The Stability of Mean Wood Specific Gravity across Stand Age in US Forests Despite Species Turnover


Abstract
1. Introduction
- (1)
- (2)
- BIOMASS (a European Space Agency Earth Explorer mission) uses P-band radar to map AGB at 200m spatial resolution [6];
- (3)
- NISAR (joint mission between NASA and the Indian Space Research Organisation) operates an L-band and an S-band synthetic aperture radar that enables observation of biomass change at hectare scales [7]; and,
- (4)
- ICESAT-2 (NASA) uses linear tracks of pulse-counting lidar that, in combination with other instruments, allows measurement of vegetation biomass [8].
2. Methods
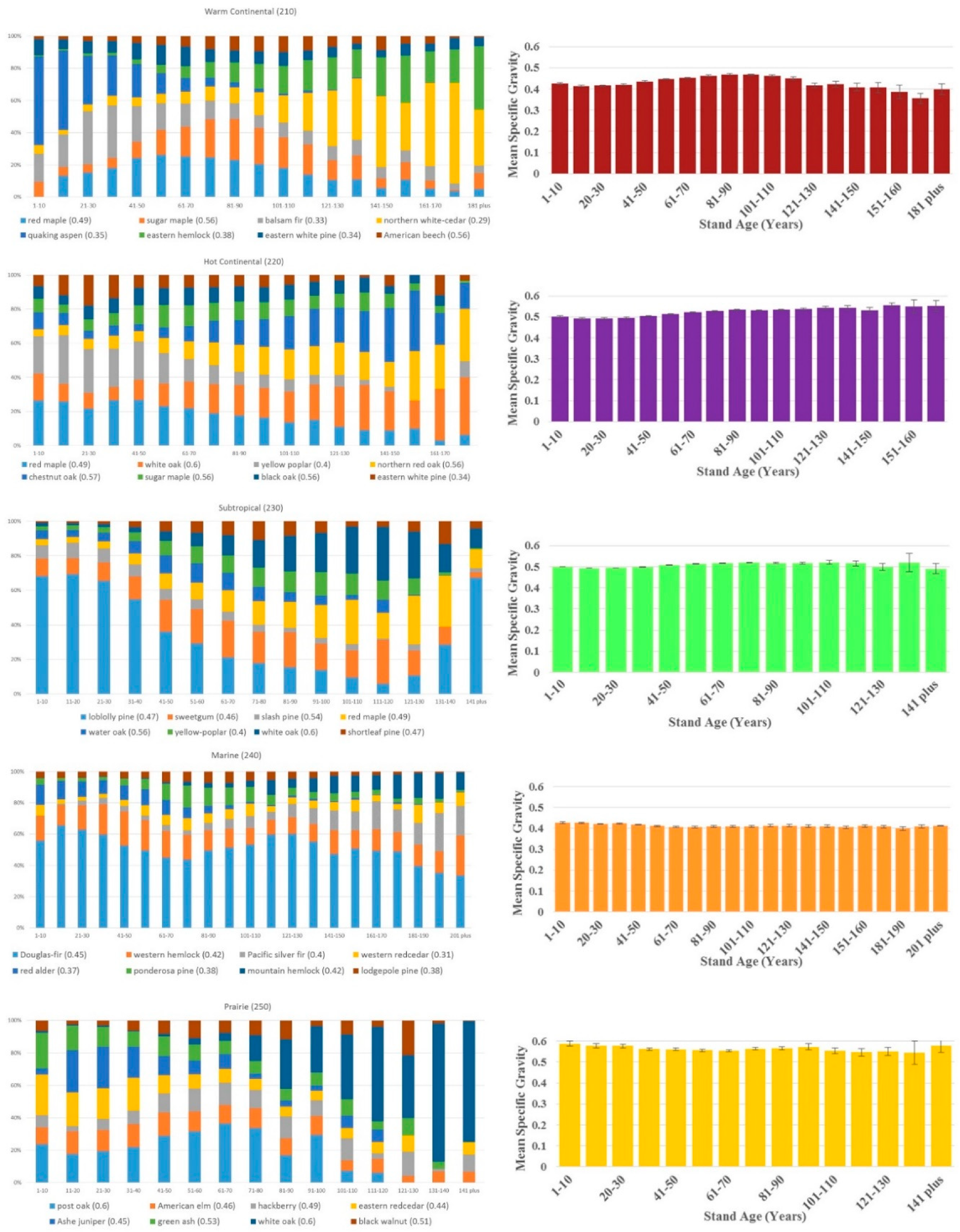
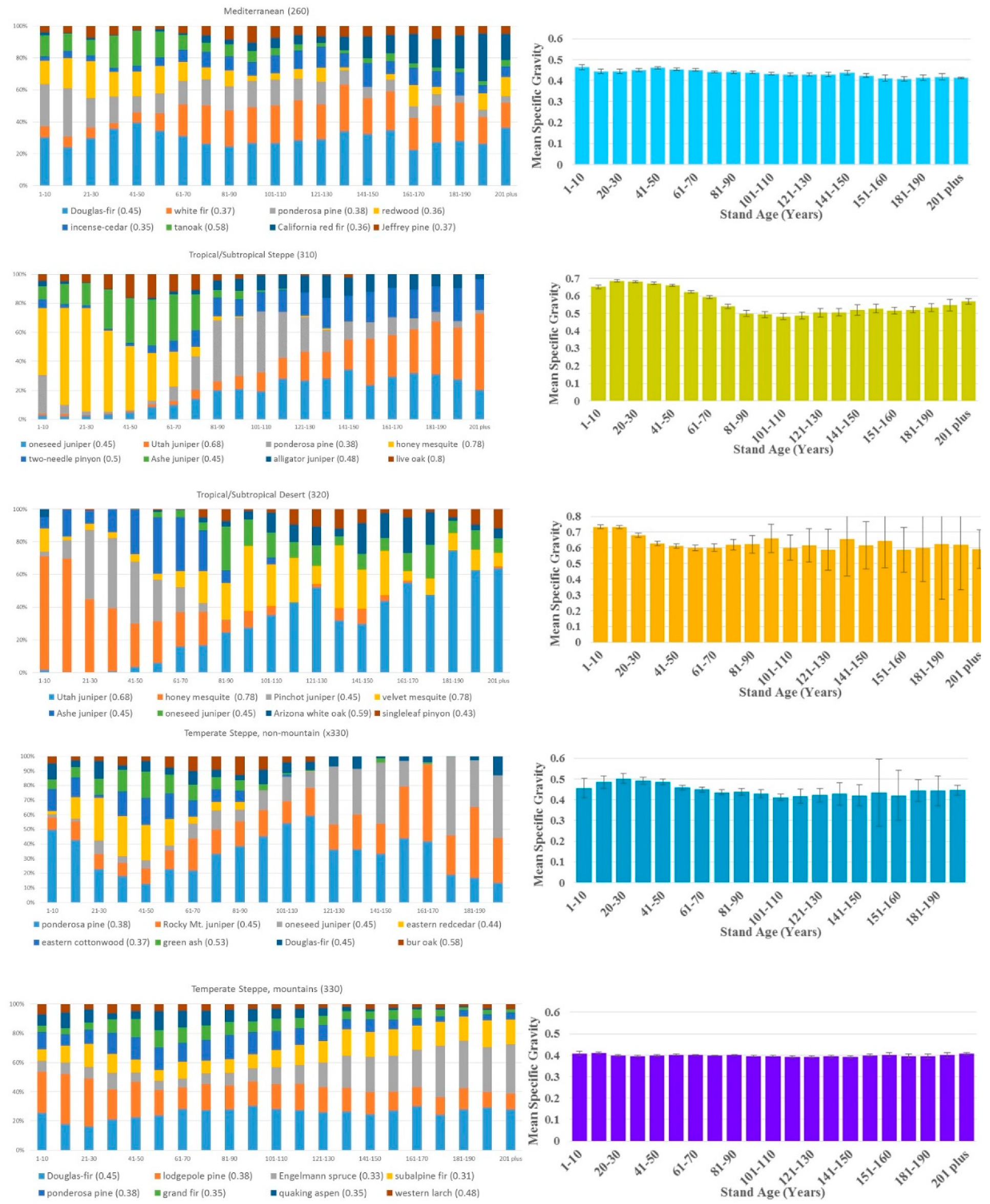
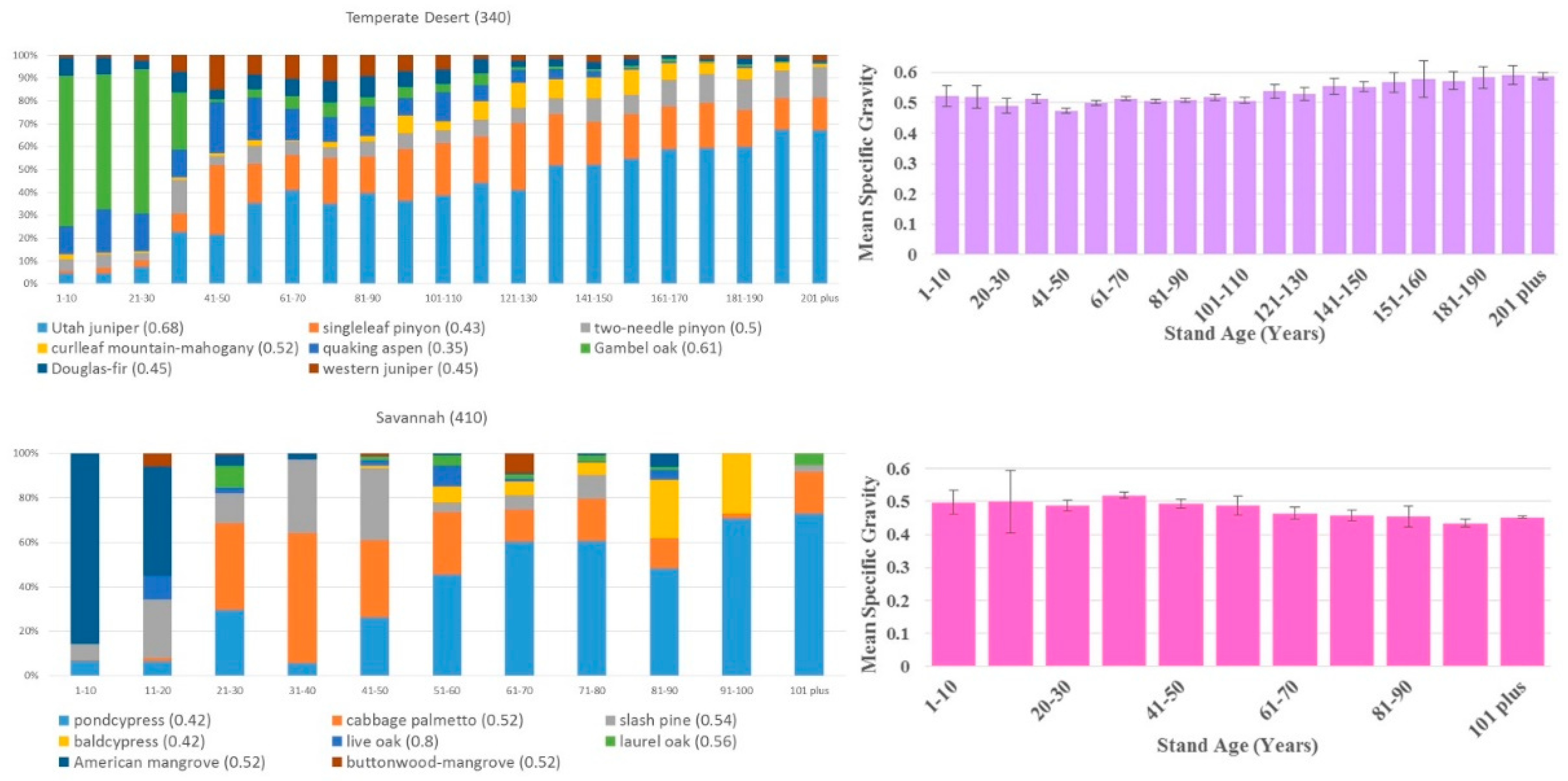
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A. Scientific Names for Cited Tree Species
| Common Name | Latin binomial | Common Name | Latin binomial |
| Pacific silver fir | Abies amabilis (Dougl. ex Louden) | lodgepole pine | Pinus contorta Douglas ex Loud.) |
| balsam fir | Abies balsamea (L.) Mill. | two-needle pinyon | Pinus edulis Engelm. |
| white fir | Abies concolor (Gord. & Glend.) Lindl. ex Hildebr. | slash pine | Pinus elliottii Engelm. |
| grand fir | Abies grandis (Dougl. ex D. Don.) Lindl. | Jeffrey pine | Pinus jeffreyi Balf. |
| subalpine fir | Abies lasiocarpa (Hook.) Nutt. | singleleaf pinyon | Pinus monophylla Torr. & Frém. |
| California red fir | Abies magnifica A. Murr. | ponderosa pine | Pinus ponderosa C. Lawson |
| bigleaf maple | Acer macrophyllum Pursh | eastern white pine | Pinus strobus L. |
| red maple | Acer rubrum L. | loblolly pine | Pinus taeda L. |
| sugar maple | Acer saccharum L. | eastern cottonwood | Populus deltoids Bartram ex Marsh. |
| red alder | Alunus rubra Bong. | quaking aspen | Populus tremuloides Michx. |
| yellow birch | Betula alleghaniensis Britton | honey mesquite | Prosopis glandulosa Torr. |
| incense cedar | Calocedrus decurrens (Torr.) Florin | velvet mesquite | Prosopis velutina Woot. |
| hackberry | Celtis occidentalis L. | black cherry | Prunus serotine Ehrh. |
| mountain-mahogany | Cercocarpus ledifolius Nutt. | Douglas-fir | Pseudotsuga menziesii |
| buttonwood-mangrove | Conocarpus erectus L. | white oak | Quercus alba L. |
| American beech | Fagus grandifolia Ehrh. | Arizona white oak | Quercus arizonica Sarg. |
| white ash | Fraxinus americana L. | canyon live oak | Quercus chrysolepis Liebm. |
| green ash | Fraxinus pennsylvanica Marsh. | southern red oak | Quercus falcata Michx. |
| black walnut | Juglans nigra L. | Gambel oak | Quercus gambelii Nutt. |
| Ashe juniper | Juniperus ashei J. Buchholz | laurel oak | Quercus laurifolia Michx. |
| alligator juniper | Juniperus deppeana Steud. | bur oak | Quercus macrocarpa Michx. |
| redberry juniper | Juniperus coahuilensis (Martiñez) Gausen ex R.P. Adams | chestnut oak | Quercus montana Willd. |
| oneseed juniper | Juniperus monosperma (Engelm.) Sarg. | water oak | Quercus nigra L. |
| western juniper | Juniperus occidentalis Hook. | northern red oak | Quercus rubra L. |
| Utah juniper | Juniperus osteosperma (Torr.) Little | post oak | Quercus stellate Wangenh. |
| Pinchot juniper | Juniperus pinchotii Sudworgh | black oak | Quercus velutina Lam. |
| Rocky Mountain juniper | Juniperus scopulorum Sarg. | live oak | Quercus virginiana Mill. |
| eastern redcedar | Juniperus virginiana L. | American mangrove | Rhizophora mangle L. |
| western larch | Larix occidentalis Nutt. | cabbage palmetto | Sabal palmetto (Walter) Lodd. ex Schult. & Schult. f. |
| sweetgum | Liquidambar styraciflua L. | redwood | Sequoia sempervirens (Lamb. ex D. Don.) Endl. |
| yellow poplar | Liriodendron tulipifera L. | pond cypress | Taxodium ascendens Brongn. |
| tanoak | Lithocarpus densiflorus (Hook. & Arn.) Rehd. | baldcypress | Taxodium distichum (L.) Rich. |
| melaleuca | Melaleuca quinquenervia (Cav.) S.F. Blake | northern white-cedar | Thuja occidentalis L. |
| swamp tupelo | Nyssa biflora Walter | western redcedar | Thuja plicata) Donn ex D. Don |
| redbay | Persea borbonia (L.) Spreng. | eastern hemlock | Tsuga Canadensis L. |
| Engelmann spruce | Picea engelmannii Parry ex Engelm. | western hemlock | Tsuga heterophylla (Raf.) Sarg. |
| red spruce | Picea rubens Sarg. | mountain hemlock | Tsuga mertensiana (Bong.) Carrière |
| shortleaf pine | Pinus echinata Mill. | American elm | Ulmus Americana L. |
References
- Houghton, R.A. Aboveground forest biomass and the global carbon balance. Glob. Chang. Biol. 2005, 11, 945–958. [Google Scholar] [CrossRef]
- Woodall, C.W.; Coulston, J.W.; Domke, G.M.; Walters, B.F.; Wear, D.N.; Smith, J.E.; Andersen, H.-E.; Clough, B.J.; Cohen, W.B.; Griffith, D.M.; et al. The U.S. Forest Carbon Accounting Framework: Stocks and Stock Change, 1990–2016; Gen. Tech. Rep. NRS-154; USDA Forest Service Northern Research Station: Newtown Square, PA, USA, 2016.
- Dubayah, R.; Goetz, S.J.; Blair, J.B.; Fatoyinbo, T.E.; Hansen, M.; Healey, S.P.; Hofton, M.A.; Hurtt, G.C.; Kellner, J.; Luthcke, S.B.; et al. The global ecosystem dynamics investigation. In AGU Fall Meeting Abstracts; American Geophysical Union: San Francisco, CA, USA, 2014. [Google Scholar]
- Saarela, S.; Holm, S.; Healey, S.P.; Andersen, H.-E.; Petersson, H.; Prentius, W.; Patterson, P.L.; Naesset, E.; Gregoire, T.E.; Ståhl, G. Generalized Hierarchical Model-Based Estimation for Aboveground Biomass Assessment Using GEDI and Landsat Data. Remote Sens. 2018, 10, 1832. [Google Scholar] [CrossRef]
- Patterson, P.L.; Healey, S.P.; Ståhl, G.; Saarela, S.; Holm, S.; Anderson, H.-E.; Dubayah, R.O.; Duncanson, L.; Hancock, S.; Armston, J.; et al. Statistical properties of hybrid estimators proposed for GEDI—NASA’s Global Ecosystem Dynamics Investigation. Environ. Res. Lett. 2019. in review. [Google Scholar]
- Carreiras, J.M.; Quegan, S.; Le Toan, T.; Minh, D.H.; Saatchi, S.S.; Carvalhais, N.; Reichstein, M.; Scipal, K. Coverage of high biomass forests by the ESA BIOMASS mission under defense restrictions. Remote Sens. Environ. 2017, 196, 154–162. [Google Scholar] [CrossRef]
- Yu, Y.; Saatchi, S. Sensitivity of L-Band SAR Backscatter to Aboveground Biomass of Global Forests. Remote Sens. 2016, 8, 522. [Google Scholar] [CrossRef]
- Glenn, N.F.; Neuenschwander, A.; Vierling, L.A.; Spaete, L.; Li, A.; Shinneman, D.J.; Pilliod, D.S.; Arkle, R.S.; McIlroy, S.K. Landsat 8 and ICESat-2: Performance and potential synergies for quantifying dryland ecosystem vegetation cover and biomass. Remote Sens. Environ. 2016, 185, 233–242. [Google Scholar] [CrossRef]
- Healey, S.P.; Patterson, P.L.; Saatchi, S.S.; Lefsky, M.A.; Lister, A.J.; Freeman, E.A. A sample design for globally consistent biomass estimation using lidar data from the Geoscience Laser Altimeter System (GLAS). Carbon Balance Manag. 2012, 7, 9. [Google Scholar] [CrossRef]
- Babcock, C.; Finley, A.O.; Cook, B.D.; Weiskittel, A.; Woodall, C.W. Modeling forest biomass and growth: Coupling long-term inventory and LiDAR data. Remote Sens. Environ. 2016, 182, 1–12. [Google Scholar] [CrossRef]
- Deo, R.K.; Domke, G.M.; Russell, M.B.; Woodall, C.W.; Andersen, H.E. Evaluating the influence of spatial resolution of Landsat predictors on the accuracy of biomass models for large-area estimation across the eastern USA. Environ. Res. Lett. 2018, 13, 055004. [Google Scholar] [CrossRef]
- Asner, G.P.; Mascaro, J. Mapping tropical forest carbon: Calibrating plot estimates to a simple LiDAR metric. Remote Sens. Environ. 2014, 140, 614–624. [Google Scholar] [CrossRef]
- Enquist, B.J.; West, G.B.; Charnov, E.L.; Brown, J.H. Allometric scaling of production and life-history variation in vascular plants. Nature 1999, 401, 907–911. [Google Scholar] [CrossRef]
- Poorter, L.; McDonald, I.; Alarcón, A.; Fichtler, E.; Licona, J.-C.; Peña-Claros, M.; Sterck, F.; Villegas, Z.; Sass-Klaassen, U. The importance of wood traits and hydraulic conductance for the performance and life history strategies of 42 rainforest tree species. New Phytol. 2010, 185, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Law, B.E.; Sun, O.J.; Campbell, J.; Van Tuyl, S.; Thornton, P.E. Changes in carbon storage and fluxes in a chronosequence of ponderosa pine. Glob. Chang. Biol. 2003, 9, 510–524. [Google Scholar] [CrossRef]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-resolution global maps of 21st-century forest cover change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Healey, S.P.; Cohen, W.B.; Yang, Z.; Brewer, C.K.; Brooks, E.B.; Gorelick, N.; Hernandez, A.J.; Huang, C.; Hughes, M.K.; Kennedy, R.E.; et al. Mapping forest change using stacked generalization: An ensemble approach. Remote Sens. Environ. 2018, 204, 717–728. [Google Scholar] [CrossRef]
- Domke, G.M.; Perry, C.H.; Walters, B.F.; Nave, L.E.; Woodall, C.W.; Swanston, C.W. Toward inventory-based estimates of soil organic carbon in forests of the United States. Ecol. Appl. 2017, 27, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Duncanson, L.I.; Dubayah, R.O.; Enquist, B.J. Assessing the general patterns of forest structure: Quantifying tree and forest allometric scaling relationships in the United States. Glob. Ecol. Biogeogr. 2015, 24, 1465–1475. [Google Scholar] [CrossRef]
- Scott, C.T.; Bechtold, W.A.; Reams, G.A.; Smith, W.D.; Westfall, J.A.; Hansen, M.H.; Moisen, G.G. Sample-Based Estimators Used by the Forest Inventory and Analysis National Information Management System; Gen. Tech. Rep. SRS-80; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2005; pp. 53–77.
- Cleland, D.T.; Freeouf, J.A.; Keyes, J.E.; Nowacki, G.J.; Carpenter, C.A.; McNabb, W.H. Ecological Subregions: Sections and Subsections for the Conterminous United States; Gen. Tech. Report WO-79D [Map on CD-ROM] (A.M. Sloan, cartographer); U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 2007.
- Bechtold, W.A.; Scott, C.T. The Forest Inventory and Analysis Plot Design; Gen. Tech. Rep. SRS-80; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2005; pp. 37–52.
- Miles, P.D.; Smith, W.B. Specific Gravity and Other Properties of Wood and Bark for 156 Tree Species Found in North America; Res. Note NRS-38; U.S. Department of Agriculture, Forest Service: Newtown Square, PA, USA, 2009.
- United States Department of Agriculture. Interior West Forest Inventory and Analysis P2 Field Procedures—V7. USDA Forest Service. Available online: https://www.fs.fed.us/rm/ogden/data-collection/pdf/P2%20Manual_70_Feb2sm.pdf (accessed on 20 November 2018).
- Williams, C.A.; Collatz, G.J.; Masek, J.; Huang, C.; Goward, S.N. Impacts of disturbance history on forest carbon stocks and fluxes: Merging satellite disturbance mapping with forest inventory data in a carbon cycle model framework. Remote Sens. Environ. 2014, 151, 57–71. [Google Scholar] [CrossRef]
- Fisher, R.A.; Koven, C.D.; Anderegg, W.R.L.; Christoffersen, B.O.; Dietze, M.C.; Farrior, C.E.; Holm, J.A.; Hurtt, G.C.; Knox, R.G.; Lawrence, P.J.; et al. Vegetation demographics in Earth System Models: A review of progress and priorities. Glob. Chang. Biol. 2018, 24, 35–54. [Google Scholar] [CrossRef]
- Clark, D.B.; Kellner, J.R. Tropical forest biomass estimation and the fallacy of misplaced concreteness. J. Veg. Sci. 2012, 23, 1191–1196. [Google Scholar] [CrossRef]
- Beets, P.N.; Gilchrist, K.; Jeffreys, M.P. Wood density of radiata pine: Effect of nitrogen supply. For. Ecol. Manag. 2001, 145, 173–180. [Google Scholar] [CrossRef]
- Lasserre, J.P.; Mason, E.G.; Watt, M.S.; Moore, J.R. Influence of initial planting spacing and genotype on microfibril angle, wood density, fibre properties and modulus of elasticity in Pinus radiata D. Don corewood. For. Ecol. Manag. 2009, 258, 1924–1931. [Google Scholar] [CrossRef]
- Jaakkola, T.; Mäkinen, H.; Saranpää, P. Wood density in Norway spruce: Changes with thinning intensity and tree age. Can. J. For. Res. 2005, 35, 1767–1778. [Google Scholar] [CrossRef]
- Lemay, A.; Krause, C.; Achim, A. Comparison of wood density in roots and stems of black spruce before and after commercial thinning. For. Ecol. Manag. 2018, 408, 94–102. [Google Scholar] [CrossRef]
- Muller-Landau, H.C. Interspecific and inter-site variation in wood specific gravity of tropical trees. Biotropica 2004, 36, 20–32. [Google Scholar] [CrossRef]
- Swenson, N.G.; Enquist, B.J. Ecological and evolutionary determinants of a key plant functional trait: Wood density and its community-wide variation across latitude and elevation. Am. J. Bot. 2007, 94, 451–459. [Google Scholar] [CrossRef]
- Compton, J.E.; Church, M.R.; Larned, S.T.; Hogsett, W.E. Nitrogen export from forested watersheds in the Oregon Coast Range: The role of N 2-fixing red alder. Ecosystems 2003, 6, 773–785. [Google Scholar] [CrossRef]
- Schultz, R.P. Loblolly Pine: The Ecology and Culture of Loblolly Pine (Pinus taeda L.); Agriculture Handbook 713; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1997; 493p.
- Mesquita, R.C.; Ickes, K.; Ganade, G.; Williamson, G.B. Alternative successional pathways in the Amazon Basin. J. Ecol. 2001, 89, 528–537. [Google Scholar] [CrossRef]
- Finegan, B. Pattern and process in neotropical secondary rain forests: The first 100 years of succession. Trends Ecol. Evol. 1996, 11, 119–124. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Division | FIA Estimate of Forestland (million ha) | Measured Trees/Seedlings | Measured Plots | Number of Surveyed Species | Mean Specific Gravity Estimate |
|---|---|---|---|---|---|
| Warm Continental (210) | 35.6 | 733,108/821,979 | 21,196 | 177 | 0.45 |
| Hot Continental (220) | 47.8 | 617,470/571,091 | 25,031 | 194 | 0.52 |
| Subtropical (230) | 67.9 | 919,398/551,384 | 32,217 | 204 | 0.50 |
| Marine (240) | 13.8 | 311,115/149,696 | 9371 | 63 | 0.42 |
| Prairie (250) | 10.9 | 81,551/72,208 | 4883 | 156 | 0.56 |
| Mediterranean (260) | 14.2 | 164,136/65,526 | 5315 | 78 | 0.44 |
| Tropical/Subtropical Steppe (310) | 27.9 | 153,967/77,679 | 9888 | 188 | 0.59 |
| Tropical/Subtropical Desert (320) | 6.5 | 18,572/8268 | 2168 | 73 | 0.65 |
| Temperate Steppe (330) without Mountain Regime | 4.5 | 27,468/23,904 | 1823 | 93 | 0.45 |
| Temperate Steppe (330) just Mountain Regime | 35.2 | 417,743/292,708 | 15,411 | 63 | 0.4 |
| Temperate Desert (340) | 12.5 | 99,703/56,268 | 5140 | 41 | 0.54 |
| Savannah (410) | 0.4 | 4034/5025 | 139 | 37 | 0.48 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Healey, S.P.; Menlove, J. The Stability of Mean Wood Specific Gravity across Stand Age in US Forests Despite Species Turnover. Forests 2019, 10, 114. https://doi.org/10.3390/f10020114
Healey SP, Menlove J. The Stability of Mean Wood Specific Gravity across Stand Age in US Forests Despite Species Turnover. Forests. 2019; 10(2):114. https://doi.org/10.3390/f10020114
Chicago/Turabian StyleHealey, Sean P., and James Menlove. 2019. "The Stability of Mean Wood Specific Gravity across Stand Age in US Forests Despite Species Turnover" Forests 10, no. 2: 114. https://doi.org/10.3390/f10020114
APA StyleHealey, S. P., & Menlove, J. (2019). The Stability of Mean Wood Specific Gravity across Stand Age in US Forests Despite Species Turnover. Forests, 10(2), 114. https://doi.org/10.3390/f10020114