
Humic Substances as Microalgal Biostimulants—Implications for Microalgal Biotechnology

 and
and 
Abstract
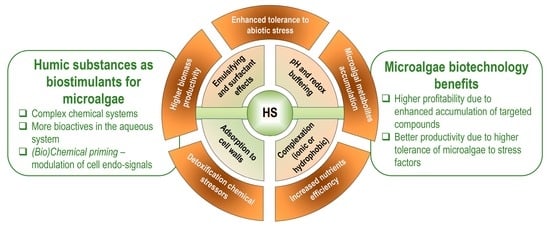
:
1. Introduction
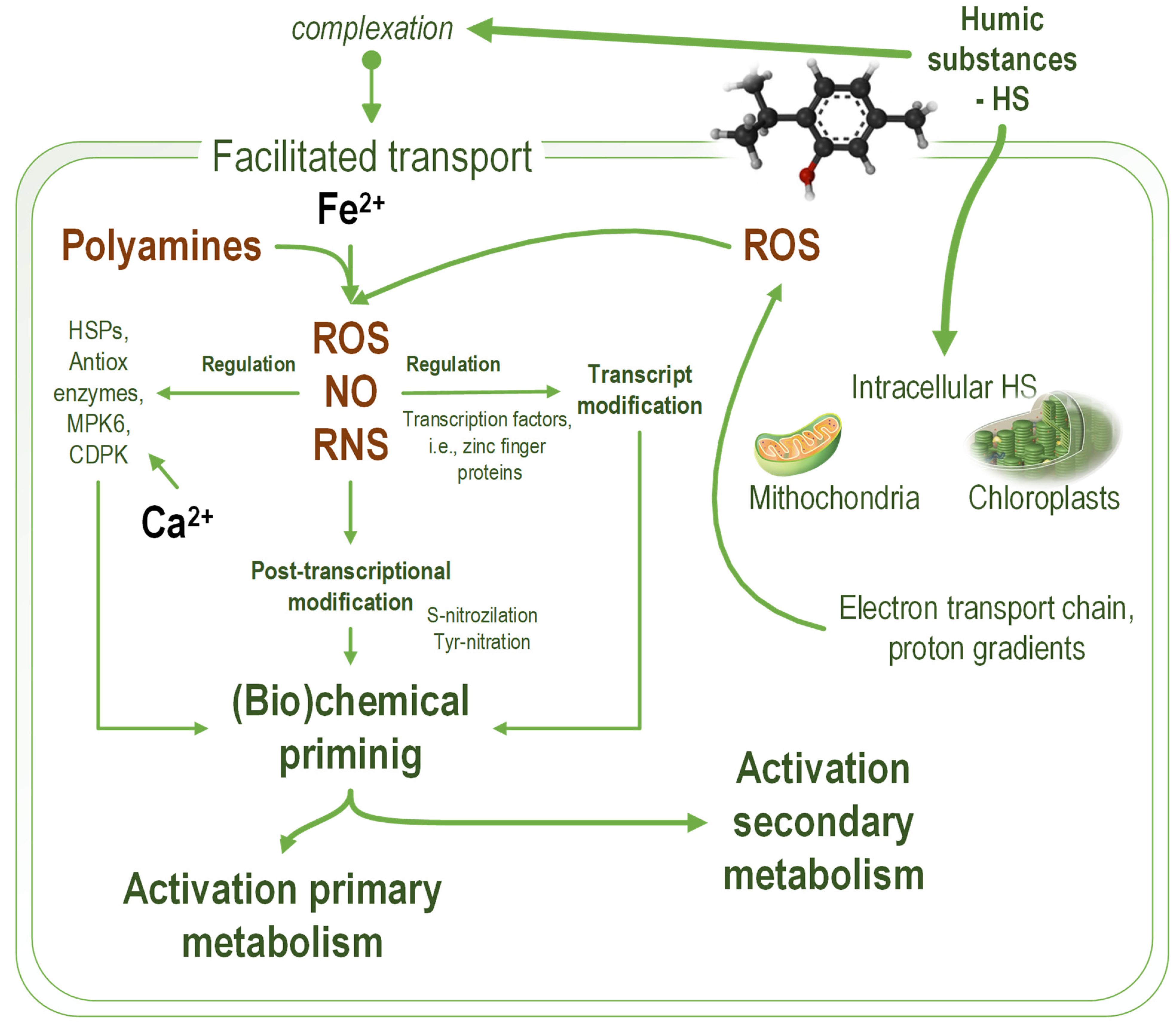
2. Hormetic Effects of Humic Substances on Microalgae
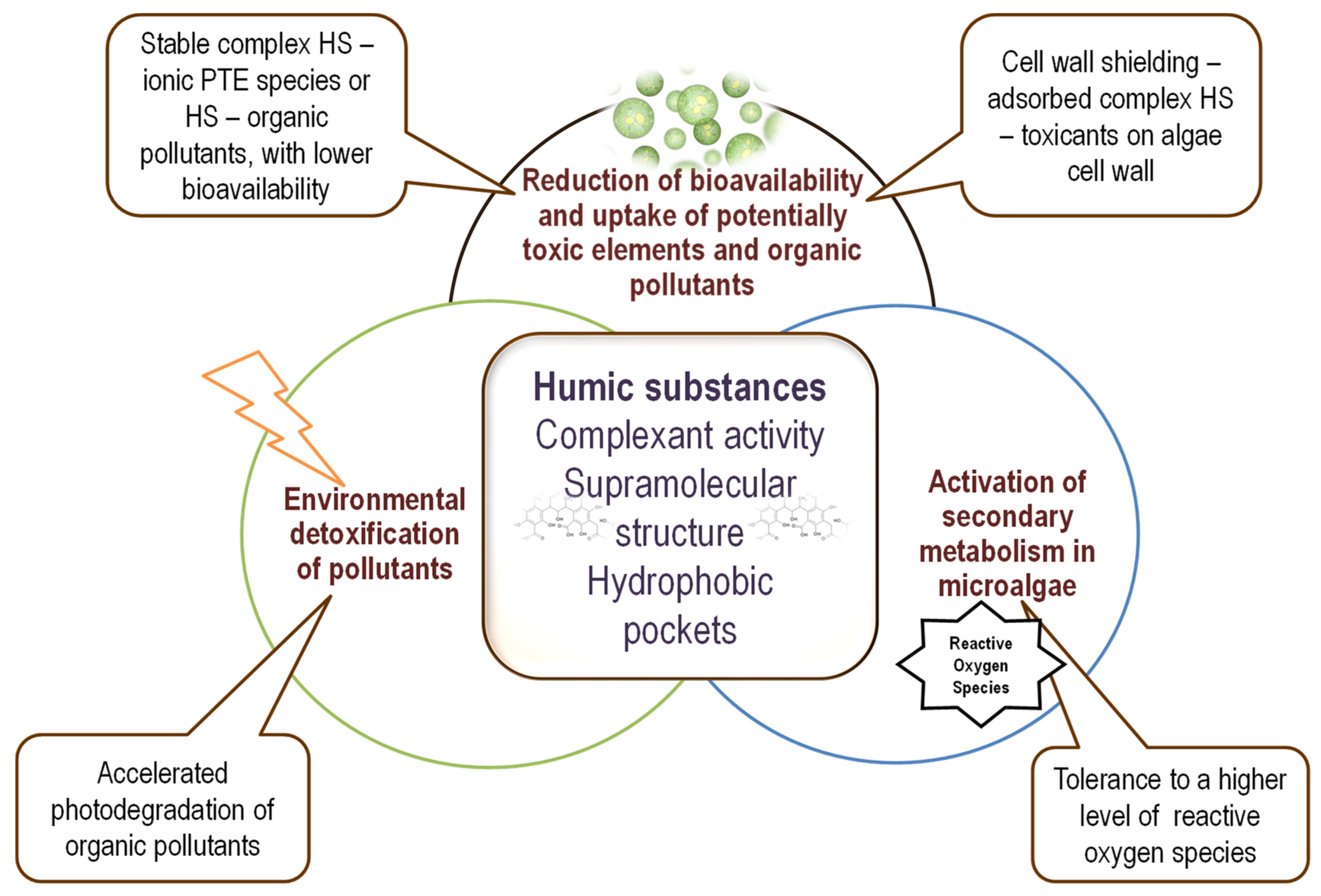
3. Protective Effects of Humic Substances on Microalgae
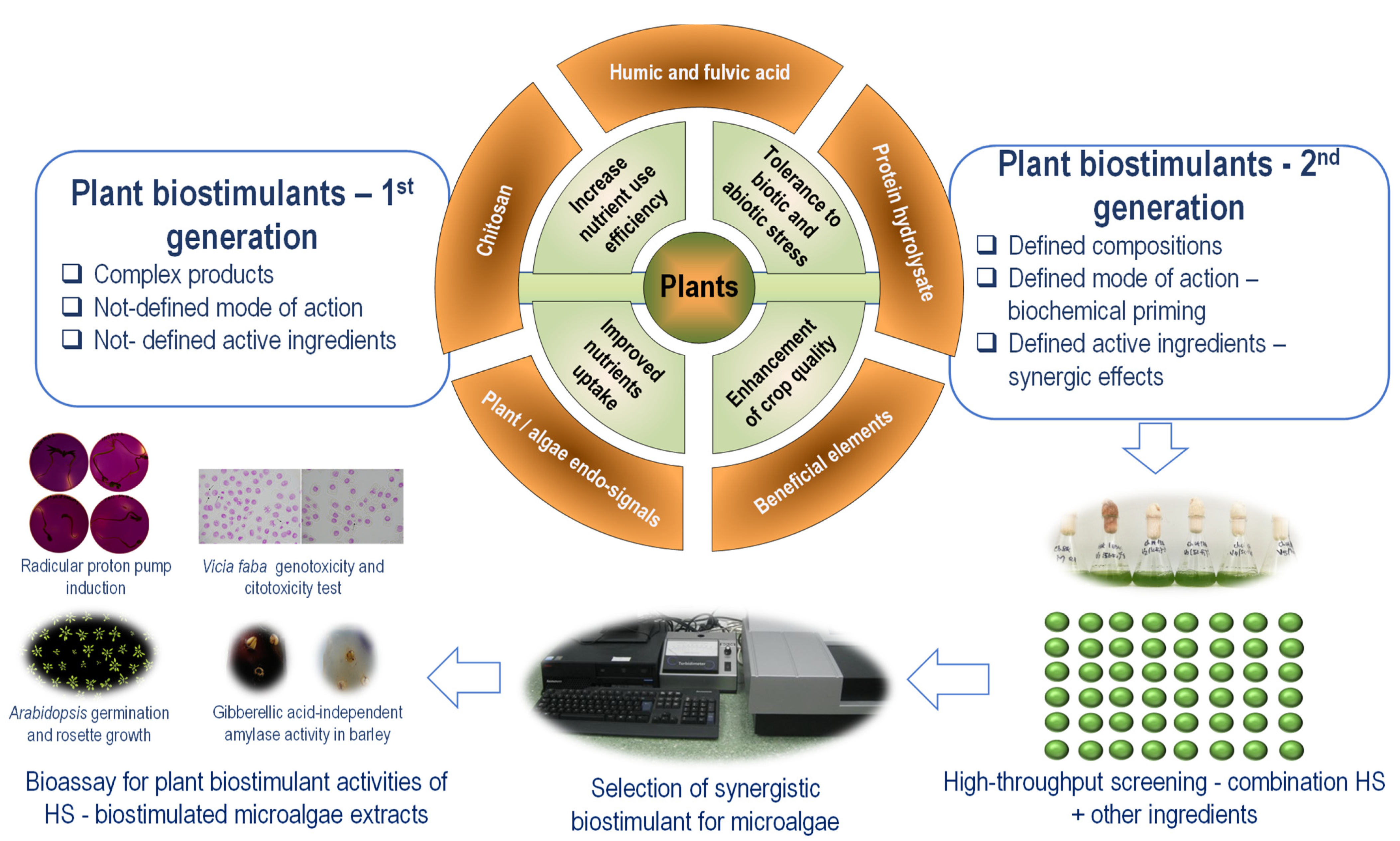
4. Humic Substances as Microalgae Biostimulants
5. Humic Substance Interactions with Microalgae Harvesting by Flocculation
6. Enhanced Biotechnological Production of Microalgae-Based High-Value Products by Humic Substances
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lehmann, J.; Kleber, M. The contentious nature of soil organic matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Regelink, I.C.; Stoof, C.R.; Rousseva, S.; Weng, L.; Lair, G.J.; Kram, P.; Nikolaidis, N.P.; Kercheva, M.; Banwart, S.; Comans, R.N.J. Linkages between aggregate formation, porosity and soil chemical properties. Geoderma 2015, 247–248, 24–37. [Google Scholar] [CrossRef]
- Yang, F.; Tang, C.; Antonietti, M. Natural and artificial humic substances to manage minerals, ions, water, and soil microorganisms. Chem. Soc. Rev. 2021, 50, 6221–6239. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.H. Solvent systems for the isolation of organic components from soils. Soil Sci. Soc. Am. J. 2006, 70, 986–994. [Google Scholar] [CrossRef] [Green Version]
- Olk, D.C.; Bloom, P.R.; Perdue, E.M.; McKnight, D.M.; Chen, Y.; Farenhorst, A.; Senesi, N.; Chin, Y.P.; Schmitt-Kopplin, P.; Hertkorn, N.; et al. Environmental and agricultural relevance of humic fractions extracted by alkali from soils and natural waters. J. Environ. Qual. 2019, 48, 217–232. [Google Scholar] [CrossRef]
- Piccolo, A. The supramolecular structure of humic substances: A novel understanding of humus chemistry and implications in soil science. Adv. Agron. 2002, 75, 57–134. [Google Scholar] [CrossRef]
- Guo, X.X.; Liu, H.T.; Wu, S.B. Humic substances developed during organic waste composting: Formation mechanisms, structural properties, and agronomic functions. Sci. Total Environ. 2019, 662, 501–510. [Google Scholar] [CrossRef]
- Nebbioso, A.; Piccolo, A. Molecular characterization of dissolved organic matter (DOM): A critical review. Anal. Bioanal. Chem. 2013, 405, 109–124. [Google Scholar] [CrossRef]
- Adani, F.; Spagnol, M.; Nierop, K.G. Biochemical origin and refractory properties of humic acid extracted from maize plants: The contribution of lignin. Biogeochemistry 2007, 82, 55–65. [Google Scholar] [CrossRef]
- Shevchenko, S.M.; Bailey, G.W. Life after death: Lignin-humic relationships reexamined. Crit. Rev. Environ. Sci. Technol. 1996, 26, 95–153. [Google Scholar] [CrossRef]
- Simpson, A.J.; Kingery, W.L.; Hayes, M.H.; Spraul, M.; Humpfer, E.; Dvortsak, P.; Kerssebaum, R.; Godejohann, M.; Hofmann, M. Molecular structures and associations of humic substances in the terrestrial environment. Naturwissenschaften 2002, 89, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Jahnel, J.B.; Frimmel, F.H. Enzymatic release of amino acids from different humic substances. Acta Hydrochim. Hydrobiol. 1995, 23, 31–35. [Google Scholar] [CrossRef]
- Ding, G.; Mao, J.; Xing, B. Characteristics of amino acids in soil humic substances. Commun. Soil Sci. Plant Anal. 2006, 32, 1991–2005. [Google Scholar] [CrossRef]
- Zou, J.; Huang, J.; Zhang, H.; Yue, D. Evolution of humic substances in polymerization of polyphenol and amino acid based on non-destructive characterization. Front. Environ. Sci. Eng. 2020, 15, 5. [Google Scholar] [CrossRef]
- Young, C.C.; Chen, L.F. Polyamines in humic acid and their effect on radical growth of lettuce seedlings. Plant Soil 1997, 195, 143–149. [Google Scholar] [CrossRef]
- Salati, S.; Papa, G.; Adani, F. Perspective on the use of humic acids from biomass as natural surfactants for industrial applications. Biotechnol. Adv. 2011, 29, 913–922. [Google Scholar] [CrossRef]
- Li, A.; Hu, J.; Li, W.; Zhang, W.; Wang, X. Polarity based fractionation of fulvic acids. Chemosphere 2009, 77, 1419–1426. [Google Scholar] [CrossRef]
- de Melo, B.A.; Motta, F.L.; Santana, M.H. Humic acids: Structural properties and multiple functionalities for novel technological developments. Mater. Sci. Eng. C Mater. Biol. Appl. 2016, 62, 967–974. [Google Scholar] [CrossRef]
- Chaaban, A.A.; Lartiges, B.; El Hayek, E.; Kazpard, V.; Plisson-Chastang, C.; Vicendo, P.; Caillet, C. Complexes of humic acid with cationic surfactants support the supramolecular view of extracted humic matter. Environ. Chem. 2021, 18, 156–167. [Google Scholar] [CrossRef]
- Gerke, J. Concepts and misconceptions of humic substances as the stable part of soil organic matter: A review. Agronomy 2018, 8, 76. [Google Scholar] [CrossRef] [Green Version]
- Piccolo, A.; Cozzolino, A.; Conte, P.; Spaccini, R. Polymerization of humic substances by an enzyme-catalyzed oxidative coupling. Naturwissenschaften 2000, 87, 391–394. [Google Scholar] [CrossRef] [PubMed]
- Spielvogel, S.; Prietzel, J.; Kgel-Knabner, I. Soil organic matter stabilization in acidic forest soils is preferential and soil type-specific. Eur. J. Soil Sci. 2008, 59, 674–692. [Google Scholar] [CrossRef]
- Yang, F.; Antonietti, M. The sleeping giant: A polymer View on humic matter in synthesis and applications. Prog. Polym. Sci. 2020, 100, 101182. [Google Scholar] [CrossRef]
- He, X.S.; Yang, C.; You, S.H.; Zhang, H.; Xi, B.D.; Yu, M.D.; Liu, S.J. Redox properties of compost-derived organic matter and their association with polarity and molecular weight. Sci. Total Environ. 2019, 665, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Haider, K.; Martin, J.P. Synthesis and transformation of phenolic compounds by Epicoccum nigrum in relation to humic acid formation. Soil Sci. Soc. Am. J. 1967, 31, 766–772. [Google Scholar] [CrossRef]
- Boguta, P.; Skic, K.; Sokolowska, Z.; Frac, M.; Sas-Paszt, L. Chemical transformation of humic acid molecules under the influence of mineral, fungal and bacterial fertilization in the context of the agricultural use of degraded soils. Molecules 2021, 26, 4921. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Dou, S.; Zhang, X.; Cui, Y.J.; Wang, T. FTIR spectroscopic analysis of humic-like substances extracted from the microbial residues. Guang Pu Xue Yu Guang Pu Fen Xi 2015, 35, 3397–3401. [Google Scholar] [CrossRef]
- Sokol, N.W.; Sanderman, J.; Bradford, M.A. Pathways of mineral-associated soil organic matter formation: Integrating the role of plant carbon source, chemistry, and point of entry. Glob. Chang. Biol. 2019, 25, 12–24. [Google Scholar] [CrossRef] [Green Version]
- Pertusatti, J.; Prado, A.G. Buffer capacity of humic acid: Thermodynamic approach. J. Colloid Interface Sci. 2007, 314, 484–489. [Google Scholar] [CrossRef]
- Rakshit, S.; Sarkar, D. Assessing redox properties of standard humic substances. Int. J. Environ. Sci. Technol. 2017, 14, 1497–1504. [Google Scholar] [CrossRef]
- Capasso, S.; Chianese, S.; Musmarra, D.; Iovino, P. Macromolecular structure of a commercial humic acid sample. Environments 2020, 7, 32. [Google Scholar] [CrossRef] [Green Version]
- Gheorghe, D.-I.; Constantinescu-Aruxandei, D.; Lupu, C.; Oancea, F. Emulsifying effect of fulvic acids from shilajit. Chem. Proc. 2022, 7, 23. [Google Scholar]
- Kucerik, J.; Smejkalova, D.; Cechlovska, H.; Pekar, M. New insights into aggregation and conformational behaviour of humic substances: Application of high resolution ultrasonic spectroscopy. Org. Geochem. 2007, 38, 2098–2110. [Google Scholar] [CrossRef]
- Piccolo, A. The nature of soil organic matter and innovative soil managements to fight global changes and maintain agricultural productivity. In Carbon Sequestration in Agricultural Soils; Springer: New York, NY, USA, 2012; pp. 1–19. [Google Scholar]
- Verrillo, M.; Salzano, M.; Savy, D.; Di Meo, V.; Valentini, M.; Cozzolino, V.; Piccolo, A. Antibacterial and antioxidant properties of humic substances from composted agricultural biomasses. Chem. Biol. Technol. Agric. 2022, 9, 28. [Google Scholar] [CrossRef]
- Rose, M.T.; Patti, A.F.; Little, K.R.; Brown, A.L.; Jackson, W.R.; Cavagnaro, T.R. A meta-analysis and review of plant-growth response to humic substances: Practical implications for agriculture. Adv. Agron. 2014, 124, 37–89. [Google Scholar]
- Canellas, L.P.; Olivares, F.L.; Okorokova-Facanha, A.L.; Facanha, A.R. Humic acids isolated from earthworm compost enhance root elongation, lateral root emergence, and plasma membrane H+-ATPase activity in maize roots. Plant Physiol. 2002, 130, 1951–1957. [Google Scholar] [CrossRef] [Green Version]
- Nardi, S.; Pizzeghello, D.; Muscolo, A.; Vianello, A. Physiological effects of humic substances on higher plants. Soil Biol. Biochem. 2002, 34, 1527–1536. [Google Scholar] [CrossRef]
- Quaggiotti, S.; Ruperti, B.; Pizzeghello, D.; Francioso, O.; Tugnoli, V.; Nardi, S. Effect of low molecular size humic substances on nitrate uptake and expression of genes involved in nitrate transport in maize (Zea mays L.). J. Exp. Bot. 2004, 55, 803–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavares, O.; Santos, L.; Ferreira, L.; Sperandio, M.; da Rocha, J.; García, A.; Dobbss, L.; Berbara, R.; de Souza, S.; Fernandes, M. Humic acid differentially improves nitrate kinetics under low-and high-affinity systems and alters the expression of plasma membrane H+-ATPases and nitrate transporters in rice. Ann. Appl. Biol. 2017, 170, 89–103. [Google Scholar] [CrossRef] [Green Version]
- Jindo, K.; Soares, T.S.; Peres, L.E.P.; Azevedo, I.G.; Aguiar, N.O.; Mazzei, P.; Spaccini, R.; Piccolo, A.; Olivares, F.L.; Canellas, L.P. Phosphorus speciation and high-affinity transporters are influenced by humic substances. J. Plant Nutr. Soil Sci. 2016, 179, 206–214. [Google Scholar] [CrossRef]
- Jannin, L.; Arkoun, M.; Ourry, A.; Laîné, P.; Goux, D.; Garnica, M.; Fuentes, M.; Francisco, S.S.; Baigorri, R.; Cruz, F. Microarray analysis of humic acid effects on Brassica napus growth: Involvement of N, C and S metabolisms. Plant Soil 2012, 359, 297–319. [Google Scholar] [CrossRef]
- Zanin, L.; Tomasi, N.; Cesco, S.; Varanini, Z.; Pinton, R. Humic substances contribute to plant iron nutrition acting as chelators and biostimulants. Front. Plant Sci. 2019, 10, 675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanin, L.; Tomasi, N.; Zamboni, A.; Sega, D.; Varanini, Z.; Pinton, R. Water-extractable humic substances speed up transcriptional response of maize roots to nitrate. Environ. Exp. Bot. 2018, 147, 167–178. [Google Scholar] [CrossRef]
- Trevisan, S.; Botton, A.; Vaccaro, S.; Vezzaro, A.; Quaggiotti, S.; Nardi, S. Humic substances affect Arabidopsis physiology by altering the expression of genes involved in primary metabolism, growth and development. Environ. Exp. Bot. 2011, 74, 45–55. [Google Scholar] [CrossRef]
- Aguiar, N.O.; Olivares, F.L.; Novotny, E.H.; Canellas, L.P. Changes in metabolic profiling of sugarcane leaves induced by endophytic diazotrophic bacteria and humic acids. Peerj 2018, 6, e5445. [Google Scholar] [CrossRef] [PubMed]
- Othibeng, K.; Nephali, L.; Ramabulana, A.T.; Steenkamp, P.; Petras, D.; Kang, K.B.; Opperman, H.; Huyser, J.; Tugizimana, F. A metabolic choreography of maize plants treated with a humic substance-based biostimulant under normal and starved conditions. Metabolites 2021, 11, 403. [Google Scholar] [CrossRef] [PubMed]
- Zandonadi, D.B.; Santos, M.P.; Dobbss, L.B.; Olivares, F.L.; Canellas, L.P.; Binzel, M.L.; Okorokova-Facanha, A.L.; Facanha, A.R. Nitric oxide mediates humic acids-induced root development and plasma membrane H+-ATPase activation. Planta 2010, 231, 1025–1036. [Google Scholar] [CrossRef]
- Ramos, A.C.; Dobbss, L.B.; Santos, L.A.; Fernandes, M.S.; Olivares, F.L.; Aguiar, N.O.; Canellas, L.P. Humic matter elicits proton and calcium fluxes and signaling dependent on Ca2+-dependent protein kinase (CDPK) at early stages of lateral plant root development. Chem. Biol. Technol. Agric. 2015, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Canellas, L.P.; Canellas, N.O.A.; da Irineu, L.E.S.; Olivares, F.L.; Piccolo, A. Plant chemical priming by humic acids. Chem. Biol. Technol. Agric. 2020, 7, 12. [Google Scholar] [CrossRef]
- Jindo, K.; Olivares, F.L.; Malcher, D.; Sanchez-Monedero, M.A.; Kempenaar, C.; Canellas, L.P. From lab to field: Role of humic substances under open-field and greenhouse conditions as biostimulant and biocontrol agent. Front. Plant Sci. 2020, 11, 426. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front. Plant Sci. 2016, 7, 2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.; Liu, Y.; Leverage, W.T.; Yin, J.J.; White, I.M.; Bentley, W.E.; Payne, G.F. Context-dependent redox properties of natural phenolic materials. Biomacromolecules 2014, 15, 1653–1662. [Google Scholar] [CrossRef] [PubMed]
- Stoica, R.; Oancea, F.; Minca, I.; Doncea, S.M.; Ganea, R.; Capra, L.; Senin, R.; Mateescu, M.; Ciobanu, I.; Ivan, G. Spectroscopic, textural and thermal characterization methods of biostimulants based on sodium humate. Rev. Chim. 2018, 69, 3477–3482. [Google Scholar] [CrossRef]
- Puglisi, I.; Barone, V.; Sidella, S.; Coppa, M.; Broccanello, C.; Gennari, M.; Baglieri, A. Biostimulant activity of humic-like substances from agro-industrial waste on Chlorella vulgaris and Scenedesmus quadricauda. Eur. J. Phycol. 2018, 53, 433–442. [Google Scholar] [CrossRef]
- Faraon, V.A.; Popa, D.G.; Tudor-Popa, I.; Mihăilă, E.G.; Oancea, F. Effect of humic acids from biomass biostimulant on microalgae growth. Chem. Proc. 2022, 7, 57. [Google Scholar]
- Saito, S. Separation approaches towards understanding supramolecular aggregate formation of humic acid. Anal. Sci. 2022, 38, 233–234. [Google Scholar] [CrossRef]
- Prakash, A.a.; Rashid, M. Influence of humic substances on the growth of marine phytoplankton: Dinoflagellates. Limnol. Oceanogr. 1968, 13, 598–606. [Google Scholar] [CrossRef]
- Zheng, X.; Xu, Z.; Zhao, D.; Luo, Y.; Lai, C.; Huang, B.; Pan, X. Double-dose responses of Scenedesmus capricornus microalgae exposed to humic acid. Sci. Total Environ. 2022, 806, 150547. [Google Scholar] [CrossRef]
- Toropkina, M.; Ryumin, A.; Kechaikina, I.; Chukov, S. Effect of humic acids on the metabolism of Chlorella vulgaris in a model experiment. Eurasian Soil Sci. 2017, 50, 1294–1300. [Google Scholar] [CrossRef]
- Bährs, H.; Steinberg, C.E. Impact of two different humic substances on selected coccal green algae and cyanobacteria—Changes in growth and photosynthetic performance. Environ. Sci. Pollut. Res. 2012, 19, 335–346. [Google Scholar] [CrossRef]
- Prokhotskaya, V.Y.; Steinberg, C.E. Differential sensitivity of a coccal green algal and a Cyanobacterial species to dissolved natural organic matter (NOM). Environ. Sci. Pollut. Res. 2007, 14, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Pouneva, I. Effect of humic substances on the growth of microalgal cultures. Russ. J. Plant Physiol. 2005, 52, 410–413. [Google Scholar] [CrossRef]
- Steinberg, C.E.; Kamara, S.; Prokhotskaya, V.Y.; Manusadžianas, L.; Karasyova, T.A.; Timofeyev, M.A.; Jie, Z.; Paul, A.; Meinelt, T.; Farjalla, V.F. Dissolved humic substances–ecological driving forces from the individual to the ecosystem level? Freshw. Biol. 2006, 51, 1189–1210. [Google Scholar] [CrossRef]
- González, P.M.; Malanga, G.; Puntarulo, S. Cellular oxidant/antioxidant network: Update on the environmental effects over marine organisms. Open Mar. Biol. J. 2015, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Vigneault, B.; Percot, A.; Lafleur, M.; Campbell, P.G. Permeability changes in model and phytoplankton membranes in the presence of aquatic humic substances. Environ. Sci. Technol. 2000, 34, 3907–3913. [Google Scholar] [CrossRef]
- Hasegawa, H.; Tate, Y.; Ogino, M.; Maki, T.; Begum, Z.A.; Ichijo, T.; Rahman, I.M. Laboratory culture experiments to study the effect of lignite humic acid fractions on iron solubility and iron uptake rates in phytoplankton. J. Appl. Phycol. 2017, 29, 903–915. [Google Scholar] [CrossRef] [Green Version]
- Orlowska, E.; Roller, A.; Pignitter, M.; Jirsa, F.; Krachler, R.; Kandioller, W.; Keppler, B.K. Synthetic iron complexes as models for natural iron-humic compounds: Synthesis, characterization and algal growth experiments. Sci. Total Environ. 2017, 577, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Benderliev, K.M.; Ivanova, N.I.; Pilarski, P.S. Singlet oxygen and other reactive oxygen species are involved in regulation of release of iron-binding chelators from Scenedesmus cells. Biol. Plant. 2003, 47, 523–526. [Google Scholar] [CrossRef]
- Liu, W.; Au, D.W.T.; Anderson, D.M.; Lam, P.K.S.; Wu, R.S.S. Effects of nutrients, salinity, pH and light:dark cycle on the production of reactive oxygen species in the alga Chattonella marina. J. Exp. Mar. Biol. Ecol. 2007, 346, 76–86. [Google Scholar] [CrossRef]
- Lee, J.; Park, J.H.; Shin, Y.S.; Lee, B.C.; Chang, N.I.; Cho, J.; Kim, S.D. Effect of dissolved organic matter on the growth of algae, Pseudokirchneriella subcapitata, in Korean lakes: The importance of complexation reactions. Ecotoxicol. Environ. Saf. 2009, 72, 335–343. [Google Scholar] [CrossRef]
- Perez-Perez, M.E.; Lemaire, S.D.; Crespo, J.L. Reactive oxygen species and autophagy in plants and algae. Plant Physiol. 2012, 160, 156–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamaeva, A.; Taliansky, M.; Filippova, A.; Love, A.J.; Golub, N.; Fesenko, I. The role of chloroplast protein remodeling in stress responses and shaping of the plant peptidome. New Phytol. 2020, 227, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.C.; Burch-Smith, T.M. Chloroplasts as mediators of plant biotic interactions over short and long distances. Curr. Opin. Plant Biol. 2019, 50, 148–155. [Google Scholar] [CrossRef]
- Unal, D.; García-Caparrós, P.; Kumar, V.; Dietz, K.-J. Chloroplast-associated molecular patterns as concept for fine-tuned operational retrograde signalling. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190443. [Google Scholar] [CrossRef]
- Wang, Y.; Selinski, J.; Mao, C.; Zhu, Y.; Berkowitz, O.; Whelan, J. Linking mitochondrial and chloroplast retrograde signalling in plants. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190410. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wang, Y.; Chan, K.X.; Marchant, D.B.; Franks, P.J.; Randall, D.; Tee, E.E.; Chen, G.; Ramesh, S.; Phua, S.Y.; et al. Evolution of chloroplast retrograde signaling facilitates green plant adaptation to land. Proc. Natl. Acad. Sci. USA 2019, 116, 5015–5020. [Google Scholar] [CrossRef] [Green Version]
- Rea, G.; Antonacci, A.; Lambreva, M.D.; Mattoo, A.K. Features of cues and processes during chloroplast-mediated retrograde signaling in the alga Chlamydomonas. Plant Sci. 2018, 272, 193–206. [Google Scholar] [CrossRef]
- Gilbert, M.; Bährs, H.; Steinberg, C.E.; Wilhelm, C. The artificial humic substance HS1500 does not inhibit photosynthesis of the green alga Desmodesmus armatus in vivo but interacts with the photosynthetic apparatus of isolated spinach thylakoids in vitro. Photosynth. Res. 2018, 137, 403–420. [Google Scholar] [CrossRef]
- Allen, J. Photosynthesis of ATP-electrons, proton pumps, rotors, and poise. Cell 2002, 110, 273–276. [Google Scholar] [CrossRef] [Green Version]
- Heinze, T.; Bährs, H.; Gilbert, M.; Steinberg, C.E.; Wilhelm, C. Selected coccal green algae are not affected by the humic substance Huminfeed® in term of growth or photosynthetic performance. Hydrobiologia 2012, 684, 215–224. [Google Scholar] [CrossRef]
- Steinberg, C.E.; Timofeyev, M.A.; Menzel, R. Dissolved Humic Substances—Interactions with Organisms; Academic Press Oxford: Oxford, UK, 2009. [Google Scholar]
- Peng, J.; Zhang, Y.; Li, J.; Wu, X.; Wang, M.; Gong, Z.; Gao, S. Interference of dissolved organic matter and its constituents on the accurate determination of hydrogen peroxide in water. Sci. Rep. 2021, 11, 22613. [Google Scholar] [CrossRef] [PubMed]
- Amblard, C.; Carrias, J.-F.o.; Bourdier, G.; Maurin, N. The microbial loop in a humic lake: Seasonal and vertical variations in the structure of the different communities. Hydrobiologia 1995, 300–301, 71–84. [Google Scholar] [CrossRef]
- Barone, M.E.; Parkes, R.; Herbert, H.; McDonnell, A.; Conlon, T.; Aranyos, A.; Fierli, D.; Fleming, G.T.A.; Touzet, N. Comparative response of marine microalgae to H2O2-induced oxidative stress. Appl. Biochem. Biotechnol. 2021, 193, 4052–4067. [Google Scholar] [CrossRef] [PubMed]
- Koukal, B.; Gueguen, C.; Pardos, M.; Dominik, J. Influence of humic substances on the toxic effects of cadmium and zinc to the green alga Pseudokirchneriella subcapitata. Chemosphere 2003, 53, 953–961. [Google Scholar] [CrossRef]
- Spierings, J.; Worms, I.A.; Mieville, P.; Slaveykova, V.I. Effect of humic substance photoalteration on lead bioavailability to freshwater microalgae. Environ. Sci. Technol. 2011, 45, 3452–3458. [Google Scholar] [CrossRef]
- Shi, Z.; Xu, H.; Wang, Z.; Du, H.; Fu, X. Effects of co-exposure to copper and humic acids on microalga Chlorella vulgaris: Growth inhibition, oxidative stress, and extracellular secretion. Environ. Pollut. Bioavailable 2021, 33, 415–424. [Google Scholar] [CrossRef]
- Hanachi, P.; Khoshnamvand, M.; Walker, T.R.; Hamidian, A.H. Nano-sized polystyrene plastics toxicity to microalgae Chlorella vulgaris: Toxicity mitigation using humic acid. Aquat. Toxicol. 2022, 245, 106123. [Google Scholar] [CrossRef]
- Tong, M.; Li, X.; Luo, Q.; Yang, C.; Lou, W.; Liu, H.; Du, C.; Nie, L.; Zhong, Y. Effects of humic acids on biotoxicity of tetracycline to microalgae Coelastrella sp. Algal Res. 2020, 50, 101962. [Google Scholar] [CrossRef]
- Zhao, J.; Li, Y.; Cao, X.; Guo, C.; Xu, L.; Wang, Z.; Feng, J.; Yi, H.; Xing, B. Humic acid mitigated toxicity of graphene-family materials to algae through reducing oxidative stress and heteroaggregation. Environ. Sci. Nano 2019, 6, 1909–1920. [Google Scholar] [CrossRef]
- Slaveykova, V.I.; Wilkinson, K.J. Predicting the bioavailability of metals and metal complexes: Critical review of the biotic ligand model. Environ. Chem. 2005, 2, 9–24. [Google Scholar] [CrossRef]
- Lamelas, C.; Pinheiro, J.P.; Slaveykova, V.I. Effect of humic acid on Cd(II), Cu(II), and Pb(II) uptake by freshwater algae: Kinetic and cell wall speciation considerations. Environ. Sci. Technol. 2009, 43, 730–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Porcher, C.; Campbell, P.G.; Fortin, C. Influence of humic acid on algal uptake and toxicity of ionic silver. Environ. Sci. Technol. 2013, 47, 8835–8842. [Google Scholar] [CrossRef] [PubMed]
- Nanayama, Y.; Sazawa, K.; Yustiawati, Y.; Syawal, M.S.; Fukushima, M.; Kuramitz, H. Effect of humic acids on the toxicity of pollutants to Chlamydomonas reinhardtii: Investigation by a microscale algal growth inhibition test. Environ. Sci. Pollut. Res. Int. 2021, 28, 211–219. [Google Scholar] [CrossRef]
- Pandey, A.K.; Pandey, S.D.; Misra, V. Stability constants of metal-humic acid complexes and its role in environmental detoxification. Ecotoxicol. Environ. Saf. 2000, 47, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Manuel Ostera, J.; Puntarulo, S.; Malanga, G. Oxidative effects of glyphosate on the lipophobic intracellular environment in the microalgae. Biocell 2022, 46, 795–802. [Google Scholar] [CrossRef]
- Liu, J.; Fan, J.; He, T.; Xu, X.; Ai, Y.; Tang, H.; Gu, H.; Lu, T.; Liu, Y.; Liu, G. The mechanism of aquatic photodegradation of organophosphorus sensitized by humic acid-Fe(3+) complexes. J. Hazard Mater. 2020, 384, 121466. [Google Scholar] [CrossRef]
- Zhang, Y.; Meng, T.; Guo, X.; Yang, R.; Si, X.; Zhou, J. Humic acid alleviates the ecotoxicity of graphene-family materials on the freshwater microalgae Scenedesmus obliquus. Chemosphere 2018, 197, 749–758. [Google Scholar] [CrossRef]
- Yin, J.; Dong, Z.; Liu, Y.; Wang, H.; Li, A.; Zhuo, Z.; Feng, W.; Fan, W. Toxicity of reduced graphene oxide modified by metals in microalgae: Effect of the surface properties of algal cells and nanomaterials. Carbon 2020, 169, 182–192. [Google Scholar] [CrossRef]
- Ouyang, S.; Zhou, Q.; Zeng, H.; Wang, Y.; Hu, X. Natural Nanocolloids Mediate the Phytotoxicity of Graphene Oxide. Environ. Sci. Technol. 2020, 54, 4865–4875. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- du Jardin, P.; Xu, L.; Geelen, D. Agricultural Functions and Action Mechanisms of Plant Biostimulants (PBs): An Introduction. In The Chemical Biology of Plant Biostimulants; Geelen, D., Xu, L., Eds.; Wiley: Hoboken, NJ, USA, 2020; pp. 1–30. [Google Scholar]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Muscolo, A.; Nardi, S. Alfalfa plant-derived biostimulant stimulate short-term growth of salt stressed Zea mays L. plants. Plant Soil 2013, 364, 145–158. [Google Scholar] [CrossRef]
- Godlewska, K.; Pacyga, P.; Michalak, I.; Biesiada, A.; Szumny, A.; Pachura, N.; Piszcz, U. Effect of botanical extracts on the growth and nutritional quality of field-grown white head cabbage (Brassica oleracea var. capitata). Molecules 2021, 26, 1992. [Google Scholar] [CrossRef] [PubMed]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and fulvic acids as biostimulants in horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and crop responses: A review. Biol. Agric. Hortic. 2015, 31, 1–17. [Google Scholar] [CrossRef]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.; Cardarelli, M. Biostimulant action of a plant-derived protein hydrolysate produced through enzymatic hydrolysis. Front. Plant Sci. 2014, 5, 448. [Google Scholar] [CrossRef] [Green Version]
- Pichyangkura, R.; Chadchawan, S. Biostimulant activity of chitosan in horticulture. Sci. Hortic. 2015, 196, 49–65. [Google Scholar] [CrossRef]
- Malerba, M.; Cerana, R. Recent advances of chitosan applications in plants. Polymers 2018, 10, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rachidi, F.; Benhima, R.; Kasmi, Y.; Sbabou, L.; El Arroussi, H. Evaluation of microalgae polysaccharides as biostimulants of tomato plant defense using metabolomics and biochemical approaches. Sci. Rep. 2021, 11, 930. [Google Scholar]
- Zhang, C.; Wang, W.; Zhao, X.; Wang, H.; Yin, H. Preparation of alginate oligosaccharides and their biological activities in plants: A review. Carbohydr. Res. 2020, 494, 108056. [Google Scholar] [CrossRef] [PubMed]
- Salachna, P.; Pietrak, A. Evaluation of carrageenan, xanthan gum and depolymerized chitosan based coatings for pineapple lily plant production. Horticulturae 2021, 7, 19. [Google Scholar] [CrossRef]
- Savvas, D.; Ntatsi, G. Biostimulant activity of silicon in horticulture. Sci. Hortic. 2015, 196, 66–81. [Google Scholar] [CrossRef]
- Laane, H.-M. The effects of foliar sprays with different silicon compounds. Plants 2018, 7, 45. [Google Scholar] [CrossRef] [Green Version]
- Bărbieru, O.-G.; Dimitriu, L.; Călin, M.; Răut, I.; Constantinescu-Aruxandei, D.; Oancea, F. Plant biostimulants based on selenium nanoparticles biosynthesized by Trichoderma strains. Proceedings 2019, 29, 95. [Google Scholar]
- Dima, S.-O.; Neamțu, C.; Desliu-Avram, M.; Ghiurea, M.; Capra, L.; Radu, E.; Stoica, R.; Faraon, V.-A.; Zamfiropol-Cristea, V.; Constantinescu-Aruxandei, D.; et al. Plant biostimulant effects of baker’s yeast vinasse and selenium on tomatoes through foliar fertilization. Agronomy 2020, 10, 133. [Google Scholar] [CrossRef] [Green Version]
- García-García, A.L.; García-Machado, F.J.; Borges, A.A.; Morales-Sierra, S.; Boto, A.; Jiménez-Arias, D. Pure organic active compounds against abiotic stress: A biostimulant overview. Front. Plant Sci. 2020, 11, 5819. [Google Scholar] [CrossRef]
- Kerchev, P.; van der Meer, T.; Sujeeth, N.; Verlee, A.; Stevens, C.V.; Van Breusegem, F.; Gechev, T. Molecular priming as an approach to induce tolerance against abiotic and oxidative stresses in crop plants. Biotechnol. Adv. 2020, 40, 107503. [Google Scholar] [CrossRef]
- Velea, S.; Oancea, F.; Fischer, F. Heterotrophic and mixotrophic microalgae cultivation. In Microalgae-Based Biofuels and Bioproducts, Cristina; Gonzalez-Fernandez, R.M., Ed.; Woodhead Publishing—Elsevier: Duxford, UK, 2017; pp. 45–65. [Google Scholar]
- Rouphael, Y.; Colla, G. Toward a sustainable agriculture through plant biostimulants: From experimental data to practical applications. Agronomy 2020, 10, 1461. [Google Scholar] [CrossRef]
- Zhao, Y.; Xing, H.; Li, X.; Geng, S.; Ning, D.; Ma, T.; Yu, X. Physiological and metabolomics analyses reveal the roles of fulvic acid in enhancing the production of astaxanthin and lipids in Haematococcus pluvialis under abiotic stress conditions. J. Agric. Food Chem. 2019, 67, 12599–12609. [Google Scholar] [CrossRef] [PubMed]
- Constantinescu-Aruxandei, D.; Vlaicu, A.; Marinas, I.C.; Vintila, A.C.N.; Dimitriu, L.; Oancea, F. Effect of betaine and selenium on the growth and photosynthetic pigment production in Dunaliella salina as biostimulants. Fems Microbiol. Lett. 2019, 366, fnz257. [Google Scholar] [CrossRef] [PubMed]
- Fan, P.; Xu, P.; Zhu, Y.; Tu, X.; Song, G.; Zuo, Y.; Bi, Y. Addition of humic acid accelerates the growth of Euglena pisciformis AEW501 and the accumulation of lipids. J. Appl. Phycol. 2022, 34, 51–63. [Google Scholar] [CrossRef]
- Hunt, R.W.; Chinnasamy, S.; Bhatnagar, A.; Das, K.C. Effect of biochemical stimulants on biomass productivity and metabolite content of the microalga, Chlorella sorokiniana. Appl. Biochem. Biotechnol. 2010, 162, 2400–2414. [Google Scholar] [CrossRef]
- Zhu, J.; Wakisaka, M. Application of lignosulfonate as the growth promotor forfreshwater microalga Euglena gracilis to increase productivity of biomass and lipids. Fuel 2021, 283, 118920. [Google Scholar] [CrossRef]
- Zhu, J.; Tan, X.; Hafid, H.S.; Wakisaka, M. Enhancement of biomass yield and lipid accumulation of freshwater microalga Euglena gracilis by phenolic compounds from basic structures of lignin. Bioresour. Technol. 2021, 321, 124441. [Google Scholar] [CrossRef]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [Green Version]
- Kumawat, K.C.; Nagpal, S.; Sharma, P. Potential of plant growth-promoting rhizobacteria-plant interactions in mitigating salt stress for sustainable agriculture: A review. Pedosphere 2022, 32, 223–245. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L. Physiological responses to humic substances as plant growth promoter. Chem. Biol. Technol. Agric. 2014, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Stirk, W.A.; van Staden, J. Potential of phytohormones as a strategy to improve microalgae productivity for biotechnological applications. Biotechnol. Adv. 2020, 44, 107612. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.D.; Xu, J. Phytohormones in microalgae: A new opportunity for microalgal biotechnology? Trends Plant Sci. 2015, 20, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zeng, H.; Bartocci, P.; Fantozzi, F.; Yan, Y. Phytohormones and effects on growth and metabolites of microalgae: A review. Fermentation 2018, 4, 25. [Google Scholar] [CrossRef] [Green Version]
- Delaux, P.-M.; Xie, X.; Timme, R.E.; Puech-Pages, V.; Dunand, C.; Lecompte, E.; Delwiche, C.F.; Yoneyama, K.; Bécard, G.; Séjalon-Delmas, N. Origin of strigolactones in the green lineage. New Phytol. 2012, 195, 857–871. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Zhao, C.; Liu, J.; Zhao, Y.; Sun, S.; Wei, J. Simultaneous biogas upgrading and biogas slurry treatment by different microalgae-based technologies under various strigolactone analog (GR24) concentrations. Bioresour. Technol. 2022, 351, 127033. [Google Scholar]
- Xu, M.; Xue, Z.; Zhao, Y.; Sun, S.; Liu, J. Enhancement of the photosynthetic and removal performance for microalgae-based technologies by co-culture strategy and strigolactone induction. Bioresour. Technol. 2021, 339, 125579. [Google Scholar] [CrossRef]
- Song, X.; Zhao, Y.; Li, T.; Han, B.; Zhao, P.; Xu, J.-W.; Yu, X. Enhancement of lipid accumulation in Monoraphidium sp. QLY-1 by induction of strigolactone. Bioresour. Technol. 2019, 288, 121607. [Google Scholar] [CrossRef]
- Shen, X.; Xue, Z.; Sun, L.; Zhao, C.; Sun, S.; Liu, J.; Zhao, Y.; Liu, J. Effect of GR24 concentrations on biogas upgrade and nutrient removal by microalgae-based technology. Bioresour. Technol. 2020, 312, 123563. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.X.; Zhao, H.J.; Zhu, B.L.; Xu, J.L.; Xu, F.; Liu, S.X.; Li, X.H.; Zhou, C.X. Phloroglucinol promotes fucoxanthin synthesis by activating the cis-zeatin and brassinolide pathways in Thalassiosira pseudonana. Appl. Environ. Microbiol. 2022, 88, e0216021. [Google Scholar] [CrossRef]
- Da Silva, J.A.T.; Dobranszki, J.; Ross, S. Phloroglucinol in plant tissue culture. Vitr. Cell. Dev. Biol.-Plant 2013, 49, 1–16. [Google Scholar] [CrossRef]
- Petti, C. Phloroglucinol mediated plant regeneration of Ornithogalum dubium as the sole "hormone-like supplement" in plant tissue culture long-term experiments. Plants 2020, 9, 929. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Xu, J.; Yu, X.; Zhao, P.; Che, R. Method for improving oil content of oil-producing microalgae based on fulvic acid. CN105671095-A, 15 June 2016. [Google Scholar]
- Yu, X.; Li, D.; Zhao, Y. Use of Melatonin for Increasing oil Content of Oil-Producing Microalgae and Promoting Accumulation of Oil in Microalgae. CN107475171-A, 15 December 2017. [Google Scholar]
- Palacios, O.A.; López, B.R.; de-Bashan, L.E. Microalga Growth-Promoting Bacteria (MGPB): A formal term proposed for beneficial bacteria involved in microalgal–bacterial interactions. Algal Res. 2022, 61, 102585. [Google Scholar] [CrossRef]
- Ramanan, R.; Kim, B.-H.; Cho, D.-H.; Oh, H.-M.; Kim, H.-S. Algae–bacteria interactions: Evolution, ecology and emerging applications. Biotechnol. Adv. 2016, 34, 14–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.-Y.; Hu, H.-Y.; Wu, Y.-H.; Zhuang, L.-L.; Xu, X.-Q.; Wang, X.-X.; Dao, G.-H. Promising solutions to solve the bottlenecks in the large-scale cultivation of microalgae for biomass/bioenergy production. Renew. Sustain. Energy Rev. 2016, 60, 1602–1614. [Google Scholar] [CrossRef]
- De-Bashan, L.E.; Moreno, M.; Hernandez, J.P.; Bashan, Y. Removal of ammonium and phosphorus ions from synthetic wastewater by the microalgae Chlorella vulgaris coimmobilized in alginate beads with the microalgae growth-promoting bacterium Azospirillum brasilense. Water Res. 2002, 36, 2941–2948. [Google Scholar] [CrossRef]
- De-Bashan, L.E.; Hernandez, J.P.; Morey, T.; Bashan, Y. Microalgae growth-promoting bacteria as "helpers" for microalgae: A novel approach for removing ammonium and phosphorus from municipal wastewater. Water Res. 2004, 38, 466–474. [Google Scholar] [CrossRef]
- Do Nascimento, M.; Dublan, M.d.l.A.; Ortiz-Marquez, J.C.F.; Curatti, L. High lipid productivity of an Ankistrodesmus—Rhizobium artificial consortium. Bioresour. Technol. 2013, 146, 400–407. [Google Scholar] [CrossRef]
- Sapp, M.; Schwaderer, A.S.; Wiltshire, K.H.; Hoppe, H.G.; Gerdts, G.; Wichels, A. Species-specific bacterial communities in the phycosphere of microalgae? Microb. Ecol. 2007, 53, 683–699. [Google Scholar] [CrossRef]
- Ramanan, R.; Kang, Z.; Kim, B.-H.; Cho, D.-H.; Jin, L.; Oh, H.-M.; Kim, H.-S. Phycosphere bacterial diversity in green algae reveals an apparent similarity across habitats. Algal Res. 2015, 8, 140–144. [Google Scholar] [CrossRef]
- Ruzzi, M.; Aroca, R. Plant growth-promoting rhizobacteria act as biostimulants in horticulture. Sci. Hortic. 2015, 196, 124–134. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuentes, J.L.; Garbayo, I.; Cuaresma, M.; Montero, Z.; González-del-Valle, M.; Vílchez, C. Impact of microalgae-bacteria interactions on the production of algal biomass and associated compounds. Mar. Drugs 2016, 14, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinberg, C.E.W.; Bach, S. Growth promotion by a groundwater fulvic acid in a bacteria/algae system. Acta Hydrochim. Hydrobiol. 1996, 24, 98–100. [Google Scholar] [CrossRef]
- Zhao, X.; Zhou, Y.; Huang, S.; Qiu, D.Y.; Schideman, L.; Chai, X.L.; Zhao, Y.C. Characterization of microalgae-bacteria consortium cultured in landfill leachate for carbon fixation and lipid production. Bioresour. Technol. 2014, 156, 322–328. [Google Scholar] [CrossRef]
- Fuqua, C.; Greenberg, E.P. Listening in on bacteria: Acyl-homoserine lactone signalling. Nat. Rev. Mol. Cell Biol. 2002, 3, 685–695. [Google Scholar] [CrossRef]
- Dow, L. How do quorum-sensing signals mediate algae–bacteria interactions? Microorganisms 2021, 9, 1391. [Google Scholar] [CrossRef]
- Xu, Y.-Y.; Yang, J.-S.; Liu, C.; Wang, E.-T.; Wang, R.-N.; Qiu, X.-Q.; Li, B.-Z.; Chen, W.-F.; Yuan, H.-L. Water-soluble humic materials regulate quorum sensing in Sinorhizobium meliloti through a novel repressor of expR. Front. Microbiol. 2018, 9, 3194. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.J.; Ji, M.; Wang, F.; Tian, Z.K.; Wang, T.Y.; Wang, S.Y.; Wang, S.Y.; Yan, Z. Insight into the short-term effect of fulvic acid on nitrogen removal performance and N-acylated-L-homoserine lactones (AHLs) release in the anammox system. Sci. Total Environ. 2020, 704, 135285. [Google Scholar] [CrossRef]
- Sandilya, A.A.; Natarajan, U.; Priya, M.H. Molecular View into the Cyclodextrin Cavity: Structure and Hydration. ACS Omega 2020, 5, 25655–25667. [Google Scholar] [CrossRef]
- Verschoor, A.M.; Van Der Stap, I.; Helmsing, N.R.; Lürling, M.; Van Donk, E. Inducible colony formation within the Scenedesmaceae: Adaptive responses to infochemicals from two different herbivore taxa. J. Phycol. 2004, 40, 808–814. [Google Scholar] [CrossRef]
- Roccuzzo, S.; Beckerman, A.P.; Pandhal, J. The use of natural infochemicals for sustainable and efficient harvesting of the microalgae Scenedesmus spp. for biotechnology: Insights from a meta-analysis. Biotechnol. Lett. 2016, 38, 1983–1990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roccuzzo, S.; Couto, N.; Karunakaran, E.; Kapoore, R.V.; Butler, T.O.; Mukherjee, J.; Hansson, E.M.; Beckerman, A.P.; Pandhal, J. Metabolic insights into infochemicals induced colony formation and flocculation in Scenedesmus subspicatus unraveled by quantitative proteomics. Front. Microbiol. 2020, 11, 792. [Google Scholar] [CrossRef] [PubMed]
- Albini, D.; Fowler, M.S.; Llewellyn, C.; Tang, K.W. Reversible colony formation and the associated costs in Scenedesmus obliquus. J. Plankton Res. 2019, 41, 419–429. [Google Scholar] [CrossRef]
- Peng, Q.; Zhao, M.; Shen, G.; Gan, X.; Li, M. Linear alkylbenzene sulfonate (LAS) promotes sedimentation and lipid accumulation in Scenedesmus obliquus. RSC Adv. 2017, 7, 9244–9250. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.X.; Wang, Z.S.; Sun, Y.F.; Gu, L.; Zhang, L.; Wang, J.; Huang, Y.; Yang, Z. Surfactants at environmentally relevant concentrations interfere the inducible defense of Scenedesmus obliquus and the implications for ecological risk assessment. Environ. Pollut. 2020, 261, 114131. [Google Scholar] [CrossRef]
- Oda, Y.; Sakamoto, M.; Miyabara, Y. Colony formation in three species of the family Scenedesmaceae (Desmodesmus subspicatus, Scenedesmus acutus, Tetradesmus dimorphus) exposed to sodium dodecyl sulfate and its interference with grazing of Daphnia galeata. Arch. Environ. Contam. Toxicol. 2022, 82, 37–47. [Google Scholar] [CrossRef]
- Lamar, R.T.; Monda, H.; Sleighter, R. Use of ore-derived humic acids with diverse chemistries to elucidate Structure-Activity Relationships (SAR) of humic acids in plant phenotypic expression. Front. Plant Sci. 2021, 12, 758424. [Google Scholar] [CrossRef]
- Nardi, S.; Schiavon, M.; Francioso, O. Chemical structure and biological activity of humic substances define their role as plant growth promoters. Molecules 2021, 26, 2256. [Google Scholar] [CrossRef]
- Van Tol de Castro, T.A.; Berbara, R.L.L.; Tavares, O.C.H.; Mello, D.F.d.G.; Pereira, E.G.; Souza, C.d.C.B.d.; Espinosa, L.M.; García, A.C. Humic acids induce a eustress state via photosynthesis and nitrogen metabolism leading to a root growth improvement in rice plants. Plant Physiol. Biochem. 2021, 162, 171–184. [Google Scholar] [CrossRef]
- Savarese, C.; di Meo, V.; Cangemi, S.; Verrillo, M.; Savy, D.; Cozzolino, V.; Piccolo, A. Bioactivity of two different humic materials and their combination on plants growth as a function of their molecular properties. Plant Soil 2022, 472, 509–526. [Google Scholar] [CrossRef]
- Savy, D.; Brostaux, Y.; Cozzolino, V.; Delaplace, P.; du Jardin, P.; Piccolo, A. Quantitative structure-activity relationship of humic-like biostimulants derived from agro-industrial byproducts and energy crops. Front. Plant Sci. 2020, 11, 581. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Xu, C.B.; Xu, Y.Z.; Bai, Y.C.; Guo, F. Molecular signatures of humic acids from different sources as revealed by ultrahigh resolution mass spectrometry. J. Chem. 2020, 2020, 7171582. [Google Scholar] [CrossRef]
- Monda, H.; McKenna, A.M.; Fountain, R.; Lamar, R.T. Bioactivity of humic acids extracted from shale ore: Molecular characterization and structure-activity relationship with tomato plant yield under nutritional stress. Front. Plant Sci. 2021, 12, 660224. [Google Scholar] [CrossRef]
- Aeschbacher, M.; Graf, C.; Schwarzenbach, R.P.; Sander, M. Antioxidant properties of humic substances. Environ. Sci. Technol. 2012, 46, 4916–4925. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.J.; Chae, A.N.; Song, K.G.; Park, J.; Lee, B.C. Effect of trophic conditions on microalga growth, nutrient removal, algal organic matter, and energy storage products in Scenedesmus (Acutodesmus) obliquus KGE-17 cultivation. Bioprocess Biosyst. Eng. 2019, 42, 1225–1234. [Google Scholar] [CrossRef]
- Khan, S.I.; Zamyadi, A.; Rao, N.R.H.; Li, X.; Stuetz, R.M.; Henderson, R.K. Fluorescence spectroscopic characterisation of algal organic matter: Towards improved in-situ fluorometer development. Environ. Sci. Water Res. Technol. 2019, 5, 417–432. [Google Scholar] [CrossRef]
- Khan, W.; Park, J.W.; Maeng, S.K. Fluorescence descriptors for algal organic matter and microalgae disintegration during ultrasonication. J. Water Process Eng. 2022, 45, 102517. [Google Scholar] [CrossRef]
- Wu, M.C.; Du, M.; Wu, G.M.; Lu, F.M.; Li, J.; Lei, A.P.; Zhu, H.; Hu, Z.L.; Wang, J.X. Water reuse and growth inhibition mechanisms for cultivation of microalga Euglena gracilis. Biotechnol. Biofuels 2021, 14, 132. [Google Scholar] [CrossRef]
- Sha, J.; Lu, Z.; Ye, J.; Wang, G.; Hu, Q.; Chen, Y.; Zhang, X. The inhibition effect of recycled Scenedesmus acuminatus culture media: Influence of growth phase, inhibitor identification and removal. Algal Res. 2019, 42, 101612. [Google Scholar] [CrossRef]
- Zhang, X.; Lu, Z.; Wang, Y.; Wensel, P.; Sommerfeld, M.; Hu, Q. Recycling Nannochloropsis oceanica culture media and growth inhibitors characterization. Algal Res. 2016, 20, 282–290. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Lam, M.K.; Lee, K.T. Microalgae biofuels: A critical review of issues, problems and the way forward. Biotechnol Adv. 2012, 30, 673–690. [Google Scholar] [CrossRef]
- Christenson, L.; Sims, R. Production and harvesting of microalgae for wastewater treatment, biofuels, and bioproducts. Biotechnol Adv. 2011, 29, 686–702. [Google Scholar] [CrossRef]
- Rawat, I.; Ranjith Kumar, R.; Mutanda, T.; Bux, F. Biodiesel from microalgae: A critical evaluation from laboratory to large scale production. Appl. Energy 2013, 103, 444–467. [Google Scholar] [CrossRef]
- Chisti, Y. Constraints to commercialization of algal fuels. J. Biotechnol. 2013, 167, 201–214. [Google Scholar] [CrossRef]
- Uduman, N.; Qi, Y.; Danquah, M.K.; Forde, G.M.; Hoadley, A. Dewatering of microalgal cultures: A major bottleneck to algae-based fuels. J. Renew. Sustain. Energy 2010, 2, 012701. [Google Scholar] [CrossRef]
- Barros, A.I.; Gonçalves, A.L.; Simões, M.; Pires, J.C.M. Harvesting techniques applied to microalgae: A review. Renew. Sustain. Energy Rev. 2015, 41, 1489–1500. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, G.; Hossain, A.K.; Sharma, V.; Duraisamy, G. Key Targets for Improving Algal Biofuel Production. Clean Technol. 2021, 3, 711–742. [Google Scholar] [CrossRef]
- Liber, J.A.; Bryson, A.E.; Bonito, G.; Du, Z.Y. Harvesting microalgae for food and energy products. Small Methods 2020, 4, 2000349. [Google Scholar] [CrossRef]
- Fasaei, F.; Bitter, J.H.; Slegers, P.M.; van Boxtel, A.J.B. Techno-economic evaluation of microalgae harvesting and dewatering systems. Algal Res. 2018, 31, 347–362. [Google Scholar] [CrossRef]
- Mathimani, T.; Mallick, N. A comprehensive review on harvesting of microalgae for biodiesel—Key challenges and future directions. Renew. Sustain. Energy Rev. 2018, 91, 1103–1120. [Google Scholar] [CrossRef]
- Khan, S.; Naushad, M.; Iqbal, J.; Bathula, C.; Sharma, G. Production and harvesting of microalgae and an efficient operational approach to biofuel production for a sustainable environment. Fuel 2022, 311, 122543. [Google Scholar] [CrossRef]
- Min, K.H.; Kim, D.H.; Ki, M.R.; Pack, S.P. Recent progress in flocculation, dewatering, and drying technologies for microalgae utilization: Scalable and low-cost harvesting process development. Bioresour. Technol. 2022, 344, 126404. [Google Scholar] [CrossRef] [PubMed]
- Huh, Y.S.; Lee, Y.C. Harvesting Oleaginous Microorganisms Using Aminoclay-Induced Humic Acid. KR1567474-B1, 11 November 2015. [Google Scholar]
- Lee, Y.C.; Oh, S.Y.; Lee, H.U.; Kim, B.; Lee, S.Y.; Choi, M.H.; Lee, G.W.; Park, J.Y.; Oh, Y.K.; Ryu, T.; et al. Aminoclay-induced humic acid flocculation for efficient harvesting of oleaginous Chlorella sp. Bioresour Technol. 2014, 153, 365–369. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, H.; Cheng, S.; Zhang, W.; Zhang, X. Enhanced microalgal harvesting using microalgae-derived extracellular polymeric substance as flocculation aid. ACS Sustain. Chem. Eng. 2020, 8, 4069–4075. [Google Scholar] [CrossRef]
- Cheng, Y.L.; Juang, Y.C.; Liao, G.Y.; Tsai, P.W.; Ho, S.H.; Yeh, K.L.; Chen, C.Y.; Chang, J.S.; Liu, J.C.; Chen, W.M.; et al. Harvesting of Scenedesmus obliquus FSP-3 using dispersed ozone flotation. Bioresour. Technol. 2011, 102, 82–87. [Google Scholar] [CrossRef]
- Beuckels, A.; Depraetere, O.; Vandamme, D.; Foubert, I.; Smolders, E.; Muylaert, K. Influence of organic matter on flocculation of Chlorella vulgaris by calcium phosphate precipitation. Biomass Bioenergy 2013, 54, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Borowitzka, M.A. High-value products from microalgae—Their development and commercialisation. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Mohammady, N.G.-E. Humic acid stimulation of growth and optimization of biochemical profiles on two microalgal species proposed as live feeds in aquaculture. Int. J. Recirc. Aquac. 2008, 9, 13–28. [Google Scholar] [CrossRef]
- Zhao, Y.T.; Shang, M.M.; Xu, J.W.; Zhao, P.; Li, T.; Yu, X.Y. Enhanced astaxanthin production from a novel strain of Haematococcus pluvialis using fulvic acid. Process. Biochem. 2015, 50, 2072–2077. [Google Scholar] [CrossRef]
- Meinelt, T.; Schreckenbach, K.; Pietrock, M.; Heidrich, S.; Steinberg, C.E. Humic substances. Part 1: Dissolved humic substances (HS) in aquaculture and ornamental fish breeding. Environ. Sci. Pollut. Res. Int. 2008, 15, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Vetvicka, V.; Baigorri, R.; Zamarreno, A.M.; Garcia-Mina, J.M.; Yvin, J.C. Glucan and humic acid: Synergistic effects on the immune system. J. Med. Food 2010, 13, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Colla, G.; Rouphael, Y. Microalgae: New source of plant biostimulants. Agronomy 2020, 10, 1240. [Google Scholar] [CrossRef]
- González-Pérez, B.K.; Rivas-Castillo, A.M.; Valdez-Calderón, A.; Gayosso-Morales, M.A. Microalgae as biostimulants: A new approach in agriculture. World J. Microbiol. Biotechnol. 2022, 38, 1–12. [Google Scholar] [CrossRef]
- Chiaiese, P.; Corrado, G.; Colla, G.; Kyriacou, M.C.; Rouphael, Y. Renewable sources of plant biostimulation: Microalgae as a sustainable means to improve crop performance. Front. Plant Sci. 2018, 9, 1782. [Google Scholar] [CrossRef] [Green Version]
- Oancea, F.; Velea, S.; Fãtu, V.; Mincea, C.; Ilie, L. Micro-algae based plant biostimulant and its effect on water stressed tomato plants. Rom. J. Plant Prot. 2013, 6, 104–117. [Google Scholar]
- Mógor, Á.F.; Ördög, V.; Lima, G.P.P.; Molnár, Z.; Mógor, G. Biostimulant properties of cyanobacterial hydrolysate related to polyamines. J. Appl. Phycol. 2017, 30, 453–460. [Google Scholar] [CrossRef]
- Creelman, R.A.; Mullet, J.E. Oligosaccharins, brassinolides, and jasmonates: Nontraditional regulators of plant growth, development, and gene expression. Plant Cell 1997, 9, 1211–1223. [Google Scholar] [CrossRef] [Green Version]
- Abdul Malik, N.A.; Kumar, I.S.; Nadarajah, K. Elicitor and receptor molecules: Orchestrators of plant defense and immunity. Int. J. Mol. Sci. 2020, 21, 963. [Google Scholar] [CrossRef] [Green Version]
- El Arroussi, H.; Benhima, R.; Elbaouchi, A.; Sijilmassi, B.; El Mernissi, N.; Aafsar, A.; Meftah-Kadmiri, I.; Bendaou, N.; Smouni, A. Dunaliella salina exopolysaccharides: A promising biostimulant for salt stress tolerance in tomato (Solanum lycopersicum). J. Appl. Phycol. 2018, 30, 2929–2941. [Google Scholar] [CrossRef]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant action of protein hydrolysates: Unraveling their effects on plant physiology and microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurya, R.; Paliwal, C.; Chokshi, K.; Pancha, I.; Ghosh, T.; Satpati, G.G.; Pal, R.; Ghosh, A.; Mishra, S. Hydrolysate of lipid extracted microalgal biomass residue: An algal growth promoter and enhancer. Bioresour. Technol. 2016, 207, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Zandonadi, D.B.; Santos, M.P.; Caixeta, L.S.; Marinho, E.B.; Peres, L.E.P.; Façanha, A.R. Plant proton pumps as markers of biostimulant action. Sci. Agric. 2016, 73, 24–28. [Google Scholar] [CrossRef] [Green Version]
- Klein, P.; Chauvey, L.; Kallerhoff, J.; Pinelli, E.; Morard, M.; Silvestre, J. A Tool derived from the Vicia faba micronucleus assay, to assess genotoxicity, cytotoxicity or biostimulation of novel compounds used in agriculture. Agronomy 2021, 11, 321. [Google Scholar] [CrossRef]
- Ugena, L.; Hýlová, A.; Podlešáková, K.; Humplík, J.F.; Doležal, K.; Diego, N.D.; Spíchal, L. Characterization of biostimulant mode of action using novel multi-trait high-throughput screening of Arabidopsis germination and rosette growth. Front. Plant Sci. 2018, 9, 1327. [Google Scholar] [CrossRef] [Green Version]
- Rayorath, P.; Khan, W.; Palanisamy, R.; MacKinnon, S.L.; Stefanova, R.; Hankins, S.D.; Critchley, A.T.; Prithiviraj, B. Extracts of the brown seaweed Ascophyllum nodosum induce gibberellic acid (GA3)-independent amylase activity in barley. J. Plant Growth Regul. 2008, 27, 370–379. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef] [Green Version]
- Che, R.Q.; Huang, L.; Xu, J.W.; Zhao, P.; Li, T.; Ma, H.X.; Yu, X.Y. Effect of fulvic acid induction on the physiology, metabolism, and lipid biosynthesis-related gene transcription of Monoraphidium sp. FXY-10. Bioresour. Technol. 2017, 227, 324–334. [Google Scholar] [CrossRef]
- Cui, N.; Xiao, J.M.; Feng, Y.J.; Zhao, Y.T.; Yu, X.Y.; Xu, J.W.; Li, T.; Zhao, P. Antioxidants enhance lipid productivity in Heveochlorella sp. Yu. Algal Res. Biomass Biofuels Bioprod. 2021, 55, 102235. [Google Scholar] [CrossRef]
- Martini, S.; D’Addario, C.; Bonechi, C.; Leone, G.; Tognazzi, A.; Consumi, M.; Magnani, A.; Rossi, C. Increasing photostability and water-solubility of carotenoids: Synthesis and characterization of β-carotene–humic acid complexes. J. Photochem. Photobiol. B Biol. 2010, 101, 355–361. [Google Scholar] [CrossRef]
- Vetvicka, V.; Garcia-Mina, J.M.; Proctor, M.; Yvin, J.C. Humic Acid and glucan: Protection against liver injury induced by carbon tetrachloride. J. Med. Food 2015, 18, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Hamza, Z.K.; Hathout, A.S.; Ostroff, G.; Soto, E.; Sabry, B.A.; El-Hashash, M.A.; Hassan, N.S.; Aly, S.E. Assessment of the protective effect of yeast cell wall beta-glucan encapsulating humic acid nanoparticles as an aflatoxin B-1 adsorbent in vivo. J. Biochem. Mol. Toxicol. 2022, 36, e22941. [Google Scholar] [CrossRef] [PubMed]
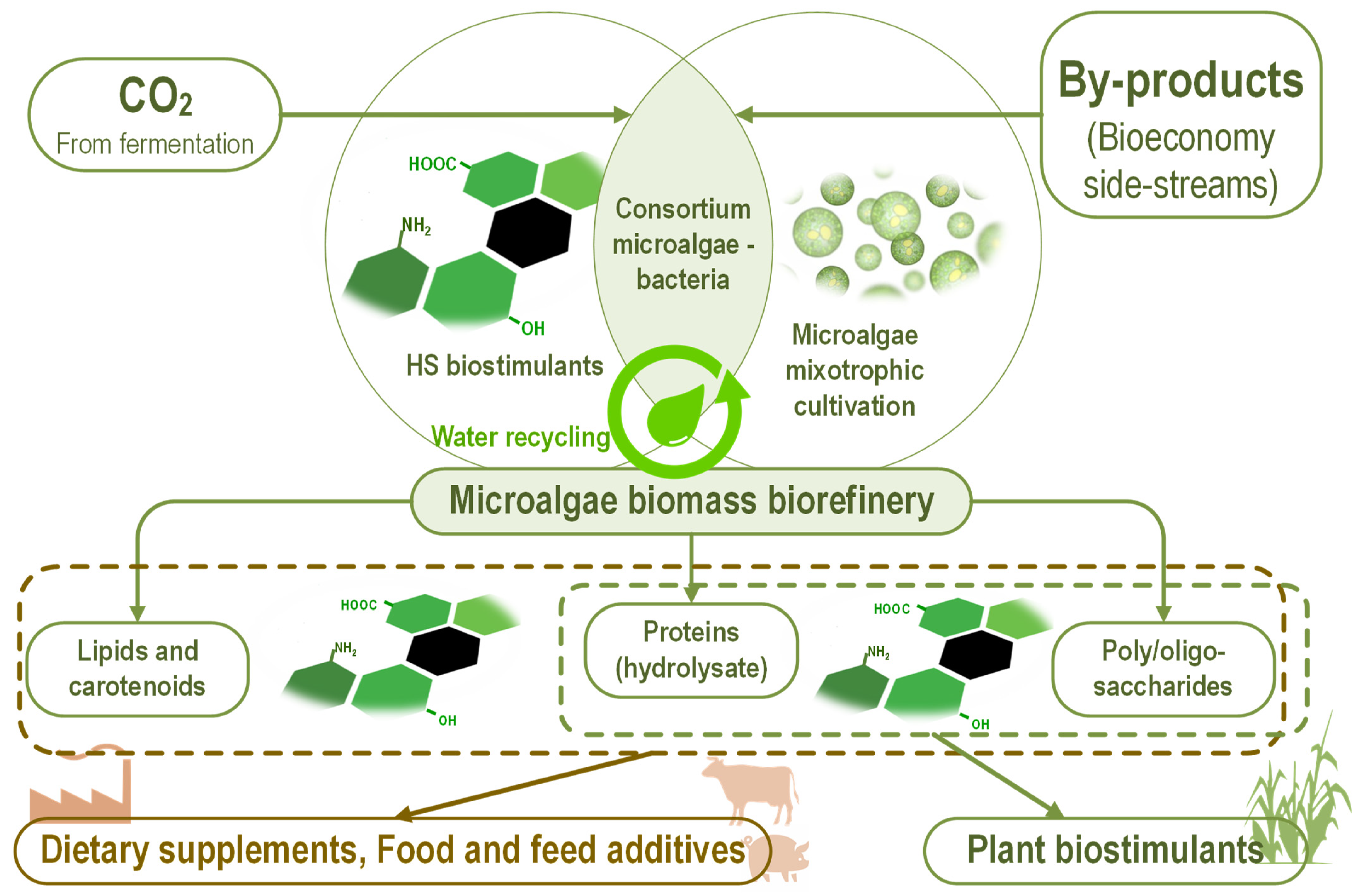

{kind=link}
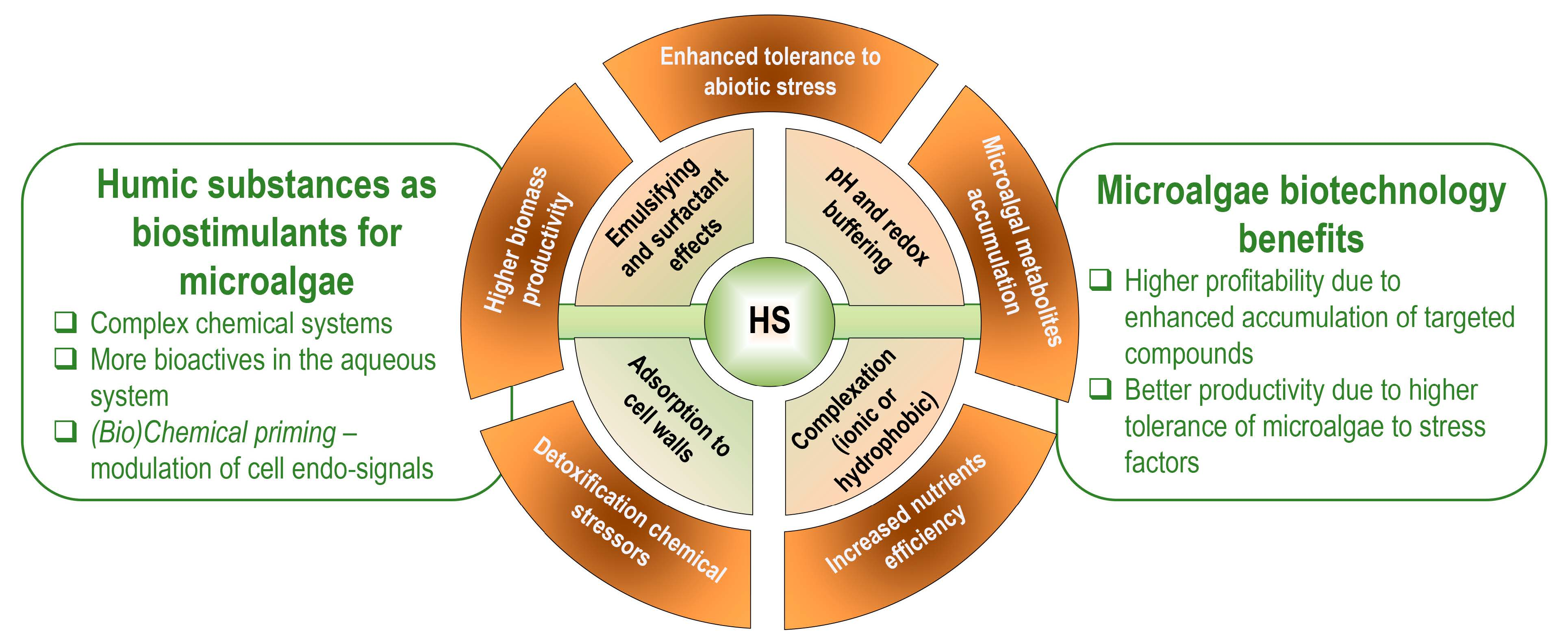
{kind=link}
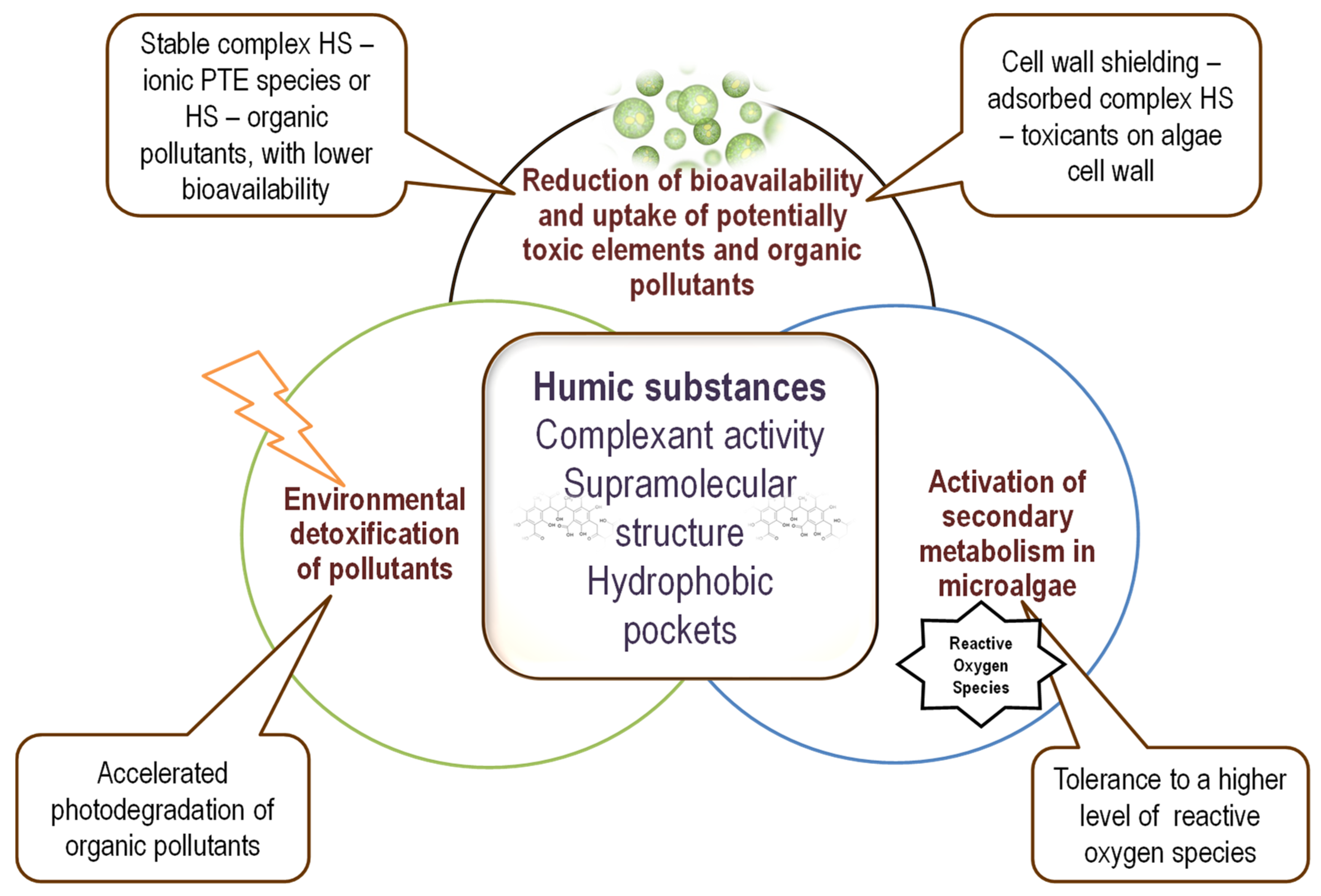{kind=link}
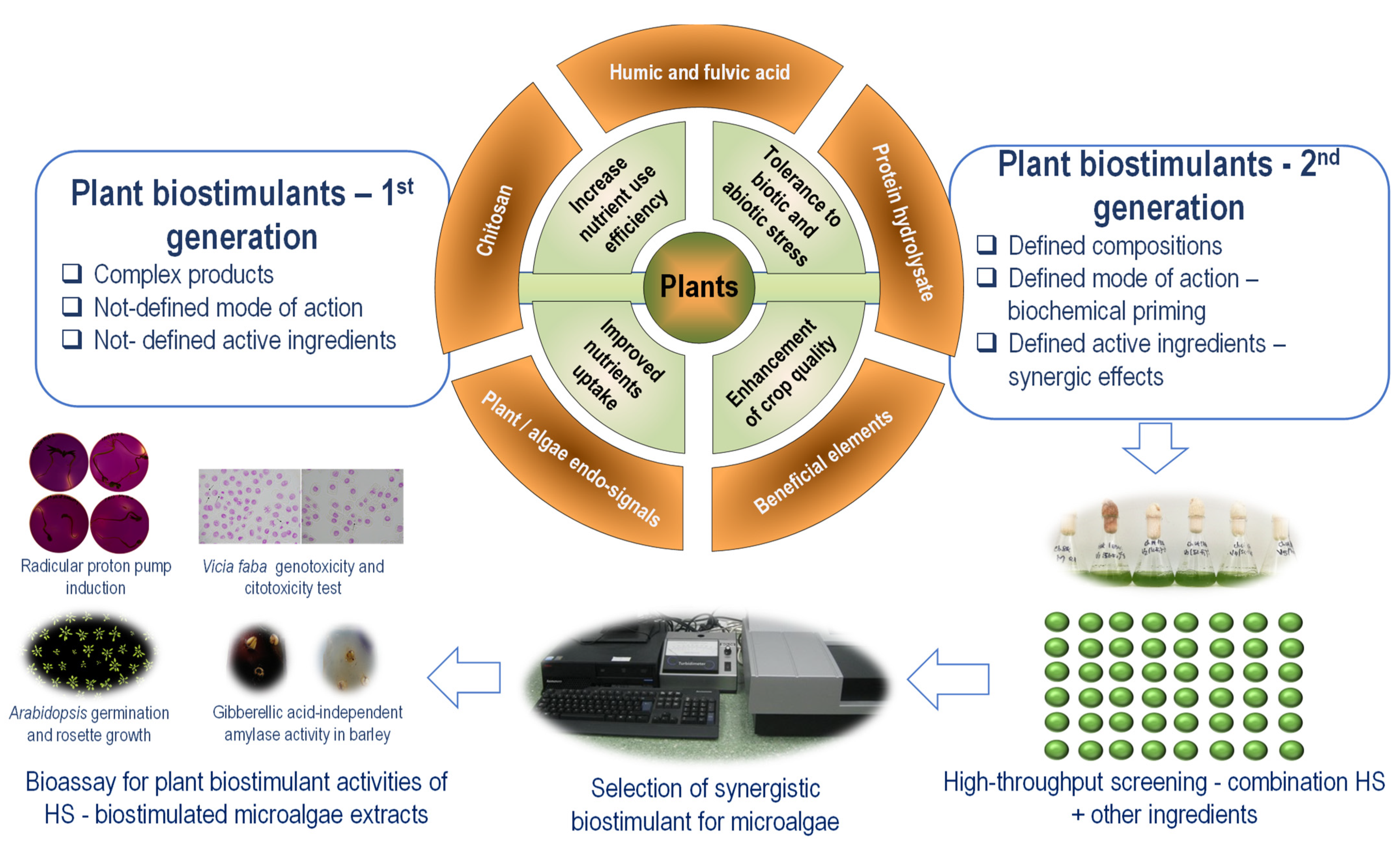{kind=link}
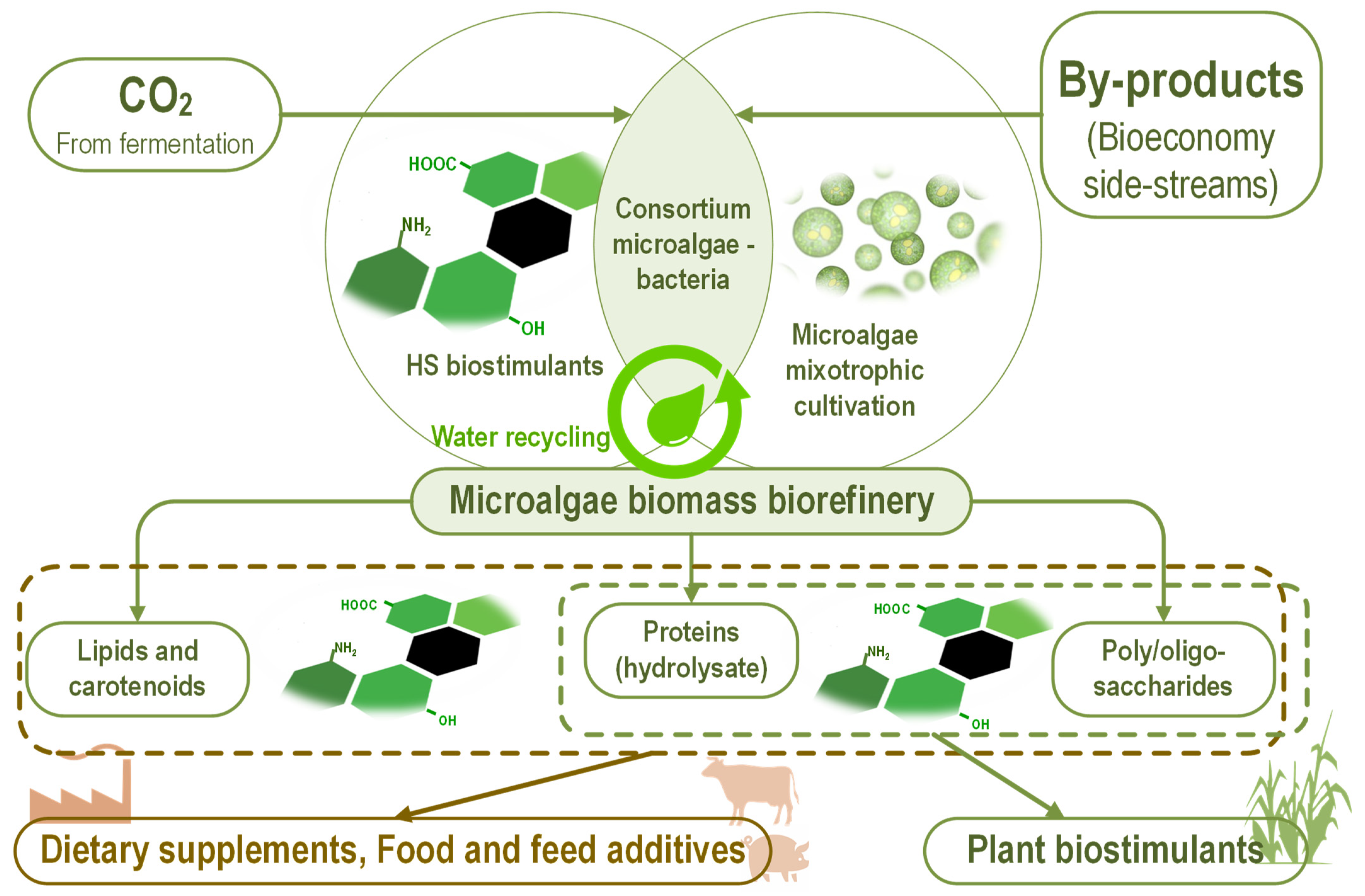{kind=link}
| Humic Substances | Tested Microalgae | Concentration–Effect | Reference |
|---|---|---|---|
| Humic acid extracted from lignite | Scenedesmus acutus Meyen Tomaselli 8 Chlorella vulgaris C-3 Anabaena variabilis 786 Nostoc commune | Up to 10 mg L−1 enhances biomass accumulation 100 mg L−1 enhances protein accumulation 1 g L−1—inhibition | Pouneva, 2005 [64] |
| Humic acids extracted from lake sediments | Desmodesmus communis 41.71 Chroococcus minutus 276-4b | 0.3 mg L−1 increases the number of D. communis cells Inhibitory effects on C. minutus | Prokhotskaya and Steinberg, 2007 [63] |
| Humic acid extracted from lignite, Artificial humic acid | Raphidocelis subcapitata 61.81 Monoraphidium braunii 2006 Synechocystis PCC 6803 Microcystis aeruginosa PCC 7806 | 0.17 mM stimulates photosynthesis 4.7 mM reduces cell development and inhibits photosynthesis | Bährs et al., 2012 [62] |
| Humic acids extracted from soils | Chlorella vulgaris co. 157 | 0.01–0.03% activation >0.03% inhibition | Toropkina et al., 2017 [61] |
| Commercial (Suwannee River) humic acids | Scenedesmus capricornus FACHB-271 Chlorella spp. FACHB-271 | 0.05–0.1 mg L−1 stimulation 1.0 mg L−1 inhibition | Zheng et al., 2022 [60] |
| Aquatic Pollutant | Tested Microalgae | Main Mechanism | Reference |
|---|---|---|---|
| Cd2+, Zn2+ | Raphidocelis subcapitata | Supramolecular structure adsorbed in the cell wall surface, which complexes toxic ions | Koukal et al., 2003 [87] |
| Pb2+ | Chlorella kesslerii | HS–Pb2+ complexes are adsorbed on microalgae cell walls. HS photoalteration reduce the adsorption of HS–Pb2+ to microalgae surface | Spierings et al., 2011 [88] |
| Cu2+ | Chlorella vulgaris | HS addition reduces bioavailability of Cu2+ and decreases the secretion of exopolysaccharide matrix involved in Cu2+ toxicity | Shi et al., 2021 [89] |
| Microplastics | Chlorella vulgaris | HA decrease electrostatic interactions between polystyrene nanoplastics and microalgae and ameliorate cellular aggregation | Hanachi et al., 2022 [90] |
| Tetracycline | Coelastrella sp. | Reduction of oxidative stress damage (due to biochemical priming) | Tong et al., 2020 [91] |
| Graphene family materials (GFMs) | Chlorella pyrenoidosa | Reduction of absorption due to steric hindrance HA- GFMs Reduction of oxidative stress damage | Zhao et al., 2019 [92] |
| Compound | Tested Microalgae | Main Effects | Reference |
|---|---|---|---|
| Humic-like extract of anaerobic digestate (D-HL) Humic-like extract of residues from rapeseed oil production (B-HL) Humic-like extract of tomato plants (T-HL) | Chlorella vulgaris CCAP 211/11C Scenedesmus quadricauda | Increased biomass production (~25–40%) by DH-L and TH-L Increased oil accumulation (~60–90%) by DH-L and TH-L Increased unsaturated fatty acid content by B-HL Increased carbohydrate content by B-HL | Puglisi et al., 2018 [56] |
| Fulvic acid | Haematococcus pluvialis KM115647 | Increased astaxanthin and lipid content | Zhao et al., 2019 [127] |
| Selenium and betaine | Dunaliella salina | Increased carotenoid and antioxidant activity | Constantinescu-Aruxandei et al., 2019 [128] |
| Humic acids | Euglena pisciformis AEW501 | Increased biomass yield Higher lipid content Higher content of unsaturated fatty acids | Fan et al., 2022 [129] |
| Humic and fulvic acid (commercial preparation) | Chlorella sorokiana UTEX2805 | Increased biomass yield Increased metabolite accumulation | Hunt et al., 2010 [130] |
| Lignosulfonate | Euglena gracilis NIES-48 | Increased biomass yield Higher lipid content | Zhu and Wakisaka, 2021 [131] |
| Phenolic precursors of lignin | Zhu et al., 2021 [132] |
| High Value-Added Compounds | Market Estimation | Price Range (USD kg−1) | |
|---|---|---|---|
| Estimated Value (mio.US$) | Compound Annual Growth Rate—CAGR | ||
| Plant biostimulants | 3200 (2021) a | 12.1% (2021–2026) | 60–90 a |
| Carotenoids (total) | 1500 (2019) b | 4.2% (2019–2027) b | - |
| Beta-carotene | 532 (2019) a | 3.3% (2014–2019) b | 300–1500 b |
| Lutein | 314 (2019) a | 3.6% (2014–2019) b | - |
| Astaxanthin | 423 (2019) a | 2.3% (2014–2019) b | 200–7000 b |
| Canthaxanthin | 117 (2019) a | 3.7% (2014–2019) b | 100–500 d |
| Omega-3 fatty acids | 2100 (2020) c | 7.4% (2020–2028) c | 80–160 d |
| Active Ingredients | Microalgae | Main Mechanism | Reference |
|---|---|---|---|
| Polysaccharides | Dunaliella salina MS002, Porphyridium sp. MS081, D. salina strain MS067, Phaeodactylum tricornutum MS023, Desmodesmus sp., Arthrospira. platensis MS001 | Elicitation of the plant defense mechanisms and activation of secondary metabolism | Rachidi et al., 2021 [116] |
| Osmoprotectants—glycine-betaine and proline | Nannochloris sp. 424-1, CCAP 251/10 | Protection of plants against hydric stress, enhanced water use efficiency | Oancea et al., 2013 [214] |
| Osmoprotectants—polyamines | A. platensis | Increased biomass yield Higher lipid content Higher content of unsaturated fatty acids | Mógor et al., 2018 [215] |
| Protein hydrolysate | Chlorella vulgaris | Activation of primary metabolism, Increased nutrient uptake and nutrient use efficiency | Maurya et al., 2016 [220] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popa, D.G.; Lupu, C.; Constantinescu-Aruxandei, D.; Oancea, F. Humic Substances as Microalgal Biostimulants—Implications for Microalgal Biotechnology. Mar. Drugs 2022, 20, 327. https://doi.org/10.3390/md20050327
Popa DG, Lupu C, Constantinescu-Aruxandei D, Oancea F. Humic Substances as Microalgal Biostimulants—Implications for Microalgal Biotechnology. Marine Drugs. 2022; 20(5):327. https://doi.org/10.3390/md20050327
Chicago/Turabian StylePopa, Daria Gabriela, Carmen Lupu, Diana Constantinescu-Aruxandei, and Florin Oancea. 2022. "Humic Substances as Microalgal Biostimulants—Implications for Microalgal Biotechnology" Marine Drugs 20, no. 5: 327. https://doi.org/10.3390/md20050327
APA StylePopa, D. G., Lupu, C., Constantinescu-Aruxandei, D., & Oancea, F. (2022). Humic Substances as Microalgal Biostimulants—Implications for Microalgal Biotechnology. Marine Drugs, 20(5), 327. https://doi.org/10.3390/md20050327