Term Transformation of Submontane Spruce-Beech Forests in the Jizerské Hory Mts.: Dynamics of Natural Regeneration. Cent. Eur. For. J. 2017, 63, 212–224. [Google Scholar] [CrossRef]
- Bilek, L.; Remes, J.; Zahradnik, D. Managed versus Unmanaged. Structure of Beech Forest Stands (Fagus sylvatica L.) after 50 Years of Development, Central Bohemian. For. Syst. 2011, 20, 122–138. [Google Scholar] [CrossRef]
- Fuchs, Z.; Vacek, Z.; Vacek, S.; Gallo, J. Effect of Game Browsing on Natural Regeneration of European Beech (Fagus sylvatica L.) Forests in the Krušné Hory Mts. (Czech Republic and Germany). Cent. Eur. For. J. 2021, 67, 166–180. [Google Scholar] [CrossRef]
- Trotsiuk, V.; Hobi, M.L.; Commarmot, B. Age Structure and Disturbance Dynamics of the Relic Virgin Beech Forest Uholka (Ukrainian Carpathians). Ecol. Manag. 2012, 265, 181–190. [Google Scholar] [CrossRef]
- Atici, E.; Colak, A.H. The Structure and Volume Parameters of Uneven-Aged Oriental Beech (Fagus orientalis Lipsky) Forests in Turkey. Arboric. J. 2008, 31, 137–160. [Google Scholar] [CrossRef]
- Amiri, M.; Rahmani, R.; Sagheb-Talebi, K.; Habashi, H. Dynamics and Structural Characteristics of a Natural Unlogged Oriental Beech (Fagus orientalis Lipsky) Stand during a 5-Year Period in Shast Kalate Forest, Northern Iran. Int. J. Environ. Resour. Res. 2013, 1, 107–129. [Google Scholar]
- Talebi-Sagheb, K.; Schütz, J.-P. The Structure of Natural Oriental Beech (Fagus orientalis) Forests in the Caspian Region of Iran and Potential for the Application of the Group Selection System. Forestry 2002, 75, 465–472. [Google Scholar] [CrossRef]
- Özçelík, R.; Brooks, J.R.; Diamantopoulou, M.J.; Wiant, H.V. Estimating Breast Height Diameter and Volume from Stump Diameter for Three Economically Important Species in Turkey. Scand. J. Res. 2010, 25, 32–45. [Google Scholar] [CrossRef]
- Valipour, A.; Namiranian, M.; Ghazanfari, H.; Heshmatol Vaezin, S.; Lexer, M.; Plieninger, T. Relationships between Forest Structure and Tree’s Dimensions with Physiographical Factors in Armardeh Forests (Northern Zagros). Iran. J. For. Poplar Res. 2013, 21, 30–47. [Google Scholar]
- Rugani, T.; Diaci, J.; Hladnik, D. Gap Dynamics and Structure of Two Old-Growth Beech Forest Remnants in Slovenia. PLoS ONE 2013, 8, e52641. [Google Scholar] [CrossRef]
- Moridi, M.; Sefidi, K.; Etemad, V. Stand Characteristics of Mixed Oriental Beech (Fagus orientalis Lipsky) Stands in the Stem Exclusion Phase, Northern Iran. Eur. J. Res. 2015, 134, 693–703. [Google Scholar] [CrossRef]
- Podrázský, V.; Baláš, M.; Linda, R.; Křivohlavý, O. State of Beech Pole Stands Established at the Clear-Cut and in the Underplanting. J. For. Sci. 2019, 65, 256–262. [Google Scholar] [CrossRef]
- Khalatbari Limaki, M.; Es-hagh Nimvari, M.; Alavi, S.J.; Mataji, A.; Kazemnezhad, F. Potential Elevation Shift of Oriental Beech (Fagus orientalis L.) in Hyrcanian Mixed Forest Ecoregion under Future Global Warming. Ecol. Model. 2021, 455, 109637. [Google Scholar] [CrossRef]
- Černý, J.; Špulák, O.; Sýkora, P.; Novosadová, K.; Kadlec, J.; Kománek, M. The Significance of European Beech in Central Europe in the Period of Climate Change: An Overview of Current Knowledge. Zprávy Lesn. Výzk 2024, 69, 74–88. [Google Scholar]
- Hrivnák, R.; Gömöry, D.; Slezák, M.; Ujházy, K.; Hédl, R.; Jarčuška, B.; Ujházyová, M. Species Richness Pattern along Altitudinal Gradient in Central European Beech Forests. Folia Geobot. 2014, 49, 425–441. [Google Scholar] [CrossRef]
- Rahbek, C. The Role of Spatial Scale and the Perception of Large-Scale Species-Richness Patterns. Ecol. Lett. 2005, 8, 224–239. [Google Scholar] [CrossRef]
- Larcher, W. Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups; Springer Science & Business Media: Berlin, Germany, 2003; ISBN 3540435166. [Google Scholar]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems; Springer: Berlin/Heidelberg, Germany, 1999. [Google Scholar]
- Sanders, N.J.; Moss, J.; Wagner, D. Patterns of Ant Species Richness along Elevational Gradients in an Arid Ecosystem. Glob. Ecol. Biogeogr. 2003, 12, 93–102. [Google Scholar] [CrossRef]
- Vannoppen, A.; Treydte, K.; Boeckx, P.; Kint, V.; Ponette, Q.; Verheyen, K.; Muys, B. Tree Species Diversity Improves Beech Growth and Alters Its Physiological Response to Drought. Trees—Struct. Funct. 2020, 34, 1059–1073. [Google Scholar] [CrossRef]
- Pretzsch, H.; Hilmers, T. Structural Diversity and Carbon Stock of Forest Stands: Tradeoff as Modified by Silvicultural Thinning. Eur. J. Res. 2024, 1–22. [Google Scholar] [CrossRef]
- Köse, N.; Güner, H.T. The Effect of Temperature and Precipitation on the Intra-Annual Radial Growth of Fagus orientalis Lipsky in Artvin, Turkey. Turk. J. Agric. For. 2012, 36, 501–509. [Google Scholar] [CrossRef]
- Oladi, R.; Pourtahmasi, K. Intra-Annual Secondary Growth Rate-Climate Relations of Fagus orientalis Lipsky in the Center of Hyrcanian Forests. Not. Sci. Biol. 2012, 4, 136–140. [Google Scholar] [CrossRef]
- Peñuelas, J.; Ogaya, R.; Boada, M.; Jump, A.S. Migration, Invasion and Decline: Changes in Recruitment and Forest Structure in a Warming-Linked Shift of European Beech Forest in Catalonia (NE Spain). Ecography 2007, 30, 829–837. [Google Scholar] [CrossRef]
- Dorado-Liñán, I.; Akhmetzyanov, L.; Menzel, A. Climate Threats on Growth of Rear-Edge European Beech Peripheral Populations in Spain. Int. J. Biometeorol. 2017, 61, 2097–2110. [Google Scholar] [CrossRef] [PubMed]
- Sedmáková, D.; Sedmák, R.; Bosela, M.; Ježík, M.; Blaženec, M.; Hlásny, T.; Marušák, R. Growth-Climate Responses Indicate Shifts in the Competitive Ability of European Beech and Norway Spruce under Recent Climate Warming in East-Central Europe. Dendrochronologia 2019, 54, 37–48. [Google Scholar] [CrossRef]
- Pavlović, L.; Stojanović, D.; Mladenović, E.; Lakićević, M.; Orlović, S. Potential Elevation Shift of the European Beech Stands (Fagus sylvatica L.) in Serbia. Front. Plant Sci. 2019, 10, 849. [Google Scholar] [CrossRef]
- Vannoppen, A.; Kint, V.; Ponette, Q.; Verheyen, K.; Muys, B. Tree Species Diversity Impacts Average Radial Growth of Beech and Oak Trees in Belgium, Not Their Long-Term Growth Trend. Ecosyst 2019, 6, 10. [Google Scholar] [CrossRef]
- Jump, A.S.; Hunt, J.M.; Peñuelas, J. Rapid Climate Change-Related Growth Decline at the Southern Range Edge of Fagus sylvatica. Glob. Change Biol. 2006, 12, 2163–2174. [Google Scholar] [CrossRef]
- Piovesan, G.; Biondi, F.; Di Filippo, A.; Alessandrini, A.; Maugeri, M. Drought-Driven Growth Reduction in Old Beech (Fagus sylvatica L.) Forests of the Central Apennines, Italy. Glob. Change Biol. 2008, 14, 1265–1281. [Google Scholar] [CrossRef]
- Kint, V.; Aertsen, W.; Campioli, M.; Vansteenkiste, D.; Delcloo, A.; Muys, B. Radial Growth Change of Temperate Tree Species in Response to Altered Regional Climate and Air Quality in the Period 1901–2008. Clim. Change 2012, 115, 343–363. [Google Scholar] [CrossRef]
- Charru, M.; Seynave, I.; Hervé, J.-C.; Bertrand, R.; Bontemps, J.-D. Recent Growth Changes in Western European Forests Are Driven by Climate Warming and Structured across Tree Species Climatic Habitats. Ann. Sci. 2017, 74, 33. [Google Scholar] [CrossRef]
- Young, D.J.N.; Stevens, J.T.; Earles, J.M.; Moore, J.; Ellis, A.; Jirka, A.L.; Latimer, A.M. Long-Term Climate and Competition Explain Forest Mortality Patterns under Extreme Drought. Ecol. Lett. 2017, 20, 78–86. [Google Scholar] [CrossRef]
- Whitman, E.; Parisien, M.A.; Thompson, D.K.; Flannigan, M.D. Short-Interval Wildfire and Drought Overwhelm Boreal Forest Resilience. Sci. Rep. 2019, 9, 18796. [Google Scholar] [CrossRef] [PubMed]
- Kolář, T.; Čermák, P.; Trnka, M.; Žid, T.; Rybníček, M. Temporal Changes in the Climate Sensitivity of Norway Spruce and European Beech along an Elevation Gradient in Central Europe. Agric. Meteorol. 2017, 239, 24–33. [Google Scholar] [CrossRef]
- Sefidi, K.; Pourgoli, Z.; Sagheb-Talebi, K.; Behjou, F.K. Stand Characteristics of Gap Formation Phase Through the Development of Oriental Beech (Fagus orientalis Lipsky) Stands in the Hyrcanian Forests, Northern Iran. Austrian J. For. Sci. 2018, 2, 137–158. [Google Scholar]
- Alavi, S.J.; Ahmadi, K.; Hosseini, S.M.; Tabari, M.; Nouri, Z. The Response of English Yew (Taxus baccata L.) to Climate Change in the Caspian Hyrcanian Mixed Forest Ecoregion. Reg. Environ. Change 2019, 19, 1495–1506. [Google Scholar] [CrossRef]
- Taleshi, H.; Jalali, S.G.; Alavi, S.J.; Hosseini, S.M.; Naimi, B.; Zimmermann, N.E. Climate Change Impacts on the Distribution and Diversity of Major Tree Species in the Temperate Forests of Northern Iran. Reg. Environ. Change 2019, 19, 2711–2728. [Google Scholar] [CrossRef]
- Yaman, B.; Köse, N.; Özel, H.B.; Şahan, E.A. The Effect of Climate on the Radial Growth of Oriental Beech in the Southern Limit of Its Distribution Area. Forestist 2020, 70, 53–59. [Google Scholar] [CrossRef]
- Akkemik, Ü.; Demir, D. Tree Ring Analysis on Eastern Beech (Fagus orientalis Lipsky.) in the Belgrade Forest. Forestist 2003, 53, 33–36. [Google Scholar]
- Martin-Benito, D.; Pederson, N.; Köse, N.; Doğan, M.; Bugmann, H.; Mosulishvili, M.; Bigler, C. Pervasive Effects of Drought on Tree Growth across a Wide Climatic Gradient in the Temperate Forests of the Caucasus. Glob. Ecol. Biogeogr. 2018, 27, 1314–1325. [Google Scholar] [CrossRef]
- Kehl, H. A Multi-Disciplinary Project on Causes and Diversity of Extrazonal Temperate Flora & Vegetation in the Amanos Mountains (SE-Turkey 1988-1997). Available online: https://www.science-e-publishing.de/project/loekat/e-loekat.htm (accessed on 16 December 2024).
- Skomarkova, M.V.; Vaganov, E.A.; Mund, M.; Knohl, A.; Linke, P.; Boerner, A.; Schulze, E.D. Inter-Annual and Seasonal Variability of Radial Growth, Wood Density and Carbon Isotope Ratios in Tree Rings of Beech (Fagus sylvatica) Growing in Germany and Italy. Trees—Struct. Funct. 2006, 20, 571–586. [Google Scholar] [CrossRef]
- Ježík, M.; Blaženec, M.; Kučera, J.; Střelcová, K.; Ditmarová, L. The Response of Intra-Annual Stem Circumference Increase of Young European Beech Provenances to 2012–2014 Weather Variability. IForest 2016, 9, 960–969. [Google Scholar] [CrossRef]
- Rohner, B.; Weber, P.; Thürig, E. Bridging Tree Rings and Forest Inventories: How Climate Effects on Spruce and Beech Growth Aggregate over Time. Ecol. Manag. 2016, 360, 159–169. [Google Scholar] [CrossRef]
- Fonti, P.; García-González, I. Suitability of Chestnut Earlywood Vessel Chronologies for Ecological Studies. New Phytol. 2004, 163, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Drobyshev, I.; Övergaard, R.; Saygin, I.; Niklasson, M.; Hickler, T.; Karlsson, M.; Sykes, M.T. Masting Behaviour and Dendrochronology of European Beech (Fagus sylvatica L.) in Southern Sweden. Ecol. Manag. 2010, 259, 2160–2171. [Google Scholar] [CrossRef]
- Foest, J.J.; Bogdziewicz, M.; Pesendorfer, M.B.; Ascoli, D.; Cutini, A.; Nussbaumer, A.; Verstraeten, A.; Beudert, B.; Chianucci, F.; Mezzavilla, F.; et al. Widespread Breakdown in Masting in European Beech Due to Rising Summer Temperatures. Glob. Change Biol. 2024, 30, e17307. [Google Scholar] [CrossRef]
- Dittmar, C.; Fricke, W.; Elling, W. Impact of Late Frost Events on Radial Growth of Common Beech (Fagus sylvatica L.) in Southern Germany. Eur. J. Res. 2006, 125, 249–259. [Google Scholar] [CrossRef]
- Latte, N.; Perin, J.; Kint, V.; Lebourgeois, F.; Claessens, H. Major Changes in Growth Rate and Growth Variability of Beech (Fagus sylvatica L.) Related to Soil Alteration and Climate Change in Belgium. Forests 2016, 7, 174. [Google Scholar] [CrossRef]
- González de Andrés, E.; Camarero, J.J.; Blanco, J.A.; Imbert, J.B.; Lo, Y.H.; Sangüesa-Barreda, G.; Castillo, F.J. Tree-to-Tree Competition in Mixed European Beech–Scots Pine Forests Has Different Impacts on Growth and Water-Use Efficiency Depending on Site Conditions. J. Ecol. 2018, 106, 59–75. [Google Scholar] [CrossRef]
- Pluess, A.R.; Weber, P. Drought-Adaptation Potential in Fagus sylvatica: Linking Moisture Availability with Genetic Diversity and Dendrochronology. PLoS ONE 2012, 7, e33636. [Google Scholar] [CrossRef]
- Martinez del Castillo, E.; Zang, C.S.; Buras, A.; Hacket-Pain, A.; Esper, J.; Serrano-Notivoli, R.; Hartl, C.; Weigel, R.; Klesse, S.; de Dios, V.R.; et al. Climate-Change-Driven Growth Decline of European Beech Forests. Commun. Biol. 2022, 5, 163. [Google Scholar] [CrossRef]
- Ruosteenoja, K.; Markkanen, T.; Venäläinen, A.; Räisänen, P.; Peltola, H. Seasonal Soil Moisture and Drought Occurrence in Europe in CMIP5 Projections for the 21st Century. Clim. Dyn. 2018, 50, 1177–1192. [Google Scholar] [CrossRef]
- Innes, J.L. Theoretical and Practical Criteria for the Selection of Ecosystem Monitoring Plots in Swiss Forests. Environ. Monit. Assess. 1995, 36, 271–294. [Google Scholar] [CrossRef] [PubMed]
- Carrer, M.; Castagneri, D.; Popa, I.; Pividori, M.; Lingua, E. Tree Spatial Patterns and Stand Attributes in Temperate Forests: The Importance of Plot Size, Sampling Design, and Null Model. Ecol. Manag. 2018, 407, 125–134. [Google Scholar] [CrossRef]
- Mousavisangdehi, A.; Oladi, R.; Pourtahmasi, K.; Etemad, V.; Koprowski, M.; Tumajer, J. Higher Temperatures Promote Intra-Annual Radial Growth of Oriental Beech (Fagus orientalis Lipsky) in the Humid Hyrcanian Forests. Trees—Struct. Funct. 2024, 38, 1569–1580. [Google Scholar] [CrossRef]
- Pourtahmasi, K.; Najafi Harsini, F.; Naderi, M.; Kousha, M.; Oladi, R.; Nadi, M. Comparative Investigation of the Radial Growth of Oriental Beech (Fagus orientalis Lipsky) in the Altitude Gradient of the HyrcanianForest, Iran. Iran. J. For. Poplar Res. 2023, 3, 1–15. [Google Scholar] [CrossRef]
- Diaconu, D.; Kahle, H.P.; Spiecker, H. Thinning Increases Drought Tolerance of European Beech: A Case Study on Two Forested Slopes on Opposite Sides of a Valley. Eur. J. Res. 2017, 136, 319–328. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European Tree Species to Drought Stress in Mixed versus Pure Forests: Evidence of Stress Release by Inter-Specific Facilitation. Plant Biol. 2013, 15, 483–495. [Google Scholar] [CrossRef]
- Antonucci, S.; Santopuoli, G.; Marchetti, M.; Tognetti, R.; Chiavetta, U.; Garfì, V. What Is Known About the Management of European Beech Forests Facing Climate Change? A Review. Curr. For. Rep. 2021, 7, 321–333. [Google Scholar] [CrossRef]
- Gömöry, D.; Krajmerová, D.; Hrivnák, M.; Longauer, R. Assisted Migration vs. Close-to-Nature Forestry: What Are the Prospects for Tree Populations under Climate Change? Cent. Eur. For. J. 2020, 66, 63–70. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PRP | Name of PRP | Latitude (N) | Longitude (E) | Species | Stand Age (Years) | Elevation (m) | Aspect | Slope (°) | Clim. Class. |
|---|---|---|---|---|---|---|---|---|---|
| Fagus sylvatica | |||||||||
| 1 | FagSyl510 | 50°50′11.8′′ | 15°38′32.5′′ | FS | 193 | 510 | NW | 15 | Cfb |
| 2 | FagSyl620 | 50°44′21.8′′ | 15°25′15.1′′ | FS, PA | 195 | 620 | SW | 22 | Cfb |
| 3 | FagSyl760 | 50°39′57.7″ | 15°53′05.2″ | FS, AP | 162 | 760 | NE | 35 | Dfb |
| 4 | FagSyl940 | 50°44′06.1′′ | 15°32′21.0′′ | FS, PA | 245 | 940 | E | 24 | Dfb |
| 5 | FagSyl1170 | 50°44′42.7′′ | 15°32′46.9′′ | FS, PA | 208 | 1170 | SW | 17 | Dfb |
| 6 | FagSyl1310 | 50°44′46.4′′ | 15°32′58.9′′ | FS, PM, SA | 146 | 1310 | SW | 21 | Dfc |
| Fagus orientalis | |||||||||
| 7 | FagOri360 | 40°54′43.9′′ | 31°12′27.6′′ | FO, QP, CB | 119 | 360 | NW | 23 | Cfb |
| 8 | FagOri570 | 40°51′42.5′′ | 31°18′10.8′′ | FO, QP, | 189 | 570 | N | 30 | Cfb |
| 9 | FagOri740 | 40°57′15.0′′ | 31°14′28.5′′ | FO, QP | 124 | 740 | SE | 5 | Cfb |
| 10 | FagOri950 | 40°49′37.7′′ | 31°25′05.6′′ | FO | 180 | 950 | W | 21 | Cfb |
| 11 | FagOri1150 | 40°46′52.8′′ | 31°28′00.3′′ | FO, AN, CB | 198 | 1150 | SW | 13 | Dfb |
| 12 | FagOri1430 | 40°47′41.8′′ | 31°28′16.2′′ | FO, AN | 239 | 1430 | NW | 16 | Dfb |
| PRP | dbh | h | v | HDR | N | BA | V | PAI | CC | SDI | BIO | CBIO |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| cm | m | m3 | Trees·ha−1 | m2·ha−1 | m3·ha−1 | m3·ha−1·y−1 | % | t·ha−1 | t·ha−1 | |||
| Fagus sylvatica | ||||||||||||
| 1 | 47.6 | 24.94 | 3.021 | 52.4 | 272 | 48.2 | 822 | 4.37 | 91.1 | 0.61 | 695 | 361 |
| 2 | 52.2 | 33.37 | 3.153 | 63.9 | 208 | 44.2 | 656 | 3.45 | 92.9 | 0.83 | 539 | 280 |
| 3 | 41.3 | 21.21 | 1.502 | 51.4 | 256 | 34.3 | 375 | 2.39 | 85.5 | 0.81 | 340 | 177 |
| 4 | 37.5 | 20.40 | 1.566 | 54.4 | 656 | 44.4 | 619 | 2.58 | 91.4 | 0.75 | 524 | 273 |
| 5 | 46.6 | 22.89 | 1.996 | 49.1 | 184 | 31.3 | 362 | 1.78 | 97.1 | 0.66 | 333 | 173 |
| 6 | 24.0 | 7.93 | 0.223 | 33.0 | 396 | 17.9 | 88 | 0.62 | 66.0 | 0.30 | 72 | 37 |
| Fagus orientalis | ||||||||||||
| 7 | 27.2 | 21.36 | 0.626 | 78.5 | 484 | 28.0 | 303 | 2.78 | 86.0 | 0.53 | 242 | 126 |
| 8 | 39.3 | 24.06 | 1.550 | 61.2 | 381 | 46.1 | 589 | 3.29 | 98.8 | 0.75 | 469 | 241 |
| 9 | 21.5 | 18.86 | 0.387 | 87.7 | 1080 | 39.2 | 418 | 3.67 | 94.6 | 0.81 | 318 | 166 |
| 10 | 35.5 | 19.31 | 1.302 | 54.4 | 276 | 27.2 | 359 | 2.11 | 78.7 | 0.46 | 306 | 160 |
| 11 | 29.2 | 19.05 | 0.793 | 65.3 | 284 | 18.9 | 225 | 1.20 | 79.7 | 0.35 | 173 | 91 |
| 12 | 26.9 | 14.04 | 0.596 | 52.1 | 360 | 20.5 | 215 | 0.93 | 84.3 | 0.40 | 168 | 87 |
| PRP | Cores (n) | Age Min–Max (Years) | RW Mean (mm) | RWI SD | EPS | Negative Pointer Years |
|---|---|---|---|---|---|---|
| 1 | 29 | 82–124 | 1.993 c | 0.174 | 0.94 | 2003, 2011, 2016, 2018, 2020 |
| 2 | 24 | 73–167 | 1.960 c | 0.194 | 0.88 | 2011, 2020 |
| 3 | 30 | 128–249 | 1.018 b | 0.221 | 0.92 | 1978, 2004, 2011, 2020 |
| 4 | 28 | 177–216 | 0.863 ab | 0.278 | 0.89 | 1948, 1952, 1956, 1981, 1984, 1985, 1996, 2020 |
| 5 | 30 | 161–260 | 0.946 ab | 0.286 | 0.93 | 1952, 1953, 1981, 1985, 1996, 2000 |
| 6 | 29 | 52–102 | 0.637 a | 0.251 | 0.91 | 1981, 2018, 2020 |
| 7 | 28 | 62–104 | 1.796 c | 0.198 | 0.92 | 1994, 2014 |
| 8 | 25 | 109–188 | 1.625 c | 0.279 | 0.85 | 1993, 2008 |
| 9 | 25 | 66–104 | 1.067 b | 0.288 | 0.88 | 1948, 1994, 2014 |
| 10 | 24 | 93–170 | 1.413 bc | 0.291 | 0.85 | 1955, 1977, 1993, 1994 |
| 11 | 26 | 79–140 | 1.034 b | 0.393 | 0.88 | 1963, 1993, 1994 |
| 12 | 23 | 155–234 | 0.679 a | 0.371 | 0.86 | 1947, 1993, 2007 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuchs, Z.; Vacek, Z.; Vacek, S.; Černý, J.; Cukor, J.; Šimůnek, V.; Gallo, J.; Hájek, V. Growth Responses of European Beech (Fagus sylvatica L.) and Oriental Beech (Fagus orientalis Lipsky) Along an Elevation Gradient Under Global Climate Change. Forests 2025, 16, 655. https://doi.org/10.3390/f16040655
Fuchs Z, Vacek Z, Vacek S, Černý J, Cukor J, Šimůnek V, Gallo J, Hájek V. Growth Responses of European Beech (Fagus sylvatica L.) and Oriental Beech (Fagus orientalis Lipsky) Along an Elevation Gradient Under Global Climate Change. Forests. 2025; 16(4):655. https://doi.org/10.3390/f16040655
Chicago/Turabian StyleFuchs, Zdeněk, Zdeněk Vacek, Stanislav Vacek, Jakub Černý, Jan Cukor, Václav Šimůnek, Josef Gallo, and Vojtěch Hájek. 2025. "Growth Responses of European Beech (Fagus sylvatica L.) and Oriental Beech (Fagus orientalis Lipsky) Along an Elevation Gradient Under Global Climate Change" Forests 16, no. 4: 655. https://doi.org/10.3390/f16040655
APA StyleFuchs, Z., Vacek, Z., Vacek, S., Černý, J., Cukor, J., Šimůnek, V., Gallo, J., & Hájek, V. (2025). Growth Responses of European Beech (Fagus sylvatica L.) and Oriental Beech (Fagus orientalis Lipsky) Along an Elevation Gradient Under Global Climate Change. Forests, 16(4), 655. https://doi.org/10.3390/f16040655