Analysis of the Effect of Climate Warming on Paludification Processes: Will Soil Conditions Limit the Adaptation of Northern Boreal Forests to Climate Change? A Synthesis



Abstract
1. Introduction
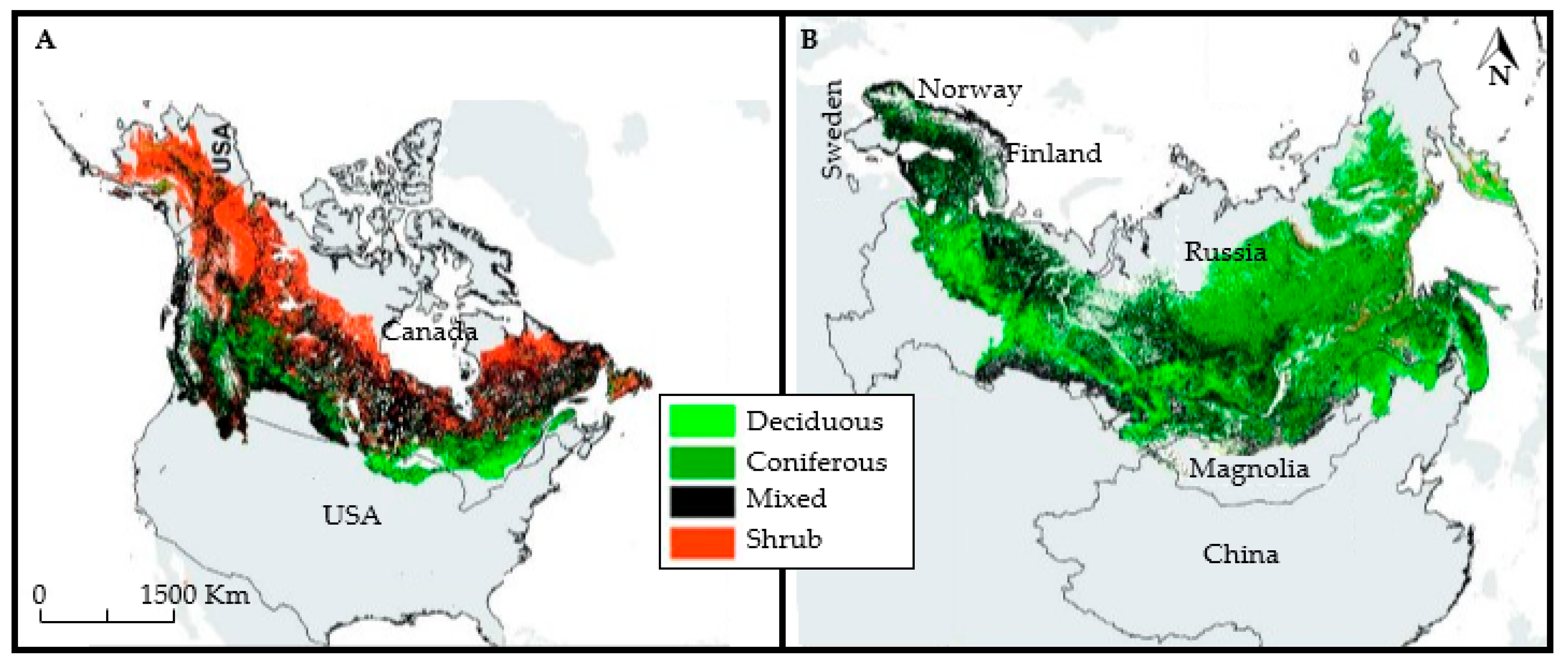
2. Study Area and Boreal Forest Soils
2.1. Sudy Area
2.2. Boreal Forest Soils and Classification
3. Expected Effect of Climate Warming on Boreal Soil Processes
4. Anticipated Forest Soils and Species Distribution in Response to Global Warming
5. Potential Effects of Climate Warming on Paludification in the Clay Belt Region
5.1. The Anticipated Effects on Paludification and Related Changes in Vegetation
5.2. Silvicultural Treatment Changes under the Three Scenarios
6. Will Soil Conditions Limit the Adaptation of Boreal Ecosystems to Climate Change? Concluding Synthesis
6.1. Boreal Ecosystem Response to Climate-Induced Changes
6.2. Effects in Future Silvicultural Treatments in the Context of Expected Changes of Paludification under the Three Selected Scenarios
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thiffault, E. Chapter 5—Boreal forests and soils. Dev. Soil Sci. 2019, 36, 59–82. [Google Scholar] [CrossRef]
- Mason, K.E.; Oakley, S.; Street, L.E.; Arro´niz-Crespo, M.; Jones, D.L.; DeLuca, T.H.; Ostle, N.J. Boreal Forest Floor Greenhouse Gas Emissions Across a Pleurozium schreberi-Dominated, Wildfire-Disturbed Chronosequence. Ecosystems 2019, 22, 1381–1392. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- McNamara, N.P.; Gregg, R.; Oakley, S.; Stott, A.; Rahman, M.T.; Murrell, J.C.; Wardle, D.A.; Bardgett, R.D.; Ostle, N.J. Soil Methane Sink Capacity Response to a Long-Term Wildfire Chronosequence in Northern Sweden. PLoS ONE 2015, 10, e0129892. [Google Scholar] [CrossRef] [PubMed]
- Van Cleve, K.; Powers, R.F. Soil carbon, soil formation, and ecosystem development. In Carbon Forms and Functions in Forest Soils. Madison (WI); McFee, W.W., Kelly, J.M., Eds.; Soil Science Society of America: Madison WI, USA, 1995. [Google Scholar] [CrossRef]
- Wickland, K.P.; Neff, J.C. Decomposition of soil organic matter from boreal black spruce forest: Environmental and chemical controls. Biogeochemistry 2008, 87, 29–47. [Google Scholar] [CrossRef]
- Gauthier, S.; Bernier, P.; Burton, P.J.; Edwards, J.; Isaac, K.; Isabel, N.; Jayen, K.; Le Goff, H.; Nelson, E.A. Climate change vulnerability and adaptation in the managed. Environ. Rev. 2014, 22, 256–285. [Google Scholar] [CrossRef]
- Hamann, A.; Wang, T. Potential effects of climate change on ecosystem and tree species distribution in British Columbia. Ecology 2006, 87, 2773–2786. [Google Scholar] [CrossRef]
- Hannah, L.; Roehrdanz, P.R.; Krishna Bahadur, K.C.; Fraser, E.D.; Donatti, C.I.; Saenz, L.; Wright, T.M.; Hijmans, R.J.; Mulligan, M.; Berg, A.; et al. The environmental consequences of climate-driven agricultural frontiers. PLoS ONE 2020, 15, e0228305. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.H.; Tian, H.; Xu, X.; Pan, Y.; Chen, G.; Lin, W. Extension of the growing season due to delayed autumn over mid and high latitudes in North America during 1982–2006. Glob. Ecol. Biogeogr. 2012, 21, 260–271. [Google Scholar] [CrossRef]
- D’Orangeville, L.; Houle, D.; Duchesne, L.; Phillips, R.P.; Bergeron, Y.; Kneeshaw, D. Beneficial effects of climate warming on boreal tree growth may be transitory. Nat. Commun. 2018, 9, 3213. [Google Scholar] [CrossRef]
- Reichsten, M. Impact of climate change on forest soils carbon: Principles, factors, models, uncertainties. For. Clim. Chang. 2007, 125–135. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Callaghan, T.V.; Bergeron, Y.; Fukuda, M.; Johnstone, J.F.; Juday, G.; Zimov, S.A. Global change and the boreal forest: Thresholds, shifting states or gradual change? Ambio 2004, 33, 361–365. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, K.E.B.; Gower, S.T.; Norman, J.M. Comparison of net primary production and light-use dynamics of two boreal black spruce forest communities. Ecosystems 2003, 6, 236–247. [Google Scholar] [CrossRef]
- Bergeron, Y.; Cyr, D.; Girardin, M.P.; Carcaillet, C. Will climate change drive 21st century burn rates in Canadian boreal forests outside of natural variability: Collating global climate model experiments with sedimentary charcoal data. Int. J. Wildland Fire 2010, 19, 1127–1139. [Google Scholar] [CrossRef]
- Roy, V.; Bernier, P.Y.; Plamondon, A.P.; Ruel, J.C. Effect of drainage and microtopography in forested wetlands on the microenvironment and growth of planted black spruce seedlings. Can. J. For. Res. 1999, 29, 563–574. [Google Scholar] [CrossRef]
- Oechel, W.C.; Van Cleve, K. The role of bryophytes in nutrient cycling in the taiga. In Forest Ecosystems in the Alaskan Taiga: A Synthesis of Structure and Function; van Cleve, K., Chapin, F.S., III, Flanagan, P.W., Viereck, L.A., Dyrness, C.T., Eds.; Springer: New York, NY, USA, 1986; pp. 121–137. [Google Scholar]
- Johnstone, J.F.; Kasischke, E.S. Stand–level effects of soil burn severity on postfire regeneration in a recently burned black spruce forest. Can. J. For. Res. 2005, 35, 2151–2163. [Google Scholar] [CrossRef]
- Vogel, J.G.; Valentine, D.W.; Ruess, R.W. Soil and root respiration in mature Alaskan black spruce forests that vary in soil organic matter decomposition rates. Can. J. For. Res. 2005, 35, 161–174. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Mitigation of Climate Change. Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Edenhofer, O., Pichs-Madruga, R., Sokona, Y., Farahani, E., Kadner, S., Seyboth, K., Adler, A., Baum, I., Brunner, S., Eickemeier, P., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Akinremi, O.O.; McGinn, S.M.; Cutforth, H.W. Precipitation trends on the Canadian prairies. J. Clim. 1999, 12, 2996–3003. [Google Scholar] [CrossRef]
- Osterkamp, T.E.; Romanovsky, V.E. Evidence for warming and thawing of discontinuous permafrost in Alaska. Permafr. Periglac. Process. 1999, 10, 17–37. [Google Scholar] [CrossRef]
- Oechel, W.C.; Vourlitis, G.L.; Hastings, S.J.; Ault, R.P., Jr.; Bryant, P. The effects of water table manipulation and elevated temperature on the net CO2 flux of wet sedge tundra ecosystems. Glob. Chang. Biol. 1998, 4, 77–90. [Google Scholar] [CrossRef]
- Beniston, M. Variations of snow depth and duration in the Swiss Alps over the last 50 years: Links to changes in large–scale climatic forcings. Clim. Chang. 1997, 36, 281–300. [Google Scholar] [CrossRef]
- Sazonova, T.S.; Romanovsky, V.E.; Walsh, J.E.; Sergueev, D.O. Permafrost dynamics in the 20th and 21st centuries along the East Siberian transect. J. Geophys. Res. Atmos. 2004, 109, 1–20. [Google Scholar] [CrossRef]
- Zhang, T.; Frauenfeld, O.W.; Serreze, M.C.; Etringer, A.; Oelke, C.; McCreight, J.; Barry, R.G.; Gilichinsky, D.; Yang, D.; Ye, H.; et al. Spatial and temporal variability in active layer thickness over the Russian Arctic drainage basin. J. Geophys. Res. Atmos. 2005, 110, 1–14. [Google Scholar] [CrossRef]
- Carpino, O.A.; Berg, A.A.; Quinton, W.L.; Adams, J.R. Climate change and permafrost thaw-induced boreal forest loss in northwestern Canada. Environ. Res. Lett. 2018, 13, 084018. [Google Scholar] [CrossRef]
- Gillett, N.P.; Weaver, A.J.; Zwiers, F.W.; Flannigan, M.D. Detecting the effect of climate change on Canadian forest fires. Geophys. Res. Lett. 2004, 31, 1–4. [Google Scholar] [CrossRef]
- Flannigan, M.D.; Logan, K.A.; Amiro, B.D.; Skinner, W.R.; Stocks, B.J. Future area burned in Canada. Clim. Chang. 2005, 72, 1–16. [Google Scholar] [CrossRef]
- Keyser, A.R.; Kimball, J.S.; Nemani, R.R.; Running, S.W. Simulating the effects of climate change on the carbon balance of North American high–latitude forests. Glob. Chang. Biol. 2000, 6, 185–195. [Google Scholar] [CrossRef]
- McGuire, A.D.; Sitch, S.; Clein, J.S.; Dargaville, R.; Esser, G.; Foley, J.; Heimann, M.; Joos, F.; Kaplan, J.; Kicklighter, D.W.; et al. Carbon balance of the terrestrial biosphere in the twentieth century: Analyses of CO2, climate and land use effects with four process–based ecosytem models. Glob. Biogeochem. Cycles 2001, 15, 183–206. [Google Scholar] [CrossRef]
- Lafleur, B.; Paré, D.; Fenton, N.J.; Bergeron, Y. Growth and nutrition of black spruce seedlings in response to disruption of Pleurozium and Sphagnum moss carpets in boreal forested peatlands. Plant. Soil 2011, 345, 141–153. [Google Scholar] [CrossRef]
- Kirschbaum, M.U.F. Forest growth and species distribution in a changing climate. Tree Physiol. 2000, 20, 309–322. [Google Scholar] [CrossRef]
- Trainor, S.F.; Calef, M.; Natcher, D.; Chapin Iii, F.S.; McGuire, A.D.; Huntington, O.; Duffy, P.; Rupp, T.S.; Dewilde, L.; Kwart, M.; et al. Vulnerability and adaptation to climate–related fire impacts in rural and urban interior Alaska. Polar Res. 2009, 28, 100–118. [Google Scholar] [CrossRef]
- Bradshaw, C.J.A.; Warkentin, I.G.; Sodhi, N.S. Urgent preservation of boreal carbon stocks and biodiversity. Trends Ecol. Evol. 2009, 24, 541–548. [Google Scholar] [CrossRef]
- DeLuca, T.H.; Boisvenue, C. Boreal forest soil carbon: Distribution, function, and modelling. Forestry 2012, 85, 161–184. [Google Scholar] [CrossRef]
- Hawkins, B.A. Summer vegetation, deglaciation and the anomalous bird diversity gradient in eastern North America. Glob. Ecol. Biogeogr. 2004, 13, 321–325. [Google Scholar] [CrossRef]
- Gorham, E. Northern peatlands: Role in the carbon cycle and probable responses to climatic warming. Ecol. Appl. 1991, 1, 182–195. [Google Scholar] [CrossRef]
- Bonan, G.B.; Shugart, H.H. Environmental factors and ecological processes in boreal forests. Annu. Rev. Ecol. Syst. 1989, 20, 1–28. [Google Scholar] [CrossRef]
- Moore, T.R. Soil Formation in Northeastern Canada. Ann. Assoc. Am. Geogr. 1978, 68, 518–534. [Google Scholar] [CrossRef]
- Stützer, A. Podzolisation as a soil forming process in the alpine belt of Rondane, Norway. Geoderma 1999, 91, 237–248. [Google Scholar] [CrossRef]
- Lundström, U.S.; Van Breemen, N.; Bain, D. The podzolization process. A review. Geoderma 2000, 94, 91–107. [Google Scholar] [CrossRef]
- Sauer, D.; Sponagel, H.; Sommer, M.; Giani, L.; Jahn, R.; Stahr, K. Podzol: Soil of the year A review on its genesis, occurrence, and functions. J. Plant Nutr. Soil Sci. 2007, 170, 581–597. [Google Scholar] [CrossRef]
- Seibert, J.; Stendahl, J.; Sørensen, R. Topographical influences on soil properties in boreal forests. Geoderma 2007, 141, 139–148. [Google Scholar] [CrossRef]
- Lavoie, M.; Paré, D.; Fenton, N.; Groot, A.; Taylor, K. Paludification and management of forested peatlands in Canada: A literature review. Environ. Rev. 2005, 13, 21–50. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources2014, Update 2015, International Soil Classification System for Naming Soils; World Soil Resources Report 106; Food and Agriculture Organization of the United Nations: Rome, Italy, 2015. [Google Scholar]
- Soil Classification Working Group. The Canadian System of Soil Classification, 3rd ed.; Agriculture and Agri-Food Canada Publication 1646; Agriculture and Agri-Food Canada: Ottawa, ON, Canada, 1998; p. 187.
- Domisch, T.; Finér, L.; Laine, J.; Laiho, R. Decomposition and nitrogen dynamics of litter in peat soils from two climatic regions under different temperature regimes. Eur. J. Soil Biol. 2006, 42, 74–81. [Google Scholar] [CrossRef]
- Liski, J.; Nissinen, A.; Erhard, M.; Taskinen, O. Climatic effects on litter decomposition from arctic tundra to tropical rainforest. Glob. Chang. Biol. 2003, 9, 575–584. [Google Scholar] [CrossRef]
- Shaw, M.R.; Harte, J. Control of litter decomposition in a subalpine meadow–sagebrush steppe ecotone under climate change. Ecol. Appl. 2001, 11, 1206–1223. [Google Scholar]
- Goulden, M.L.; Wofsy, S.C.; Harden, J.W.; Trumbore, S.E.; Crill, P.M.; Gower, S.T.; Fries, T.; Daube, B.C.; Fan, S.M.; Sutton, D.J.; et al. Sensitivity of boreal forest carbon balance to soil thaw. Science 1998, 279, 214–217. [Google Scholar] [CrossRef]
- Agren, G.I.; Bosatta, E.; Magill, A.H. Combining theory and experiment to understand effects of inorganic nitrogen on litter decomposition. Oecologia 2001, 128, 94–98. [Google Scholar] [CrossRef]
- Gauthier, S.; Bernier, P.; Kuuluvainen, T.; Shvidenko, A.Z.; Schepaschenko, D.G. Boreal Forest Health and Global Change. Science 2015, 349, 819–822. [Google Scholar] [CrossRef]
- Aerts, R. The freezer defrosting: Global warming and litter decomposition rates in cold biomes. J. Ecol. 2006, 94, 713–724. [Google Scholar] [CrossRef]
- Zhang, D.; Hui, D.; Luo, Y.; Zhou, G. Rates of litter decomposition in terrestrial ecosystems: Global patterns and controlling factors. J. Plant Ecol. 2008, 1, 85–93. [Google Scholar] [CrossRef]
- Rustad, L.E.; Campbell, J.L.; Marion, G.M.; Norby, R.J.; Mitchell, M.J.; Hartley, A.E.; Cornelissen, J.H.C.; Gurevitch, J.; Alward, R.; Beier, C.; et al. A meta–analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia 2001, 126, 543–562. [Google Scholar] [CrossRef] [PubMed]
- Dabros, A.; Fyles, J.W. Effects of open–top chambers and substrate type on biogeochemical processes at disturbed boreal forest sites in northwestern Quebec. Plant Soil 2010, 327, 465–469. [Google Scholar] [CrossRef]
- Pajari, B. Soil respiration in a poor upland site of Scots pine stand subjected to elevated temperatures and atmospheric carbon concentration. Plant Soil 1995, 168–169, 563–570. [Google Scholar] [CrossRef]
- Robinson, C.H.; Wookey, P.A.; Parsons, A.N.; Potter, J.A.; Callaghan, T.V.; Lee, J.A.; Press, M.C.; Welker, J.M. Responses of plant litter decomposition and nitrogen mineralisation to simulated environmental change in a high arctic polar semi–desert and a subarctic dwarf shrub heath. Oikos 1995, 74, 503–512. [Google Scholar] [CrossRef]
- Eliasson, P.E.; McMurtrie, R.E.; Pepper, D.A.; Strömgren, M.; Linder, S.; Agren, G.I. The response of heterotrophic CO2 flux to soil warming. Global. Chang. Biol. 2005, 11, 167–181. [Google Scholar] [CrossRef]
- Kaplan, J.O.; Bigelow, N.H.; Prentice, I.C.; Harrison, S.P.; Bartlein, P.J.; Christensen, T.R.; Cramer, W.; Matveyeva, N.V.; McGuire, A.D.; Murray, D.F.; et al. Climate change and Arctic ecosystems: 2. Modeling, paleodata–model comparisons, and future projections. J. Geophys. Res. Atmos. 2003, 108, 8171. [Google Scholar] [CrossRef]
- Gamache, I.; Payette, S. Latitudinal response of subarctic tree lines to recent climate change in eastern Canada. J. Biogeogr. 2005, 32, 849–862. [Google Scholar] [CrossRef]
- Dunn, A.L.; Wofsy, S.C.; Bright, A.V.H. Landscape heterogeneity, soil climate, and carbon exchange in a boreal black spruce forest. Ecol. Appl. 2009, 19, 495–504. [Google Scholar] [CrossRef]
- Kellomaki, S.; Kolstrom, M. Simulation of tree species composition and organic matter accumulation in Finnish boreal forests under changing climatic conditions. Vegetation 1992, 102, 47–68. [Google Scholar] [CrossRef]
- Côté, L.; Brown, S.; Paré, D.; Fyles, J.; Bauhus, J. Dynamics of carbon and nitrogen mineralization in relation to stand type, stand age and soil texture in the boreal mixed wood. Soil Biol. Biochem. 2000, 32, 1079–1090. [Google Scholar] [CrossRef]
- Jin, X.Y.; Jin, H.J.; Iwahana, G.; Marchenko, S.S.; Luo, D.L.; Li, X.Y.; Liang, S.H. Impacts of climate-induced permafrost degradation on vegetation: A review. Adv. Clim. Chang. Res. 2020. [Google Scholar] [CrossRef]
- Serreze, M.C.; Walsh, J.E.; Chapin Iii, F.S.; Osterkamp, T.; Dyurgerov, M.; Romanovsky, V.; Oechel, W.C.; Morison, J.; Zhang, T.; Barry, R.G. Observational evidence of recent change in the northern high–latitude environment. Clim. Chang. 2000, 46, 159–207. [Google Scholar] [CrossRef]
- Chapin, F.S., III; McGuire, A.D.; Randerson, J.; Pielke, R., Sr.; Baldocchi, D.; Hobbie, S.E.; Roulet, N.; Eugster, W.; Kasischke, E.; Rastetter, E.B.; et al. Arctic and boreal ecosystems of western North America as components of the climate system. Glob. Chang. Biol. 2000, 6, 211–223. [Google Scholar] [CrossRef]
- Crawford, R.M.M.; Jeffree, C.E.; Rees, W.G. Paludification and forest retreat in northern oceanic environments. Ann. Bot. 2003, 91, 213–226. [Google Scholar] [CrossRef]
- Lavoie, M.; Paré, D.; Bergeron, Y. Impact of global change and forest management on carbon sequestration in northern forested peatlands. Environ. Rev. 2005, 13, 199–240. [Google Scholar] [CrossRef]
- Vygodskaya, N.N.; Groisman, P.Y.; Tchebakova, N.M.; Kurbatova, J.A.; Panfyorov, O.; Parfenova, E.I.; Sogachev, A.F. Ecosystems and climate interactions in the boreal zone of northern Eurasia. Environ. Res. Lett. 2007, 2, 045033. [Google Scholar] [CrossRef]
- Fenton, N.J.; Bergeron, Y. Sphagnum community change after partial harvest in black spruce boreal forests. For. Ecol. Manag. 2007, 242, 24–33. [Google Scholar] [CrossRef]
- Skre, O.; Baxter, R.; Crawford, R.M.M.; Callaghan, T.V.; Fedorkov, A. How will the tundra–taiga interface respond to climate change? Ambio 2002, 31, 37–46. [Google Scholar]
- Veillette, J.J. Evolution and paleohydrology of glacial Lakes Barlow and Ojibway. Quat. Sci. Rev. 1994, 13, 945–971. [Google Scholar] [CrossRef]
- Laamrani, A.; Valeria, O.; Fenton, N.; Bergeron, Y. Landscape-scale influence of topography on organic layer accumulation in paludified boreal forests. For. Sci. 2014, 60, 579–590. [Google Scholar] [CrossRef]
- Laamrani, A.; Valeria, O.; Fenton, N.; Bergeron, Y.; Cheng, L.Z. The role of mineral soil topography on the spatial distribution of organic layer thickness in a paludified boreal landscape. Geoderma 2014, 221–222, 70–78. [Google Scholar] [CrossRef]
- Laamrani, A.; Valeria, O. Ranking Importance of Topographical Surface and Subsurface Parameters on Paludification in Northern Boreal Forests Using Very High Resolution Remotely Sensed Datasets. Sustainability 2020, 12, 577. [Google Scholar] [CrossRef]
- MacIver, D.C.; Isaac, J.L. Profils Bioclimatiques Pour le Canada, 1951–1980; Environnement Canada, Service de L’environnement Atmosphérique: Toronto, ON, Canada, 1989. [Google Scholar]
- Bush, E.; Lemmen, D.S. (Eds.) Canada’s Changing Climate Report; Government of Canada: Ottawa, ON, Canada, 2019; p. 444. Available online: https://changingclimate.ca/CCCR2019/ (accessed on 28 September 2020).
- Houghton, J.T.A. Climate Change 2001: The Scientific Basis. In Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: New York, NY, USA, 2001; p. 892. [Google Scholar]
- Laamrani, A. Effect of Climate Change on Boreal Forest Soils: Will Soil Conditions Limit the Adaptation of Boreal Ecosystems to Climate Change? Ph.D. Thesis, University of Quebec in Abitibi-Témiscamingue (UQAT), Rouyn-Noranda, QC, Canada, 2010. Environmental Synthesis Report, Environmental Sciences Program. Available online: https://www.researchgate.net/publication/263966215_Effect_of_climate_change_on_boreal_forest_soils_Will_soil_conditions_limit_the_adaptation_of_boreal_ecosystems_to_climate_change (accessed on 29 September 2020).
- Bonan, G.B.; Shugart, H.H.; Urban, D.L. The sensitivity of some high-latitude boreal forests to climatic parameters. Clim. Chang. 1990, 16, 9–29. [Google Scholar] [CrossRef]
- Helbig, M.; Waddington, J.M.; Alekseychik, P.; Amiro, B.D.; Aurela, M.; Barr, A.G.; Black, T.A.; Blanken, P.D.; Carey, S.K.; Chen, J.; et al. Increasing contribution of peatlands to boreal evapotranspiration in a warming climate. Nat. Clim. Chang. 2020, 10, 555–560. [Google Scholar] [CrossRef]
- Spittlehouse, D.L.; Stewart, R.B. Adapting to climate change in forest management. BC Ecosyst. Manag. 2003, 4, 7–17. [Google Scholar]
- Marschall, M.; Proctor, M.C.F. Are Bryophytes Shade Plants? Photosynthetic Light Responses and Proportions of Chlorophyll a, Chlorophyll b and Total Carotenoids. Ann. Bot. 2004, 94, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Lafleur, B.; Fenton, N.J.; Bergeron, Y. Forecasting the development of boreal paludified forests in response to climate change: A case study using Ontario ecosite classification. For. Ecosyst. 2015, 2, 3. [Google Scholar] [CrossRef]
- Spittlehouse, D.L. Integrating climate change adaptation into forest management. For. Chron. 2005, 81, 691–695. [Google Scholar] [CrossRef]
- Bernier, P.; Schoene, D. Adapting forests and their management to climate change: An overview. Unasylva 2009, 60, 5–11. [Google Scholar]
- Laamrani, A.; Valeria, O.; Fenton, N.; Bergeron, Y.; Cheng, L.Z. Effects of topography and thickness of organic layer on productivity of black spruce boreal forests of the Canadian Clay Belt region. For. Ecol. Manag. 2014, 330, 144–157. [Google Scholar] [CrossRef]
- Lafleur, B.; Fenton, N.J.; Paré, D.; Simard, M.; Bergeron, Y. Contrasting effects of season and method of harvest on soil properties and the growth of black spruce regeneration in the boreal forested Peatlands of Eastern Canada. Silva Fenn. 2010, 44, 799–813. [Google Scholar] [CrossRef]
- Simard, M.; Bernier, P.Y.; Bergeron, Y.; Paré, D.; Guérine, L. Paludification dynamics in the boreal forest of the James Bay Lowlands: Effect of time since fire and topography. Can. J. For. Res. 2009, 39, 546–552. [Google Scholar] [CrossRef]
- Henneb, M.; Valeria, O.; Thiffault, N.; Fenton, N.J.; Bergeron, Y. Effects of Mechanical Site Preparation on Microsite Availability and Growth of Planted Black Spruce in Canadian Paludified Forests. Forests 2019, 10, 670. [Google Scholar] [CrossRef]
- Lafleur, B.; Fenton, N.J.; Simard, M.; Leduc, A.; Pare, D.; Valeria, O.; Bergeron, Y. Ecosystem management in paludified boreal forests: Enhancing wood production, biodiversity, and carbon sequestration at the landscape level. For. Ecosyst. 2018, 5, 27. [Google Scholar] [CrossRef]
- Splawinski, T.B.; Schab, A.; Leduc, A.; Valeria, O.; Cyr, D.; Pascual Puigdevall, J.; Gauthier, S.; Bergeron, Y. Ajustement des stratégies de production de bois dans certaines portions sensibles de la forêt boréale. In Rapport Présenté au Ministère des Forêts, de la Faune et des Parcs Par la Chaire Industrielle CRSNG UQAT-UQAM en Aménagement Forestier Durable; Ministère des Forêts, de la Faune et des Parcs du Québec: Quebec, QC, Canada, 2019; pp. 1–120. [Google Scholar]
- Mansuy, N.; Valeria, O.; Laamrani, A.; Fenton, N.; Guindon, L.; Bergeron, Y.; Beaudoin, A.; Légaré, S. Digital mapping of paludification in soils under black spruce forests of eastern Canada. Geoderma Reg. 2018, 15, e00194. [Google Scholar] [CrossRef]
- Thompson, D.K.; Simpson, B.N.; Whitman, E.; Barber, Q.E.; Parisien, M.-A. Peatland Hydrological Dynamics as A Driver of Landscape Connectivity and Fire Activity in the Boreal Plain of Canada. Forests 2019, 10, 534. [Google Scholar] [CrossRef]
- Le Stum-Boivin, É.; Magnan, G.; Garneau, M.; Fenton, N.J.; Grondin, P.; Bergeron, Y. Spatiotemporal evolution of paludification associated with autogenic and allogenic factors in the black spruce-moss boreal forest of Québec, Canada. Quat. Res. 2019, 91, 650–664. [Google Scholar] [CrossRef]
- Kuhry, P. Palsa and peat plateau development in the Hudson Bay Lowlands, Canada: Timing, pathways and causes. Boreas 2008, 37, 316–327. [Google Scholar] [CrossRef]
- Jump, A.S.; Penuelas, J. Running to stand still: Adaptation and the response of plants to rapid climate change. Ecol. Lett. 2005, 8, 1010–1020. [Google Scholar] [CrossRef]


{kind=link}
| Soil Characteristics/Drainage Conditions | Horizon | Features/Composition |
|---|---|---|
| Organic/ Well-drained | L | Fresh organic residues, recognizable plant material, i.e., leaves, on the surface of the forest floor. |
| F | Decomposed plant material, i.e., roots, but the origins of plant residues are still distinguishable. | |
| H | Humified plant material where plant residues are not recognizable, with the exception of some roots or wood. | |
| Organic/ Poorly drained | Of | Fibric horizon where there are more roots and amorphous material than moss detritus |
| Om | Mesic horizon where plant residues are partly decomposed/amorphous. Intermediate decomposition between Of and Oh. | |
| Oh | Humic horizon with highly decomposed organics, amorphous and unrecognizable organics. | |
| Mineral | Ah | Typically a brown silt loam, sometimes grading into sandy loam or loam, occasionally with charcoal and/or rocks |
| Scenarios | Site Response | Vegetation Changes † | Silvicultural Treatments ‡ | |
|---|---|---|---|---|
| Indicators | Paludification | |||
| S1: ↑T & ↑ P | ↑ ET & WTL = | = or slightly favored | BS + MW + WS | TC + PB |
| S2: ↑T & ↓P | ↑ ET & ↓WTL ↑ decomp. rate ↓ Sphagnum growth | ↓ & ↑ soil nutrient availability | AS + JP + WS | TC + PC + Rep.: JP, WS, Po |
| S3: ↑T & P = | ↑ ET & ↓WTL ↑ decomp. rate ↓Sphagnum growth | ↓ stands prone to paludification | AS + MW + BS | TC + PB + Rep.: JP, WS, Po |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laamrani, A.; Valeria, O.; Chehbouni, A.; Bergeron, Y. Analysis of the Effect of Climate Warming on Paludification Processes: Will Soil Conditions Limit the Adaptation of Northern Boreal Forests to Climate Change? A Synthesis. Forests 2020, 11, 1176. https://doi.org/10.3390/f11111176
Laamrani A, Valeria O, Chehbouni A, Bergeron Y. Analysis of the Effect of Climate Warming on Paludification Processes: Will Soil Conditions Limit the Adaptation of Northern Boreal Forests to Climate Change? A Synthesis. Forests. 2020; 11(11):1176. https://doi.org/10.3390/f11111176
Chicago/Turabian StyleLaamrani, Ahmed, Osvaldo Valeria, Abdelghani Chehbouni, and Yves Bergeron. 2020. "Analysis of the Effect of Climate Warming on Paludification Processes: Will Soil Conditions Limit the Adaptation of Northern Boreal Forests to Climate Change? A Synthesis" Forests 11, no. 11: 1176. https://doi.org/10.3390/f11111176
APA StyleLaamrani, A., Valeria, O., Chehbouni, A., & Bergeron, Y. (2020). Analysis of the Effect of Climate Warming on Paludification Processes: Will Soil Conditions Limit the Adaptation of Northern Boreal Forests to Climate Change? A Synthesis. Forests, 11(11), 1176. https://doi.org/10.3390/f11111176