Plot-Scale Agroforestry Modeling Explores Tree Pruning and Fertilizer Interactions for Maize Production in a Faidherbia Parkland


Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Field Experiments for Model Calibration and Testing
2.2.1. Experimental Design and Maize Establishment
2.2.2. Measurements
2.2.3. Modeling
2.2.4. Virtual Experiments
3. Results
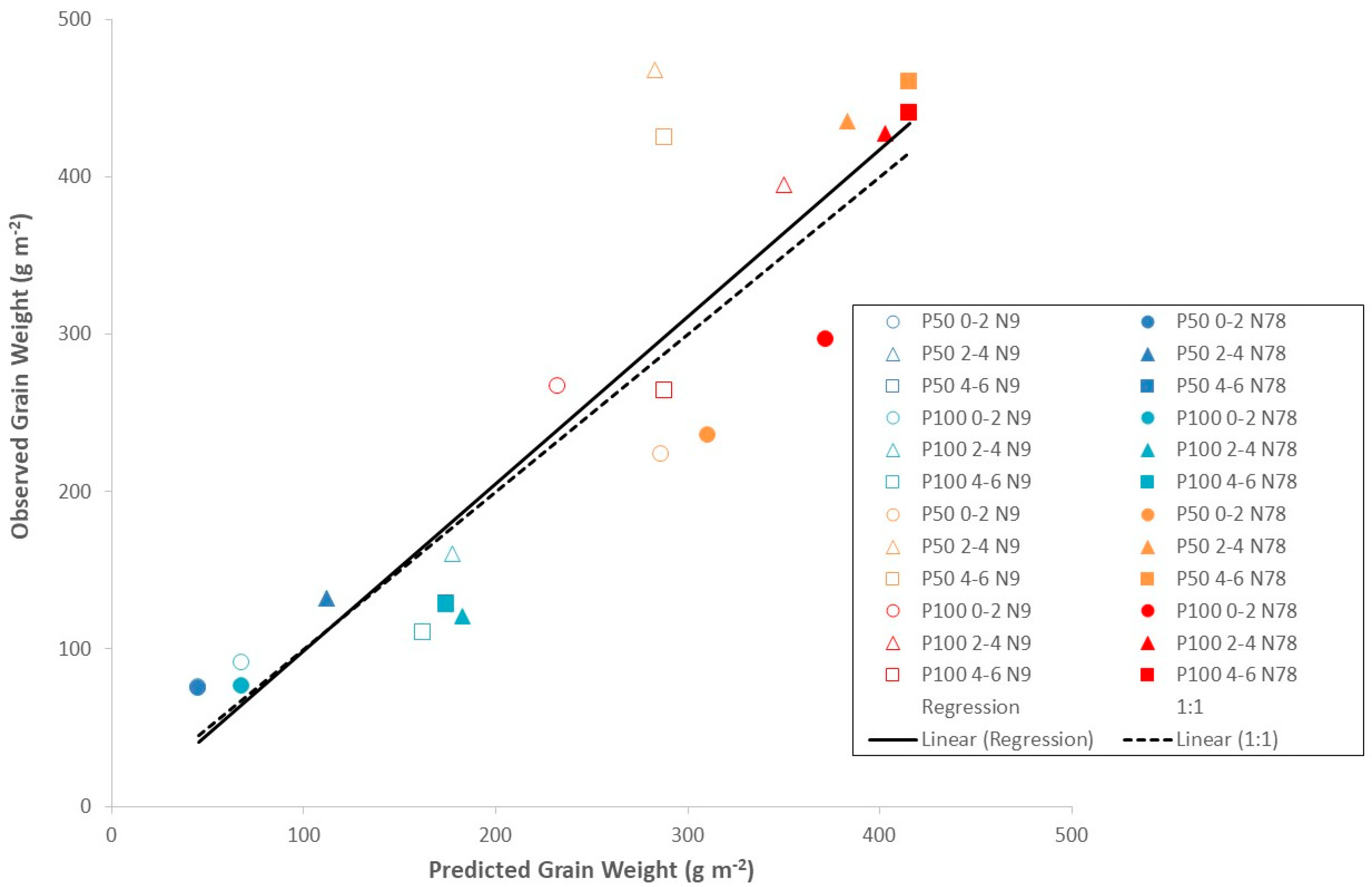
3.1. Evaluation of Model Performance
3.2. Virtual Experiments
3.2.1. Light Experiment
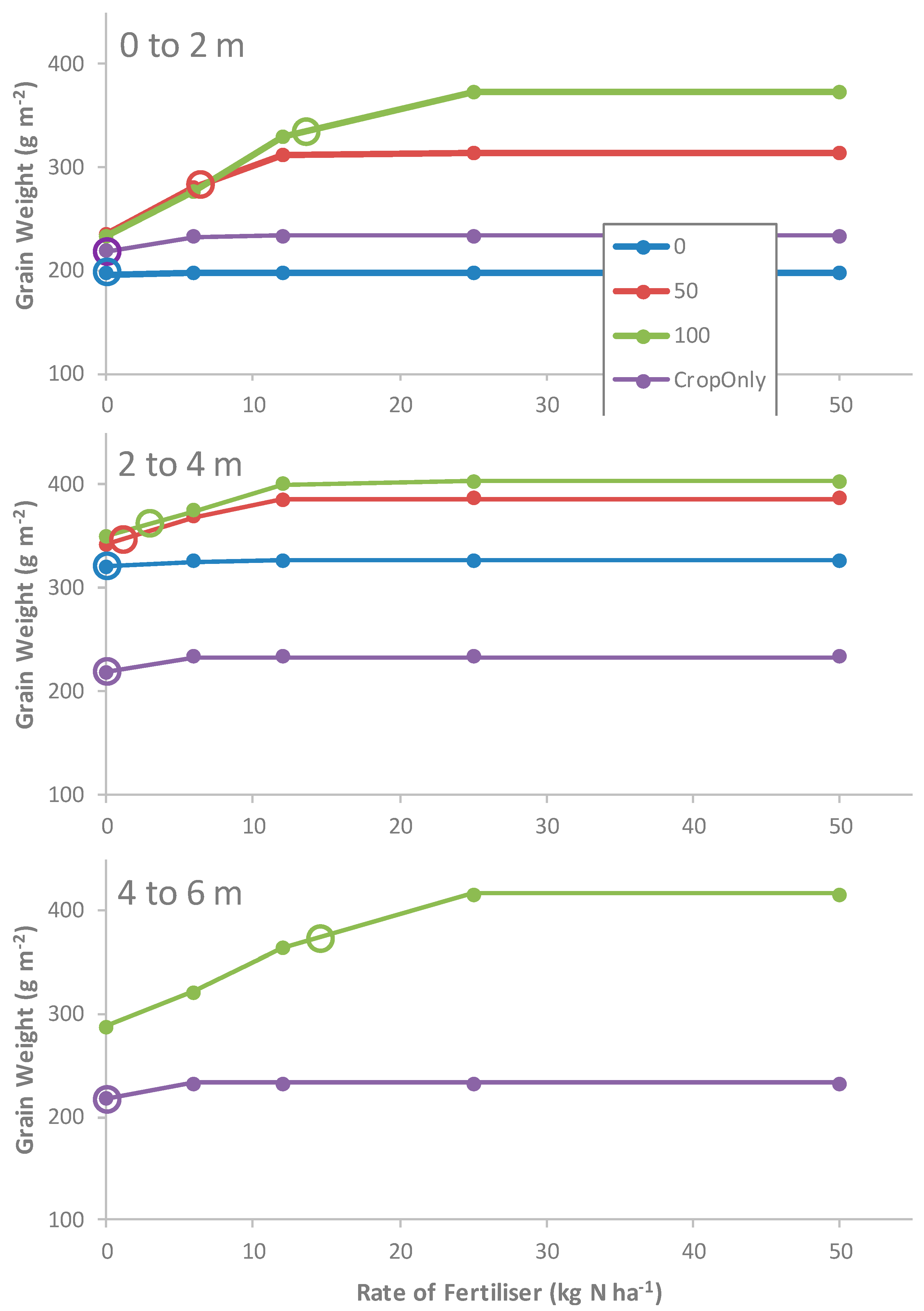
3.2.2. N Fertilizer, Pruning, and Zone Experiment
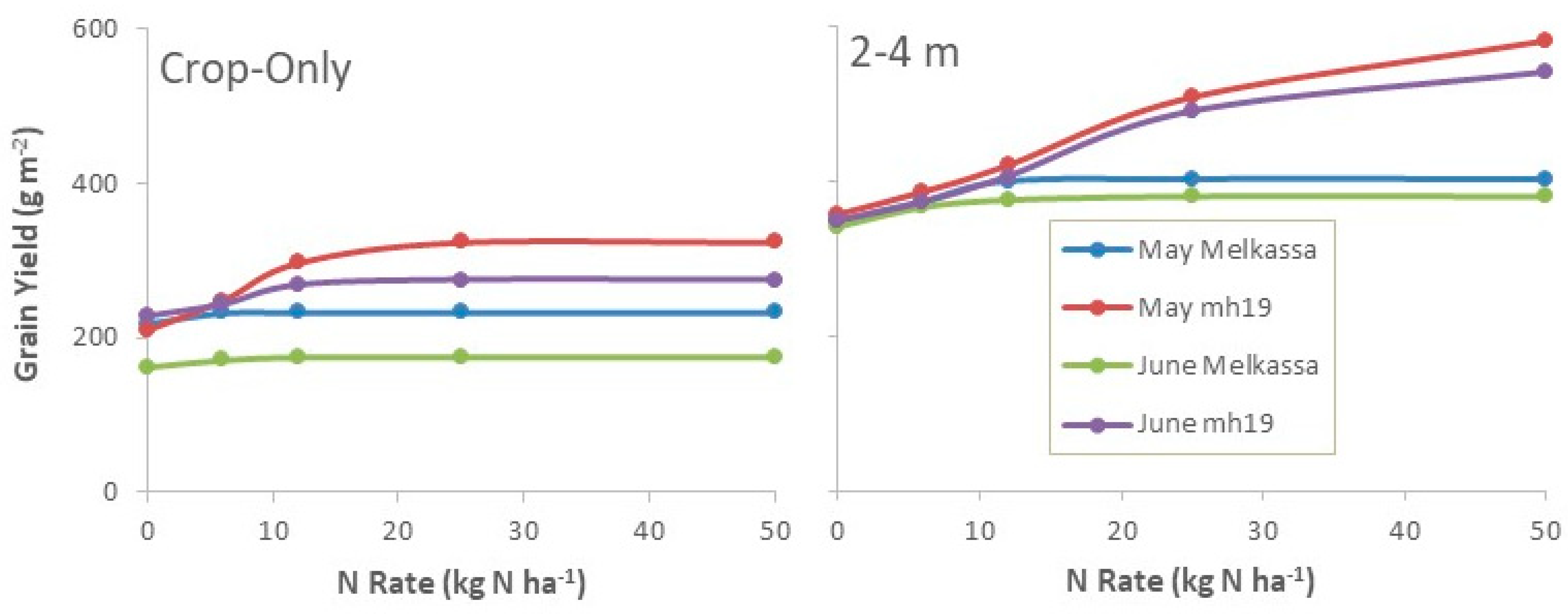
3.2.3. Genotype and Sowing Date Experiment
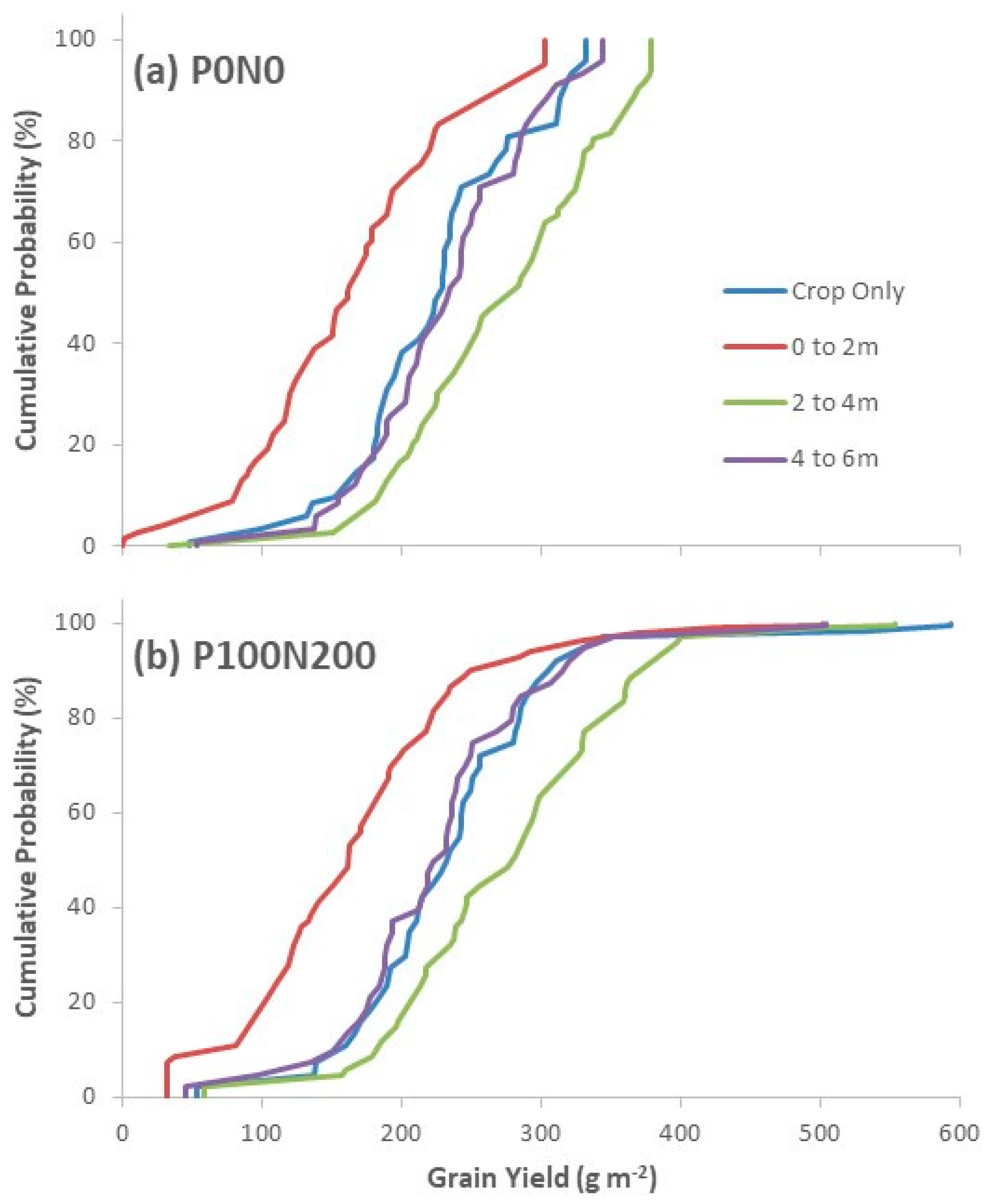
3.2.4. Climate Variability Experiment
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tittonell, P.; Giller, K.E. When yield gaps are poverty traps: The paradigm of ecological intensification in African smallholder agriculture. Field Crop Res. 2013, 143, 76–90. [Google Scholar] [CrossRef]
- Vågen, T.-G.; Winowiecki, L.A.; Tondoh, J.E.; Desta, L.T.; Gumbricht, T. Mapping of soil properties and land degradation risk in Africa using MODIS reflectance. Geoderma 2016, 263, 216–225. [Google Scholar] [CrossRef]
- Cooper, P.; Dimes, J.; Rao, K.; Shapiro, B.; Shiferaw, B.; Twomlow, S. Coping better with current climatic variability in the rain-fed farming systems of sub-Saharan Africa: An essential first step in adapting to future climate change? Agric. Ecosyst. Environ. 2008, 126, 24–35. [Google Scholar] [CrossRef]
- Thornton, P.K.; Jones, P.G.; Ericksen, P.J.; Challinor, A.J. Agriculture and food systems in sub-Saharan Africa in a 4 C+ world. Philos. Trans. R. Soc. Lond. A Math. Phys. Eng. Sci. 2011, 69, 117–136. [Google Scholar] [CrossRef]
- Legesse, W.; Mosisa, W.; Berhanu, T.; Girum, A.; Wende, A.; Solomon, A.; Tolera, K.; Dagne, W.; Girma, D.; Temesgen, C.; et al. Genetic improvement of maize for mid-altitude and lowland sub-humid agro-ecologies of Ethiopia. In Meeting the Challenges of Global Climate Change and Food Security through Innovative Maize Research; CIMMYT: Mexico City, Mexico, 2011; p. 24. [Google Scholar]
- Abate, T.; Shiferaw, B.; Menkir, A.; Wegary, D.; Kebede, Y.; Tesfaye, K.; Kassie, M.; Bogale, G.; Tadesse, B.; Keno, T. Factors that transformed maize productivity in Ethiopia. Food Secur. 2015, 7, 965–981. [Google Scholar] [CrossRef]
- Dilla, A.M.; Smethurst, P.J.; Barry, K.; Parsons, D.; Denboba, M.A. Tree pruning, zone and fertiliser interactions determine maize productivity in the Faidherbia albida (Delile) A. Chev parkland agroforestry system of Ethiopia. Agrofor. Syst. 2019, 93, 1897–1907. [Google Scholar] [CrossRef]
- Snapp, S.S.; Grabowski, P.; Chikowo, R.; Smith, A.; Anders, E.; Sirrine, D.; Chimonyo, V.; Bekunda, M. Maize yield and profitability tradeoffs with social, human and environmental performance: Is sustainable intensification feasible? Agric. Syst. 2018, 162, 77–88. [Google Scholar] [CrossRef]
- Lin, B.B. Agroforestry management as an adaptive strategy against potential microclimate extremes in coffee agriculture. Agric. For. Meteorol. 2007, 144, 85–94. [Google Scholar] [CrossRef]
- Sileshi, G.W.; Nyoka, B.I.; Beedy, T.L.; Chanyenga, T. Modelling the scaling of stem growth with crown size and optimum stocking densities for systematic grid plantation of Faidherbia albida. New For. 2014, 45, 699–714. [Google Scholar] [CrossRef]
- Bayala, J.; Sanou, J.; Teklehaimanot, Z.; Ouedraogo, S.; Kalinganire, A.; Coe, R.; Van Noordwijk, M. Advances in knowledge of processes in soil–tree–crop interactions in parkland systems in the West African Sahel: A review. Agric. Ecosyst. Environ. 2015, 205, 25–35. [Google Scholar] [CrossRef]
- García-Barrios, L.; Ong, C. Ecological interactions, management lessons and design tools in tropical agroforestry systems. In New Vistas in Agroforestry; Springer: Berlin/Heidelberg, Germany, 2004; pp. 221–236. [Google Scholar]
- Semwal, R.; Maikhuri, R.; Rao, K.; Singh, K.; Saxena, K. Crop productivity under differently lopped canopies of multipurpose trees in Central Himalaya, India. Agrofor. Syst. 2002, 56, 57–63. [Google Scholar] [CrossRef]
- Siriri, D.; Ong, C.; Wilson, J.; Boffa, J.; Black, C. Tree species and pruning regime affect crop yield on bench terraces in SW Uganda. Agrofor. Syst. 2010, 78, 65. [Google Scholar] [CrossRef]
- Demeke, A.B.; Keil, A.; Zeller, M. Using panel data to estimate the effect ofrainfall shocks on smallholders food security and vulnerability in rural Ethiopia. Clim. Chang. 2011, 108, 185–206. [Google Scholar] [CrossRef]
- Kassie, B.; Van Ittersum, M.; Hengsdijk, H.; Asseng, S.; Wolf, J.; Rötter, R.P. Climate-induced yield variability and yield gaps of maize (Zea mays L.) in the Central Rift Valley of Ethiopia. Field Crop Res. 2014, 160, 41–53. [Google Scholar] [CrossRef]
- Keating, B.A.; Carberry, P.S.; Hammer, G.L.; Probert, M.E.; Robertson, M.J.; Holzworth, D.; Huth, N.I.; Hargreaves, J.N.; Meinke, H.; Hochman, Z. An overview of APSIM, a model designed for farming systems simulation. Eur. J. Agron. 2003, 18, 267–288. [Google Scholar] [CrossRef]
- Holzworth, D.; Huth, N.I.; Fainges, J.; Brown, H.; Zurcher, E.; Cichota, R.; Verrall, S.; Herrmann, N.I.; Zheng, B.; Snow, V. APSIM Next Generation: Overcoming challenges in modernising a farming systems model. Environ. Model. Softw. 2018, 103, 43–51. [Google Scholar] [CrossRef]
- Smethurst, P.J.; Huth, N.I.; Masikati, P.; Sileshi, G.W.; Akinnifesi, F.K.; Wilson, J.; Sinclair, F. Accurate crop yield predictions from modelling tree-crop interactions in gliricidia-maize agroforestry. Agric. Syst. 2017, 155, 70–77. [Google Scholar] [CrossRef]
- Dilla, A.; Smethurst, P.J.; Parsons, D.; Barry, K.; Denboba, M. Potential of the APSIM model to simulate impacts of shading on maize productivity. Agrofor. Syst. 2018, 92, 1699–1709. [Google Scholar] [CrossRef]
- Knörzer, H.; Lawes, R.; Robertson, M.; Graeff-Hönninger, S.; Claupein, W. Evaluation and performance of the APSIM crop growth model for German winter wheat, maize and fieldpea varieties in monocropping and intercropping systems. J. Agric. Sci. Technol. B 2011, 9, 698–717. [Google Scholar]
- Dufour, L.; Metay, A.; Talbot, G.; Dupraz, C. Assessing light competition for cereal production in temperate agroforestry systems using experimentation and crop modelling. J. Agron. Crop Sci. 2013, 199, 217–227. [Google Scholar] [CrossRef]
- Cooper, M.; Messina, C.D.; Podlich, D.; Totir, L.R.; Baumgarten, A.; Hausmann, N.J.; Wright, D.; Graham, G. Predicting the future of plant breeding: Complementing empirical evaluation with genetic prediction. Crop Pasture Sci. 2014, 65, 311–336. [Google Scholar] [CrossRef]
- Seyoum, S.; Rachaputi, R.; Chauhan, Y.; Prasanna, B.; Fekybelu, S. Application of the APSIM model to exploit G× E× M interactions for maize improvement in Ethiopia. Field Crop Res. 2018, 217, 113–124. [Google Scholar] [CrossRef]
- Dilla, A.M.; Smethurst, P.J.; Barry, K.; Parsons, D. Preliminary estimate of carbon sequestration potential of Faidherbia albida (Delile) A. Chev in an agroforestry parkland in the Central Rift Valley of Ethiopia. For. Trees Livelihoods 2019, 28, 79–89. [Google Scholar] [CrossRef]
- Goma, M. Characterization of Climate Variability and Water Harvesting System for Crop Production in Adulala Watershed, Central Rift Valley of Ethiopia; Haramaya University: Haramaya, Oromia, Ethiopia, 2015. [Google Scholar]
- Argaw, M.; Teketay, D.; Olsson, M. Soil seed flora, germination and regeneration pattern of woody species in an Acacia woodland of the Rift Valley in Ethiopia. J. Arid Environ. 1999, 43, 411–435. [Google Scholar] [CrossRef]
- Endale, Y.; Derero, A.; Argaw, M.; Muthuri, C. Farmland tree species diversity and spatial distribution pattern in semi-arid East Shewa, Ethiopia. For. Trees Livelihoods 2017, 26, 199–214. [Google Scholar] [CrossRef]
- Araya, A.; Girma, A.; Getachew, F. Exploring impacts of climate change on maize yield in two contrasting agro-ecologies of Ethiopia. Asian J. Appl. Sci. Eng. 2015, 4, 26–36. [Google Scholar]
- Dawson, I.K.; Lengkeek, A.; Weber, J.C.; Jamnadass, R. Managing genetic variation in tropical trees: Linking knowledge with action in agroforestry ecosystems for improved conservation and enhanced livelihoods. Biodivers. Conserv. 2009, 18, 969–986. [Google Scholar] [CrossRef]
- Moriasi, D.N.; Arnold, J.G.; Liew, M.W.V.; Bingner, R.L.; Harmel, R.D.; Veith, T.L. Model evaluation guidelines for systematic quantification of accuracy in watershed simulations. Trans. ASABE 2007, 50, 885–900. [Google Scholar] [CrossRef]
- Yang, J.M.; Yang, J.Y.; Liu, S.; Hoogenboom, G. An evaluation of the statistical methods for testing the performance of crop models with observed data. Agric. Syst. 2014, 127, 81–89. [Google Scholar] [CrossRef]
- Ritter, A.; Muñoz-Carpena, R. Performance evaluation of hydrological models: Statistical significance for reducing subjectivity in goodness-of-fit assessments. J. Hydrol. 2013, 480, 33–45. [Google Scholar] [CrossRef]
- MacCarthy, D.S.; Sommer, R.; Vlek, P.L. Modeling the impacts of contrasting nutrient and residue management practices on grain yield of sorghum (Sorghum bicolor (L.) Moench) in a semi-arid region of Ghana using APSIM. Field Crop Res. 2009, 113, 105–115. [Google Scholar] [CrossRef]
- Robertson, M.; Sakala, W.; Benson, T.; Shamudzarira, Z. Simulating response of maize to previous velvet bean (Mucuna pruriens) crop and nitrogen fertiliser in Malawi. Field Crop Res. 2005, 91, 91–105. [Google Scholar] [CrossRef]
- Sida, T.S.; Baudron, F.; Kim, H.; Giller, K.E. Climate-smart agroforestry: Faidherbia albida trees buffer wheat against climatic extremes in the Central Rift Valley of Ethiopia. Agric. For. Meteorol. 2018, 248, 339–347. [Google Scholar] [CrossRef]
- Ndoli, A.; Baudron, F.; Schut, A.G.; Mukuralinda, A.; Giller, K.E. Disentangling the positive and negative effects of trees on maize performance in smallholdings of Northern Rwanda. Field Crop Res. 2017, 213, 1–11. [Google Scholar] [CrossRef]
- Kho, R.; Yacouba, B.; Yayé, M.; Katkoré, B.; Moussa, A.; Iktam, A.; Mayaki, A. Separating the effects of trees on crops: The case of Faidherbia albida and millet in Niger. Agrofor. Syst. 2001, 52, 219–238. [Google Scholar] [CrossRef]
- Sileshi, G.W. The magnitude and spatial extent of influence of Faidherbia albida trees on soil properties and primary productivity in drylands. J. Arid Environ. 2016, 132, 1–14. [Google Scholar] [CrossRef]
- Tiwari, T.P.; Brook, R.M.; Wagstaff, P.; Sinclair, F.L. Effects of light environment on maize in hillside agroforestry systems of Nepal. Food Secur. 2012, 4, 103–114. [Google Scholar] [CrossRef]
- Jose, S.; Gillespie, A.; Seifert, J.; Biehle, D. Defining competition vectors in a temperate alley cropping system in the midwestern USA: 2. Competition for water. Agrofor. Syst. 2000, 48, 41–59. [Google Scholar] [CrossRef]
- Pardon, P.; Reubens, B.; Reheul, D.; Mertens, J.; De Frenne, P.; Coussement, T.; Janssens, P.; Verheyen, K. Trees increase soil organic carbon and nutrient availability in temperate agroforestry systems. Agric. Ecosyst. Environ. 2017, 247, 98–111. [Google Scholar] [CrossRef]
- Mekonnen, K.; Amede, T.; Duncan, A.; Gebrekirstos, A. Sustainable agricultural intensification and its role on the climate resilient green economy initiative in Ethiopia. In Proceedings of the 3rd National Platform Meeting on Land and Water Management in Ethiopia, Addis Ababa, Ethiopia, 23–24 July 2012; International Livestock Research Institute: Nairobi, Kenya, 2013. [Google Scholar]
- Makumba, W.; Janssen, B.; Oenema, O.; Akinnifesi, F.K.; Mweta, D.; Kwesiga, F. The long-term effects of a gliricidia–maize intercropping system in Southern Malawi, on gliricidia and maize yields, and soil properties. Agric. Ecosyst. Environ. 2006, 116, 85–92. [Google Scholar] [CrossRef]
- Boffa, J.-M. Agroforestry Parklands in Sub-Saharan Africa; FAO: Rome, Italy, 1999. [Google Scholar]
- Rodriguez, D.; Cox, H.; Power, B. A participatory whole farm modelling approach to understand impacts and increase preparedness to climate change in Australia. Agric. Syst. 2014, 126, 50–61. [Google Scholar] [CrossRef]
- Yin, X.; Kersebaum, K.C.; Beaudoin, N.; Constantin, J.; Chen, F.; Louarn, G.; Manevski, K.; Hoffmann, M.; Kollas, C.; Armas-Herrera, C.M.; et al. Uncertainties in simulating N uptake, net N mineralization, soil mineral N and N leaching in European crop rotations using process-based models. Field Crop Res. 2020, 255, 107863. [Google Scholar] [CrossRef]
- Akinnifesi, F.K.; Chirwa, P.W.; Ajayi, O.C.; Sileshi, G.; Matakala, P.; Kwesiga, F.R.; Harawa, H.; Makumba, W. Contributions of agroforestry research to livelihood of smallholder farmers in Southern Africa: 1. Taking stock of the adaptation, adoption and impact of fertilizer tree options. Agric. J. 2008, 3, 58–75. [Google Scholar]
- Mafongoya, P.; Bationo, A.; Kihara, J.; Waswa, B.S. Appropriate technologies to replenish soil fertility in southern Africa. Nutr. Cycl. Agroecosyst. 2006, 76, 137–151. [Google Scholar] [CrossRef]
- Vanlauwe, B.; Aihou, K.; Aman, S.; Iwuafor, E.N.; Tossah, B.K.; Diels, J.; Sanginga, N.; Lyasse, O.; Merckx, R.; Deckers, J. Maize yield as affected by organic inputs and urea in the West African moist savannah. Agron. J. 2001, 93, 1191–1199. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pruning Level | Shade (%) Zone (m) | ||||
|---|---|---|---|---|---|
| 0–2 | 2–4 | 4–6 | 6–8 | Crop-Only | |
| Unpruned | 46.3 | 41.7 | 27.4 | 0 | 0 |
| 50% Pruned | 31 | 24 | 14 | 0 | 0 |
| 100% Pruned | 17 | 9 | 9 | 0 | 0 |
| Depth (cm) | Zone (m) | ||||
|---|---|---|---|---|---|
| 0–2 | 2–4 | 4–6 | 6–8 | Crop-Only | |
| 0–20 | 0.51 | 0.44 | 0.24 | 0.10 | 0 |
| 20–40 | 0.48 | 0.30 | 0.23 | 0.14 | 0 |
| 40–60 | 0.16 | 0.06 | 0.14 | 0.02 | 0 |
| 60–80 | 0.06 | 0.03 | 0.06 | 0.01 | 0 |
| 80–120 | 0.03 | 0.01 | 0.01 | 0.01 | 0 |
| 120–200 | 0.02 | 0.01 | 0.01 | 0.01 | 0 |
| 200–350 | 0.01 | 0.01 | 0.01 | 0.01 | 0 |
| Zone | Depth | BD 1,2 | OC | NO3 | NH4 | Fbiom | Finert | DUL | LLmaize | KLmaize | PAWCmaize |
|---|---|---|---|---|---|---|---|---|---|---|---|
| (g cm−3) | (%) | (µg g−1) | (µg g−1) | (mm mm−1) | (d) | (mm mm−1) | |||||
| 0–2 m | 0–20 | 1.150 | 1.700 | 2.174 | 0.870 | 0.020 | 0.650 | 0.350 | 0.160 | 0.005 | 0.190 |
| 20–40 | 1.150 | 1.400 | 0.870 | 0.435 | 0.020 | 0.700 | 0.350 | 0.170 | 0.004 | 0.180 | |
| 40–60 | 1.340 | 1.300 | 0.373 | 0.187 | 0.010 | 0.800 | 0.350 | 0.200 | 0.004 | 0.150 | |
| 60–80 | 1.340 | 1.200 | 0.187 | 0.000 | 0.010 | 0.900 | 0.350 | 0.210 | 0.004 | 0.140 | |
| 2–4 m | 0–20 | 1.150 | 1.48 | 2.174 | 0.870 | 0.020 | 0.650 | 0.350 | 0.160 | 0.006 | 0.190 |
| 20–40 | 1.150 | 1.46 | 0.870 | 0.435 | 0.020 | 0.700 | 0.350 | 0.170 | 0.005 | 0.180 | |
| 40–60 | 1.340 | 1.42 | 0.373 | 0.187 | 0.010 | 0.800 | 0.350 | 0.200 | 0.004 | 0.150 | |
| 60–80 | 1.340 | 1.35 | 0.187 | 0.000 | 0.010 | 0.900 | 0.350 | 0.210 | 0.003 | 0.140 | |
| 4–6 m | 0–20 | 1.150 | 1.33 | 2.174 | 0.870 | 0.020 | 0.650 | 0.350 | 0.160 | 0.005 | 0.190 |
| 20–40 | 1.150 | 1.12 | 0.870 | 0.435 | 0.020 | 0.700 | 0.350 | 0.170 | 0.004 | 0.180 | |
| 40–60 | 1.340 | 0.90 | 0.373 | 0.187 | 0.010 | 0.800 | 0.350 | 0.200 | 0.004 | 0.150 | |
| 60–80 | 1.340 | 0.76 | 0.187 | 0.000 | 0.010 | 0.900 | 0.350 | 0.210 | 0.004 | 0.140 | |
| Crop-only | 0–20 | 1.150 | 1.07 | 2.174 | 0.870 | 0.010 | 0.600 | 0.230 | 0.180 | 0.005 | 0.050 |
| 20–40 | 1.150 | 0.86 | 0.870 | 0.435 | 0.010 | 0.650 | 0.260 | 0.190 | 0.004 | 0.070 | |
| 40–60 | 1.340 | 0.79 | 0.373 | 0.187 | 0.001 | 0.650 | 0.297 | 0.220 | 0.004 | 0.077 | |
| 60–80 | 1.340 | 0.70 | 0.187 | 0.000 | 0.000 | 0.750 | 0.330 | 0.230 | 0.004 | 0.100 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dilla, A.M.; Smethurst, P.J.; Huth, N.I.; Barry, K.M. Plot-Scale Agroforestry Modeling Explores Tree Pruning and Fertilizer Interactions for Maize Production in a Faidherbia Parkland. Forests 2020, 11, 1175. https://doi.org/10.3390/f11111175
Dilla AM, Smethurst PJ, Huth NI, Barry KM. Plot-Scale Agroforestry Modeling Explores Tree Pruning and Fertilizer Interactions for Maize Production in a Faidherbia Parkland. Forests. 2020; 11(11):1175. https://doi.org/10.3390/f11111175
Chicago/Turabian StyleDilla, Aynalem M., Philip J. Smethurst, Neil I. Huth, and Karen M. Barry. 2020. "Plot-Scale Agroforestry Modeling Explores Tree Pruning and Fertilizer Interactions for Maize Production in a Faidherbia Parkland" Forests 11, no. 11: 1175. https://doi.org/10.3390/f11111175
APA StyleDilla, A. M., Smethurst, P. J., Huth, N. I., & Barry, K. M. (2020). Plot-Scale Agroforestry Modeling Explores Tree Pruning and Fertilizer Interactions for Maize Production in a Faidherbia Parkland. Forests, 11(11), 1175. https://doi.org/10.3390/f11111175