
Wide Range Applications of Spirulina: From Earth to Space Missions

 , , ,
, , ,  ,
,  ,
,  , ,
, , 

Abstract
1. Introduction
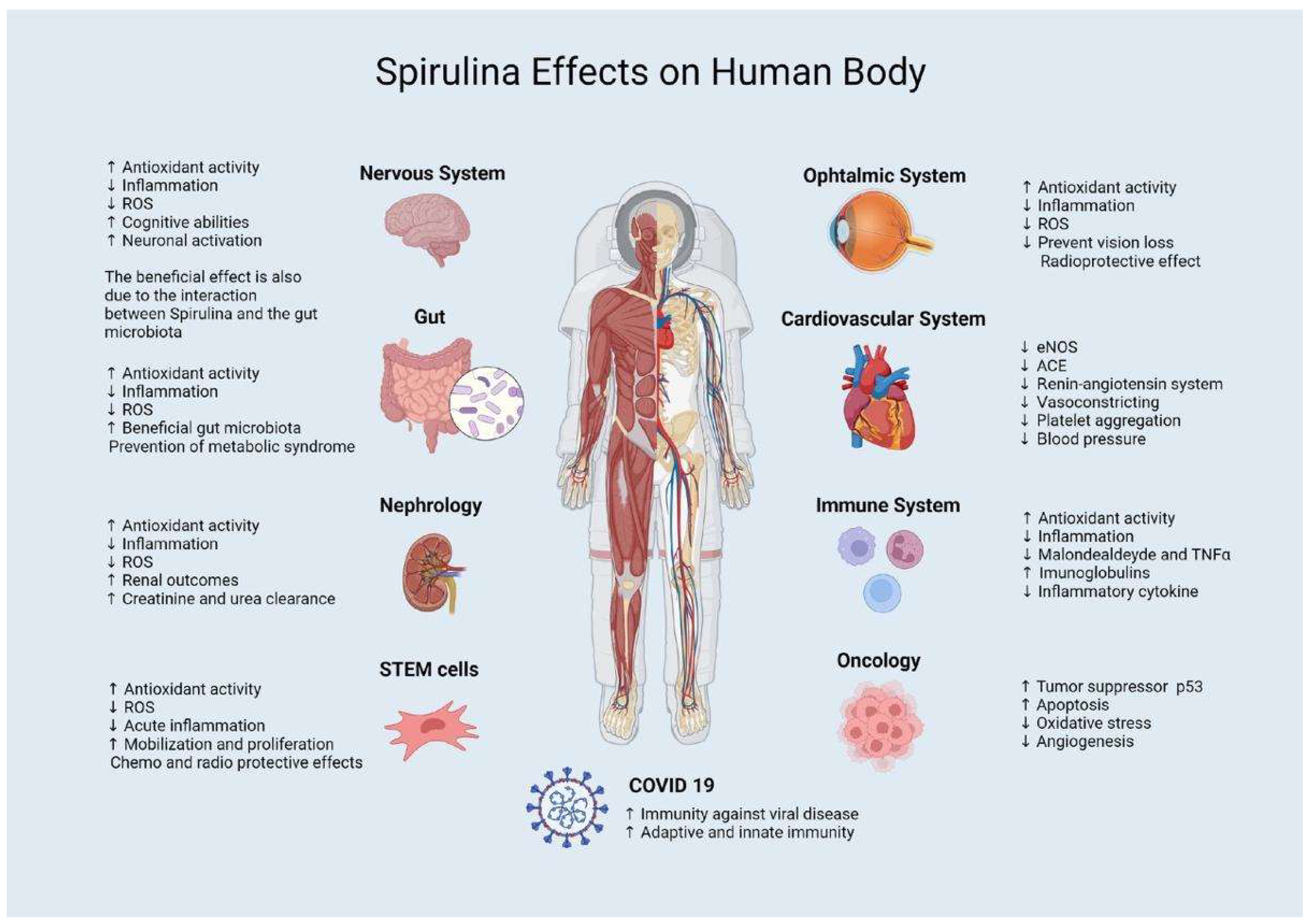
2. Clinical Applications on Earth
2.1. Probiotic Effect
2.2. Antihypertensive Effect
2.3. Antioxidant and Anti-Inflammatory Effects
2.4. Antiviral Effect
2.5. Antihistamine Effect
3. Applications in Ophthalmology
3.1. Therapeutic Effect of Spirulina in Corneal Inflammation
3.2. Radioprotective Effect of Spirulina on Lacrimal Glands
3.3. The Protection of Spirulina on the Visual Function
4. Applications in Oncology
4.1. Breast Cancer
4.2. Hepatocarcinoma
4.3. Lung Cancer
5. Applications in Central and Peripheral Nervous System
6. Applications in COVID-19 Infection
7. Interaction with Stem Cells
8. Nephrological Involvement
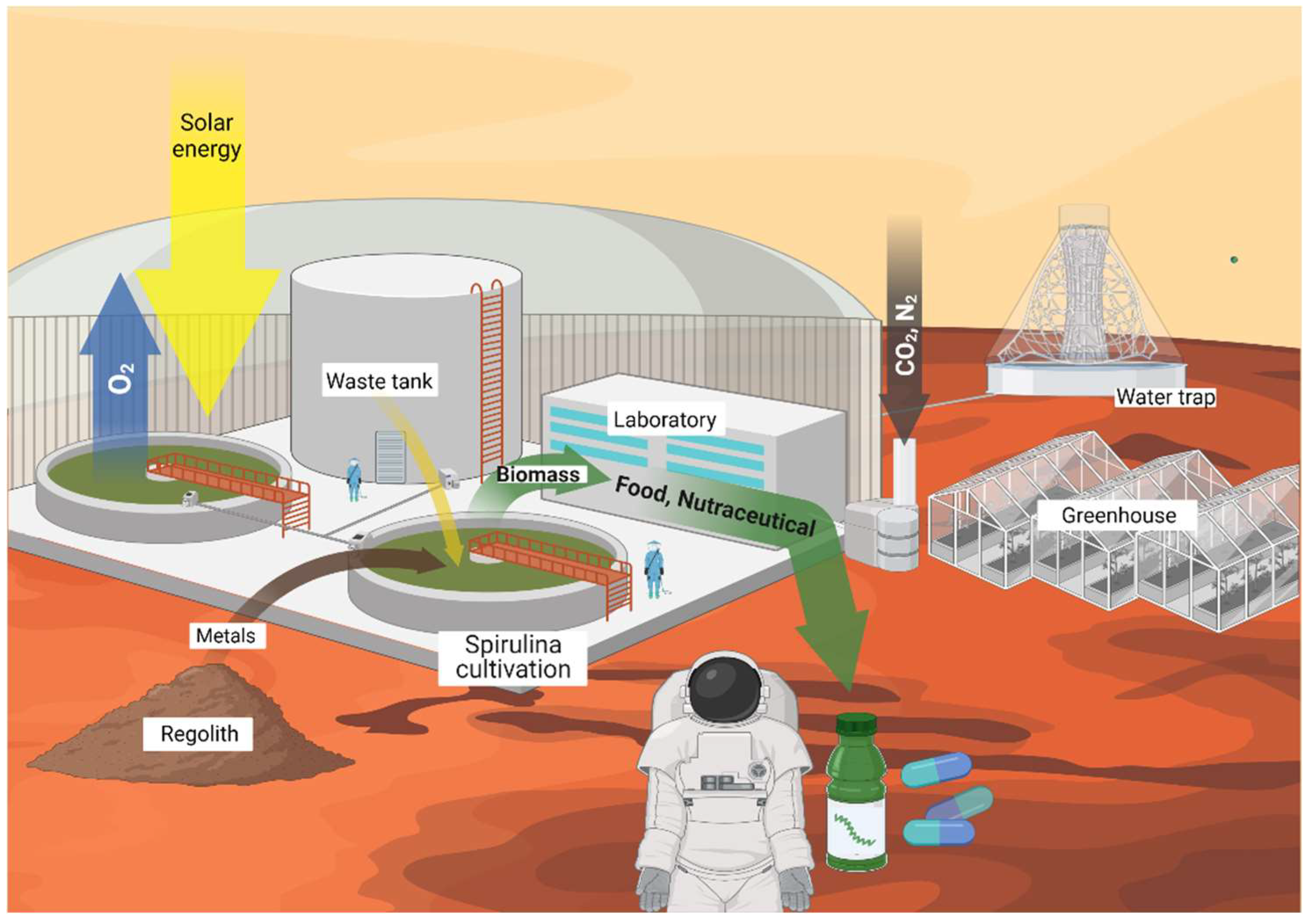
9. Beyond Earth: Space Sustainability
10. Spirulina for Astronauts’ Healthcare
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Komárek, J.; Kaštovský, J.; Mareš, J.; Johansen, J.R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach Taxonomické hodnocení cyanoprokaryot (cyanobakteriální rody) v roce 2014 podle polyfázického přístupu. Preslia 2014, 86, 295–335. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. 2022. Available online: https://www.algaebase.org/search/genus/detail/?genus_id=43076 (accessed on 7 March 2022).
- Hoseini, S.M.; Khosravi-Darani, K.; Mozafari, M.R. Nutritional and Medical Applications of Spirulina Microalgae. Mini-Rev. Med. Chem. 2013, 13, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Banji, D.; Banji, O.J.F.; Pratusha, N.G.; Annamalai, A.R. Investigation on the role of Spirulina platensis in ameliorating behavioural changes, thyroid dysfunction and oxidative stress in offspring of pregnant rats exposed to fluoride. Food Chem. 2013, 140, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, B.; Fletcher, I.R.; Brocks, J.J.; Kilburn, M.R. Reassessing the first appearance of eukaryotes and cyanobacteria. Nature 2008, 455, 1101–1104. [Google Scholar] [CrossRef] [PubMed]
- Gaysina, L.A.; Saraf, A.; Singh, P. Cyanobacteria in Diverse Habitats; Elsevier Inc.: Amsterdam, The Netherlands, 2018; ISBN 9780128146682. [Google Scholar]
- Thomas, S.S. The Role of Parry Organic Spirulina in Health Management; Valensa International: Eustis, FL, USA, 2010. [Google Scholar]
- Pyne, S.K.; Bhattacharjee, P.; Srivastav, P.P. Microalgae (Spirulina Platensis) and Its Bioactive Molecules: Review. Indian J. Nutr. 2017, 4, 160. [Google Scholar]
- Soni, R.A.; Sudhakar, K.; Rana, R.S. Spirulina—From growth to nutritional product: A review. Trends Food Sci. Technol. 2017, 69, 157–171. [Google Scholar] [CrossRef]
- Colla, L.M.; Oliveira Reinehr, C.; Reichert, C.; Costa, J.A.V. Production of biomass and nutraceutical compounds by Spirulina platensis under different temperature and nitrogen regimes. Bioresour. Technol. 2007, 98, 1489–1493. [Google Scholar] [CrossRef]
- Vaz, B.S.; Moreira, J.B.; Morais, M.G.; Costa, J.A.V. Microalgae as a new source of bioactive compounds in food supplement. Curr. Opin. Food Sci. 2016, 7, 73–77. [Google Scholar] [CrossRef]
- De Morais, M.G.; da Silva Vaz, B.; Greque de Morais, E.; Vieira Costa, J.A. Biologically Active Metabolites Synthesized by Microalgae. BioMed Res. Int. 2015, 2015, 835761. [Google Scholar] [CrossRef]
- Ciferri, O. Spirulina, the edible microorganism. Microbiol. Rev. 1983, 47, 551. [Google Scholar] [CrossRef]
- Dillon, J.C.; Phuc, A.P.; Dubacq, J.P. Nutritional Value of the Alga Spirulina. World Rev. Nutr. Diet. 1995, 77, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Dillon, J.C.; Phan, P.A. Spirulina as a source of proteins in human nutrition. Bulletin de l’Institut Océanographique 1993, 12, 103–107. [Google Scholar]
- Braune, S.; Krüger-Genge, A.; Kammerer, S.; Jung, F.; Küpper, J.H. Phycocyanin from arthrospira platensis as potential anti-cancer drug: Review of in vitro and in vivo studies. Life 2021, 11, 91. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Liu, L.; Miron, A.; Klímová, B.; Wan, D.; Kuča, K. The antioxidant, immunomodulatory, and anti-inflammatory activities of Spirulina: An overview. Arch. Toxicol. 2016, 90, 1817–1840. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Blankenship, R.E. On the interface of light-harvesting antenna complexes and reaction centers in oxygenic photosynthesis. Biochim. Biophys. Acta-Bioenerg. 2019, 1860, 148079. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Oliveira, O.; Oliveira, S.; Mendes, A.M.; Caetano, N.S. Lipid content and productivity of arthrospira platensis and chlorella vulgaris under mixotrophic conditions and salinity stress. Chem. Eng. Trans. 2016, 49, 187–192. [Google Scholar] [CrossRef]
- Ramadan, M.F.; Asker, M.M.S.; Ibrahim, Z.K. Functional bioactive compounds and biological activities of spirulina platensis lipids. Czech J. Food Sci. 2008, 26, 211–222. [Google Scholar] [CrossRef]
- Kulshreshtha, A.; Anish, J.Z.; Jarouliya, U.; Bhadauriya, P.; Prasad, G.; Bisen, P. Spirulina in Health Care Management. Curr. Pharm. Biotechnol. 2008, 9, 400–405. [Google Scholar] [CrossRef]
- Mühling, M.; Belay, A.; Whitton, B.A. Variation in fatty acid composition of Arthrospira (Spirulina) strains. J. Appl. Phycol. 2005, 17, 137–146. [Google Scholar] [CrossRef]
- Sotiroudis, T.G.; Sotiroudis, G.T. Health aspects of Spirulina (Arthrospira) microalga food supplement. J. Serb. Chem. Soc. 2013, 78, 395–405. [Google Scholar] [CrossRef]
- Diraman, H.; Koru, E.; Dibeklioglu, H. Fatty acid profile of Spirulina platensis used as a food supplement. Isr. J. Aquac.-Bamidgeh 2009, 61, 134–142. [Google Scholar] [CrossRef]
- Sánchez, M.; Bernal-Castillo, J.; Rozo, C.; Rodríguez, I. SPIRULINA (ARTHROSPIRA): AN EDIBLE MICROORGANISM: A REVIEW. Univ. Sci. 2003, 8, 7–24. Available online: https://revistas.javeriana.edu.co/index.php/scientarium/article/view/4842 (accessed on 7 March 2022).
- Matos, J.; Cardoso, C.; Bandarra, N.M.; Afonso, C. Microalgae as healthy ingredients for functional food: A review. Food Funct. 2017, 8, 2672–2685. [Google Scholar] [CrossRef] [PubMed]
- Pugh, N.; Ross, S.A.; Elsohly, H.N.; Elsohly, M.A.; Pasco, D.S. Isolation of three high molecular weight polysaccharide preparations with potent immunostimulatory activity from Spirulina platensis, aphanizomenon flos-aquae and Chlorella pyrenoidosa. Planta Med. 2001, 67, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Sukenik, A.; Hadas, O.; Kaplan, A.; Quesada, A. Invasion of Nostocales (cyanobacteria) to subtropical and temperate freshwater lakes—physiological, regional, and global driving forces. Front. Microbiol. 2012, 3, 86. [Google Scholar] [CrossRef] [PubMed]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Zahra, Z.; Choo, D.H.; Lee, H.; Parveen, A. Cyanobacteria: Review of current potentials and applications. Environments 2020, 7, 13. [Google Scholar] [CrossRef]
- Abdulqader, G.; Barsanti, L.; Tredici, M.R. Harvest of Arthrospira platensis from Lake Kossorom (Chad) and its household usage among the Kanembu. J. Appl. Phycol. 2000, 12, 493–498. [Google Scholar] [CrossRef]
- Ramírez, L.; Olvera, R. Uso tradicional y actual de Spirulina sp. (Arthrospira sp.). Interciencia 2006, 31, 657–663. [Google Scholar]
- Salazar, M.; Chamorro, G.A.; Salazar, S.; Steele, C.E. Effect of Spirulina maxima consumption on reproduction and peri- and postnatal development in rats. Food Chem. Toxicol. 1996, 34, 353–359. [Google Scholar] [CrossRef]
- Chamorro, G.; Salazar, S.; Steele, C.; Salazar, M. Reproduction and peri- and postnatal evaluation of Spirulina maxima in mice. Toxicol. Lett. 1996, 88, 67. [Google Scholar] [CrossRef]
- Salazar, M.; Martínez, E.; Madrigal, E.; Ruiz, L.E.; Chamorro, G.A. Subchronic toxicity study in mice fed Spirulina maxima. J. Ethnopharmacol. 1998, 62, 235–241. [Google Scholar] [CrossRef]
- Simpore, J.; Kabore, F.; Zongo, F.; Dansou, D.; Bere, A.; Pignatelli, S.; Biondi, D.M.; Ruberto, G.; Musumeci, S. Nutrition rehabilitation of undernourished children utilizing Spiruline and Misola. Nutr. J. 2006, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Marles, R.J.; Barrett, M.L.; Barnes, J.; Chavez, M.L.; Gardiner, P.; Ko, R.; Mahady, G.B.; Dog, T.L.; Sarma, N.D.; Giancaspro, G.I.; et al. United states pharmacopeia safety evaluation of spirulina. Crit. Rev. Food Sci. Nutr. 2011, 51, 593–604. [Google Scholar] [CrossRef]
- Tarantino LM Agency Response Letter GRASS Notice No. GRN 000127. Available online: https://www.algbiotek.com/spirulina-sertifikalar/spirulina-gras-2012.pdf (accessed on 10 March 2022).
- Moorhead, K.; Capelli, B.; Cysewski, G.R. SPIRULINA Nature’s Superfood; Cyanotech Corporation: Kailua-Kona, HI, USA, 2013; Volume 53, ISBN 9788578110796. Available online: https://www.terapiaclark.es/Docs/spirulina_book.pdf (accessed on 10 March 2022).
- Milia, M.; Corrias, F.; Addis, P.; Zitelli, G.C.; Cicchi, B.; Torzillo, G.; Andreotti, V.; Angioni, A. Influence of Different Light Sources on the Biochemical Composition of Arthrospira spp. Grown in Model Systems. Foods 2022, 11, 399. [Google Scholar] [CrossRef]
- Çelekli, A.; Yavuzatmaca, M.; Bozkurt, H. Modeling of biomass production by Spirulina platensis as function of phosphate concentrations and pH regimes. Bioresour. Technol. 2009, 100, 3625–3629. [Google Scholar] [CrossRef]
- Mathur, M. Molécules bioactives de la spiruline: Un complément alimentaire. In Bioactive Molecules in Food; Mérillon, J.M., Ramawat, K., Eds.; Série de Référence en Phytochimie; Springer: Cham, Switzerland, 2018; pp. 1621–1642. [Google Scholar]
- Hernández Lepe, M.A.; Wall-Medrano, A.; Juárez-Oropeza, M.A.; Ramos-Jiménez, A.; Hernández-Torres, R.P. Spirulina Y Su Efecto Hipolipemiante Y Antioxidante En Humanos: Una Revisión Sistemática [Spirulina and Its Hypolipidemic And Antioxidant Effects In Humans: A Systematic Review] (in Spanish). Nutr. Hosp. 2015, 32, 494–500. [Google Scholar]
- Mysliwa-Kurdziel, B.; Solymosi, K. Phycobilins and Phycobiliproteins Used in Food Industry and Medicine. Mini-Rev. Med. Chem. 2017, 17, 1173–1193. [Google Scholar] [CrossRef]
- Karkos, P.D.; Leong, S.C.; Karkos, C.D.; Sivaji, N.; Assimakopoulos, D.A. Spirulina in clinical practice: Evidence-based human applications. Evidence-based Complement. Altern. Med. 2011, 2011, 4–7. [Google Scholar] [CrossRef]
- Tranquille, N.; Emeis, J.J.; de Chambure, D.; Binot, R.; Tamponnet, C. Spirulina acceptability trials in rats. A study for the “Melissa” life-support system. Adv. Sp. Res. 1994, 14, 167–170. [Google Scholar] [CrossRef]
- Soni, R.A.; Sudhakar, K.; Rana, R.S.; Baredar, P. Food Supplements Formulated with Spirulina. Algae 2021, 201–226. [Google Scholar] [CrossRef]
- Jung, F.; Krüger-Genge, A.; Waldeck, P.; Küpper, J.H. Spirulina platensis, a super food? J. Cell. Biotechnol. 2019, 5, 43–54. [Google Scholar] [CrossRef]
- Ciani, M.; Lippolis, A.; Fava, F.; Rodolfi, L.; Niccolai, A.; Tredici, M.R. Microbes: Food for the Future. Foods 2021, 10, 971. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Rodrigues, M.M.; Estrada-Beristain, C.; Metri-Ojeda, J.; Pérez-Alva, A.; Baigts-Allende, D.K. Spirulina platensis protein as sustainable ingredient for nutritional food products development. Sustainability 2021, 13, 6849. [Google Scholar] [CrossRef]
- Afshinnekoo, E.; Scott, R.T.; MacKay, M.J.; Pariset, E.; Cekanaviciute, E.; Barker, R.; Gilroy, S.; Hassane, D.; Smith, S.M.; Zwart, S.R.; et al. Fundamental Biological Features of Spaceflight: Advancing the Field to Enable Deep-Space Exploration. Cell 2020, 183, 1162–1184. [Google Scholar] [CrossRef]
- Garrett-Bakelman, F.E.; Darshi, M.; Green, S.J.; Gur, R.C.; Lin, L.; Macias, B.R.; McKenna, M.J.; Meydan, C.; Mishra, T.; Nasrini, J.; et al. The NASA twins study: A multidimensional analysis of a year-long human spaceflight. Science 2019, 364, eaau8650. [Google Scholar] [CrossRef]
- Stein, T.P.; Leskiw, M.J. Oxidant damage during and after spaceflight. Am. J. Physiol.-Endocrinol. Metab. 2000, 278, 375–382. [Google Scholar] [CrossRef]
- Tominaga, H.; Kodama, S.; Matsuda, N.; Suzuki, K.; Watanabe, M. Involvement of reactive oxygen species (ROS) in the induction of genetic instability by radiation. J. Radiat. Res. 2004, 45, 181–188. [Google Scholar] [CrossRef]
- Yatagai, F.; Honma, M.; Dohmae, N.; Ishioka, N. Biological effects of space environmental factors: A possible interaction between space radiation and microgravity. Life Sci. Sp. Res. 2019, 20, 113–123. [Google Scholar] [CrossRef]
- Concas, A.; Corrias, G.; Orrù, R.; Licheri, R.; Pisu, M.; Cao, G. Remarks on ISRU and ISFR technologies for manned missions on moon and mars. Eurasian Chem. J. 2012, 14, 243–248. [Google Scholar] [CrossRef]
- Poughon, L.; Laroche, C.; Creuly, C.; Dussap, C.G.; Paille, C.; Lasseur, C.; Monsieurs, P.; Heylen, W.; Coninx, I.; Mastroleo, F.; et al. Limnospira indica PCC8005 growth in photobioreactor: Model and simulation of the ISS and ground experiments. Life Sci. Sp. Res. 2020, 25, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Alemany, L.; Peiro, E.; Arnau, C.; Garcia, D.; Poughon, L.; Cornet, J.F.; Dussap, C.G.; Gerbi, O.; Lamaze, B.; Lasseur, C.; et al. Continuous controlled long-term operation and modeling of a closed loop connecting an air-lift photobioreactor and an animal compartment for the development of a life support system. Biochem. Eng. J. 2019, 151, 107323. [Google Scholar] [CrossRef]
- Verseux, C.; Baqué, M.; Lehto, K.; De Vera, J.P.P.; Rothschild, L.J.; Billi, D. Sustainable life support on Mars—The potential roles of cyanobacteria. Int. J. Astrobiol. 2016, 15, 65–92. [Google Scholar] [CrossRef]
- Verseux, C.; Heinicke, C.; Ramalho, T.P.; Determann, J.; Duckhorn, M.; Smagin, M.; Avila, M. A Low-Pressure, N2/CO2 Atmosphere Is Suitable for Cyanobacterium-Based Life-Support Systems on Mars. Front. Microbiol. 2021, 12, 611798. [Google Scholar] [CrossRef]
- Hendrickx, L.; De Wever, H.; Hermans, V.; Mastroleo, F.; Morin, N.; Wilmotte, A.; Janssen, P.; Mergeay, M. Microbial ecology of the closed artificial ecosystem MELiSSA (Micro-Ecological Life Support System Alternative): Reinventing and compartmentalizing the Earth’s food and oxygen regeneration system for long-haul space exploration missions. Res. Microbiol. 2006, 157, 77–86. [Google Scholar] [CrossRef]
- Deng, R.; Chow, T.-J. Hypolipidemic, Antioxidant, and Antiinflammatory Activities of Microalgae Spirulina. Cardiovasc. Ther. 2010, 28, e33–e45. [Google Scholar] [CrossRef]
- Yousefi, R.; Saidpour, A.; Mottaghi, A. The effects of Spirulina supplementation on metabolic syndrome components, its liver manifestation and related inflammatory markers: A systematic review. Complement. Ther. Med. 2019, 42, 137–144. [Google Scholar] [CrossRef]
- Kaushik, P.; Chauhan, A. In vitro antibacterial activity of laboratory grown culture of Spirulina platensis. Indian J. Microbiol. 2008, 48, 348–352. [Google Scholar] [CrossRef]
- Parada, J.L.; Zulpa de Caire, G.; Zaccaro de Mulé, M.C.; Storni de Cano, M.M. Lactic acid bacteria growth promoters from Spirulina platensis. Int. J. Food Microbiol. 1998, 45, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.D. The Gut Microbiome and Its Role in Obesity. Nutr. Today 2016, 51, 167–174. [Google Scholar] [CrossRef]
- Schroeder Bjoern, O.; Bäckhed, F. Signals from the gut microbiota to distant organs in physiology and disease. Nat. Med. 2016, 22, 1079–1089. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.K.; Yang, Y.S.; Chen, Y.; Yuan, J.; Sun, G.; Peng, L.H. Intestinal microbiota pathogenesis and fecal microbiota transplantation for inflammatory bowel disease. World J. Gastroenterol. 2014, 20, 14805–14820. [Google Scholar] [CrossRef] [PubMed]
- Dhalla, N.S.; Temsah, R.M.; Netticadan, T. Role of oxidative stress in cardiovascular diseases. J. Hypertens. 2000, 18, 655–673. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, D. Low density lipoprotein oxidation and its pathobiological significance. J. Biol. Chem. 1997, 272, 20963–20966. [Google Scholar] [CrossRef]
- Chisolm, G.M.; Steinberg, D. The oxidative modification hypothesis of atherogenesis: An overview. Free. Radic. Biol. Med. 2000, 28, 1815–1826. [Google Scholar] [CrossRef]
- Lusis, A.J. Atherosclerosis. Nature 2000, 407, 233–241. [Google Scholar] [CrossRef]
- Glass, C.K.; Witztum, J.L. Atherosclerosis. the road ahead. Cell 2001, 104, 503–516. [Google Scholar] [CrossRef]
- Arthur-Ataam, J.; Bideaux, P.; Charrabi, A.; Sicard, P.; Fromy, B.; Liu, K.; Eddahibi, S.; Pasqualin, C.; Jouy, N.; Richard, S.; et al. Dietary supplementation with silicon-enriched spirulina improves arterial remodeling and function in hypertensive rats. Nutrients 2019, 11, 2574. [Google Scholar] [CrossRef]
- Romay, C.; Delgado, R.; Remirez, D.; González, R.; Rojas, A. Effects of phycocyanin extract on tumor necrosis factor-alpha and nitrite levels in serum of mice treated with endotoxin. Arzneimittel-Forschung 2001, 51, 733–736. [Google Scholar] [CrossRef]
- Schafer, F.Q.; Wang, H.P.; Kelley, E.E.; Cueno, K.L.; Martin, S.M.; Buettner, G.R. Comparing beta-carotene, vitamin E and nitric oxide as membrane antioxidants. Biol. Chem. 2002, 383, 671–681. [Google Scholar] [CrossRef]
- Yung, L.M.; Leung, F.P.; Yao, X.; Chen, Z.Y.; Huang, Y. Reactive oxygen species in vascular wall. Cardiovasc. Hematol. Disord. Drug Targets 2006, 6, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Dartsch, P.C. Antioxidant potential of selected Spirulina platensis preparations. Phytother. Res. PTR 2008, 22, 627–633. [Google Scholar] [CrossRef]
- Grosshagauer, S.; Kraemer, K.; Somoza, V. The True Value of Spirulina. J. Agric. Food Chem. 2020, 68, 4109–4115. [Google Scholar] [CrossRef] [PubMed]
- Mao, T.K.; van de Water, J.; Gershwin, M.E. Effects of a Spirulina-based dietary supplement on cytokine production from allergic rhinitis patients. J. Med. Food 2005, 8, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.; Choi, W.Y.; Park, A.; Lee, Y.J.; Lee, Y.; Park, G.H.; Lee, S.J.; Lee, W.K.; Ryu, Y.K.; Kang, D.H. Marine cyanobacterium Spirulina maxima as an alternate to the animal cell culture medium supplement. Sci. Rep. 2021, 11, 4906. [Google Scholar] [CrossRef]
- Abd El-Baky, H.H.; El-Baroty, G.S. Spirulina maxima L-asparaginase: Immobilization, Antiviral and Antiproliferation Activities. Recent Pat Biotechnol. 2020, 14, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Ratha, S.K.; Renuka, N.; Rawat, I.; Bux, F. Prospective options of algae-derived nutraceuticals as supplements to combat COVID-19 and human coronavirus diseases. Nutrition 2021, 83, 111089. [Google Scholar] [CrossRef]
- Hernández-Corona, A.; Nieves, I.; Meckes, M.; Chamorro, G.; Barron, B.L. Antiviral activity of Spirulina maxima against herpes simplex virus type 2. Antiviral Res. 2002, 56, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.S.; Ngo, D.H.; Kang, K.H.; Park, S.J.; Kim, S.K. The role of peptides derived from Spirulina maxima in downregulation of FcεRI-mediated allergic responses. Mol. Nutr. Food Res. 2014, 58, 2226–2234. [Google Scholar] [CrossRef]
- Yang, L.; Wang, Y.; Zhou, Q.; Chen, P.; Wang, Y.; Wang, Y.; Liu, T.; Xie, L. Inhibitory effects of polysaccharide extract from Spirulina platensis on corneal neovascularization. Mol. Vis. 2009, 15, 1951–1961. [Google Scholar]
- Yang, L.L.; Zhou, Q.J.; Wang, Y.; Gao, Y.; Wang, Y.Q. Comparison of the therapeutic effects of extracts from Spirulina platensis and amnion membrane on inflammation-associated corneal neovascularization. Int. J. Ophthalmol. 2012, 5, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Atilgan, H.I.; Akbulut, A.; Yazihan, N.; Yumusak, N.; Singar, E.; Koca, G.; Korkmaz, M. The Cytokines-Directed Roles of Spirulina for Radioprotection of Lacrimal Gland. Ocul. Immunol. Inflamm. 2022, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, T.; Kawashima, H.; Osada, H.; Toda, E.; Homma, K.; Nagai, N.; Imai, Y.; Tsubota, K.; Ozawa, Y. Dietary spirulina supplementation protects visual function from photostress by suppressing retinal neurodegeneration in mice. Transl. Vis. Sci. Technol. 2019, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Amato, R.; Canovai, A.; Melecchi, A.; Pezzino, S.; Corsaro, R.; Dal Monte, M.; Rusciano, D.; Bagnoli, P.; Cammalleri, M. Dietary Supplementation of Antioxidant Compounds Prevents Light-Induced Retinal Damage in a Rat Model. Biomedicines 2021, 9, 1177. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.M.; Jo, Y.D.; Choung, S.Y. Protective Effects of Spirulina maxima against Blue Light-Induced Retinal Damages in A2E-Laden ARPE-19 Cells and Balb/c Mice. Nutrients 2022, 14, 401. [Google Scholar] [CrossRef]
- Kefayat, A.; Ghahremani, F.; Safavi, A.; Hajiaghababa, A.; Moshtaghian, J. Spirulina extract enriched for Braun-type lipoprotein (Immulina®) for inhibition of 4T1 breast tumors’ growth and metastasis. Phyther. Res. 2020, 34, 368–378. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, Y.; Liu, G.; Liu, H.; Zhu, F.; Ji, H.; Li, B. C-Phycocyanin exerts anti-cancer effects via the MAPK signaling pathway in MDA-MB-231 cells. Cancer Cell Int. 2018, 18, 12. [Google Scholar] [CrossRef]
- Subramaiam, H.; Chu, W.L.; Radhakrishnan, A.K.; Chakravarthi, S.; Selvaduray, K.R.; Kok, Y.Y. Evaluating anticancer and immunomodulatory effects of spirulina (Arthrospira) platensis and gamma-tocotrienol supplementation in a syngeneic mouse model of breast cancer. Nutrients 2021, 13, 2320. [Google Scholar] [CrossRef]
- Mahmoud, Y.I.; Shehata, A.M.M.; Fares, N.H.; Mahmoud, A.A. Spirulina inhibits hepatocellular carcinoma through activating p53 and apoptosis and suppressing oxidative stress and angiogenesis. Life Sci. 2021, 265, 118827. [Google Scholar] [CrossRef]
- Czerwonka, A.; Kaławaj, K.; Sławińska-Brych, A.; Lemieszek, M.K.; Bartnik, M.; Wojtanowski, K.K.; Zdzisińska, B.; Rzeski, W. Anticancer effect of the water extract of a commercial Spirulina (Arthrospira platensis) product on the human lung cancer A549 cell line. Biomed. Pharmacother. 2018, 106, 292–302. [Google Scholar] [CrossRef]
- Kennedy, D.O. B vitamins and the brain: Mechanisms, dose and efficacy—A review. Nutrients 2016, 8, 68. [Google Scholar] [CrossRef] [PubMed]
- Rosa, G.P.; Tavares, W.R.; Sousa, P.M.C.; Pagès, A.K.; Seca, A.M.L.; Pinto, D.C.G.A. Seaweed secondary metabolites with beneficial health effects: An overview of successes in in vivo studies and clinical trials. Mar. Drugs 2020, 18, 8. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.; Valado, A. The Seaweed Diet in Prevention and Treatment of the. Mar. Drugs 2021, 19, 128. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.S.; Cintra, R.G.; Barros, S.B.M.; Mancini-Filho, J. Antioxidant activity of the microalga Spirulina maxima. Braz. J. Med. Biol. Res. 1998, 31, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Cornish, M.L.; Critchley, A.T.; Mouritsen, O.G. Consumption of seaweeds and the human brain. J. Appl. Phycol. 2017, 29, 2377–2398. [Google Scholar] [CrossRef]
- Makkar, R.; Behl, T.; Bungau, S.; Zengin, G.; Mehta, V.; Kumar, A.; Uddin, M.S.; Ashraf, G.M.; Abdel-Daim, M.M.; Arora, S.; et al. Nutraceuticals in neurological disorders. Int. J. Mol. Sci. 2020, 21, 4424. [Google Scholar] [CrossRef] [PubMed]
- Morais, L.H.; Schreiber, H.L.; Mazmanian, S.K. The gut microbiota–brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef]
- Sorrenti, V.; Castagna, D.A.; Fortinguerra, S.; Buriani, A.; Scapagnini, G.; Willcox, D.C. Spirulina microalgae and brain health: A scoping review of experimental and clinical evidence. Mar. Drugs 2021, 19, 293. [Google Scholar] [CrossRef]
- Hu, J.; Li, Y.; Pakpour, S.; Wang, S.; Pan, Z.; Liu, J.; Wei, Q.; She, J.; Cang, H.; Zhang, R.X. Dose Effects of Orally Administered Spirulina Suspension on Colonic Microbiota in Healthy Mice. Front. Cell. Infect. Microbiol. 2019, 9, 243. [Google Scholar] [CrossRef]
- Neyrinck, A.M.; Taminiau, B.; Walgrave, H.; Daube, G.; Cani, P.D.; Bindels, L.B.; Delzenne, N.M. Spirulina protects against hepatic inflammation in aging: An effect related to the modulation of the gut microbiota? Nutrients 2017, 9, 633. [Google Scholar] [CrossRef]
- Westfall, S.; Lomis, N.; Kahouli, I.; Dia, S.Y.; Singh, S.P.; Prakash, S. Microbiome, probiotics and neurodegenerative diseases: Deciphering the gut brain axis. Cell. Mol. Life Sci. 2017, 74, 3769–3787. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.H.; Lee, I.-T.; Jeng, K.C.; Wang, M.F.; Hou, R.C.W.; Wu, S.M.; Chan, Y.C. Spirulina prevents memory dysfunction, reduces oxidative stress damage and augments antioxidant activity in senescence-accelerated mice. J. Nutr. Sci. Vitaminol. 2011, 57, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Pabon, M.M.; Jernberg, J.N.; Morganti, J.; Contreras, J.; Hudson, C.E.; Klein, R.L.; Bickford, P.C. A Spirulina-Enhanced Diet Provides Neuroprotection in an α-Synuclein Model of Parkinson’s Disease. PLoS ONE 2012, 7, e90972. [Google Scholar] [CrossRef]
- Lima, F.A.V.; Joventino, I.P.; Joventino, F.P.; de Almeida, A.C.; Neves, K.R.T.; do Carmo, M.R.; Leal, L.K.A.M.; de Andrade, G.M.; de Barros Viana, G.S. Neuroprotective Activities of Spirulina platensis in the 6-OHDA Model of Parkinson’s Disease Are Related to Its Anti-Inflammatory Effects. Neurochem. Res. 2017, 42, 3390–3400. [Google Scholar] [CrossRef]
- Koh, E.J.; Kim, K.J.; Song, J.H.; Choi, J.; Lee, H.Y.; Kang, D.H.; Heo, H.J.; Lee, B.Y. Spirulina maxima extract ameliorates learning and memory impairments via inhibiting GSK-3β phosphorylation induced by intracerebroventricular injection of amyloid-β 1-42 in mice. Int. J. Mol. Sci. 2017, 18, 2401. [Google Scholar] [CrossRef]
- Moradi-Kor, N.; Ghanbari, A.; Rashidipour, H.; Bandegi, A.R.; Yousefi, B.; Barati, M.; Kokhaei, P.; Rashidy-Pour, A. Therapeutic effects of spirulina platensis against adolescent stress-induced oxidative stress, brain-derived neurotrophic factor alterations and morphological remodeling in the amygdala of adult female rats. J. Exp. Pharmacol. 2020, 12, 75–85. [Google Scholar] [CrossRef]
- Park, H.J.; Lee, Y.J.; Ryu, H.K.; Kim, M.H.; Chung, H.W.; Kim, W.Y. A randomized double-blind, placebo-controlled study to establish the effects of spirulina in elderly Koreans. Ann. Nutr. Metab. 2008, 52, 322–328. [Google Scholar] [CrossRef]
- Reid, S.N.S.; Ryu, J.K.; Kim, Y.; Jeon, B.H. The Effects of Fermented Laminaria japonica on Short-Term Working Memory and Physical Fitness in the Elderly. Evidence-based Complement. Altern. Med. 2018, 2018, 8109621. [Google Scholar] [CrossRef]
- Haskell-Ramsay, C.F.; Jackson, P.A.; Dodd, F.L.; Forster, J.S.; Bérubé, J.; Levinton, C.; Kennedy, D.O. Acute post-prandial cognitive effects of Brown seaweed extract in humans. Nutrients 2018, 10, 85. [Google Scholar] [CrossRef]
- Pentón-Rol, G.; Lagumersindez-Denis, N.; Muzio, L.; Bergami, A.; Furlan, R.; Fernández-Massó, J.R.; Nazabal-Galvez, M.; Llópiz-Arzuaga, A.; Herrera-Rolo, T.; Veliz-Rodriguez, T.; et al. Comparative Neuroregenerative Effects of C-Phycocyanin and IFN-Beta in a Model of Multiple Sclerosis in Mice. J. Neuroimmune Pharmacol. 2016, 11, 153–167. [Google Scholar] [CrossRef]
- Heppner, F.L.; Greter, M.; Marino, D.; Falsig, J.; Raivich, G.; Hövelmeyer, N.; Waisman, A.; Rülicke, T.; Prinz, M.; Priller, J.; et al. Experimental autoimmune encephalomyelitis repressed by microglial paralysis. Nat. Med. 2005, 11, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Pattern Recognition Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed]
- Monk, C.; Georgieff, M.K.; Osterholm, E.A. Research Review: Maternal prenatal distress and poor nutrition—Mutually influencing risk factors affecting infant neurocognitive development. J. Child Psychol. Psychiatry Allied Discip. 2013, 54, 115–130. [Google Scholar] [CrossRef]
- Sinha, S.; Patro, N.; Tiwari, P.K.; Patro, I.K. Maternal Spirulina supplementation during pregnancy and lactation partially prevents oxidative stress, glial activation and neuronal damage in protein malnourished F1 progeny. Neurochem. Int. 2020, 141, 104877. [Google Scholar] [CrossRef] [PubMed]
- Patro, N.; Sharma, A.; Kariaya, K.; Patro, I. Spirulina platensis protects neurons via suppression of glial activation and peripheral sensitization leading to restoration of motor function in collagen-induced arthritic rats. Indian J. Exp. Biol. 2011, 49, 739–748. [Google Scholar] [PubMed]
- Tzachor, A.; Rozen, O.; Khatib, S.; Jensen, S.; Avni, D. Photosynthetically Controlled Spirulina, but Not Solar Spirulina, Inhibits TNF-α Secretion: Potential Implications for COVID-19-Related Cytokine Storm Therapy. Mar. Biotechnol. 2021, 23, 149–155. [Google Scholar] [CrossRef]
- Baindara, P.; Chakraborty, R.; Holliday, Z.M.; Mandal, S.M.; Schrum, A.G. Oral probiotics in coronavirus disease 2019: Connecting the gut-lung axis to viral pathogenesis, inflammation, secondary infection and clinical trials. New Microbes New Infect. 2021, 40, 100837. [Google Scholar] [CrossRef]
- Arab, S.; Ghasemi, S.; Ghanbari, A.; Bahraminasab, M.; Satari, A.; Mousavi, M.; Dehcheshme, H.G.; Asgharzade, S. Chemopreventive effect of spirulina microalgae on an animal model of glioblastoma via down-regulation of PI3K/AKT/mTOR and up-regulation of miR-34a/miR-125B expression. Phyther. Res. 2021, 35, 6452–6461. [Google Scholar] [CrossRef]
- Díaz Domínguez, G.; Marsán Suárez, V.; del Valle Pérez, L.O. Principales propiedades inmunomoduladoras y antinflamatorias de la ficobiliproteína C-ficocianina. Rev. Cuba. Hematol. Inmunol. Hemoter. 2016, 32, 447–454. [Google Scholar]
- Ponce-Canchihuamán, J.C.; Pérez-Méndez, O.; Hernández-Mũoz, R.; Torres-Durán, P.V.; Juárez-Oropeza, M.A. Protective effects of Spirulina maxima on hyperlipidemia and oxidative-stress induced by lead acetate in the liver and kidney. Lipids Health Dis. 2010, 9, 35. [Google Scholar] [CrossRef]
- Oh, S.H.; Ahn, J.; Kang, D.H.; Lee, H.Y. The Effect of Ultrasonificated Extracts of Spirulina maxima on the Anticancer Activity. Mar. Biotechnol. 2011, 13, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Koh, E.J.; Seo, Y.J.; Choi, J.; Lee, H.Y.; Kang, D.H.; Kim, K.J.; Lee, B.Y. Spirulina maxima extract prevents neurotoxicity via promoting activation of BDNF/CREB signaling pathways in neuronal cells and mice. Molecules 2017, 22, 1363. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Park, A.; Kim, M.J.; Lim, H.J.; Rha, Y.A.; Kang, H.G. Spirulina extract enhanced a protective effect in type 1 diabetes by anti-apoptosis and anti-ROS production. Nutrients 2017, 9, 1363. [Google Scholar] [CrossRef] [PubMed]
- Bachstetter, A.D.; Jernberg, J.; Schlunk, A.; Vila, J.L.; Hudson, C.; Cole, M.J.; Shytle, R.D.; Tan, J.; Sanberg, P.R.; Sanberg, C.D.; et al. Spirulina promotes stem cell genesis and protects against LPS induced declines in neural stem cell proliferation. PLoS ONE 2010, 5, e10496. [Google Scholar] [CrossRef] [PubMed]
- Steffens, D.; Lersch, M.; Rosa, A.; Scher, C.; Crestani, T.; Morais, M.G.; Costa, J.A.V.; Pranke, P. A new biomaterial of nanofibers with the microalga spirulina as scaffolds to cultivate with stem cells for use in tissue engineering. J. Biomed. Nanotechnol. 2013, 9, 710–718. [Google Scholar] [CrossRef]
- Bickford, P.C.; Tan, J.; Douglas Shytle, R.; Sanberg, C.D.; El-Badri, N.; Sanberg, P.R. Nutraceuticals synergistically promote proliferation of human stem cells. Stem Cells Dev. 2006, 15, 118–123. [Google Scholar] [CrossRef]
- Jensen, G.S.; Hart, A.N.; Zaske, L.A.M.; Drapeau, C.; Gupta, N.; Schaeffer, D.J.; Cruickshank, J.A. Mobilization of human CD34+CD133+ and CD34+CD133- stem cells in vivo by consumption of an extract from Aphanizomenon flos-aquae-related to modulation of CXCR4 expression by an L-selectin ligand? Cardiovasc. Revascularization Med. 2007, 8, 189–202. [Google Scholar] [CrossRef]
- Kuhad, A.; Tirkey, N.; Pilkhwal, S.; Chopra, K. Renoprotective effect of Spirulina fusiformis on cisplatin-induced oxidative stress and renal dysfunction in rats. Ren. Fail. 2006, 28, 247–254. [Google Scholar] [CrossRef]
- Avdagić, N.; Ćosović, E.; Nakaš-Ičindič, E.; Mornjaković, Z.; Začiragić, A.; Hadžovič-Džuvo, A. Spirulina platensis protects against renal injury in rats with gentamicin-induced acute tubular necrosis. Bosn. J. Basic Med. Sci. 2008, 8, 331–336. [Google Scholar] [CrossRef]
- Hamad, E.M.; Mousa, H.M.; Ashoush, I.S.; Abdel-Salam, A.M. Nephroprotective effect of camel milk and spirulina platensis in gentamicin-induced nephrotoxicity in rats. Int. J. Pharmacol. 2018, 14, 559–565. [Google Scholar] [CrossRef]
- Khan, M.; Shobha, J.C.; Mohan, I.K.; Rao Naidu, M.U.; Prayag, A.; Kutala, V.K. Spirulina attenuates cyclosporine-induced nephrotoxicity in rats. J. Appl. Toxicol. 2006, 27, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Carbajal, M.C.; Torres-Duràn, P.V.; Dìaz-Zagoya, J.C.; Mascher, D.; Juárez-Oropeza, M.A. Effects of dietary spirulina maxima on endothelium dependent vasomotor responses of rat aortic rings. Life Sci. 1997, 61, 211–219. [Google Scholar] [CrossRef]
- Memije-Lazaro, I.N.; Blas-Valdivia, V.; Franco-Colín, M.; Cano-Europa, E. Arthrospira maxima (Spirulina) and C-phycocyanin prevent the progression of chronic kidney disease and its cardiovascular complications. J. Funct. Foods 2018, 43, 37–43. [Google Scholar] [CrossRef]
- Kovic, M. Risks of space colonization. Futures 2021, 126, 102638. [Google Scholar] [CrossRef]
- Nangle, S.N.; Wolfson, M.Y.; Hartsough, L.; Ma, N.J.; Mason, C.E.; Merighi, M.; Nathan, V.; Silver, P.A.; Simon, M.; Swett, J.; et al. The case for biotech on Mars. Nat. Biotechnol. 2020, 38, 401–407. [Google Scholar] [CrossRef]
- Revellame, E.D.; Aguda, R.; Chistoserdov, A.; Fortela, D.L.; Hernandez, R.A.; Zappi, M.E. Microalgae cultivation for space exploration: Assessing the potential for a new generation of waste to human life-support system for long duration space travel and planetary human habitation. Algal Res. 2021, 55, 102258. [Google Scholar] [CrossRef]
- Belz, S.; Buchert, M.; Bretschneider, J.; Nathanson, E.; Fasoulas, S. Physicochemical and biological technologies for future exploration missions. Acta Astronaut. 2014, 101, 170–179. [Google Scholar] [CrossRef]
- Cycil, L.M.; Hausrath, E.M.; Ming, D.W.; Adcock, C.T.; Raymond, J.; Remias, D.; Ruemmele, W.P. Investigating the Growth of Algae Under Low Atmospheric Pressures for Potential Food and Oxygen Production on Mars. Front. Microbiol. 2021, 12, 733244. [Google Scholar] [CrossRef]
- Yang, C.; Liu, H.; Li, M.; Yu, C.; Yu, G. Treating urine by Spirulina platensis. Acta Astronaut. 2008, 63, 1049–1054. [Google Scholar] [CrossRef]
- Rapp, D. Use of Extraterrestrial Resources for Human Space Missions to Moon or Mars; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar] [CrossRef]
- De Man, P. In-Situ Resource Utilization: Legal Aspects. In Promoting Productive Cooperation Between Space Lawyers and Engineers; Pecujlic, A.N., Tugnoli, M., Eds.; IGI Global: Hershey, PA, USA, 2019; pp. 211–224. [Google Scholar] [CrossRef]
- Olsson-Francis, K.; Cockell, C.S. Use of cyanobacteria for in-situ resource use in space applications. Planet. Space Sci. 2010, 58, 1279–1285. [Google Scholar] [CrossRef]
- Billi, D.; Gallego Fernandez, B.; Fagliarone, C.; Chiavarini, S.; Rothschild, L.J. Exploiting a perchlorate-tolerant desert cyanobacterium to support bacterial growth for in situ resource utilization on Mars. Int. J. Astrobiol. 2021, 20, 29–35. [Google Scholar] [CrossRef]
- Cao, G.; Concas, A.; Fais, G.; Gabrielli, G.; Manca, A.; Pantaleo, A. Process and Kit to Investigate Microgravity Effect on Animal/Vegetable Cells under Extraterrestrial Cultivation Conditions and Cultivation Process thereof to Sustain Manned Space Missions 2021. PCT/IB2012/053754, 2021. Available online: https://patentscope2.wipo.int/search/en/detail.jsf?docId=WO2013014606 (accessed on 10 March 2022).
- Nickerson, C.A.; Ott, C.M.; Wilson, J.W.; Ramamurthy, R.; Pierson, D.L. Microbial Responses to Microgravity and Other Low-Shear Environments. Microbiol. Mol. Biol. Rev. 2004, 68, 345–361. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.W.; Ott, C.M.; Ramamurthy, R.; Quick, L.; Porwollik, S.; Cheng, P.; Mcclelland, M.; Ho, K.; Tsaprailis, G.; Radabaugh, T.; et al. Space flight alters bacterial gene expression and virulence and reveals a role for global regulator Hfq. Proc. Natl. Acad. Sci. USA 2007, 104, 16299–16304. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.W.; Ott, C.M.; Quick, L.; Davis, R.; zu Bentrup, K.H.; Crabbé, A.; Richter, E.; Sarker, S.; Barrila, J.; Porwollik, S.; et al. Media ion composition controls regulatory and virulence response of Salmonella in spaceflight. PLoS ONE 2008, 3, e3923. [Google Scholar] [CrossRef] [PubMed]
- Barrila, J.; Radtke, A.L.; Crabbé, A.; Sarker, S.F.; Herbst-Kralovetz, M.M.; Ott, C.M.; Nickerson, C.A. Organotypic 3D cell culture models: Using the rotating wall vessel to study host-pathogen interactions. Nat. Rev. Microbiol. 2010, 8, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.T.; Dowd, S.E.; Galley, J.D.; Hufnagle, A.R.; Allen, R.G.; Lyte, M. Exposure to a social stressor alters the structure of the intestinal microbiota: Implications for stressor-induced immunomodulation. Brain. Behav. Immun. 2011, 25, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Castro, S.L.; Nelman-Gonzalez, M.; Nickerson, C.A.; Ott, C.M. Induction of attachment-independent biofilm formation and repression of hfq expression by low-fluid-shear culture of Staphylococcus aureus. Appl. Environ. Microbiol. 2011, 77, 6368–6378. [Google Scholar] [CrossRef]
- Crabbé, A.; Schurr, M.J.; Monsieurs, P.; Morici, L.; Schurr, J.; Wilson, J.W.; Ott, C.M.; Tsaprailis, G.; Pierson, D.L.; Stefanyshyn-Piper, H.; et al. Transcriptional and proteomic responses of Pseudomonas aeruginosa PAO1 to spaceflight conditions involve Hfq regulation and reveal a role for oxygen. Appl. Environ. Microbiol. 2011, 77, 1221–1230. [Google Scholar] [CrossRef]
- Turroni, S.; Magnani, M.; KC, P.; Lesnik, P.; Vidal, H.; Heer, M. Gut Microbiome and Space Travelers’ Health: State of the Art and Possible Pro/Prebiotic Strategies for Long-Term Space Missions. Front. Physiol. 2020, 11, 553929. [Google Scholar] [CrossRef]
- Crucian, B.; Sams, C. Immune system dysregulation during spaceflight: Clinical risk for exploration-class missions. J. Leukoc. Biol. 2009, 86, 1017–1018. [Google Scholar] [CrossRef]
- Siddiqui, R.; Akbar, N.; Khan, N.A. Gut microbiome and human health under the space environment. J. Appl. Microbiol. 2021, 130, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.; Wu, H.; Choi, J.U.; Rowan, A.E.; Zhang, H.; Poole, K.; Lauko, J.; Chou, J. Modeling the Impact of Microgravity at the Cellular Level: Implications for Human Disease. Front. Cell Dev. Biol. 2020, 8, 96. [Google Scholar] [CrossRef] [PubMed]
- Da Silveira, W.A.; Fazelinia, H.; Rosenthal, S.B.; Laiakis, E.C.; Kim, M.S.; Meydan, C.; Kidane, Y.; Rathi, K.S.; Smith, S.M.; Stear, B.; et al. Comprehensive Multi-omics Analysis Reveals Mitochondrial Stress as a Central Biological Hub for Spaceflight Impact. Cell 2020, 183, 1185–1201.e20. [Google Scholar] [CrossRef]
- Powers, S.K.; Ji, L.L.; Kavazis, A.N.; Jackson, M.J. Reactive oxygen species: Impact on skeletal muscle. Compr. Physiol. 2011, 1, 941–969. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Villanueva, M.; Wong, M.; Lu, T.; Zhang, Y.; Wu, H. Interplay of space radiation and microgravity in DNA damage and DNA damage response. NPJ Microgravity 2017, 3, 14. [Google Scholar] [CrossRef] [PubMed]
- Richter, C. Reactive oxygen and DNA damage in mitochondria. Mutat. Res. DNAging 1992, 275, 249–255. [Google Scholar] [CrossRef]
- Goodwin, T.J.; Christofidou-Solomidou, M. Oxidative Stress and Space Biology: An Organ-Based Approach. Int. J. Mol. Sci. 2018, 19, 959. [Google Scholar] [CrossRef]
- Schimmerling, W. Space and radiation protection: Scientific requirements for space research. Radiat. Environ. Biophys. 1995, 34, 133–137. [Google Scholar] [CrossRef]
- Belobrajdic, B.; Melone, K.; Diaz-Artiles, A. Planetary extravehicular activity (EVA) risk mitigation strategies for long-duration space missions. NPJ Microgravity 2021, 7, 16. [Google Scholar] [CrossRef]
- Buonanno, M.; De Toledo, S.M.; Howell, R.W.; Azzam, E.I. Low-dose energetic protons induce adaptive and bystander effects that protect human cells against DNA damage caused by a subsequent exposure to energetic iron ions. J. Radiat. Res. 2015, 56, 502–508. [Google Scholar] [CrossRef]
- Bigley, A.B.; Agha, N.H.; Baker, F.L.; Spielmann, G.; Kunz, H.E.; Mylabathula, P.L.; Rooney, B.V.; Laughlin, M.S.; Mehta, S.K.; Pierson, D.L.; et al. NK cell function is impaired during long-duration spaceflight. J. Appl. Physiol. 2019, 126, 842–853. [Google Scholar] [CrossRef] [PubMed]
- Crucian, B.; Stowe, R.P.; Mehta, S.; Quiriarte, H.; Pierson, D.; Sams, C. Alterations in adaptive immunity persist during long-duration spaceflight. NPJ Microgravity 2015, 1, 15013. [Google Scholar] [CrossRef] [PubMed]
- Martinez, E.M.; Yoshida, M.C.; Candelario, T.L.T.; Hughes-Fulford, M. Spaceflight and simulated microgravity cause a significant reduction of key gene expression in early T-cell activation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R480–R488. [Google Scholar] [CrossRef] [PubMed]
- Pecaut, M.J.; Mao, X.W.; Bellinger, D.L.; Jonscher, K.R.; Stodieck, L.S.; Ferguson, V.L.; Bateman, T.A.; Mohney, R.P.; Gridley, D.S. Is spaceflight-induced immune dysfunction linked to systemic changes in metabolism? PLoS ONE 2017, 12, e0174174. [Google Scholar] [CrossRef]
- Paul, A.M.; Cheng-Campbell, M.; Blaber, E.A.; Anand, S.; Bhattacharya, S.; Zwart, S.R.; Crucian, B.E.; Smith, S.M.; Meller, R.; Grabham, P.; et al. Beyond Low-Earth Orbit: Characterizing Immune and microRNA Differentials following Simulated Deep Spaceflight Conditions in Mice. iScience 2020, 23, 101747. [Google Scholar] [CrossRef]
- Pavlakou, P.; Dounousi, E.; Roumeliotis, S.; Eleftheriadis, T.; Liakopoulos, V. Oxidative Stress and the Kidney in the Space Environment. Int. J. Mol. Sci. 2018, 19, 3176. [Google Scholar] [CrossRef]
- Roumelioti, M.-E.; Glew, R.H.; Khitan, Z.J.; Rondon-Berrios, H.; Argyropoulos, C.P.; Malhotra, D.; Raj, D.S.; Agaba, E.I.; Rohrscheib, M.; Murata, G.H.; et al. Fluid balance concepts in medicine: Principles and practice. World J. Nephrol. 2018, 7, 1–28. [Google Scholar] [CrossRef]
- Drummer, C.; Gerzer, R.; Baisch, F.; Heer, M. Body fluid regulation in micro-gravity differs from that on Earth: An overview. Pflugers Arch. 2000, 441, R66–R72. [Google Scholar] [CrossRef]
- Whitson, P.A.; Pietrzyk, R.A.; Jones, J.A.; Nelman-Gonzalez, M.; Hudson, E.K.; Sams, C.F. Effect of potassium citrate therapy on the risk of renal stone formation during spaceflight. J. Urol. 2009, 182, 2490–2496. [Google Scholar] [CrossRef]
- Pietrzyk, R.A.; Jones, J.A.; Sams, C.F.; Whitson, P.A. Renal stone formation among astronauts. Aviat. Space. Environ. Med. 2007, 78 (Suppl. S4), A9–A13. [Google Scholar]
- Liakopoulos, V.; Leivaditis, K.; Eleftheriadis, T.; Dombros, N. The kidney in space. Int. Urol. Nephrol. 2012, 44, 1893–1901. [Google Scholar] [CrossRef] [PubMed]
- Barcellos-Hoff, M.H.; Blakely, E.A.; Burma, S.; Fornace, A.J.; Gerson, S.; Hlatky, L.; Kirsch, D.G.; Luderer, U.; Shay, J.; Wang, Y.; et al. Concepts and challenges in cancer risk prediction for the space radiation environment. Life Sci. Sp. Res. 2015, 6, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Kim, M.H.Y.; McClellan, G.E.; Cucinotta, F.A. Modeling the acute health effects of astronauts from exposure to large solar particle events. Health Phys. 2009, 96, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Patel, S. The effects of microgravity and space radiation on cardiovascular health: From low-Earth orbit and beyond. IJC Heart Vasc. 2020, 30, 100595. [Google Scholar] [CrossRef] [PubMed]
- Melissa Gaskill Vascular Studies in Space: Good Heart, Everyone’s; 2021. p. 166. Available online: https://www.nasa.gov/sites/default/files/atoms/files/benefits-for-humanity_third.pdf (accessed on 7 March 2022).
- Meyers, C.A. Neurocognitive dysfunction in cancer patients. Oncology (Williston Park) 2000, 14, 75–85. [Google Scholar] [PubMed]
- Jandial, R.; Hoshide, R.; Waters, J.D.; Limoli, C.L. Space-brain: The negative effects of space exposure on the central nervous system. Surg. Neurol. Int. 2018, 9, 9. [Google Scholar] [CrossRef]
- Andersen, B.L.; Tewfik, H.H. Psychological reactions to radiation therapy: Reconsideration of the adaptive aspects of anxiety. J. Pers. Soc. Psychol. 1985, 48, 1024–1032. [Google Scholar] [CrossRef]
- Makale, M.T.; McDonald, C.R.; Hattangadi-Gluth, J.A.; Kesari, S. Mechanisms of radiotherapy-associated cognitive disability in patients with brain tumours. Nat. Rev. Neurol. 2017, 13, 52–64. [Google Scholar] [CrossRef]
- Tang, Y.; Luo, D.; Rong, X.; Shi, X.; Peng, Y. Psychological disorders, cognitive dysfunction and quality of life in nasopharyngeal carcinoma patients with radiation-induced brain injury. PLoS ONE 2012, 7, e36529. [Google Scholar] [CrossRef]
- Wellisch, D.K.; Kaleita, T.A.; Freeman, D.; Cloughesy, T.; Goldman, J. Predicting major depression in brain tumor patients. Psychooncology 2002, 11, 230–238. [Google Scholar] [CrossRef]
- Parihar, V.K.; Allen, B.D.; Caressi, C.; Kwok, S.; Chu, E.; Tran, K.K.; Chmielewski, N.N.; Giedzinski, E.; Acharya, M.M.; Britten, R.A.; et al. Cosmic radiation exposure and persistent cognitive dysfunction. Sci. Rep. 2016, 6, 34774. [Google Scholar] [CrossRef] [PubMed]
- Cherry, J.D.; Liu, B.; Frost, J.L.; Lemere, C.A.; Williams, J.P.; Olschowka, J.A.; O’Banion, M.K. Galactic cosmic radiation leads to cognitive impairment and increased aβ plaque accumulation in a mouse model of Alzheimer’s disease. PLoS ONE 2012, 7, e53275. [Google Scholar] [CrossRef] [PubMed]
- Parihar, V.K.; Pasha, J.; Tran, K.K.; Craver, B.M.; Acharya, M.M.; Limoli, C.L. Persistent changes in neuronal structure and synaptic plasticity caused by proton irradiation. Brain Struct. Funct. 2015, 220, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Machida, M.; Lonart, G.; Britten, R.A. Low (60 cGy) doses of (56)Fe HZE-particle radiation lead to a persistent reduction in the glutamatergic readily releasable pool in rat hippocampal synaptosomes. Radiat. Res. 2010, 174, 618–623. [Google Scholar] [CrossRef] [PubMed]
- Gaboyard, S.; Blanchard, M.-P.; Travo, C.; Viso, M.; Sans, A.; Lehouelleur, J. Weightlessness affects cytoskeleton of rat utricular hair cells during maturation in vitro. Neuroreport 2002, 13, 2139–2142. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhang, X.; Gao, Y.; Li, S.; Sun, Y. Effects of altered gravity on the cell cycle, actin cytoskeleton and proteome in Physarum polycephalum. Acta Astronaut. 2008, 63, 915–922. [Google Scholar] [CrossRef]
- Huang, Y.; Dai, Z.-Q.; Ling, S.-K.; Zhang, H.-Y.; Wan, Y.-M.; Li, Y.-H. Gravity, a regulation factor in the differentiation of rat bone marrow mesenchymal stem cells. J. Biomed. Sci. 2009, 16, 87. [Google Scholar] [CrossRef]
- Sarkar, P.; Sarkar, S.; Ramesh, V.; Hayes, B.E.; Thomas, R.L.; Wilson, B.L.; Kim, H.; Barnes, S.; Kulkarni, A.; Pellis, N.; et al. Proteomic analysis of mice hippocampus in simulated microgravity environment. J. Proteome Res. 2006, 5, 548–553. [Google Scholar] [CrossRef][Green Version]
- Clément, G.R.; Boyle, R.D.; George, K.A.; Nelson, G.A.; Reschke, M.F.; Williams, T.J.; Paloski, W.H. Challenges to the central nervous system during human spaceflight missions to Mars. J. Neurophysiol. 2020, 123, 2037–2063. [Google Scholar] [CrossRef]
- Tauber, S.; Hauschild, S.; Paulsen, K.; Gutewort, A.; Raig, C.; Hürlimann, E.; Biskup, J.; Philpot, C.; Lier, H.; Engelmann, F.; et al. Signal transduction in primary human T lymphocytes in altered gravity during parabolic flight and clinostat experiments. Cell. Physiol. Biochem. 2015, 35, 1034–1051. [Google Scholar] [CrossRef]
- Tauber, S.; Hauschild, S.; Crescio, C.; Secchi, C.; Paulsen, K.; Pantaleo, A.; Saba, A.; Buttron, I.; Thiel, C.S.; Cogoli, A.; et al. Signal transduction in primary human T lymphocytes in altered gravity—Results of the MASER-12 suborbital space flight mission. Cell Commun. Signal. 2013, 11, 32. [Google Scholar] [CrossRef]
- Mehta, S.K.; Laudenslager, M.L.; Stowe, R.P.; Crucian, B.E.; Feiveson, A.H.; Sams, C.F.; Pierson, D.L. Latent virus reactivation in astronauts on the international space station. NPJ Microgravity 2017, 3, 11. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Sánchez, J.L.; Callant, J.; Krüger, M.; Sahana, J.; Kraus, A.; Baselet, B.; Infanger, M.; Baatout, S.; Grimm, D. Cancer studies under space conditions: Finding answers abroad. Biomedicines 2022, 10, 25. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, S.; Nagamatsu, A.; Nenoi, M.; Fujimori, A.; Kakinuma, S.; Katsube, T.; Wang, B.; Tsuruoka, C.; Shirai, T.; Nakamura, A.J.; et al. Space Radiation Biology for “Living in Space”. Biomed Res. Int. 2020, 2020, 4703286. [Google Scholar] [CrossRef] [PubMed]
- Jeggo, P.; Löbrich, M. Radiation-induced DNA damage responses. Radiat. Prot. Dosim. 2006, 122, 124–127. [Google Scholar] [CrossRef]
- Santivasi, W.L.; Xia, F. Ionizing radiation-induced DNA damage, response, and repair. Antioxid. Redox Signal. 2014, 21, 251–259. [Google Scholar] [CrossRef]
- Tajvidi, E.; Nahavandizadeh, N.; Pournaderi, M.; Pourrashid, A.Z.; Bossaghzadeh, F.; Khoshnood, Z. Study the antioxidant effects of blue-green algae Spirulina extract on ROS and MDA production in human lung cancer cells. Biochem. Biophys. Rep. 2021, 28, 101139. [Google Scholar] [CrossRef]
- Leach, C.S.; Johnson, P.C. Influence of spaceflight on erythrokinetics in man. Science 1984, 225, 216–218. [Google Scholar] [CrossRef]
- Selmi, C.; Leung, P.S.C.; Fischer, L.; German, B.; Yang, C.Y.; Kenny, T.P.; Cysewski, G.R.; Gershwin, M.E. The effects of Spirulina on anemia and immune function in senior citizens. Cell. Mol. Immunol. 2011, 8, 248–254. [Google Scholar] [CrossRef]
- Mader, T.H.; Gibson, C.R.; Pass, A.F.; Kramer, L.A.; Lee, A.G.; Fogarty, J.; Tarver, W.J.; Dervay, J.P.; Hamilton, D.R.; Sargsyan, A.; et al. Optic disc edema, globe flattening, choroidal folds, and hyperopic shifts observed in astronauts after long-duration space flight. Ophthalmology 2011, 118, 2058–2069. [Google Scholar] [CrossRef]
- Zwart, S.R.; Gibson, C.R.; Mader, T.H.; Ericson, K.; Ploutz-Snyder, R.; Heer, M.; Smith, S.M. Vision Changes after Spaceflight Are Related to Alterations in Folate– and Vitamin B-12–Dependent One-Carbon Metabolism. J. Nutr. 2012, 142, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Ata, F.; Bint, I.; Bilal, A.; Javed, S.; Shabir Chaudhry, H.; Sharma, R.; Fatima Malik, R.; Choudry, H.; Bhaskaran Kartha, A. Optic neuropathy as a presenting feature of vitamin B-12 deficiency: A systematic review of literature and a case report. Ann. Med. Surg. 2020, 60, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Beetstra, S.; Thomas, P.; Salisbury, C.; Turner, J.; Fenech, M. Folic acid deficiency increases chromosomal instability, chromosome 21 aneuploidy and sensitivity to radiation-induced micronuclei. Mutat. Res. 2005, 578, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Moens, A.L.; Vrints, C.J.; Claeys, M.J.; Timmermans, J.P.; Champion, H.C.; Kass, D.A. Mechanisms and potential therapeutic targets for folic acid in cardiovascular disease. Am. J. Physiol.—Heart Circ. Physiol. 2008, 294, 1971–1977. [Google Scholar] [CrossRef] [PubMed]
- Zwart, S.R.; Gibson, C.R.; Gregory, J.F.; Mader, T.H.; Stover, P.J.; Zeisel, S.H.; Smith, S.M. Astronaut ophthalmic syndrome. FASEB J. 2017, 31, 3746–3756. [Google Scholar] [CrossRef]
- Akiyama, T.; Horie, K.; Hinoi, E.; Hiraiwa, M.; Kato, A.; Maekawa, Y.; Takahashi, A.; Furukawa, S. How does spaceflight affect the acquired immune system? NPJ Microgravity 2020, 6, 14. [Google Scholar] [CrossRef]
- Makedonas, G.; Mehta, S.K.; Scheuring, R.A.; Haddon, R.; Crucian, B.E. SARS-CoV-2 Pandemic Impacts on NASA Ground Operations to Protect ISS Astronauts. J. Allergy Clin. Immunol. Pract. 2020, 8, 3247–3250. [Google Scholar] [CrossRef]
- Blaber, E.A.; Finkelstein, H.; Dvorochkin, N.; Sato, K.Y.; Yousuf, R.; Burns, B.P.; Globus, R.K.; Almeida, E.A.C. Microgravity Reduces the Differentiation and Regenerative Potential of Embryonic Stem Cells. Stem Cells Dev. 2015, 24, 2605–2621. [Google Scholar] [CrossRef]
- Blaber, E.; Sato, K.; Almeida, E.A.C. Stem cell health and tissue regeneration in microgravity. Stem Cells Dev. 2014, 23, 73–78. [Google Scholar] [CrossRef]
- Tadros, M.G. Characterization of Spirulina Biomass for CELSS Diet Potential; NASA Contract: NCC 2-501; US Gov.: Washington, DC, USA, 1988; 53p. Available online: https://ntrs.nasa.gov/citations/19890016190 (accessed on 24 April 2022).
- Oguchi, M.; Nitta, K.; Hatayaina, S.; Technology, S. Food production and gas exchange system using blue-green alga (Spirulina) for CELSS. Adv. Space Res. 1987, 7, 7–10. [Google Scholar] [CrossRef]
- García, J.L.; de Vicente, M.; Galán, B. Microalgae, old sustainable food and fashion nutraceuticals. Microb. Biotechnol. 2017, 10, 1017–1024. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Macronutrients | % d.w. |
| Proteins | 50–70 |
| Carbohydrates | 15–20 |
| Lipids | 7–16 |
| Vitamins | mg/100 g d.w. |
| Carotene | 140 |
| Biotin | 0.005 |
| Folic acid | 0.01 |
| Niacin | 14 |
| Riboflavin | 4 |
| Thiamin B1 | 3.5 |
| Vitamin E | 100 |
| Vitamin B12 | 0.32 |
| Vitamin K | 2.2 |
| Carbohydrates | mg/100 g d.w. |
| Glucose | 54.4 |
| Galactose | 2.6 |
| Mannose | 9.3 |
| Rhamnose | 22.3 |
| Xylose | 7 |
| Lipids | g/100 g d.w. |
| Arachidic | 0.048 |
| Gamma linolenic (GLA) | 1 |
| Myristic | 0.041 |
| Oleic | 0.017 |
| Palmitic | 2 |
| Amino acids | g/100 g d.w. |
| Leucine | 4.94 |
| Isoleucine | 3.2 |
| Lysine | 3.02 |
| Methionine | 1.15 |
| Phenylalaline | 2.78 |
| Threonine | 2.97 |
| Tryptophan | 0.93 |
| Valine | 3.51 |
| Phytonutrients | g/100 g d.w. |
| cis beta-carotene | 0.073 |
| Chlorophyll-a | 1 |
| Phycocyanin | 12 |
| trans beta-carotene | 0.26 |
| Minerals | mg/100 g d.w. |
| Calcium | 700 |
| Copper | 1.2 |
| Iron | 100 |
| Magnesium | 400 |
| Manganese | 5 |
| Phosphorus | 800 |
| Potassium | 1400 |
| Sodium | 900 |
| Zinc | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fais, G.; Manca, A.; Bolognesi, F.; Borselli, M.; Concas, A.; Busutti, M.; Broggi, G.; Sanna, P.; Castillo-Aleman, Y.M.; Rivero-Jiménez, R.A.; et al. Wide Range Applications of Spirulina: From Earth to Space Missions. Mar. Drugs 2022, 20, 299. https://doi.org/10.3390/md20050299
Fais G, Manca A, Bolognesi F, Borselli M, Concas A, Busutti M, Broggi G, Sanna P, Castillo-Aleman YM, Rivero-Jiménez RA, et al. Wide Range Applications of Spirulina: From Earth to Space Missions. Marine Drugs. 2022; 20(5):299. https://doi.org/10.3390/md20050299
Chicago/Turabian StyleFais, Giacomo, Alessia Manca, Federico Bolognesi, Massimiliano Borselli, Alessandro Concas, Marco Busutti, Giovanni Broggi, Pierdanilo Sanna, Yandy Marx Castillo-Aleman, René Antonio Rivero-Jiménez, and et al. 2022. "Wide Range Applications of Spirulina: From Earth to Space Missions" Marine Drugs 20, no. 5: 299. https://doi.org/10.3390/md20050299
APA StyleFais, G., Manca, A., Bolognesi, F., Borselli, M., Concas, A., Busutti, M., Broggi, G., Sanna, P., Castillo-Aleman, Y. M., Rivero-Jiménez, R. A., Bencomo-Hernandez, A. A., Ventura-Carmenate, Y., Altea, M., Pantaleo, A., Gabrielli, G., Biglioli, F., Cao, G., & Giannaccare, G. (2022). Wide Range Applications of Spirulina: From Earth to Space Missions. Marine Drugs, 20(5), 299. https://doi.org/10.3390/md20050299