

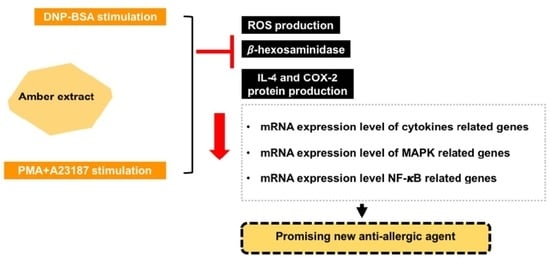
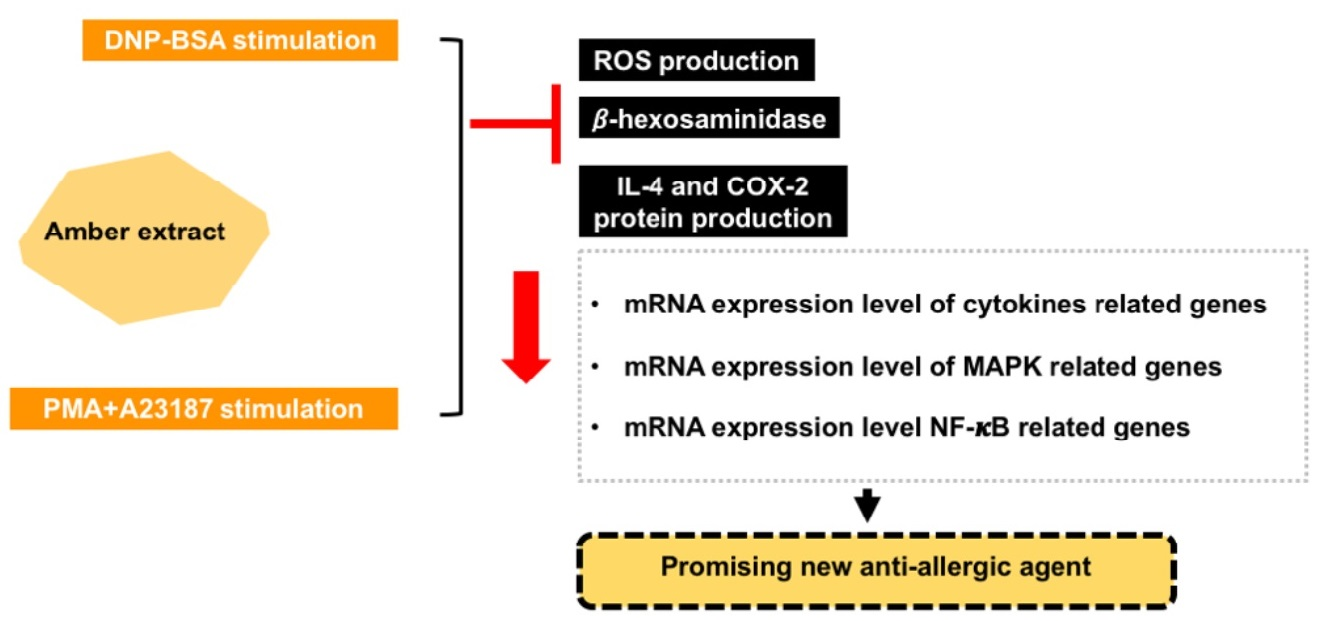
Amber Extract Suppressed Mast Cell-Mediated Allergic Inflammation via the Regulation of Allergic Mediators—An In Vitro Study
 ,
,
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Amber Extract
2.3. Cell Culture
2.4. Cell Viability Assay
2.5. Determination of Nitric Oxide (NO) Production
2.6. Determination of β-Hexosaminidase
2.7. Intracellular ROS Accumulation
2.8. Western Blotting
2.9. qRT-PCR
2.10. Statistical Analysis
3. Results
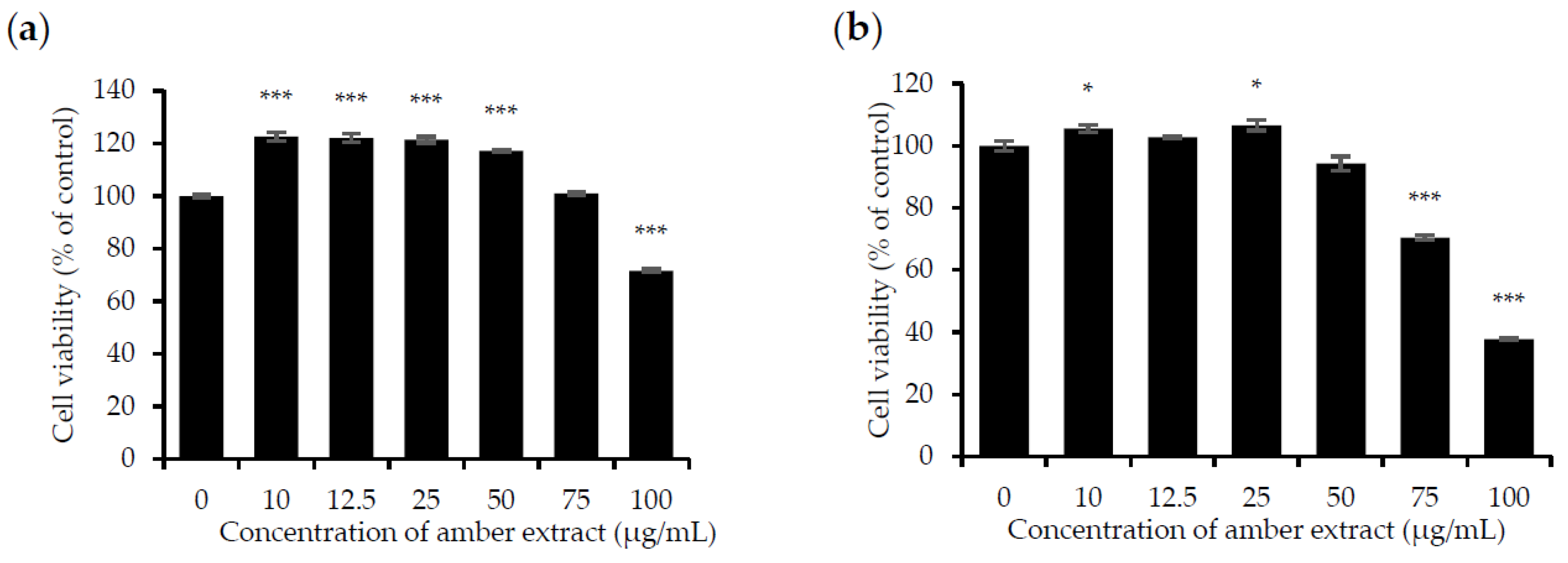
3.1. Amber Extract at Concentrations up to 50 μg/mL Showed No Significant Cytotoxic Effects on RAW 264.7 and RBL-2H3 Cells
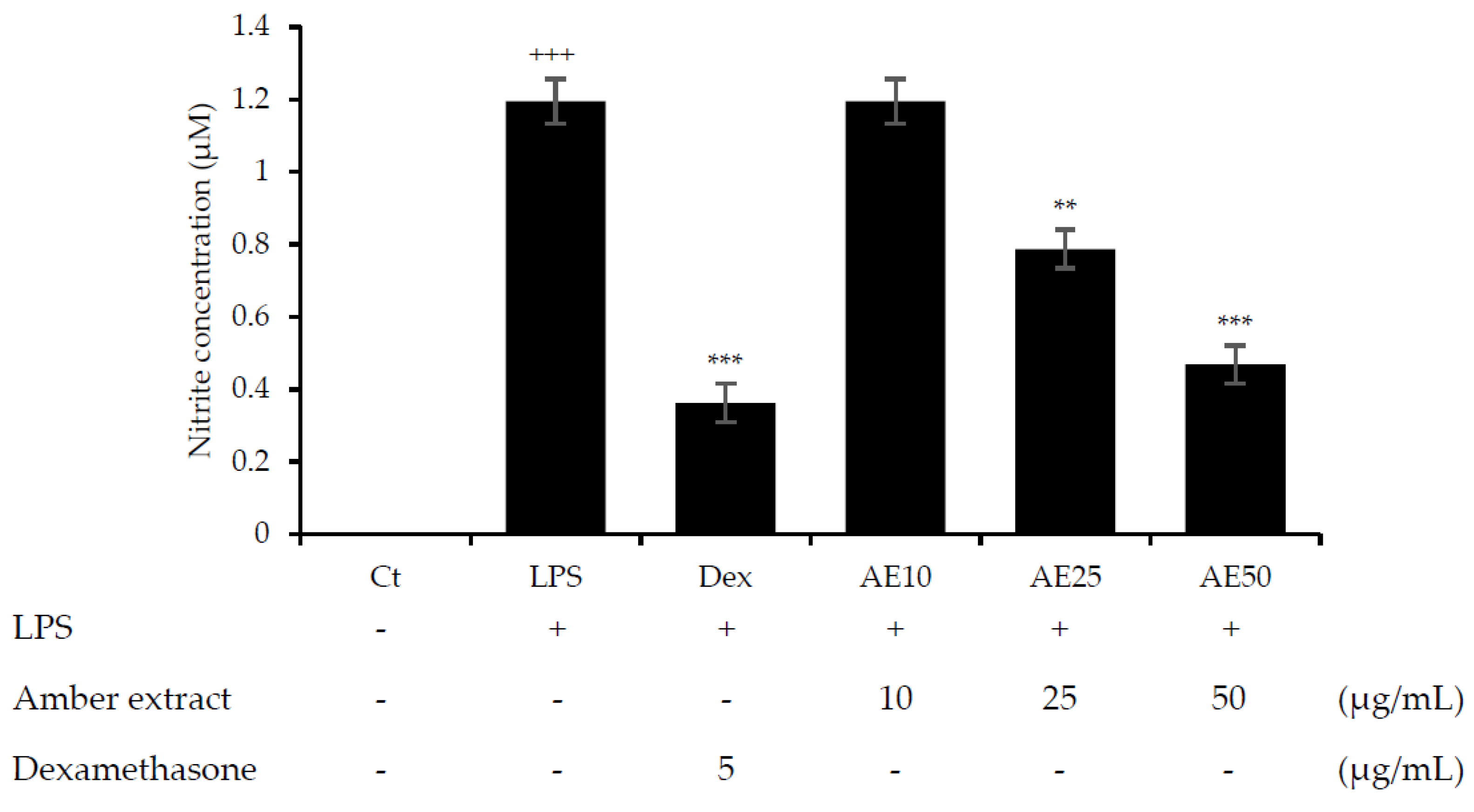
3.2. Amber Extract Significantly Inhibited NO Production
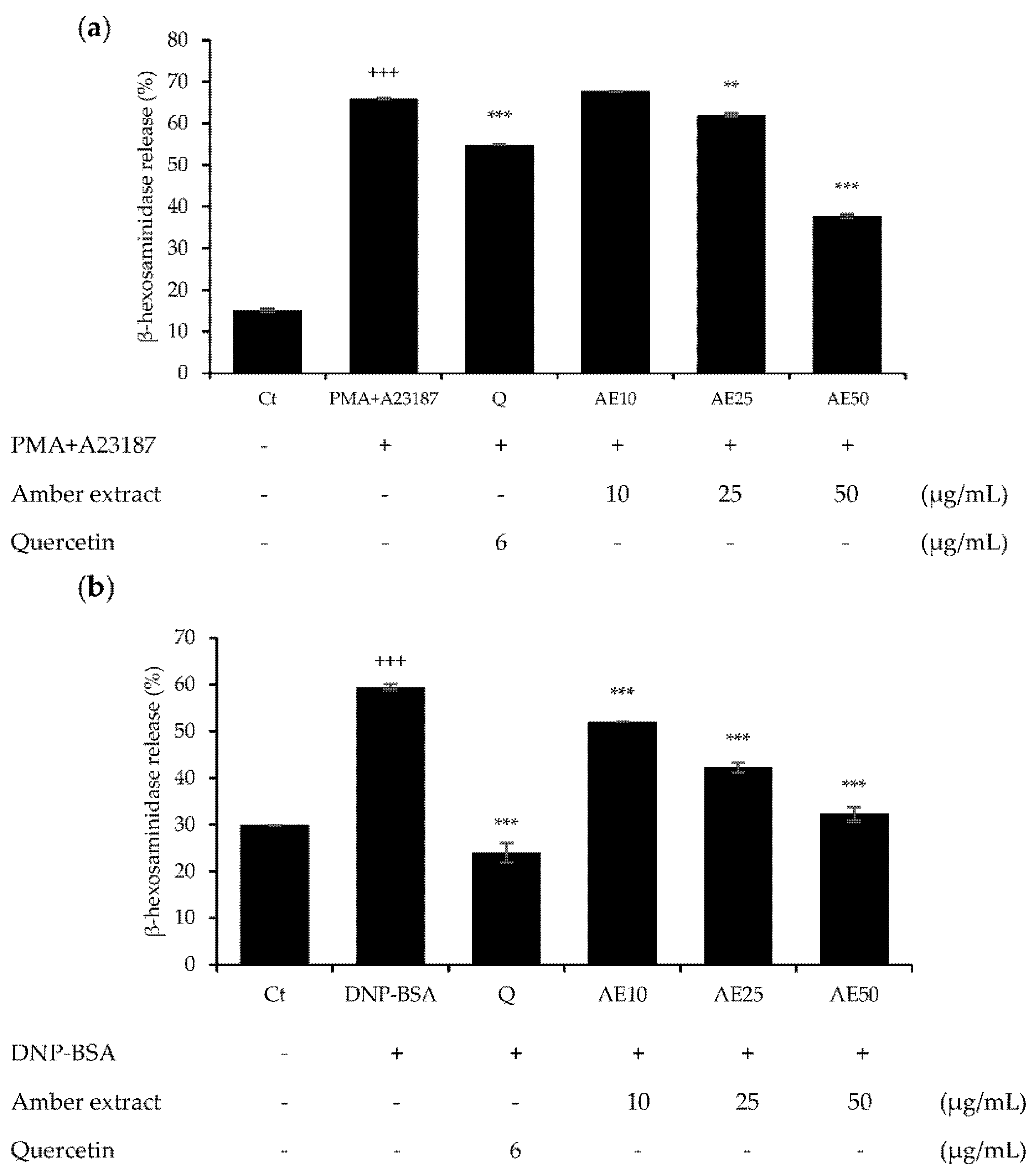
3.3. Effect of Amber Extract on β-Hexosaminidase Secretion in RBL-2H3 Cells
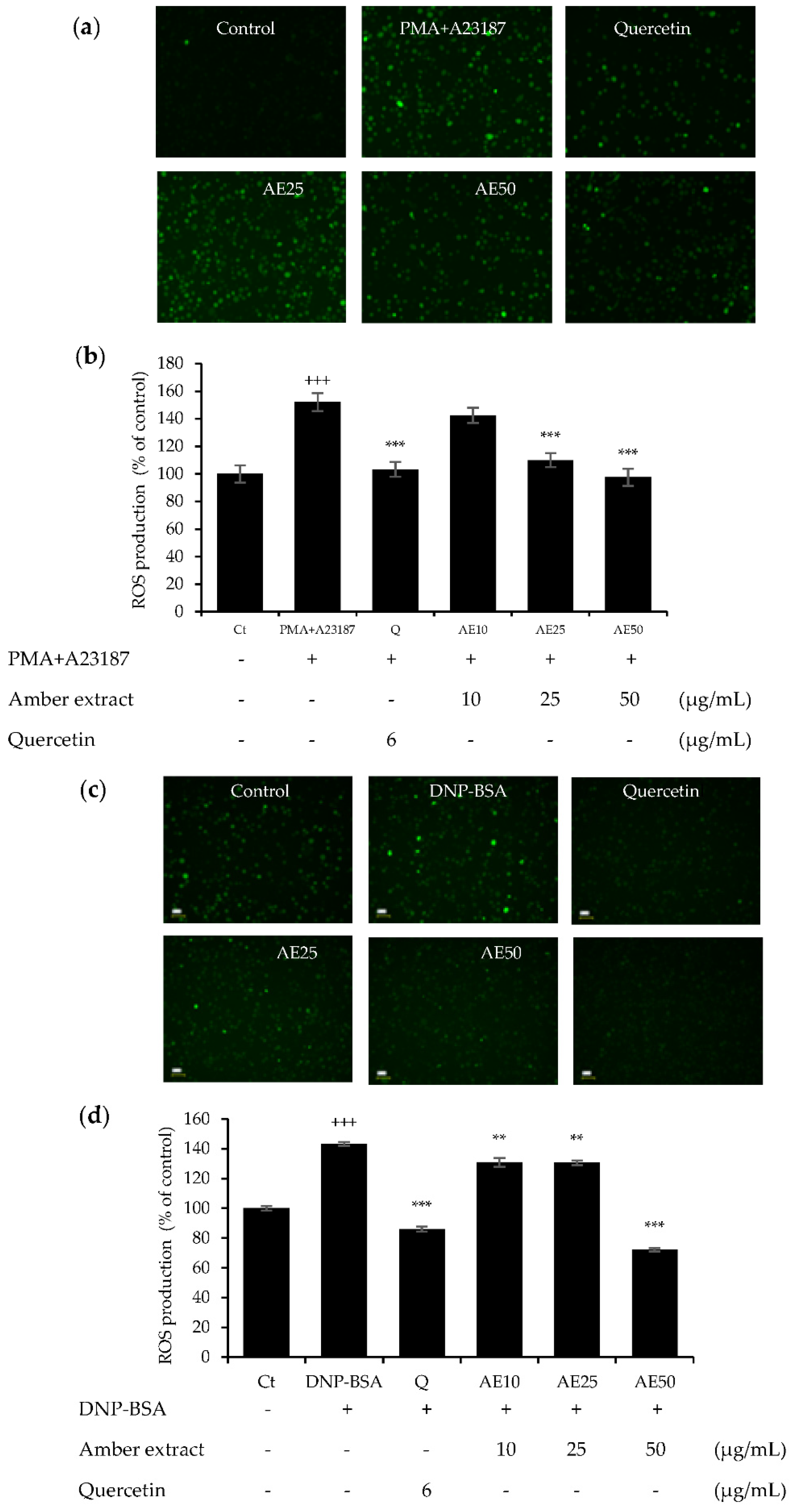
3.4. Amber Extract Inhibited Reactive Oxygen Species (ROS) Generation in RBL-2H3 Cells
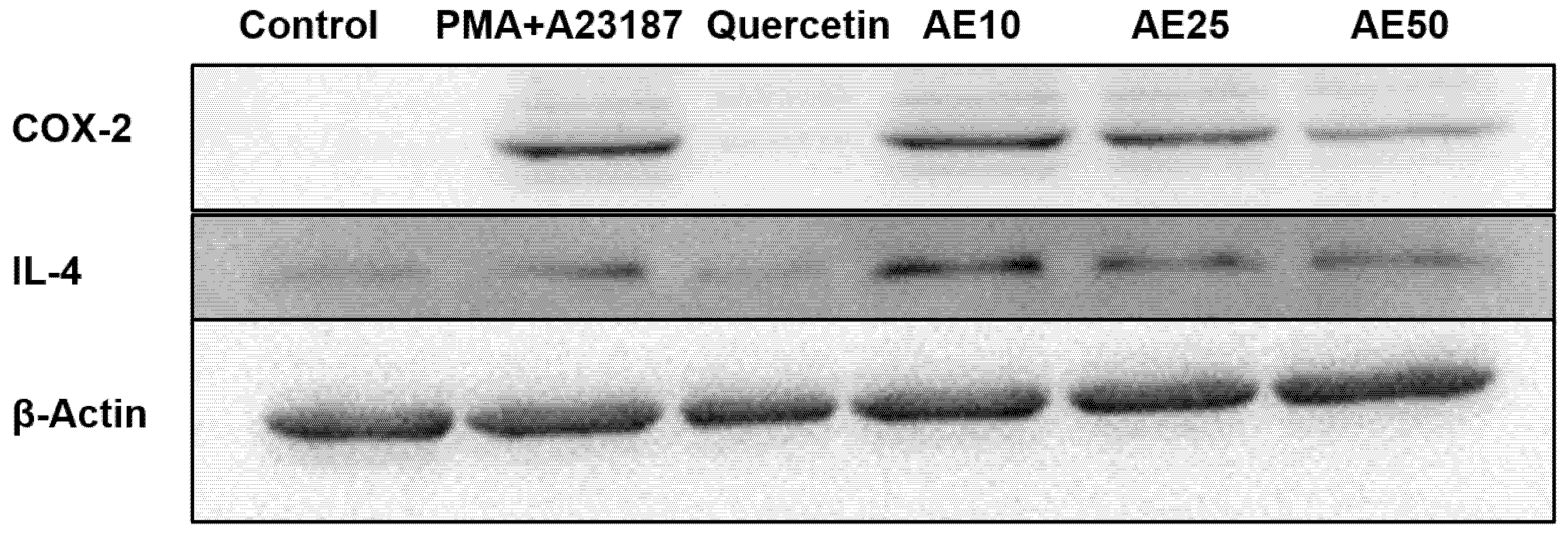
3.5. Amber Extract Inhibited the Inflammatory Mediators
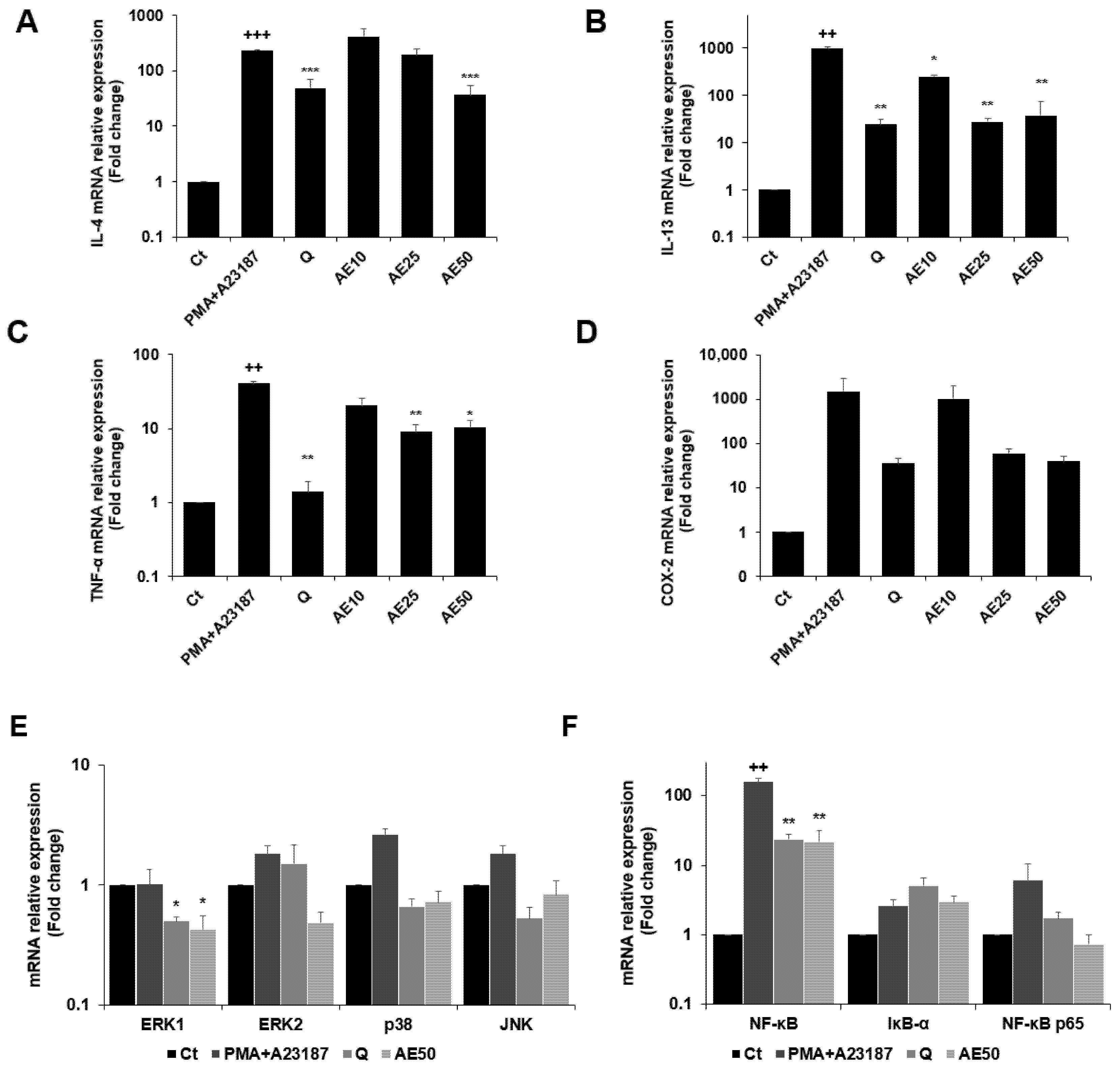
3.6. Effects of Amber Extract on Cytokine-, MAPK-, and Nuclear Factor (NF)-κB-Related Gene Expressions in RBL-2H3 Cells Stimulated with PMA + A23187
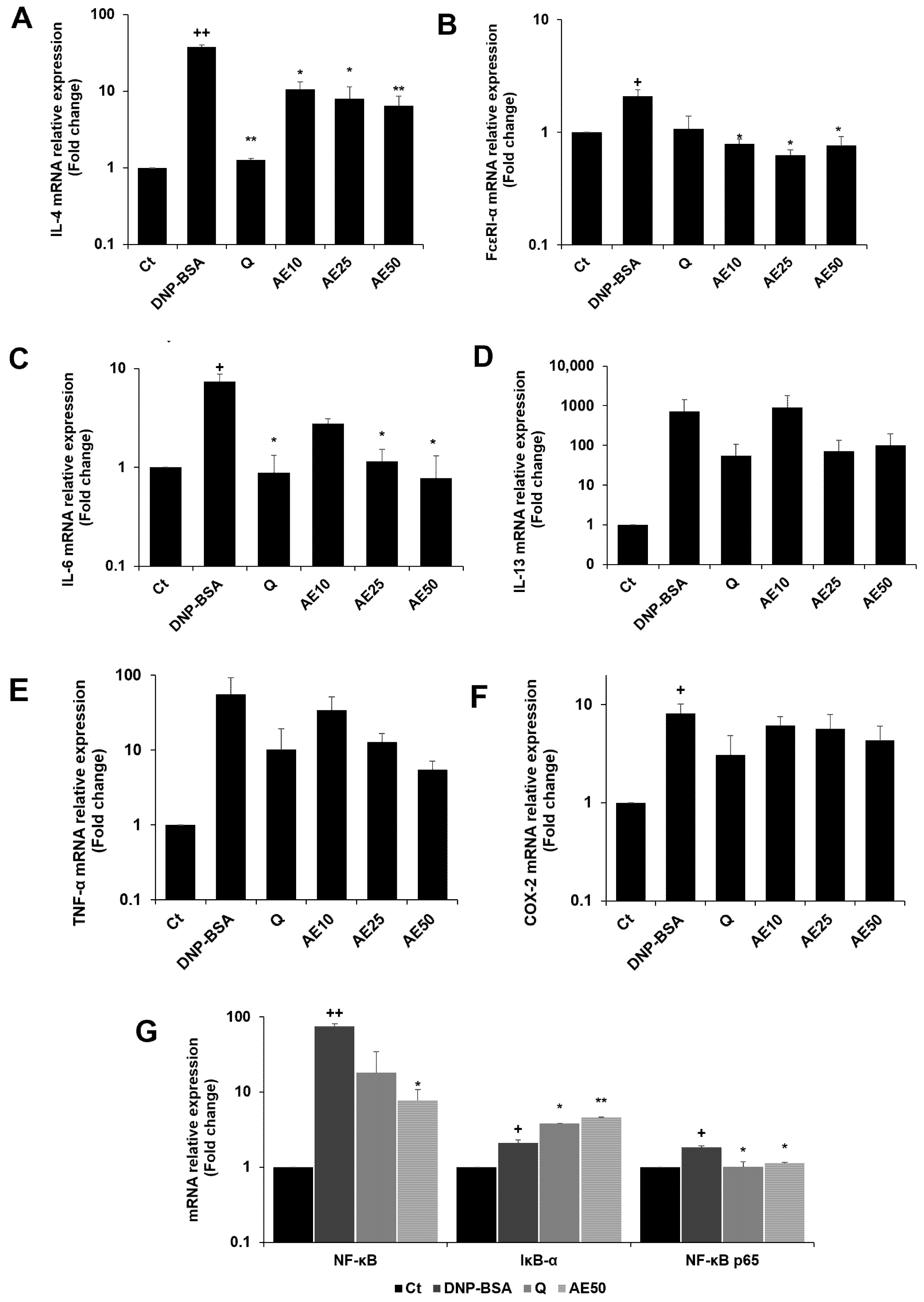
3.7. Effects of Amber Extract on Cytokine- and NF-κB-Related Gene Expressions in RBL-2H3 Cells Stimulated with DNP-BSA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, Y.Y.; Je, I.G.; Kim, M.J.; Kang, B.C.; Choi, Y.A.; Baek, M.C.; Lee, B.; Choi, J.K.; Park, H.R.; Shin, T.Y.; et al. 2-Hydroxy-3-methoxybenzoic acid attenuates mast cell-mediated allergic reaction in mice via modulation of the FcεRI signaling pathway. Acta Pharmacol. Sin. 2017, 38, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast cell: A multi-functional master cell. Front. Immunol. 2015, 6, 620. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Kwon, O.; Kim, D.Y.; Taketomi, Y.; Murakami, M.; Chang, H.W. NecroX-5 suppresses IgE/Ag-stimulated anaphylaxis and mast cell activation by regulating the SHP-1- Syk signaling module. Allergy 2016, 71, 198–209. [Google Scholar] [CrossRef] [PubMed]
- De Benedetto, A.; Agnihothri, R.; McGirt, L.Y.; Bankova, L.G.; Beck, L.A. Atopic dermatitis: A disease caused by innate immune defects? J. Investig. Dermatol. 2009, 129, 14–30. [Google Scholar] [CrossRef] [PubMed]
- Ogi, K.; Takabayashi, T.; Yamada, T.; Sakashita, M.; Kanno, M.; Narita, N.; Fujieda, S. Trimellitic anhydride induces low-grade mast cell degranulation without specific IgE. Toxicol. Rep. 2016, 3, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Bansode, R.R.; Plundrich, N.J.; Randolph, P.D.; Lila, M.A.; Williams, L.L. Peanut flour aggregation with polyphenolic extracts derived from peanut skin inhibits IgE binding capacity and attenuates RBL-2H3 cells degranulation via MAPK signaling pathway. Food Chem. 2018, 263, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zhou, S.; Haeiwa, H.; Takeda, R.; Okazaki, K.; Sekita, M.; Yamamoto, T.; Yamano, M.; Sakamoto, K. Role of amber extract in protecting SHSY5Y cells against amyloid β1-42-induced neurotoxicity. Biomed. Pharmacother. 2021, 141, 111804. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zhou, S.; Takeda, R.; Okazaki, K.; Sekita, M.; Sakamoto, K. Anti-inflammatory activities of amber extract in lipopolysaccharide-induced RAW 264.7 macrophages. Biomed. Pharmacother. 2021, 141, 111854. [Google Scholar] [CrossRef]
- Sogo, E.; Zhou, S.; Haeiwa, H.; Takeda, R.; Okazaki, K.; Sekita, M.; Yamamoto, T.; Yamano, M.; Sakamoto, K. Amber extract reduces lipid content in mature 3T3-L1 adipocytes by activating the lipolysis pathway. Molecules 2021, 26, 4630. [Google Scholar] [CrossRef]
- Suzuki, S.; Abe, J.; Kudo, Y.; Shirai, M.; Kimura, K.I. Inhibition of melanin production and promotion of collagen production by the extract of Kuji amber. Biosci. Biotechnol. Biochem. 2020, 84, 518–525. [Google Scholar] [CrossRef]
- Luo, Y.; Zhou, S.; Takeda, R.; Okazaki, K.; Sekita, M.; Sakamoto, K. Protective effect of amber extract on human dopaminergic cells against 6-hydroxydopamine-induced neurotoxicity. Molecules 2022, 27, 1817. [Google Scholar] [CrossRef] [PubMed]
- Fukuishi, N.; Murakami, S.; Ohno, A.; Yamanaka, N.; Matsui, N.; Fukutsuji, K.; Yamada, S.; Itoh, K.; Akagi, M. Does β-hexosaminidase function only as a degranulation indicator in mast cells? The primary role of β-hexosaminidase in mast cell granules. J. Immunol. 2014, 193, 1886–1894. [Google Scholar] [CrossRef] [PubMed]
- Vávra, N. The Chemistry of amber—facts, findings and opinions. Ann. Naturhist. Mus. Wien 2009, 111, 445–473. [Google Scholar]
- Kim, H.M.; Jung, H.S.; Shin, H.Y.; Jung, K.Y. Inhibition of mast cell-dependent anaphylaxis by succinic acid. Pharmaco. Toxicol. 1999, 84, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, F.; Jiang, H.; Xu, D.; Deng, D. Fumaric acid and succinic acid treat gestational hypertension by downregulating the expression of KCNMB1 and TET1. Exp. Ther. Med. 2021, 22, 1072. [Google Scholar] [CrossRef]
- Sova, M. Antioxidant and antimicrobial activities of cinnamic acid derivatives. Mini Rev. Med. Chem. 2012, 12, 749–767. [Google Scholar] [CrossRef]
- Nguyen, T.T.M.; Nguyen, T.T.; Lee, H.S.; Lee, B.; Min, B.S.; Kim, J.A. Anti-allergic and cytotoxic effects of sesquiterpenoids and phenylpropanoids isolated from Magnolia Biondii. Nat. Prod. Commun. 2017, 12, 1543–1545. [Google Scholar] [CrossRef]
- Urbanski, T.; Molak, W. Chemistry of Baltic amber: Part VII. Chemistry 1984, 32, 3–7. [Google Scholar]
- Takahashi, N.; Yao, R.; Kang, M.; Senda, M.; Ando, C.; Nishimura, K.; Goto, T.; Hirai, S.; Ezaki, Y.; Kawada, T. Dehydroabietic acid activates peroxisome proliferator-activated receptor-γ and stimulates insulin-dependent glucose uptake into 3T3-L1 adipocytes. BioFactors 2011, 37, 309–314. [Google Scholar] [CrossRef]
- Zhang, H.L.; Gan, X.Q.; Fan, Q.F.; Yang, J.J.; Zhang, P.; Hu, H.B.; Song, Q.S. Chemical constituents and anti-inflammatory activities of Maqian (Zanthoxylum myriacanthum var. pubescens) bark extracts. Sci. Rep. 2017, 7, 45805. [Google Scholar] [CrossRef]
- Saito, H.; Ishizaka, T.; Ishizaka, K. Mast cells and IgE: From history to today. Allergol. Int. 2013, 62, 3–12. [Google Scholar] [CrossRef]
- Yu, Y.; Blokhuis, B.R.; Garssen, J.; Redegeld, F.A. Non-IgE mediated mast cell activation. Eur. J. Pharmacol. 2016, 778, 33–43. [Google Scholar] [CrossRef]
- Matsuda, H.; Nakamura, S.; Yoshikawa, M. Degranulation inhibitors from medicinal plants in antigen-stimulated rat basophilic leukemia (RBL-2H3) cells. Chem. Pharm. Bull. 2016, 64, 96–103. [Google Scholar] [CrossRef]
- Huang, L.; Li, T.; Zhou, H.; Qiu, P.; Wu, J.; Liu, L. Sinomenine potentiates degranulation of RBL-2H3 basophils via up-regulation of phospholipase A2 phosphorylation by annexin A1 cleavage and ERK phosphorylation without influencing on calcium mobilization. Int. Immunopharmacol. 2015, 28, 945–951. [Google Scholar] [CrossRef]
- Suzuki, Y.; Yoshimaru, T.; Inoue, T.; Niide, O.; Ra, C. Role of oxidants in mast cell activation. Chem. Immunol. Allergy. 2005, 87, 32–42. [Google Scholar] [CrossRef]
- Van Rijt, L.S.; Utsch, L.; Lutter, R.; Van Ree, R. Oxidative stress: Promoter of allergic sensitization to protease allergens? Int. J. Mol. Sci. 2017, 18, 1112. [Google Scholar] [CrossRef]
- Poulin, J.; Helwig, K. Inside amber: The structural role of succinic acid in class Ia and class Id resinite. Anal. Chem. 2014, 86, 7428–7435. [Google Scholar] [CrossRef]
- Zarubina, I.V.; Lukk, M.V.; Shabanov, P.D. Antihypoxic and antioxidant effects of exogenous succinic acid and aminothiol succinate-containing antihypoxants. Bull. Exp. Biol. Med. 2012, 153, 336–339. [Google Scholar] [CrossRef]
- Saggini, A.; Maccauro, G.; Tripodi, D.; De Lutiis, M.A.; Conti, F.; Felaco, P.; Fulcheri, M.; Galzio, R.; Caraffa, A.; Antinolfi, P.; et al. Allergic inflammation: Role of cytokines with special emphasis on IL-4. Int. J. Immunopathol. Pharmacol. 2011, 24, 305–311. [Google Scholar] [CrossRef]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer. 2013, 12, 86. [Google Scholar] [CrossRef]
- Brown, M.A.; Hural, J. Functions of IL-4 and control of its expression. Crit. Rev. Immunol. 1997, 17, 1–32. [Google Scholar] [CrossRef]
- Yang, M.; Wang, Y.; Patel, G.; Xue, Q.; Singor Njateng, G.S.; Cai, S.; Cheng, G.; Kai, G. In vitro and in vivo anti-inflammatory effects of different extracts from Epigynum auritum through down-regulation of NF-κB and MAPK signaling pathways. J. Ethnopharmacol. 2020, 261, 113105. [Google Scholar] [CrossRef]
- Kritas, S.K.; Saggini, A.; Varvara, G.; Murmura, G.; Caraffa, A.; Antinolfi, P.; Toniato, E.; Pantalone, A.; Neri, G.; Frydas, S.; et al. Impact of mast cells on the skin. Int. J. Immunopathol. Pharmacol. 2013, 26, 855–859. [Google Scholar] [CrossRef]
- Kawai, M.; Hirano, T.; Higa, S.; Arimitsu, J.; Maruta, M.; Kuwahara, Y.; Ohkawara, T.; Hagihara, K.; Yamadori, T.; Shima, Y.; et al. Flavonoids and related compounds as anti-allergic substances. Allergol. Int. 2007, 56, 113–123. [Google Scholar] [CrossRef]
- Do, H.J.; Oh, T.W.; Yang, J.H.; Park, K.I.; Ma, J.Y. Davallia mariesii moore improves FcεRI-mediated allergic responses in the rat basophilic leukemia mast cell line RBL-2H3 and passive cutaneous anaphylaxis in mice. Mediators Inflam. 2017, 2017, 8701650. [Google Scholar] [CrossRef]
- Marone, G.; Granata, F.; Pucino, V.; Pecoraro, A.; Heffler, E.; Loffredo, S.; Scadding, G.W.; Varricchi, G. The intriguing role of interleukin 13 in the pathophysiology of asthma. Front. Pharmacol. 2019, 10, 1387. [Google Scholar] [CrossRef]
- Wynn, T.A. IL-13 effector functions. Annu. Rev. Immunol. 2003, 21, 425–456. [Google Scholar] [CrossRef]
- Chelombitko, M.A.; Fedorov, A.V.; Ilyinskaya, O.P.; Zinovkin, R.A.; Chernyak, B.V. Role of reactive oxygen species in mast cell degranulation. Biochemistry (Mosc.) 2016, 81, 1564–1577. [Google Scholar] [CrossRef]
- Zhu, M.; Zou, J.; Li, T.; O’Brien, S.A.; Zhang, Y.; Ogden, S.; Zhang, W. Differential roles of phospholipase D proteins in FcεRI-mediated signaling and mast cell function. J. Immunol. 2015, 195, 4492–4502. [Google Scholar] [CrossRef]
- Gilfillan, A.M.; Tkaczyk, C. Integrated signalling pathways for mast-cell activation. Nat. Rev. Immunol. 2006, 6, 218–230. [Google Scholar] [CrossRef]
- Frossi, B.; De Carli, M.; Daniel, K.C.; Rivera, J.; Pucillo, C. Oxidative stress stimulates IL-4 and IL-6 production in mast cells by an APE/Ref-1-dependent pathway. Eur. J. Immunol. 2003, 33, 2168–2177. [Google Scholar] [CrossRef]
- Gordon, J.R.; Galli, S.J. Mast cells as a source of both preformed and immunologically inducible TNF-alpha/cachectin. Nature 1990, 346, 274–276. [Google Scholar] [CrossRef]
- Lorz, L.R.; Kim, D.; Kim, M.Y.; Cho, J.Y. Panax ginseng–derived fraction BIOGF1K reduces atopic dermatitis responses via suppression of mitogen-activated protein kinase signaling pathway. J. Ginseng Res. 2020, 44, 453–460. [Google Scholar] [CrossRef]
- Robinson, M.J.; Cobb, M.H. Mitogen-activated protein kinase pathways. Curr. Opin. Cell Biol. 1997, 9, 180–186. [Google Scholar] [CrossRef]
- Hidding, U.; Mielke, K.; Waetzig, V.; Brecht, S.; Hanisch, U.; Behrens, A.; Wagner, E.; Herdegen, T. The c-Jun N-terminal kinases in cerebral microglia: Immunological functions in the brain. Biochem. Pharmacol. 2002, 64, 781–788. [Google Scholar] [CrossRef]
- Kim, E.; Kang, Y.G.; Kim, Y.J.; Lee, T.R.; Yoo, B.C.; Jo, M.; Kim, J.H.; Kim, J.H.; Kim, D.; Cho, J.Y. Dehydroabietic acid suppresses inflammatory response via suppression of Src-, Syk-, and TAK1-mediated pathways. Int. J. Mol. Sci. 2019, 20, 1593. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Refli, R.; Sofyana, N.T.; Haeiwa, H.; Takeda, R.; Okazaki, K.; Sekita, M.; Sakamoto, K. Amber Extract Suppressed Mast Cell-Mediated Allergic Inflammation via the Regulation of Allergic Mediators—An In Vitro Study. Nutraceuticals 2023, 3, 75-90. https://doi.org/10.3390/nutraceuticals3010006
Refli R, Sofyana NT, Haeiwa H, Takeda R, Okazaki K, Sekita M, Sakamoto K. Amber Extract Suppressed Mast Cell-Mediated Allergic Inflammation via the Regulation of Allergic Mediators—An In Vitro Study. Nutraceuticals. 2023; 3(1):75-90. https://doi.org/10.3390/nutraceuticals3010006
Chicago/Turabian StyleRefli, Redoyan, Neng Tanty Sofyana, Haruna Haeiwa, Reiko Takeda, Kazuma Okazaki, Marie Sekita, and Kazuichi Sakamoto. 2023. "Amber Extract Suppressed Mast Cell-Mediated Allergic Inflammation via the Regulation of Allergic Mediators—An In Vitro Study" Nutraceuticals 3, no. 1: 75-90. https://doi.org/10.3390/nutraceuticals3010006
APA StyleRefli, R., Sofyana, N. T., Haeiwa, H., Takeda, R., Okazaki, K., Sekita, M., & Sakamoto, K. (2023). Amber Extract Suppressed Mast Cell-Mediated Allergic Inflammation via the Regulation of Allergic Mediators—An In Vitro Study. Nutraceuticals, 3(1), 75-90. https://doi.org/10.3390/nutraceuticals3010006